
Iron Deficiency and Iron Deficiency Anemia: A Comprehensive Overview of Established and Emerging Concepts

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Prevalence, Causes, and Outcomes of Anemia, ID, and IDA
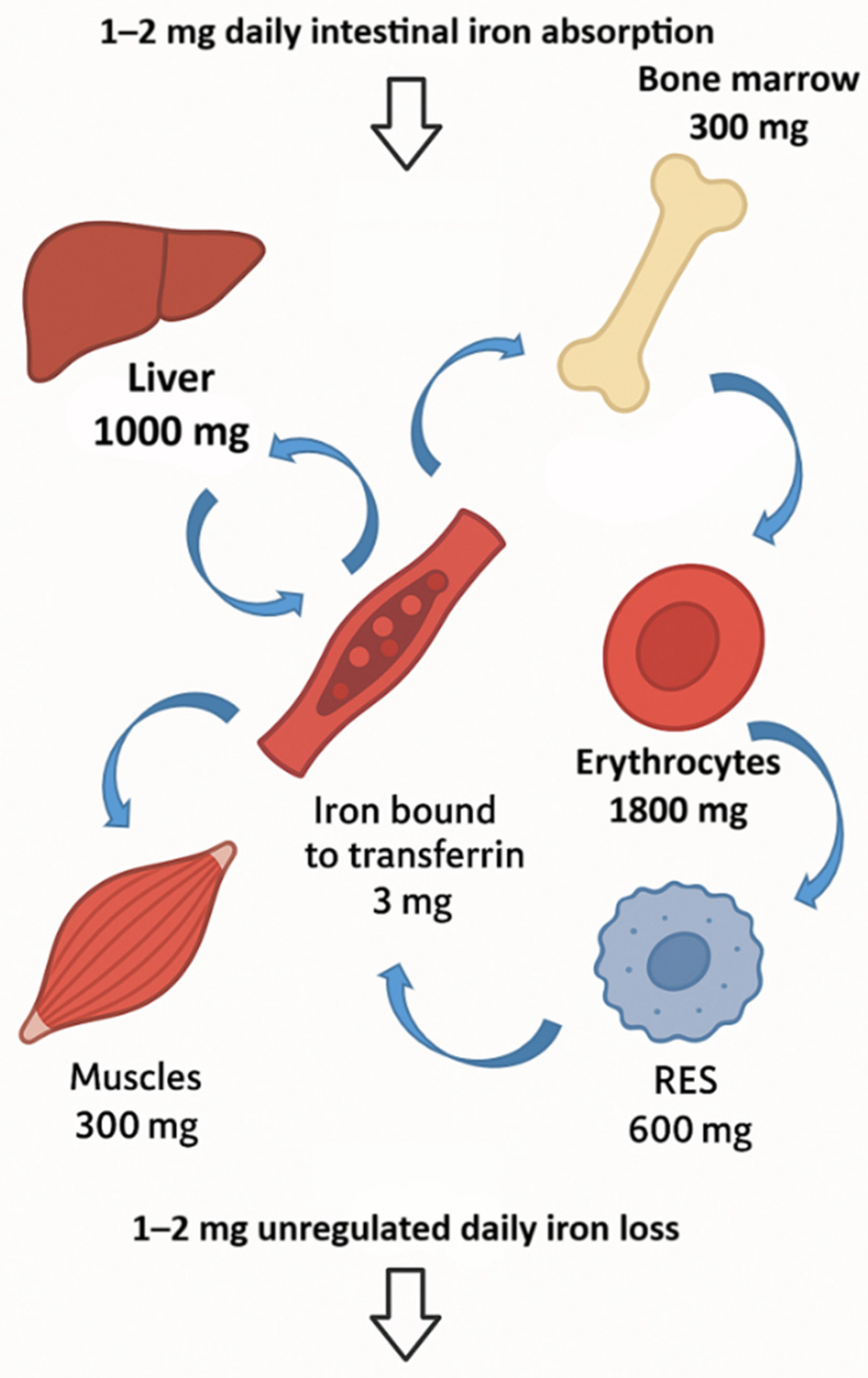
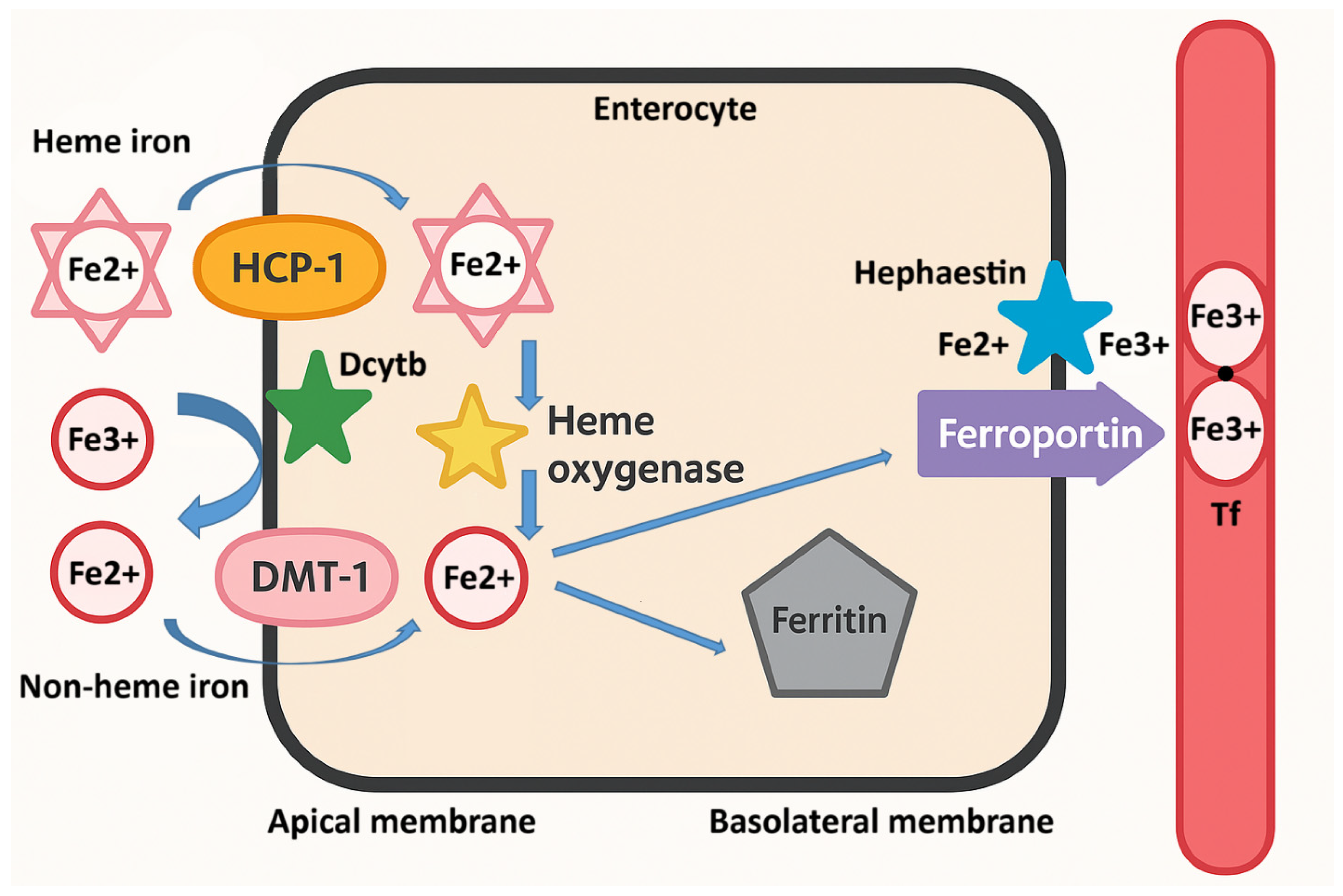
3. Iron Absorption and Metabolism

{kind=link}
{kind=link}
{kind=link}
| Name | Type | Function/Mechanism | Change in ID | Functional Consequence | Reference |
|---|---|---|---|---|---|
| Hepcidin | Gene/Protein | Regulates systemic iron homeostasis by degrading ferroportin | Downregulated | Increases iron absorption and release from stores | [61] |
| Hemojuvelin | Gene/Protein | BMP6 co-receptor, activates BMP–SMAD pathway for hepcidin induction | Downregulated | Reduces hepcidin expression, increases iron absorption | [63] |
| BMP6 | Gene/Protein | Stimulates hepcidin via SMAD signaling | Downregulated | Reduces hepcidin synthesis | [63] |
| Transferrin Receptor 2 | Gene/Protein | Iron sensor in hepatocytes; mediates hepcidin regulation | Downregulated | Decreases hepcidin synthesis, enhances iron absorption | [63] |
| Erythroferrone | Protein/Hormone | Suppresses hepcidin in response to erythropoietin | Upregulated | Increases iron availability for erythropoiesis | [65] |
| Histone Deacetylase 3 | Epigenetic Regulator | Epigenetically represses hepcidin expression | Increased activity | Decreases hepcidin transcription, enhances absorption | [64] |
| mTOR | Protein/Signaling | Regulates cell growth, autophagy, erythropoiesis; Fe-sensitive | Inhibited (decreased) | Promotes ferritinophagy and metabolic adaptation | [69] |
| Tristetraprolin | Protein/RNA-binding | Binds to mRNAs, promotes degradation; regulates inflammatory and iron metabolism mRNAs | Upregulated | Modulates inflammatory signals affecting hepcidin and iron regulation | [69] |
| Ferroportin | Gene/Protein | Iron exporter in enterocytes/macrophages | Stabilized (increased) | Enhances plasma iron availability | [67] |
| DMT-1 (SLC11A2) | Gene/Protein | Imports dietary non-heme iron into enterocytes | Upregulated | Promotes intestinal iron absorption | [65,66,67] |
| Dcytb | Gene/Protein | Reduces Fe3+ to Fe2+ at brush border | Upregulated | Facilitates iron absorption via DMT-1 | [65,66,67] |
| Transferrin receptor 1 | Gene/Protein | Cellular uptake of transferrin-bound iron | Upregulated | Increases cellular iron import | [65] |
| Transferrin | Protein | Iron transport in plasma | Decreased or unchanged | Affects iron delivery to tissues | [63] |
| SLC11A1 | Gene/Protein | Iron transporter in macrophages | Upregulated | Modulates immune-related iron handling | [25] |
| Ferritin (FTH1/FTL) | Gene/Protein | Cytosolic iron storage | Downregulated | Reflects depleted iron stores | [70] |
| NCOA4 | Gene/Protein | Mediates ferritin degradation (ferritinophagy) | Upregulated | Mobilizes stored intracellular iron | [70] |
| IRP1/IRP2 | Protein | Iron sensors; regulate translation via IREs | Active (increased) | Enhances iron uptake, suppresses storage/export | [68] |
| HRI | Protein/Kinase | Senses heme deficiency, phosphorylates eIF2α to inhibit protein synthesis | Activated | Prevents accumulation of unassembled globin; modulates erythropoiesis under ID | [73] |
| 5-aminolevulinic acid synthase 2 | Gene/Protein (Enzyme) | Catalyzes first step in heme biosynthesis in erythroid cells; essential for hemoglobin production | Downregulated or functionally impaired due to iron shortage | Limits heme and hemoglobin synthesis, contributing to anemia and ineffective erythropoiesis | [65] |
| Erythropoietin | Gene/Protein | Stimulates red blood cell production | Upregulated | Compensates for anemia | [73] |
| HIF-2α | Gene/Protein | Controls hypoxia response; induces DMT-1/Erythropoietin | Upregulated | Enhances erythropoiesis and absorption | [66] |
| Ferritin (circulating) | Protein/Metabolite | Stored iron in blood | Decreased | Key diagnostic marker of ID | [70] |
| Lactate | Metabolite | End product of anaerobic glycolysis | Increased | Suggests metabolic shift due to hypoxia | [65,66,67] |
| Succinate | Metabolite | TCA cycle intermediate | Accumulates | Activates HIFs and IRPs | [65,66,67] |
| Pyruvate | Metabolite | Glycolysis end product | Increased | Reflects altered cellular metabolism | [65,66,67] |
| ATP | Metabolite | Energy currency of the cell | Decreased | Reduced mitochondrial function | [65,66,67] |
| Reactive oxygen species | Metabolite | By-product of redox metabolism | Variable (increased) | Indicates oxidative stress | [55] |
| Iron-regulated amino acids (e.g., histidine, tryptophan) | Metabolites | Cofactors for Fe-dependent enzymes | Altered | Affected by Fe-dependent metabolic pathways | [65,66,67] |
3.1. Inhibitors of Iron Absorption in Food
| Flavonoid | Source (Fruits/Vegetables/Grains) | Modulation of Iron | Mode of Action | Disease Relevance | References |
|---|---|---|---|---|---|
| Quercetin | Onions, apples, berries, grapes | Inhibits absorption | Chelates iron; inhibits FPN; affects IRP/IRE system | Avoid in IDA; potential for iron redistribution therapy | [81] |
| Epicatechin | Tea, cocoa, apples | Promotes bioavailability (in vitro) | May increase Fe2+ availability | Potentially beneficial in iron deficiency | [88] |
| Kaempferol | Kale, beans, tea, broccoli | Promotes bioavailability (in vitro) | May increase Fe2+ availability | Potentially beneficial in iron deficiency | [88] |
| Myricetin | Berries, grapes, tea | Inhibits hepcidin expression | Inhibits BMP/SMAD pathway | May counteract hepcidin overexpression | [85] |
| Genistein | Soybeans | Increases hepcidin | Activates hepcidin expression (unknown pathway) | Useful in iron overload disorders | [83] |
| Ipriflavone | Synthetic (from daidzein) | Increases hepcidin | Synthetic analog, promotes hepcidin | Possible role in iron metabolism disorders | [84] |
| Icariin | Epimedium (Chinese herbs) | Increases hepcidin | Activates Stat3 and Smad1/5/8 pathways | Regulates iron homeostasis; potential therapeutic use | [89] |
3.2. Promoters of Iron Absorption in Food
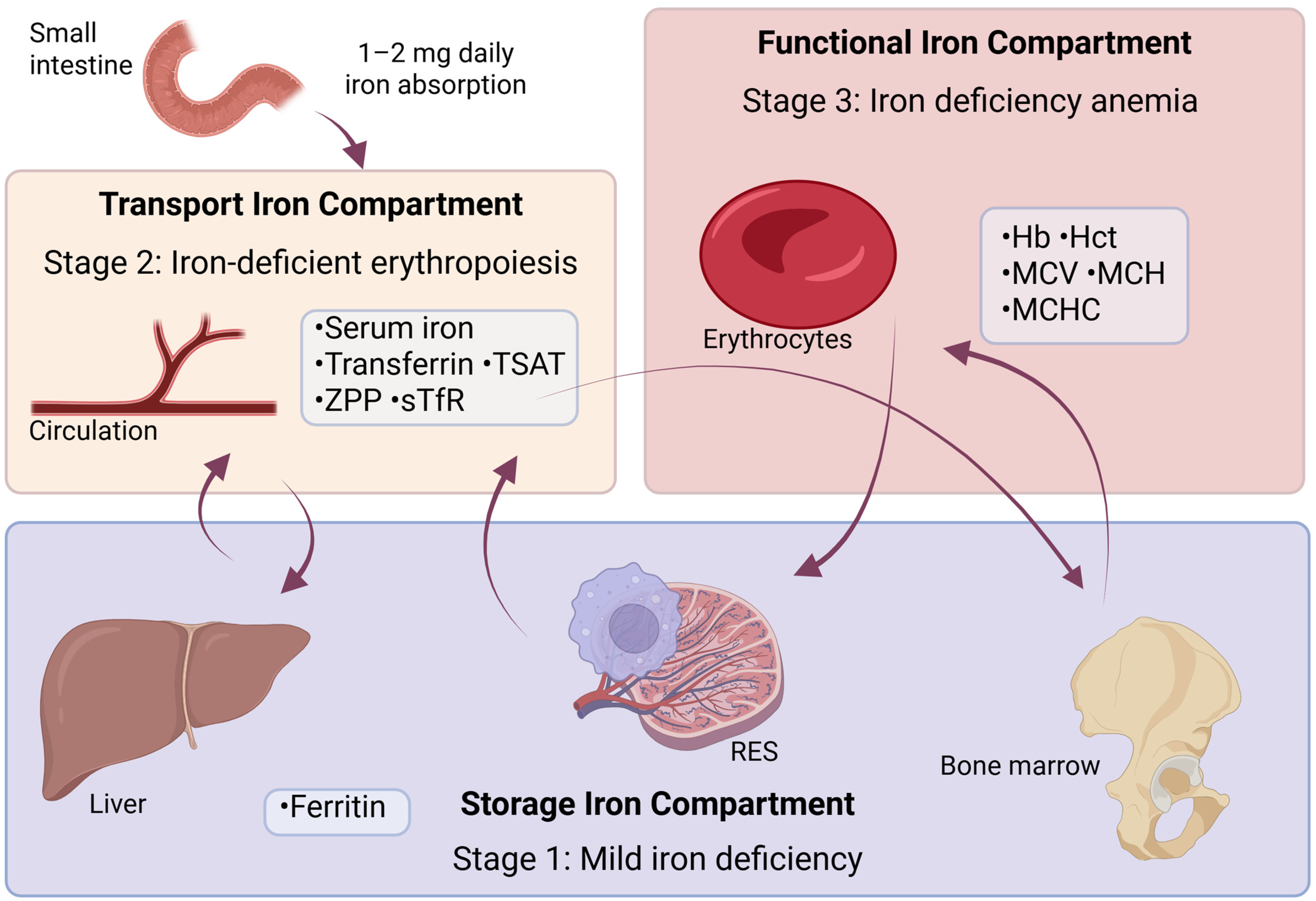
4. Diagnostic Indicators of ID and Anemia
| Stage I | Stage II | Stage III | Compartment | |
|---|---|---|---|---|
| Ferritin | ↓ | ↓↓ | ↓↓ | Storage |
| Iron | N | ↓ | ↓ | Transport |
| Transferrin | N | ↑ | ↑ | Transport |
| TIBC | N | ↑ | ↑ | Transport |
| TSAT | N | ↓ | ↓ | Transport |
| sTfR | N | ↑ | ↑ | Transport |
| ZPP | N | ↑ | ↑ | Transport |
| Hb | N | N | ↓ | Functional |
| MCV | N | N | ↓ | Functional |
| RDW | N | N | ↑ | Functional |
| Hepcidin | N | ↓ | ↓ | NA |
Diagnostic Challenges in Anemia of Inflammation
5. Addressing Iron Deficiency: Replacement Therapy and Prevention
5.1. Changes in Eating Habits
Region-Specific Strategies for Improving Iron Nutrition
| Food Group | Examples | Key Health Benefits | Protective Components/Notes | References |
|---|---|---|---|---|
| Dark-green leafy vegetables | Spinach, kale, collards | Cardiovascular health, cancer prevention, folic acid | High in folic acid, antioxidants (carotenoids, flavonoids) | [180] |
| source | ||||
| Cruciferous vegetables | Broccoli, cauliflower, Brussels sprouts | Cancer prevention | Contain dithiothiones, indoles, isothiocyanates | [181] |
| Yellow-orange vegetables | Carrots, sweet potatoes, pumpkin | Eye health, cancer prevention | Rich in carotenoids (beta-carotene) | [182] |
| Allium vegetables | Garlic, onions, leeks | Cancer prevention, cardiovascular benefits | Sulfur-containing compounds | [180] |
| Citrus fruits | Oranges, grapefruits, lemons | Immune support, cancer prevention, heart health | High in vitamin C, flavonoids, folic acid | [180] |
| Deep-yellow-orange fruits | Mango, papaya, apricot | Antioxidant support, cancer prevention | Carotenoids, vitamin C | [181] |
| General fruit intake | Apples, pears, bananas, berries | Cardiovascular health, cholesterol control, fiber source | High in fiber, antioxidants | [180] |
| Potassium-rich vegetables/fruits | Bananas, potatoes, leafy greens | Hypertension control | Potassium content | [180,183] |
| Fiber-rich fruits and vegetables | Most fruits and vegetables | Cholesterol control, diverticulosis prevention | Insoluble and soluble fiber | [180] |
5.2. Supplementation
5.3. Fortification
6. Recent Developments in Oral and Intravenous Therapies for Iron Deficiency
6.1. Oral Iron Therapies
6.2. Intravenous Iron Therapies
7. Conclusions
8. Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ID | iron deficiency |
| IDA | iron deficiency anemia |
| Hb | hemoglobin |
| WHO | World Health Organization |
| NHANES | National Health and Nutrition Examination Survey |
| GI | gastrointestinal |
| IBD | inflammatory bowel disease |
| SF | serum ferritin |
| UNICEF | United Nations International Children’s Emergency Fund |
| AE | anemia in the elderly |
| AI | anemia of inflammation |
| CKD | chronic kidney disease |
| IRIDA | iron-refractory iron deficiency anemia |
| YLDs | years lived with disability |
| ROS | reactive oxygen species |
| Dcytb | duodenal cytochrome b |
| DMT-1 | divalent metal transporter 1 |
| FPN | ferroportin |
| Heph | hephaestin |
| Tf | transferrin |
| sTfR | soluble transferrin receptor |
| TIBC | total iron-binding capacity |
| FBC | full blood count |
| MCV | mean corpuscular volume |
| MCH | mean corpuscular hemoglobin |
| MCHC | mean corpuscular hemoglobin concentration |
| RDW | red cell distribution width |
| NTBI | non-transferrin-bound iron |
| CRP | C-reactive protein |
| EHA | European Hematology Association |
| CDC | Centers for Disease Control and Prevention |
| Fe-ASP | iron conjugated with N-aspartyl-casein |
| IV | intravenous |
| FCM | ferric carboxymaltose |
| FDI | ferric derisomaltose |
| HFrEF | heart failure with reduced ejection fraction |
| ERFE | erythroferrone |
| BMP | bone morphogenetic protein |
| HIF-2α | hypoxia-inducible factor 2 alpha |
| HRI | heme-regulated inhibitor kinase |
| IREs | iron-responsive elements |
| UTRs | untranslated regions |
| CNN | convolutional neural networks |
References
- Pasricha, S.R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron Deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef]
- Abioye, A.I.; Andersen, C.T.; Sudfeld, C.R.; Fawzi, W.W. Anemia, Iron Status, and HIV: A Systematic Review of the Evidence. Adv. Nutr. 2020, 11, 1334–1363. [Google Scholar] [CrossRef] [PubMed]
- Iron Deficiency without Anemia: Indications for Treatment–GREM–Gynecological and Reproductive Endocrinology & Metabolism. Available online: https://gremjournal.com/journal/04-2020/iron-deficiency-without-anemia-indications-for-treatment/ (accessed on 6 May 2025).
- Zimmermann, M.B.; Hurrell, R.F. Nutritional Iron Deficiency. Lancet 2007, 370, 511–520. [Google Scholar] [CrossRef] [PubMed]
- WHO/UNICEF Discussion Paper: The Extension of the 2025 Maternal, Infant and Young Child Nutrition Targets to 2030–UNICEF DATA. Available online: https://data.unicef.org/resources/who-unicef-discussion-paper-nutrition-targets/ (accessed on 6 May 2025).
- Kassebaum, N.J.; Jasrasaria, R.; Naghavi, M.; Wulf, S.K.; Johns, N.; Lozano, R.; Regan, M.; Weatherall, D.; Chou, D.P.; Eisele, T.P.; et al. A Systematic Analysis of Global Anemia Burden from 1990 to 2010. Blood 2014, 123, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Gardner, W.M.; Razo, C.; McHugh, T.A.; Hagins, H.; Vilchis-Tella, V.M.; Hennessy, C.; Taylor, H.J.; Perumal, N.; Fuller, K.; Cercy, K.M.; et al. Prevalence, Years Lived with Disability, and Trends in Anaemia Burden by Severity and Cause, 1990–2021: Findings from the Global Burden of Disease Study 2021. Lancet Haematol. 2023, 10, e713–e734. [Google Scholar] [CrossRef]
- Lemoine, A.; Tounian, P. Childhood Anemia and Iron Deficiency in Sub-Saharan Africa–Risk Factors and Prevention: A Review. Arch. Pédiatr 2020, 27, 490–496. [Google Scholar] [CrossRef]
- Rakanita, Y.; Sinuraya, R.K.; Suradji, E.W.; Suwantika, A.A.; Syamsunarno, M.R.A.; Abdulah, R. The Challenges in Eradication of Iron Deficiency Anemia in Developing Countries. Syst. Rev. Pharm. 2020, 11, 383–401. [Google Scholar] [CrossRef]
- Levi, M.; Rosselli, M.; Simonetti, M.; Brignoli, O.; Cancian, M.; Masotti, A.; Pegoraro, V.; Cataldo, N.; Heiman, F.; Chelo, M.; et al. Epidemiology of Iron Deficiency Anaemia in Four European Countries: A Population-Based Study in Primary Care. Eur. J. Haematol. 2016, 97, 583–593. [Google Scholar] [CrossRef]
- Jefferds, M.E.D.; Mei, Z.; Addo, Y.; Hamner, H.C.; Perrine, C.G.; Flores-Ayala, R.; Pfeiffer, C.M.; Sharma, A.J. Iron Deficiency in the United States: Limitations in Guidelines, Data, and Monitoring of Disparities. Am. J. Public. Health 2022, 112, S826–S835. [Google Scholar] [CrossRef]
- Hercberg, S.; Preziosi, P.; Galan, P. Iron Deficiency in Europe. Public. Health Nutr. 2001, 4, 537–545. [Google Scholar] [CrossRef]
- Knowles, J.; Walters, T.; Yarparvar, A.; Brown, R. A Review of Anemia Prevalence, and Prevention and Control Strategies, in the Eastern Europe and Central Asia Region. Curr. Dev. Nutr. 2024, 8, 104477. [Google Scholar] [CrossRef]
- Milman, N.; Taylor, C.L.; Merkel, J.; Brannon, P.M. Iron Status in Pregnant Women and Women of Reproductive Age in Europe. Am. J. Clin. Nutr. 2017, 106, 1655S–1662S. [Google Scholar] [CrossRef]
- Anaemia in Women and Children. Available online: https://www.who.int/data/gho/data/themes/topics/anaemia_in_women_and_children (accessed on 7 May 2025).
- Eussen, S.; Alles, M.; Uijterschout, L.; Brus, F.; Van Der Horst-Graat, J. Iron Intake and Status of Children Aged 6–36 Months in Europe: A Systematic Review. Ann. Nutr. Metab. 2015, 66, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Tijanić, I.R. Solubilni Transferenski Receptori u Dijagnostici Anemije Zbog Deficita Gvožđa. Ph.D. Thesis, University of Niš, Niš, Serbia, 2014. [Google Scholar]
- Rakic, L.; Djokic, D.; Drakulovic, M.B.; Pejic, A.; Radojicic, Z.; Marinkovic, M. Risk Factors Associated with Anemia among Serbian Non-Pregnant Women 20 to 49 Years Old. A Cross-Sectional Study. Hippokratia 2013, 17, 47–54. [Google Scholar] [PubMed]
- Petrović, V. Zdravstveno Stanje Stanovništva AP Vojvodine 2019; Godine: Novi Sad, Serbia, 2020. [Google Scholar]
- de Benoist, B.; McLean, E.; Egli, I.; Cogswell, M. (Eds.) Worldwide Prevalence of Anaemia 1993–2005: WHO Global Database on Anaemia; World Health Organization: Geneva, Switzerland, 2008; 40p. [Google Scholar]
- Camaschella, C. Iron Deficiency. Blood 2019, 133, 30–39. [Google Scholar] [CrossRef]
- Bonovas, S.; Fiorino, G.; Allocca, M.; Lytras, T.; Tsantes, A.; Peyrin-Biroulet, L.; Danese, S. Intravenous versus Oral Iron for the Treatment of Anemia in Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Medicine 2016, 95, e2308. [Google Scholar] [CrossRef]
- Haile, K.; Yemane, T.; Tesfaye, G.; Wolde, D.; Timerga, A.; Haile, A. Anemia and Its Association with Helicobacter Pylori Infection among Adult Dyspeptic Patients Attending Wachemo University Nigist Eleni Mohammad Memorial Referral Hospital, Southwest Ethiopia: A Cross-Sectional Study. PLoS ONE 2021, 16, e0245168. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, X.; Shen, Y.; Fang, X.; Wang, Y.; Wang, F. Obesity and Iron Deficiency: A Quantitative Meta-Analysis. Obes. Rev. 2015, 16, 1081–1093. [Google Scholar] [CrossRef]
- Bardou-Jacquet, E.; Island, M.L.; Jouanolle, A.M.; Détivaud, Ĺ.; Fatih, N.; Ropert, M.; Brissot, E.; Mosser, A.; Maisonneuve, H.; Brissot, P.; et al. A Novel N491S Mutation in the Human SLC11A2 Gene Impairs Protein Trafficking and in Association with the G212V Mutation Leads to Microcytic Anemia and Liver Iron Overload. Blood Cells Mol. Dis. 2011, 47, 243–248. [Google Scholar] [CrossRef]
- Sun, B.; Tan, B.; Zhang, P.; Zhu, L.; Wei, H.; Huang, T.; Li, C.; Yang, W. Iron Deficiency Anemia: A Critical Review on Iron Absorption, Supplementation and Its Influence on Gut Microbiota. Food Funct. 2024, 15, 1144–1157. [Google Scholar] [CrossRef]
- Althunibat, O.Y.; Saghir, S.A.M.; Aladaileh, S.H.; Rawadieh, A. The Impact of Weight Loss Diet Programs on Anemia, Nutrient Deficiencies, and Organ Dysfunction Markers among University Female Students: A Cross-Sectional Study. Electron. J. Gen. Med. 2023, 20, em436. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.E.; Vassallo, R.R. How Do We Manage Iron Deficiency after Blood Donation? Br. J. Haematol. 2018, 181, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Haider, L.M.; Schwingshackl, L.; Hoffmann, G.; Ekmekcioglu, C. The Effect of Vegetarian Diets on Iron Status in Adults: A Systematic Review and Meta-Analysis. Crit. Rev. Food Sci. Nutr. 2018, 58, 1359–1374. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, N.S.; Jaceldo-Siegl, K.; Sabate, J.; Fraser, G.E. Nutrient Profiles of Vegetarian and Nonvegetarian Dietary Patterns. J. Acad. Nutr. Diet. 2013, 113, 1610–1619. [Google Scholar] [CrossRef]
- Hall, N.J.; Rubin, G.; Charnock, A. Systematic Review: Adherence to a Gluten-Free Diet in Adult Patients with Coeliac Disease. Aliment. Pharmacol. Ther. 2009, 30, 315–330. [Google Scholar] [CrossRef]
- Gasche, C.; Lomer, M.C.E.; Cavill, I.; Weiss, G. Iron, Anaemia, and Inflammatory Bowel Diseases. Gut 2004, 53, 1190–1197. [Google Scholar] [CrossRef]
- McAfee, A.J.; McSorley, E.M.; Cuskelly, G.J.; Moss, B.W.; Wallace, J.M.W.; Bonham, M.P.; Fearon, A.M. Red Meat Consumption: An Overview of the Risks and Benefits. Meat Sci. 2010, 84, 1–13. [Google Scholar] [CrossRef]
- Al Hasan, S.M.; Hassan, M.; Saha, S.; Islam, M.; Billah, M.; Islam, S. Dietary Phytate Intake Inhibits the Bioavailability of Iron and Calcium in the Diets of Pregnant Women in Rural Bangladesh: A Cross-Sectional Study. BMC Nutr. 2016, 2, 24. [Google Scholar] [CrossRef]
- Simcox, J.A.; McClain, D.A. Iron and Diabetes Risk. Cell Metab. 2013, 17, 329–341. [Google Scholar] [CrossRef]
- Engert, A.; Balduini, C.; Brand, A.; Coiffier, B.; Cordonnier, C.; Döhner, H.; De Wit, T.D.; Eichinger, S.; Fibbe, W.; Green, T.; et al. The European Hematology Association Roadmap for European Hematology Research: A Consensus Document. Haematologica 2016, 101, 115–208. [Google Scholar] [CrossRef]
- Bach, V.; Schruckmayer, G.; Sam, I.; Kemmler, G.; Stauder, R. Prevalence and Possible Causes of Anemia in the Elderly: A Cross-Sectional Analysis of a Large European University Hospital Cohort. Clin. Interv. Aging 2014, 9, 1187–1196. [Google Scholar] [CrossRef]
- Girelli, D.; Marchi, G.; Camaschella, C. Anemia in the Elderly. Hemasphere 2018, 2, e40. [Google Scholar] [CrossRef]
- Awaluddin, S.M.; Shahein, N.A.; Rahim, N.C.A.; Zaki, N.A.M.; Nasaruddin, N.H.; Saminathan, T.A.; Alias, N.; Ganapathy, S.S.; Ahmad, N.A. Anemia among Men in Malaysia: A Population-Based Survey in 2019. Int. J. Environ. Res. Public Health 2021, 18, 10922. [Google Scholar] [CrossRef]
- Rohr, M.; Brandenburg, V.; Brunner-La Rocca, H.P. How to Diagnose Iron Deficiency in Chronic Disease: A Review of Current Methods and Potential Marker for the Outcome. Eur. J. Med. Res. 2023, 28, 15. [Google Scholar] [CrossRef]
- Larson, L.M.; Kubes, J.N.; Ramírez-Luzuriaga, M.J.; Khishen, S.; H Shankar, A.; Prado, E.L. Effects of Increased Hemoglobin on Child Growth, Development, and Disease: A Systematic Review and Meta-Analysis. Ann. N. Y. Acad. Sci. 2019, 1450, 83–104. [Google Scholar] [CrossRef]
- Fu, X.Y.; Xie, X.T. Association between Iron Deficiency and Brain Developmental Disorder in Children. Zhongguo Dang Dai Er Ke Za Zhi 2018, 20, 964–967. [Google Scholar] [CrossRef]
- Hua, M.; Shi, D.; Xu, W.; Zhu, L.; Hao, X.; Zhu, B.; Shu, Q.; Lozoff, B.; Geng, F.; Shao, J. Differentiation between Fetal and Postnatal Iron Deficiency in Altering Brain Substrates of Cognitive Control in Pre-Adolescence. BMC Med. 2023, 21, 167. [Google Scholar] [CrossRef]
- Baron, D.M.; Hochrieser, H.; Posch, M.; Metnitz, B.; Rhodes, A.; Moreno, R.P.; Pearse, R.M.; Metnitz, P. Preoperative Anaemia Is Associated with Poor Clinical Outcome in Non-Cardiac Surgery Patients. Br. J. Anaesth. 2014, 113, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Verdon, F.; Burnand, B.; Fallab Stubi, C.L.; Bonard, C.; Graff, M.; Michaud, A.; Bischoff, T.; De Vevey, M.; Studer, J.P.; Herzig, L.; et al. Iron Supplementation for Unexplained Fatigue in Non-Anaemic Women: Double Blind Randomised Placebo Controlled Trial. BMJ 2003, 326, 1124. [Google Scholar] [CrossRef] [PubMed]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on Iron and Its Importance for Human Health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar] [PubMed]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and Regulatory Aspects of Intestinal Iron Absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, 397–409. [Google Scholar] [CrossRef]
- Schrier, S.L. So You Know How to Treat Iron Deficiency Anemia. Blood 2015, 126, 1971. [Google Scholar] [CrossRef] [PubMed]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron Fortification Adversely Affects the Gut Microbiome, Increases Pathogen Abundance and Induces Intestinal Inflammation in Kenyan Infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Kortman, G.A.M.; Raffatellu, M.; Swinkels, D.W.; Tjalsma, H. Nutritional Iron Turned inside out: Intestinal Stress from a Gut Microbial Perspective. FEMS Microbiol. Rev. 2014, 38, 1202–1234. [Google Scholar] [CrossRef]
- Swanson, C.A. Iron Intake and Regulation: Implications for Iron Deficiency and Iron Overload. Alcohol 2003, 30, 99–102. [Google Scholar] [CrossRef]
- Piskin, E.; Cianciosi, D.; Gulec, S.; Tomas, M.; Capanoglu, E. Iron Absorption: Factors, Limitations, and Improvement Methods. ACS Omega 2022, 7, 20441–20456. [Google Scholar] [CrossRef]
- Ganz, T. Cellular Iron: Ferroportin Is the Only Way Out. Cell Metab. 2005, 1, 155–157. [Google Scholar] [CrossRef]
- Anderson, G.J.; Vulpe, C.D. Mammalian Iron Transport. Cell. Mol. Life Sci. 2009, 66, 3241–3261. [Google Scholar] [CrossRef]
- Brissot, P.; Ropert, M.; Le Lan, C.; Loréal, O. Non-Transferrin Bound Iron: A Key Role in Iron Overload and Iron Toxicity. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2012, 1820, 403–410. [Google Scholar] [CrossRef]
- Waldvogel-Abramowski, S.; Waeber, G.; Gassner, C.; Buser, A.; Frey, B.M.; Favrat, B.; Tissot, J.D. Physiology of Iron Metabolism. Transfus. Med. Hemother. 2014, 41, 213–221. [Google Scholar] [CrossRef]
- Sharp, P.; Srai, S.K. Molecular Mechanisms Involved in Intestinal Iron Absorption. World J. Gastroenterol. 2007, 13, 4716–4724. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Ganz, T. Regulation of Iron Metabolism by Hepcidin. Annu. Rev. Nutr. 2006, 26, 323–342. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, C.C.; Skaar, E.P. Nutritional Immunity: The Battle for Nutrient Metals at the Host–Pathogen Interface. Nat. Rev. Microbiol. 2022, 20, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Hood, M.I.; Skaar, E.P. Nutritional Immunity: Transition Metals at the Pathogen-Host Interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef]
- Aschemeyer, S.; Qiao, B.; Stefanova, D.; Valore, E.V.; Sek, A.C.; Alex Ruwe, T.; Vieth, K.R.; Jung, G.; Casu, C.; Rivella, S.; et al. Structure-Function Analysis of Ferroportin Defines the Binding Site and an Alternative Mechanism of Action of Hepcidin. Blood 2018, 131, 899–910. [Google Scholar] [CrossRef]
- Kautz, L.; Meynard, D.; Monnier, A.; Darnaud, V.; Bouvet, R.; Wang, R.H.; Deng, C.; Vaulont, S.; Mosser, J.; Coppin, H.; et al. Iron Regulates Phosphorylation of Smad1/5/8 and Gene Expression of Bmp6, Smad7, Id1, and Atoh8 in the Mouse Liver. Blood 2008, 112, 1503–1509. [Google Scholar] [CrossRef]
- Silvestri, L.; Pagani, A.; Nai, A.; De Domenico, I.; Kaplan, J.; Camaschella, C. The Serine Protease Matriptase-2 (TMPRSS6) Inhibits Hepcidin Activation by Cleaving Membrane Hemojuvelin. Cell Metab. 2008, 8, 502–511. [Google Scholar] [CrossRef]
- Pasricha, S.R.; Lim, P.J.; Duarte, T.L.; Casu, C.; Oosterhuis, D.; Mleczko-Sanecka, K.; Suciu, M.; Da Silva, A.R.; Al-Hourani, K.; Arezes, J.; et al. Hepcidin Is Regulated by Promoter-Associated Histone Acetylation and HDAC3. Nat. Commun. 2017, 8, 403. [Google Scholar] [CrossRef]
- Kautz, L.; Jung, G.; Valore, E.V.; Rivella, S.; Nemeth, E.; Ganz, T. Identification of Erythroferrone as an Erythroid Regulator of Iron Metabolism. Nat. Genet. 2014, 46, 678–684. [Google Scholar] [CrossRef]
- Mastrogiannaki, M.; Matak, P.; Peyssonnaux, C. The Gut in Iron Homeostasis: Role of HIF-2 under Normal and Pathological Conditions. Blood 2013, 122, 885–892. [Google Scholar] [CrossRef]
- Zhang, D.L.; Wu, J.; Shah, B.N.; Greutélaers, K.C.; Ghosh, M.C.; Ollivierre, H.; Su, X.Z.; Thuma, P.E.; Bedu-Addo, G.; Mockenhaupt, F.P.; et al. Erythrocytic Ferroportin Reduces Intracellular Iron Accumulation, Hemolysis, and Malaria Risk. Science 2018, 359, 1520–1523. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.L.; Ghosh, M.C.; Rouault, T.A. The Physiological Functions of Iron Regulatory Proteins in Iron Homeostasis–an Update. Front. Pharmacol. 2014, 5, 124. [Google Scholar] [CrossRef] [PubMed]
- Bayeva, M.; Khechaduri, A.; Puig, S.; Chang, H.C.; Patial, S.; Blackshear, P.J.; Ardehali, H. MTOR Regulates Cellular Iron Homeostasis through Tristetraprolin. Cell Metab. 2012, 16, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Mancias, J.D.; Wang, X.; Gygi, S.P.; Harper, J.W.; Kimmelman, A.C. Quantitative Proteomics Identifies NCOA4 as the Cargo Receptor Mediating Ferritinophagy. Nature 2014, 509, 105–109. [Google Scholar] [CrossRef]
- Richardson, C.L.; Delehanty, L.L.; Bullock, G.C.; Rival, C.M.; Tung, K.S.; Kimpel, D.L.; Gardenghi, S.; Rivella, S.; Goldfarb, A.N. Isocitrate Ameliorates Anemia by Suppressing the Erythroid Iron Restriction Response. J. Clin. Investig. 2013, 123, 3614–3623. [Google Scholar] [CrossRef]
- Nai, A.; Lidonnici, M.R.; Rausa, M.; Mandelli, G.; Pagani, A.; Silvestri, L.; Ferrari, G.; Camaschella, C. The Second Transferrin Receptor Regulates Red Blood Cell Production in Mice. Blood 2015, 125, 1170–1179. [Google Scholar] [CrossRef]
- Sonnweber, T.; Nachbaur, D.; Schroll, A.; Nairz, M.; Seifert, M.; Demetz, E.; Haschka, D.; Mitterstiller, A.M.; Kleinsasser, A.; Burtscher, M.; et al. Hypoxia Induced Downregulation of Hepcidin Is Mediated by Platelet Derived Growth Factor BB. Gut 2014, 63, 1951–1959. [Google Scholar] [CrossRef]
- Hurrell, R.; Egli, I. Iron Bioavailability and Dietary Reference Values. Am. J. Clin. Nutr. 2010, 91, 1461S–1467S. [Google Scholar] [CrossRef]
- Gibson, R.S.; Bailey, K.B.; Gibbs, M.; Ferguson, E.L. A Review of Phytate, Iron, Zinc, and Calcium Concentrations in Plant-Based Complementary Foods Used in Low-Income Countries and Implications for Bioavailability. Food Nutr. Bull. 2010, 31, S134–S146. [Google Scholar] [CrossRef]
- Cook, J.D.; Dassenko, S.A.; Lynch, S.R. Assessment of the Role of Nonheme-Iron Availability in Iron Balance. Am. J. Clin. Nutr. 1991, 54, 717–722. [Google Scholar] [CrossRef]
- Hoppe, M.; Ross, A.B.; Svelander, C.; Sandberg, A.S.; Hulthén, L. Low-Phytate Wholegrain Bread Instead of High-Phytate Wholegrain Bread in a Total Diet Context Did Not Improve Iron Status of Healthy Swedish Females: A 12-Week, Randomized, Parallel-Design Intervention Study. Eur. J. Nutr. 2019, 58, 853–864. [Google Scholar] [CrossRef]
- Armah, S.M.; Boy, E.; Chen, D.; Candal, P.; Reddy, M.B. Regular Consumption of a High-Phytate Diet Reduces the Inhibitory Effect of Phytate on Nonheme-Iron Absorption in Women with Suboptimal Iron Stores. J. Nutr. 2015, 145, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health Benefits of Polyphenols: A Concise Review. J. Food Biochem. 2022, 46, e14264. [Google Scholar] [CrossRef] [PubMed]
- Lesjak, M.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.S. Quercetin Inhibits Intestinal Non-Haem Iron Absorption by Regulating Iron Metabolism Genes in the Tissues. Eur. J. Nutr. 2019, 58, 743–753. [Google Scholar] [CrossRef]
- Lesjak, M.; Hoque, R.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.S.; Sharp, P.A. Quercetin Inhibits Intestinal Iron Absorption and Ferroportin Transporter Expression In Vivo and In Vitro. PLoS ONE 2014, 9, e102900. [Google Scholar] [CrossRef]
- Dijiong, W.; Xiaowen, W.; Linlong, X.; Wenbin, L.; Huijin, H.; Baodong, Y.; Yuhong, Z. Iron Chelation Effect of Curcumin and Baicalein on Aplastic Anemia Mouse Model with Iron Overload. Iran. J. Basic. Med. Sci. 2019, 22, 660. [Google Scholar] [CrossRef]
- Zhen, A.W.; Nguyen, N.H.; Gibert, Y.; Motola, S.; Buckett, P.; Wessling-Resnick, M.; Fraenkel, E.; Fraenkel, P.G. The Small Molecule, Genistein, Increases Hepcidin Expression in Human Hepatocytes. Hepatology 2013, 58, 1315–1325. [Google Scholar] [CrossRef]
- Patchen, B.; Koppe, T.; Cheng, A.; Seo, Y.A.; Wessling-Resnick, M.; Fraenkel, P.G. Dietary Supplementation with Ipriflavone Decreases Hepatic Iron Stores in Wild Type Mice. Blood Cells Mol. Dis. 2016, 60, 36–43. [Google Scholar] [CrossRef]
- Mu, M.; An, P.; Wu, Q.; Shen, X.; Shao, D.; Wang, H.; Zhang, Y.; Zhang, S.; Yao, H.; Min, J.; et al. The Dietary Flavonoid Myricetin Regulates Iron Homeostasis by Suppressing Hepcidin Expression. J. Nutr. Biochem. 2016, 30, 53–61. [Google Scholar] [CrossRef]
- Bayele, H.K.; Balesaria, S.; Srai, S.K.S. Phytoestrogens Modulate Hepcidin Expression by Nrf2: Implications for Dietary Control of Iron Absorption. Free Radic. Biol. Med. 2015, 89, 1192–1202. [Google Scholar] [CrossRef]
- Vanhees, K.; Godschalk, R.W.; Sanders, A.; Van Waalwijk van Doorn-Khosrovani, S.B.; Van Schooten, F.J. Maternal Quercetin Intake during Pregnancy Results in an Adapted Iron Homeostasis at Adulthood. Toxicology 2011, 290, 350–358. [Google Scholar] [CrossRef]
- Hart, J.J.; Tako, E.; Kochian, L.V.; Glahn, R.P. Identification of Black Bean (Phaseolus vulgaris L.) Polyphenols That Inhibit and Promote Iron Uptake by Caco-2 Cells. J. Agric. Food Chem. 2015, 63, 5950–5956. [Google Scholar] [CrossRef]
- Lesjak, M.; Srai, S.K.S. Role of Dietary Flavonoids in Iron Homeostasis. Pharmaceuticals 2019, 12, 119. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Kim, E.Y.; Lindsay, E.A.; Han, O. Bioactive Dietary Polyphenols Inhibit Heme Iron Absorption in a Dose-Dependent Manner in Human Intestinal Caco-2 Cells. J. Food Sci. 2011, 76, H143–H150. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, L.; Rossander-Hulthèn, L.; Brune, M.; Gleerup, A. Inhibition of Haem-Iron Absorption in Man by Calcium. Br. J. Nutr. 1993, 69, 533–540. [Google Scholar] [CrossRef]
- Roughead, Z.K.; Zito, C.A.; Hunt, J.R. Inhibitory Effects of Dietary Calcium on the Initial Uptake and Subsequent Retention of Heme and Nonheme Iron in Humans: Comparisons Using an Intestinal Lavage Method. Am. J. Clin. Nutr. 2005, 82, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Gaitán, D.; Flores, S.; Saavedra, P.; Miranda, C.; Olivares, M.; Arredondo, M.; de Romaña, D.L.; Lönnerdal, B.; Pizarro, F. Calcium Does Not Inhibit the Absorption of 5 Milligrams of Nonheme or Heme Iron at Doses Less Than 800 Milligrams in Nonpregnant Women. J. Nutr. 2011, 141, 1652–1656. [Google Scholar] [CrossRef]
- Henare, S.J.; Nur Singh, N.; Ellis, A.M.; Moughan, P.J.; Thompson, A.K.; Walczyk, T. Iron Bioavailability of a Casein-Based Iron Fortificant Compared with That of Ferrous Sulfate in Whole Milk: A Randomized Trial with a Crossover Design in Adult Women. Am. J. Clin. Nutr. 2019, 110, 1362–1369. [Google Scholar] [CrossRef]
- Cámara-Martos, F.; Amaro-López, M.A. Influence of Dietary Factors on Calcium Bioavailability: A Brief Review. Biol. Trace Elem. Res. 2002, 89, 43–52. [Google Scholar] [CrossRef]
- Hurrell, R.F.; Lynch, S.R.; Trinidad, T.P.; Dassenko, S.A.; Cook, J.D. Iron Absorption in Humans: Bovine Serum Albumin Compared with Beef Muscle and Egg White. Am. J. Clin. Nutr. 1988, 47, 102–107. [Google Scholar] [CrossRef]
- Awoniyi, A.; Daniel, O.; Babatunde, O.; Awoniyi, A.; Daniel, O.; Babatunde, O. Dietary Iron Uptake and Absorption. In Metabolism Annual Volume 2024; IntechOpen: London, UK, 2024. [Google Scholar] [CrossRef]
- Khoja, K.K.; Aslam, M.F.; Sharp, P.A.; Latunde-Dada, G.O. In Vitro Bioaccessibility and Bioavailability of Iron from Fenugreek, Baobab and Moringa. Food Chem. 2021, 335, 127671. [Google Scholar] [CrossRef]
- He, W.; Li, X.; Ding, K.; Li, Y.; Li, W. Ascorbic Acid Can Reverse the Inhibition of Phytic Acid, Sodium Oxalate and Sodium Silicate on Iron Absorption in Caco-2 Cells. Int. J. Vitam. Nutr. Res. 2019, 88, 65–72. [Google Scholar] [CrossRef]
- Villaño, D.; Vilaplana, C.; Medina, S.; Algaba-Chueca, F.; Cejuela-Anta, R.; Martínez-Sanz, J.M.; Ferreres, F.; Gil-Izquierdo, A. Relationship between the Ingestion of a Polyphenol-Rich Drink, Hepcidin Hormone, and Long-Term Training. Molecules 2016, 21, 1333. [Google Scholar] [CrossRef]
- Mao, X.; Yao, G. Effect of Vitamin C Supplementations on Iron Deficiency Anemia in Chinese Children. Biomed. Environ. Sci. 1992, 5, 125–129. [Google Scholar]
- Teucher, B.; Olivares, M.; Cori, H. Enhancers of Iron Absorption: Ascorbic Acid and Other Organic Acids. Int. J. Vitam. Nutr. Res. 2004, 74, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.D.; Monsen, E.R. Vitamin C, the Common Cold, and Iron Absorption. Am. J. Clin. Nutr. 1977, 30, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.D.; Reddy, M.B. Effect of Ascorbic Acid Intake on Nonheme-Iron Absorption from a Complete Diet. Am. J. Clin. Nutr. 2001, 73, 93–98. [Google Scholar] [CrossRef]
- Layrisse, M.; Martínez-Torres, C.; Roche, M. Effect of Interaction of Various Foods on Iron Absorption. Am. J. Clin. Nutr. 1968, 21, 1175–1183. [Google Scholar] [CrossRef]
- Bœch, S.B.; Hansen, M.; Bukhave, K.; Jensen, M.; Sørensen, S.S.; Kristensen, L.; Purslow, P.P.; Skibsted, L.H.; Sandström, B. Nonheme-Iron Absorption from a Phytate-Rich Meal Is Increased by the Addition of Small Amounts of Pork Meat. Am. J. Clin. Nutr. 2003, 77, 173–179. [Google Scholar] [CrossRef]
- Navas-Carretero, S.; Pérez-Granados, A.M.; Sarriá, B.; Vaquero, M.P.; Carbajal, A.; Pedrosa, M.M.; Roe, M.A.; Fairweather-Tait, S.J. Oily Fish Increases Iron Bioavailability of a Phytate Rich Meal in Young Iron Deficient Women. J. Am. Coll. Nutr. 2008, 27, 96–101. [Google Scholar] [CrossRef]
- O’Flaherty, E.A.A.; Tsermoula, P.; O’Neill, E.E.; O’Brien, N.M. Co-Products of Beef Processing Enhance Non-Haem Iron Absorption in an in Vitro Digestion/Caco-2 Cell Model. Int. J. Food Sci. Technol. 2019, 54, 1256–1264. [Google Scholar] [CrossRef]
- Hurrell, R.F.; Reddy, M.B.; Juillerat, M.; Cook, J.D. Meat Protein Fractions Enhance Nonheme Iron Absorption in Humans. J. Nutr. 2006, 136, 2808–2812. [Google Scholar] [CrossRef] [PubMed]
- Armah, C.N.; Sharp, P.; Mellon, F.A.; Pariagh, S.; Lund, E.K.; Dainty, J.R.; Teucher, B.; Fairweather-Tait, S.J. L-α-Glycerophosphocholine Contributes to Meat’s Enhancement of Nonheme Iron Absorption. J. Nutr. 2008, 138, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Jeroense, F.M.D.; Michel, L.; Zeder, C.; Herter-Aeberli, I.; Zimmermann, M.B. Consumption of Galacto-Oligosaccharides Increases Iron Absorption from Ferrous Fumarate: A Stable Iron Isotope Study in Iron-Depleted Young Women. J. Nutr. 2019, 149, 738–746. [Google Scholar] [CrossRef]
- Giorgetti, A.; Husmann, F.M.D.; Zeder, C.; Herter-Aeberli, I.; Zimmermann, M.B. Prebiotic Galacto-Oligosaccharides and Fructo-Oligosaccharides, but Not Acacia Gum, Increase Iron Absorption from a Single High-Dose Ferrous Fumarate Supplement in Iron-Depleted Women. J. Nutr. 2022, 152, 1015–1021. [Google Scholar] [CrossRef]
- Ahmad, A.M.R.; Ahmed, W.; Iqbal, S.; Javed, M.; Rashid, S. Iahtisham-ul-Haq Prebiotics and Iron Bioavailability? Unveiling the Hidden Association–A Review. Trends Food Sci. Technol. 2021, 110, 584–590. [Google Scholar] [CrossRef]
- Yeung, C.K.; Glahn, R.P.; Welch, R.M.; Miller, D.D. Prebiotics and Iron Bioavailability—Is There a Connection? J. Food Sci. 2005, 70, R88–R92. [Google Scholar] [CrossRef]
- Husmann, F.M.D.; Zimmermann, M.B.; Herter-Aeberli, I. The Effect of Prebiotics on Human Iron Absorption: A Review. Adv. Nutr. 2022, 13, 2296–2304. [Google Scholar] [CrossRef]
- Christides, T.; Sharp, P. Sugars Increase Non-Heme Iron Bioavailability in Human Epithelial Intestinal and Liver Cells. PLoS ONE 2013, 8, e83031. [Google Scholar] [CrossRef]
- WHO. Haemoglobin Concentrations for the Diagnosis of Anaemia and Assessment of Severity. In Vitamin and Mineral Nutrition Information System; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Goddard, A.F.; James, M.W.; McIntyre, A.S.; Scott, B.B. Guidelines for the Management of Iron Deficiency Anaemia. Gut 2011, 60, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Hastka, J.; Lasserre, J.J.; Schwarzbeck, A.; Reiter, A.; Hehlmann, R. Laboratory Tests of Iron Status: Correlation or Common Sense? Clin. Chem. 1996, 42, 718–724. [Google Scholar] [CrossRef]
- Guyatt, G.H.; Oxman, A.D.; Ali, M.; Willan, A.; McIlroy, W.; Patterson, C. Laboratory Diagnosis of Iron-Deficiency Anemia–An Overview. J. Gen. Intern. Med. 1992, 7, 145–153. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Serum Ferritin Concentrations for the Assessment of Iron Status and Iron Deficiency in Populations; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Sofiantin, N.; Kurniawan, L.B.; Arif, M. Analysis of Ferritin Levels, TIBC and Fe Serum In Central Obesity And Non Central Obesity. Str. J. Ilm. Kesehat. 2021, 10, 1265–1271. Available online: https://www.sjik.org/index.php/sjik/article/view/691 (accessed on 15 May 2025).
- Khan, A.; Khan, W.M.; Ayub, M.; Humayun, M.; Haroon, M. Ferritin Is a Marker of Inflammation Rather than Iron Deficiency in Overweight and Obese People. J. Obes. 2016, 2016, 1937320. [Google Scholar] [CrossRef]
- Coenen, J.L.L.M.; Van Dieijen-Visser, M.P.; Van Pelt, J.; Van Deursen, C.T.B.M.; Fickers, M.M.F.; Van Wersch, J.W.J.; Brombacher, P.J. Measurements of Serum Ferritin Used to Predict Concentrations of Iron in Bone Marrow in Anemia of Chronic Disease. Clin. Chem. 1991, 37, 560–563. [Google Scholar] [CrossRef]
- Pfeiffer, C.M.; Looker, A.C. Laboratory Methodologies for Indicators of Iron Status: Strengths, Limitations, and Analytical Challenges. Am. J. Clin. Nutr. 2017, 106, 1606S–1614S. [Google Scholar] [CrossRef]
- Auerbach, M.; Adamson, J.W. How We Diagnose and Treat Iron Deficiency Anemia. Am. J. Hematol. 2016, 91, 31–38. [Google Scholar] [CrossRef]
- Fletcher, A.; Forbes, A.; Svenson, N.; Wayne Thomas, D. Guideline for the Laboratory Diagnosis of Iron Deficiency in Adults (Excluding Pregnancy) and Children. J. Haematol. 2022, 196, 523–529. [Google Scholar] [CrossRef]
- Wish, J.B. Assessing Iron Status: Beyond Serum Ferritin and Transferrin Saturation. J. Am. Soc. Nephrol. 2006, 1, S4–S8. [Google Scholar] [CrossRef]
- Cook, J.D. Diagnosis and Management of Iron-Deficiency Anaemia. In Best Practice & Research Clinical Haematology; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Daude, S.; Remen, T.; Chateau, T.; Danese, S.; Gastin, I.; Baumann, C.; Gueant, J.L.; Peyrin-Biroulet, L. Comparative Accuracy of Ferritin, Transferrin Saturation and Soluble Transferrin Receptor for the Diagnosis of Iron Deficiency in Inflammatory Bowel Disease. Aliment. Pharmacol Ther. 2020, 51, 1087–1095. [Google Scholar] [CrossRef]
- World Health Organization. Assessing the Iron Status of Populations, 2nd ed.; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Graham, E.A.; Felgenhauer, J.; Detter, J.C.; Labbe, R.F. Elevated Zinc Protoporphyrin Associated with Thalassemia Trait and Hemoglobin E. J. Pediatr. 1996, 129, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Asobayire, F.S.; Adou, P.; Davidsson, L.; Cook, J.D.; Hurrell, R.F. Prevalence of Iron Deficiency with and without Concurrent Anemia in Population Groups with High Prevalences of Malaria and Other Infections: A Study in Côte d’Ivoire. Am. J. Clin. Nutr. 2001, 74, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W. Sensitivity, Specificity, and Predictive Value of Serum Soluble Transferrin Receptor at Different Stages of Iron Deficiency. Ann. Clin. Lab. Sci. 2005, 35, 435–439. Available online: https://www.annclinlabsci.org/content/35/4/435.long (accessed on 20 May 2025). [PubMed]
- Shin, D.H.; Kim, H.S.; Park, M.J.; Suh, I.B.; Shin, K.S. Utility of Access Soluble Transferrin Receptor (sTfR) and sTfR/log Ferritin Index in Diagnosing Iron Deficiency Anemia. Ann. Clin. Lab. Sci. 2015, 45, 396–402. Available online: https://www.annclinlabsci.org/content/45/4/396.long (accessed on 20 May 2025). [PubMed]
- Cook, J.D.; Flowers, C.H.; Skikne, B.S. The Quantitative Assessment of Body Iron. Blood 2003, 101, 3359–3363. [Google Scholar] [CrossRef]
- Bermejo, F.; García-López, S. A Guide to Diagnosis of Iron Deficiency and Iron Deficiency Anemia in Digestive Diseases. World J. Gastroenterol. 2009, 15, 4638–4643. [Google Scholar] [CrossRef]
- Sultana, G.S.; Haque, S.A.; Sultana, T.; An, A. Value of Red Cell Distribution Width (RDW) and RBC Indices in the Detection of Iron Deficiency Anemia. Mymensingh Med. J. 2013, 22, 370–376. [Google Scholar]
- Cheng, P.P.; Jiao, X.Y.; Wang, X.H.; Lin, J.H.; Cai, Y.M. Hepcidin Expression in Anemia of Chronic Disease and Concomitant Iron-Deficiency Anemia. Clin. Exp. Med. 2011, 11, 33–42. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Iron Sequestration and Anemia of Inflammation. Semin. Hematol. 2009, 46, 387–393. [Google Scholar] [CrossRef]
- Kroot, J.J.C.; Hendriks, J.C.M.; Laarakkers, C.M.M.; Klaver, S.M.; Kemna, E.H.J.M.; Tjalsma, H.; Swinkels, D.W. (Pre)Analytical Imprecision, between-Subject Variability, and Daily Variations in Serum and Urine Hepcidin: Implications for Clinical Studies. Anal. Biochem. 2009, 389, 124–129. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, X.; Jian, N.; Chen, J.; Hu, P.; Jiang, J. A Rapid and Sensitive LC–MS–MS Method for Determination of Hepcidin-25 in Human Serum, and Measurement of Its Diurnal Rhythm for Healthy Subjects. Chromatographia 2015, 78, 73–80. [Google Scholar] [CrossRef]
- Young, M.F.; Glahn, R.P.; Ariza-Nieto, M.; Inglis, J.; Olbina, G.; Westerman, M.; O’Brien, K.O. Serum Hepcidin Is Significantly Associated with Iron Absorption from Food and Supplemental Sources in Healthy Young Women. Am. J. Clin. Nutr. 2009, 89, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.T.; Witcher, D.R.; Luan, P.; Wroblewski, V.J. Quantitation of Hepcidin from Human and Mouse Serum Using Liquid Chromatography Tandem Mass Spectrometry. Blood 2007, 110, 1048–1054. [Google Scholar] [CrossRef]
- Girelli, D.; Nemeth, E.; Swinkels, D.W. Hepcidin in the Diagnosis of Iron Disorders. Blood 2016, 127, 2809–2813. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.; Ganz, T.; Goodnough, L.T. Anemia of Inflammation. Blood 2019, 133, 40–50. [Google Scholar] [CrossRef]
- Jelkmann, W. Regulation of Erythropoietin Production. J. Physiol. 2011, 589, 1251–1258. [Google Scholar] [CrossRef]
- Seekircher, L.; Siller, A.; Amato, M.; Tschiderer, L.; Balog, A.; Astl, M.; Schennach, H.; Willeit, P. HemoCue Hb-801 Provides More Accurate Hemoglobin Assessment in Blood Donors Than OrSense NBM-200. Transfus. Med. Rev. 2024, 38, 150826. [Google Scholar] [CrossRef]
- Asare, J.W.; Appiahene, P.; Donkoh, E.T.; Dimauro, G. Iron Deficiency Anemia Detection Using Machine Learning Models: A Comparative Study of Fingernails, Palm and Conjunctiva of the Eye Images. Eng. Rep. 2023, 5, e12667. [Google Scholar] [CrossRef]
- WHO. WHO Guideline on Use of Ferritin Concentrations to Assess Iron Status in Individuals and Populations; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Iolascon, A.; Andolfo, I.; Russo, R.; Sanchez, M.; Busti, F.; Swinkels, D.; Aguilar Martinez, P.; Bou-Fakhredin, R.; Muckenthaler, M.U.; Unal, S.; et al. Recommendations for Diagnosis, Treatment, and Prevention of Iron Deficiency and Iron Deficiency Anemia. Hemasphere 2024, 8, e108. [Google Scholar] [CrossRef]
- Bouri, S.; Martin, J. Investigation of Iron Deficiency Anaemia. Clin. Med. 2018, 18, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Dahlerup, J.F.; Eivindson, M.; Jacobsen, B.A.; Jensen, N.M.; Jørgensen, S.P.; Laursen, S.B.; Rasmussen, M.; Nathan, T. Diagnosis and Treatment of Unexplained Anemia with Iron Deficiency without Overt Bleeding. Dan. Med. J. 2015, 62, C5072. [Google Scholar] [PubMed]
- Snook, J.; Bhala, N.; Beales, I.L.P.; Cannings, D.; Kightley, C.; Logan, R.P.H.; Pritchard, D.M.; Sidhu, R.; Surgenor, S.; Thomas, W.; et al. British Society of Gastroenterology Guidelines for the Management of Iron Deficiency Anaemia in Adults. Gut 2021, 70, 2030–2051. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gaxiola, A.C.; De-Regil, L.M. Intermittent Iron Supplementation for Reducing Anaemia and Its Associated Impairments in Adolescent and Adult Menstruating Women. Cochrane Database Syst. Rev. 2019, 1, CD009218. [Google Scholar] [CrossRef] [PubMed]
- Pantopoulos, K. Oral Iron Supplementation: New Formulations, Old Questions. Haematologica 2024, 109, 2790. [Google Scholar] [CrossRef]
- Melina, V.; Craig, W.; Levin, S. Position of the Academy of Nutrition and Dietetics: Vegetarian Diets. J. Acad. Nutr. Diet. 2016, 116, 1970–1980. [Google Scholar] [CrossRef]
- Thane, C.W.; Walmsley, C.M.; Bates, C.J.; Prentice, A.; Cole, T.J. Risk Factors for Poor Iron Status in British Toddlers: Further Analysis of Data from the National Diet and Nutrition Survey of Children Aged 1.5–4.5 Years. Public. Health Nutr. 2000, 3, 433–440. [Google Scholar] [CrossRef]
- Singh, A.; Bains, K.; Kaur, H. Effect of Inclusion of Key Foods on in Vitro Iron Bioaccessibility in Composite Meals. J. Food Sci. Technol. 2016, 53, 2033–2039. [Google Scholar] [CrossRef]
- Saunders, A.V.; Craig, W.J.; Baines, S.K.; Posen, J.S. Iron and Vegetarian Diets. Med. J. Aust. 2013, 199, S11. [Google Scholar] [CrossRef]
- Food Sources of Iron|Dietary Guidelines for Americans. Available online: https://www.dietaryguidelines.gov/ (accessed on 10 May 2025).
- Iron-Rich Food|List of Meats And Vegetables|Red Cross Blood. Available online: https://www.redcrossblood.org/donate-blood/blood-donation-process/before-during-after/iron-blood-donation/iron-rich-foods.html (accessed on 24 June 2025).
- Kraemer, K.; Zimmermann, M.B. Nutritional Anemia; Springer: London, UK, 2007. [Google Scholar]
- Acosta, A.; Amar, M.; Cornbluth-Szarfarc, S.C. Iron Absorption from Typical Latin American Diets. Am. J. Clin. Nutr. 1984, 39, 953–962. [Google Scholar] [CrossRef]
- Okwuonu, I.C.; Narayanan, N.N.; Egesi, C.N.; Taylor, N.J. Opportunities and Challenges for Biofortification of Cassava to Address Iron and Zinc Deficiency in Nigeria. Glob. Food Sec. 2021, 28, 100478. [Google Scholar] [CrossRef]
- Rahman, S.; Shaheen, N. Phytate-iron Molar Ratio and Bioavailability of Iron in Bangladesh. Trop. Med. Int. Health 2022, 27, 509–514. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Anaemia Action Alliance. Available online: https://www.who.int/teams/nutrition-and-food-safety/anaemia-action-alliance (accessed on 9 May 2025).
- WHO. Nutritional Anaemias: Tools for Effective Prevention; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. Anaemia: Fact Sheet. Available online: https://www.who.int/news-room/fact-sheets/detail/anaemia (accessed on 9 May 2025).
- Hooda, J.; Shah, A.; Zhang, L. Heme, an Essential Nutrient from Dietary Proteins, Critically Impacts Diverse Physiological and Pathological Processes. Nutrients 2014, 6, 1080–1102. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Feng, Y. Intakes of Heme Iron and Zinc and Colorectal Cancer Incidence: A Meta-Analysis of Prospective Studies. Cancer Causes Control 2013, 24, 1175–1183. [Google Scholar] [CrossRef]
- Lunn, J.C.; Kuhnle, G.; Mai, V.; Frankenfeld, C.; Shuker, D.E.G.; Glen, R.C.; Goodman, J.M.; Pollock, J.R.A.; Bingham, S.A. The Effect of Haem in Red and Processed Meat on the Endogenous Formation of N -Nitroso Compounds in the Upper Gastrointestinal Tract. Carcinogenesis 2007, 28, 685–690. [Google Scholar] [CrossRef]
- Matias-Guiu, X.; Prat, J. Molecular Pathology of Endometrial Carcinoma. Histopathology 2013, 62, 111–123. [Google Scholar] [CrossRef]
- Nobles, C.L.; Green, S.I.; Maresso, A.W. A Product of Heme Catabolism Modulates Bacterial Function and Survival. PLoS Pathog. 2013, 9, e1003507. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, S.; Liu, G.; Yan, F.; Ma, X.; Huang, Z.; Tian, H. Body Iron Stores and Heme-Iron Intake in Relation to Risk of Type 2 Diabetes: A Systematic Review and Meta-Analysis. PLoS ONE 2012, 7, e41641. [Google Scholar] [CrossRef]
- Tzonou, A.; Lagiou, P.; Trichopoulou, A.; Tsoutsos, V.; Trichopoulos, D. Dietary Iron and Coronary Heart Disease Risk: A Study from Greece. Am. J. Epidemiol. 1998, 147, 161–166. [Google Scholar] [CrossRef]
- Vieira, D.A.D.S.; Sales, C.H.; Cesar, C.L.G.; Marchioni, D.M.; Fisberg, R.M. Influence of Haem, Non-Haem, and Total Iron Intake on Metabolic Syndrome and Its Components: A Population-Based Study. Nutrients 2018, 10, 314. [Google Scholar] [CrossRef]
- Van Duyn, M.A.S.; Pivonka, E. Overview of the Health Benefits of Fruit and Vegetable Consumption for the Dietetics Professional: Selected Literature. J. Am. Diet. Assoc. 2000, 100, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Klerk, M.; Jansen, M.C.J.F.; van ‘t Veer, P.; Kok, F.J. Fruits and Vegetables in Chronic Disease Prevention; Wageningen Agricultural University: Wageningen, The Netherlands, 1998; 70p. [Google Scholar]
- Steinmetz, K.A.; Potter, J.D. Vegetables, Fruit, and Cancer Prevention: A Review. J. Am. Diet. Assoc. 1996, 96, 1027–1039. [Google Scholar] [CrossRef]
- Jacques, P.F.; Chylack, L.T. Epidemiologic Evidence of a Role for the Antioxidant Vitamins and Carotenoids in Cataract Prevention. Am. J. Clin. Nutr. 1991, 53, 352S–355S. [Google Scholar] [CrossRef]
- Appel, L.J.; Moore, T.J.; Obarzanek, E.; Vollmer, W.M.; Svetkey, L.P.; Sacks, F.M.; Bray, G.A.; Vogt, T.M.; Cutler, J.A.; Windhauser, M.M.; et al. A Clinical Trial of the Effects of Dietary Patterns on Blood Pressure. N. Engl. J. Med. 1997, 336, 1117–1124. [Google Scholar] [CrossRef]
- Van Doren, L.; Steinheiser, M.; Boykin, K.; Taylor, K.J.; Menendez, M.; Auerbach, M. Expert Consensus Guidelines: Intravenous Iron Uses, Formulations, Administration, and Management of Reactions. Am. J. Hematol. 2024, 99, 1338–1348. [Google Scholar] [CrossRef]
- Auerbach, M.; Macdougall, I. The Available Intravenous Iron Formulations: History, Efficacy, and Toxicology. Hemodial. Int. 2017, 21, S83–S92. [Google Scholar] [CrossRef] [PubMed]
- Stojanovic, S.; Graudins, L.V.; Aung, A.K.; Grannell, L.; Hew, M.; Zubrinich, C. Safety of Intravenous Iron Following Infusion Reactions. J. Allergy Clin. Immunol. Pract. 2021, 9, 1660–1666. [Google Scholar] [CrossRef]
- Blumenstein, I.; Shanbhag, S.; Langguth, P.; Kalra, P.A.; Zoller, H.; Lim, W. Newer Formulations of Intravenous Iron: A Review of Their Chemistry and Key Safety Aspects–Hypersensitivity, Hypophosphatemia, and Cardiovascular Safety. Expert. Opin. Drug Saf. 2021, 20, 757–769. [Google Scholar] [CrossRef]
- Wu, T.W.; Tsai, F.P. Comparison of the Therapeutic Effects and Side Effects of Oral Iron Supplements in Iron Deficiency Anemia. Drug Res. 2016, 66, 257–261. [Google Scholar] [CrossRef]
- Stoffel, N.U.; von Siebenthal, H.K.; Moretti, D.; Zimmermann, M.B. Oral Iron Supplementation in Iron-Deficient Women: How Much and How Often? Mol. Asp. Med. 2020, 75, 100865. [Google Scholar] [CrossRef]
- Kolars, B.; Minakovic, I.; Grabovac, B.; Zivanovic, D.; Jovin, V.M. Treatment Adherence and the Contemporary Approach to Treating Type 2 Diabetes Mellitus. Biomed. Pap. 2024, 168, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Tolkien, Z.; Stecher, L.; Mander, A.P.; Pereira, D.I.A.; Powell, J.J. Ferrous Sulfate Supplementation Causes Significant Gastrointestinal Side-Effects in Adults: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0117383. [Google Scholar] [CrossRef] [PubMed]
- Moretti, D.; Goede, J.S.; Zeder, C.; Jiskra, M.; Chatzinakou, V.; Tjalsma, H.; Melse-Boonstra, A.; Brittenham, G.; Swinkels, D.W.; Zimmermann, M.B. Oral Iron Supplements Increase Hepcidin and Decrease Iron Absorption from Daily or Twice-Daily Doses in Iron-Depleted Young Women. Blood 2015, 126, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, K. Iron Deficiency Anemia: Guidelines from the American Gastroenterological Association. Am. Fam. Physician 2021, 104, 211–212. [Google Scholar]
- Clark, M.A.; Goheen, M.M.; Fulford, A.; Prentice, A.M.; Elnagheeb, M.A.; Patel, J.; Fisher, N.; Taylor, S.M.; Kasthuri, R.S.; Cerami, C. Host Iron Status and Iron Supplementation Mediate Susceptibility to Erythrocytic Stage Plasmodium falciparum. Nat. Commun. 2014, 5, 4446. [Google Scholar] [CrossRef]
- Arcangelo, V.; Peterson, A. (Eds.) Pharmacotherapeutics for Advanced Practice: A Practical Approach; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2006. [Google Scholar]
- Idjradinata, P.; Watkins, W.E.; Pollitt, E. Adverse Effect of Iron Supplementation on Weight Gain of Iron-Replete Young Children. Lancet 1994, 343, 1252–1254. [Google Scholar] [CrossRef]
- Majumdar, I.; Paul, P.; Talib, V.H.; Ranga, S. The Effect of Iron Therapy on the Growth of Iron-Replete and Iron-Deplete Children. J. Trop. Pediatr. 2003, 49, 84–88. [Google Scholar] [CrossRef]
- Gahagan, S.; Delker, E.; Blanco, E.; Burrows, R.; Lozoff, B. Randomized Controlled Trial of Iron-Fortified versus Low-Iron Infant Formula: Developmental Outcomes at 16 Years. J. Pediatr. 2019, 212, 124–130.e1. [Google Scholar] [CrossRef]
- Lozoff, B.; Castillo, M.; Clark, K.M.; Smith, J.B. Iron-Fortified vs Low-Iron Infant Formula: Developmental Outcome at 10 Years. Arch. Pediatr. Adolesc. Med. 2012, 166, 208–215. [Google Scholar] [CrossRef]
- Hurrell, R.F. Safety and Efficacy of Iron Supplements in Malaria-Endemic Areas. Ann. Nutr. Metab. 2011, 59, 64–66. [Google Scholar] [CrossRef]
- Hotz, C.; McClafferty, B. From Harvest to Health: Challenges for Developing Biofortified Staple Foods and Determining Their Impact on Micronutrient Status. Food Nutr. Bull. 2007, 28, S271–S279. [Google Scholar] [CrossRef] [PubMed]
- Hurrell, R.F. Fortification: Overcoming Technical and Practical Barriers. J. Nutr. 2002, 132, 806S–812S. [Google Scholar] [CrossRef] [PubMed]
- Klassen-Wigger, P.; Barclay, D.V. Market-Driven Fortification. In Food Fortification in a Globalized World; Academic Press: Cambridge, MA, USA, 2018; pp. 63–67. [Google Scholar] [CrossRef]
- Hackl, L.; Cercamondi, C.I.; Zeder, C.; Wild, D.; Adelmann, H.; Zimmermann, M.B.; Moretti, D. Cofortification of Ferric Pyrophosphate and Citric Acid/Trisodium Citrate into Extruded Rice Grains Doubles Iron Bioavailability through in Situ Generation of Soluble Ferric Pyrophosphate Citrate Complexes. Am. J. Clin. Nutr. 2016, 103, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Diosady, L.L.; Mannar, M.G.V.; Krishnaswamy, K. Improving the Lives of Millions through New Double Fortification of Salt Technology. Matern. Child. Nutr. 2019, 15, e12773. [Google Scholar] [CrossRef]
- Van Thuy, P.; Berger, J.; Davidsson, L.; Khan, N.C.; Lam, N.T.; Cook, J.D.; Hurrell, R.F.; Khoi, H.H. Regular Consumption of NaFeEDTA-Fortified Fish Sauce Improves Iron Status and Reduces the Prevalence of Anemia in Anemic Vietnamese Women. Am. J. Clin. Nutr. 2003, 78, 284–290. [Google Scholar] [CrossRef]
- Huo, J.S.; Yin, J.Y.; Sun, J.; Huang, J.; Lu, Z.X.; Regina, M.P.; Chen, J.S.; Chen, C.M. Effect of NaFeEDTA-Fortified Soy Sauce on Anemia Prevalence in China: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Biomed. Environ. Sci. 2015, 28, 788–798. [Google Scholar] [CrossRef]
- Forsman, C.; Milani, P.; Schondebare, J.A.; Matthias, D.; Guyondet, C. Rice Fortification: A Comparative Analysis in Mandated Settings. Ann. N. Y. Acad. Sci. 2014, 1324, 67–81. [Google Scholar] [CrossRef]
- Hurrell, R.F. Ensuring the Efficacious Iron Fortification of Foods: A Tale of Two Barriers. Nutrients 2022, 14, 1609. [Google Scholar] [CrossRef]
- Iwuozor, K.O.; Mbamalu, P.S.; Olaniyi, B.O.; Anyanwu, V.U.; Emenike, E.C.; Adeniyi, A.G. Fortification of Sugar: A Call for Action. Sugar Tech 2022, 24, 1284–1294. [Google Scholar] [CrossRef]
- Suchdev, P.S.; Williams, A.M.; Mei, Z.; Flores-Ayala, R.; Pasricha, S.R.; Rogers, L.M.; Namaste, S.M.L. Assessment of Iron Status in Settings of Inflammation: Challenges and Potential Approaches. Am. J. Clin. Nutr. 2017, 106, 1626S–1633S. [Google Scholar] [CrossRef]
- Fishbane, S.; Block, G.A.; Loram, L.; Neylan, J.; Pergola, P.E.; Uhlig, K.; Chertow, G.M. Effects of Ferric Citrate in Patients with Nondialysis-Dependent CKD and Iron Deficiency Anemia. J. Am. Soc. Nephrol. 2017, 28, 1851–1858. [Google Scholar] [CrossRef]
- Schmidt, C.; Allen, S.; Kopyt, N.; Pergola, P. Iron Replacement Therapy with Oral Ferric Maltol: Review of the Evidence and Expert Opinion. J. Clin. Med. 2021, 10, 4448. [Google Scholar] [CrossRef]
- Gasche, C.; Ahmad, T.; Tulassay, Z.; Baumgart, D.C.; Bokemeyer, B.; Büning, C.; Howaldt, S.; Stallmach, A. Ferric Maltol Is Effective in Correcting Iron Deficiency Anemia in Patients with Inflammatory Bowel Disease: Results from a Phase-3 Clinical Trial Program. Inflamm. Bowel Dis. 2015, 21, 579–588. [Google Scholar] [CrossRef]
- Gómez-Ramírez, S.; Brilli, E.; Tarantino, G.; Muñoz, M. Sucrosomial® Iron: A New Generation Iron for Improving Oral Supplementation. Pharmaceuticals 2018, 11, 97. [Google Scholar] [CrossRef]
- Abbati, G.; Incerti, F.; Boarini, C.; Pileri, F.; Bocchi, D.; Ventura, P.; Buzzetti, E.; Pietrangelo, A. Safety and Efficacy of Sucrosomial Iron in Inflammatory Bowel Disease Patients with Iron Deficiency Anemia. Intern. Emerg. Med. 2019, 14, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Giordano, G.; Napolitano, M.; Di Battista, V.; Lucchesi, A. Oral High-Dose Sucrosomial Iron vs Intravenous Iron in Sideropenic Anemia Patients Intolerant/Refractory to Iron Sulfate: A Multicentric Randomized Study. Ann. Hematol. 2021, 100, 2173–2179. [Google Scholar] [CrossRef] [PubMed]
- Cesarano, D.; Borrelli, S.; Campilongo, G.; D’Ambra, A.; Papadia, F.; Garofalo, C.; De Marco, A.; Marzano, F.; Ruotolo, C.; Gesualdo, L.; et al. Efficacy and Safety of Oral Supplementation with Liposomal Iron in Non-Dialysis Chronic Kidney Disease Patients with Iron Deficiency. Nutrients 2024, 16, 1255. [Google Scholar] [CrossRef] [PubMed]
- Maladkar, M.; Sankar, S.; Yadav, A.; Maladkar, M.; Sankar, S.; Yadav, A. A Novel Approach for Iron Deficiency Anaemia with Liposomal Iron: Concept to Clinic. J. Biosci. Med. 2020, 8, 27–41. [Google Scholar] [CrossRef]
- de Alvarenga Antunes, C.V.; de Alvarenga Nascimento, C.R.; Campanha da Rocha Ribeiro, T.; de Alvarenga Antunes, P.; de Andrade Chebli, L.; Martins Gonçalves Fava, L.; Malaguti, C.; Maria Fonseca Chebli, J. Treatment of Iron Deficiency Anemia with Liposomal Iron in Inflammatory Bowel Disease: Efficacy and Impact on Quality of Life. Int. J. Clin. Pharm. 2020, 42, 895–902. [Google Scholar] [CrossRef]
- Biniwale, P.; Pal, B.; Sundari, T.; Mandrupkar, G.; Datar, N.; Khurana, A.S.; Qamra, A.; Motlekar, S.; Jain, R.; Biniwale, P.; et al. Liposomal Iron for Iron Deficiency Anemia in Women of Reproductive Age: Review of Current Evidence. Open J. Obs. Gynecol. 2018, 8, 993–1005. [Google Scholar] [CrossRef]
- Mohammed, N.I.; Wason, J.; Mendy, T.; Nass, S.A.; Ofordile, O.; Camara, F.; Baldeh, B.; Sanyang, C.; Jallow, A.T.; Hossain, I.; et al. A Novel Nano-Iron Supplement versus Standard Treatment for Iron Deficiency Anaemia in Children 6–35 Months (IHAT-GUT Trial): A Double-Blind, Randomised, Placebo-Controlled Non-Inferiority Phase II Trial in The Gambia. EClinicalMedicine 2023, 56, 101853. [Google Scholar] [CrossRef]
- Tsilika, M.; Mitrou, J.; Antonakos, N.; Tseti, I.K.; Damoraki, G.; Leventogiannis, K.; Giamarellos-Bourboulis, E.J. An Active New Formulation of Iron Carried by Aspartyl Casein for Iron-Deficiency Anemia: Results of the ACCESS Trial. Ann. Hematol. 2023, 102, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Meeting Highlights from the Committee for Medicinal Products for Human Use (CHMP) 24–27 March 2025|European Medicines Agency (EMA). Available online: https://www.ema.europa.eu/en/news/meeting-highlights-committee-medicinal-products-human-use-chmp-24-27-march-2025 (accessed on 22 May 2025).
- Scott, L.J. Ferric Carboxymaltose: A Review in Iron Deficiency. Drugs 2018, 78, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Keating, G.M. Ferric Carboxymaltose: A Review of Its Use in Iron Deficiency. Drugs 2015, 75, 101–127. [Google Scholar] [CrossRef]
- INJECTAFER® Approved in the U.S. for the Treatment of Iron Deficiency in Adult Patients with Heart Failure|HFSA. Available online: https://hfsa.org/injectaferr-approved-us-treatment-iron-deficiency-adult-patients-heart-failure (accessed on 22 May 2025).
- Zoller, H.; Wolf, M.; Blumenstein, I.; Primas, C.; Lindgren, S.; Thomsen, L.L.; Reinisch, W.; Iqbal, T. Hypophosphataemia Following Ferric Derisomaltose and Ferric Carboxymaltose in Patients with Iron Deficiency Anaemia Due to Inflammatory Bowel Disease (PHOSPHARE-IBD): A Randomised Clinical Trial. Gut 2023, 72, 644–653. [Google Scholar] [CrossRef]
- Cleland, J.G.F.; Kalra, P.A.; Pellicori, P.; Graham, F.J.; Foley, P.W.X.; Squire, I.B.; Cowburn, P.J.; Seed, A.; Clark, A.L.; Szwejkowski, B.; et al. Intravenous Iron for Heart Failure, Iron Deficiency Definitions, and Clinical Response: The IRONMAN Trial. Eur. Heart J. 2024, 45, 1410–1426. [Google Scholar] [CrossRef]
- Adkinson, N.F.; Strauss, W.E.; Macdougall, I.C.; Bernard, K.E.; Auerbach, M.; Kaper, R.F.; Chertow, G.M.; Krop, J.S. Comparative Safety of Intravenous Ferumoxytol versus Ferric Carboxymaltose in Iron Deficiency Anemia: A Randomized Trial. Am. J. Hematol. 2018, 93, 683–690. [Google Scholar] [CrossRef]
- Van Doren, L.; Auerbach, M. IV Iron Formulations and Use in Adults. Hematology 2023, 2023, 622–629. [Google Scholar] [CrossRef]
- Intravenous Iron Offers Some Benefits for Some Patients with Heart Failure–American College of Cardiology. Available online: https://www.acc.org/About-ACC/Press-Releases/2025/03/30/12/35/Intravenous-Iron-Offers-Some-Benefits-for-Some-Patients-with-Heart-Failure? (accessed on 22 May 2025).
- Derman, R.J.; Bellad, M.B.; Somannavar, M.S.; Bhandari, S.; Mehta, S.; Mehta, S.; Sharma, D.K.; Kumar, Y.; Charantimath, U.; Patil, A.P.; et al. Single-Dose Intravenous Iron vs Oral Iron for Treatment of Maternal Iron Deficiency Anemia: A Randomized Clinical Trial. Am. J. Obs. Gynecol. 2025, in press. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolarš, B.; Mijatović Jovin, V.; Živanović, N.; Minaković, I.; Gvozdenović, N.; Dickov Kokeza, I.; Lesjak, M. Iron Deficiency and Iron Deficiency Anemia: A Comprehensive Overview of Established and Emerging Concepts. Pharmaceuticals 2025, 18, 1104. https://doi.org/10.3390/ph18081104
Kolarš B, Mijatović Jovin V, Živanović N, Minaković I, Gvozdenović N, Dickov Kokeza I, Lesjak M. Iron Deficiency and Iron Deficiency Anemia: A Comprehensive Overview of Established and Emerging Concepts. Pharmaceuticals. 2025; 18(8):1104. https://doi.org/10.3390/ph18081104
Chicago/Turabian StyleKolarš, Bela, Vesna Mijatović Jovin, Nemanja Živanović, Ivana Minaković, Nemanja Gvozdenović, Isidora Dickov Kokeza, and Marija Lesjak. 2025. "Iron Deficiency and Iron Deficiency Anemia: A Comprehensive Overview of Established and Emerging Concepts" Pharmaceuticals 18, no. 8: 1104. https://doi.org/10.3390/ph18081104
APA StyleKolarš, B., Mijatović Jovin, V., Živanović, N., Minaković, I., Gvozdenović, N., Dickov Kokeza, I., & Lesjak, M. (2025). Iron Deficiency and Iron Deficiency Anemia: A Comprehensive Overview of Established and Emerging Concepts. Pharmaceuticals, 18(8), 1104. https://doi.org/10.3390/ph18081104