
Influence of Tariquidar, an ABC Transporter Inhibitor, on the Ca2+-Dependent Mitochondrial Permeability Transition Pore


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
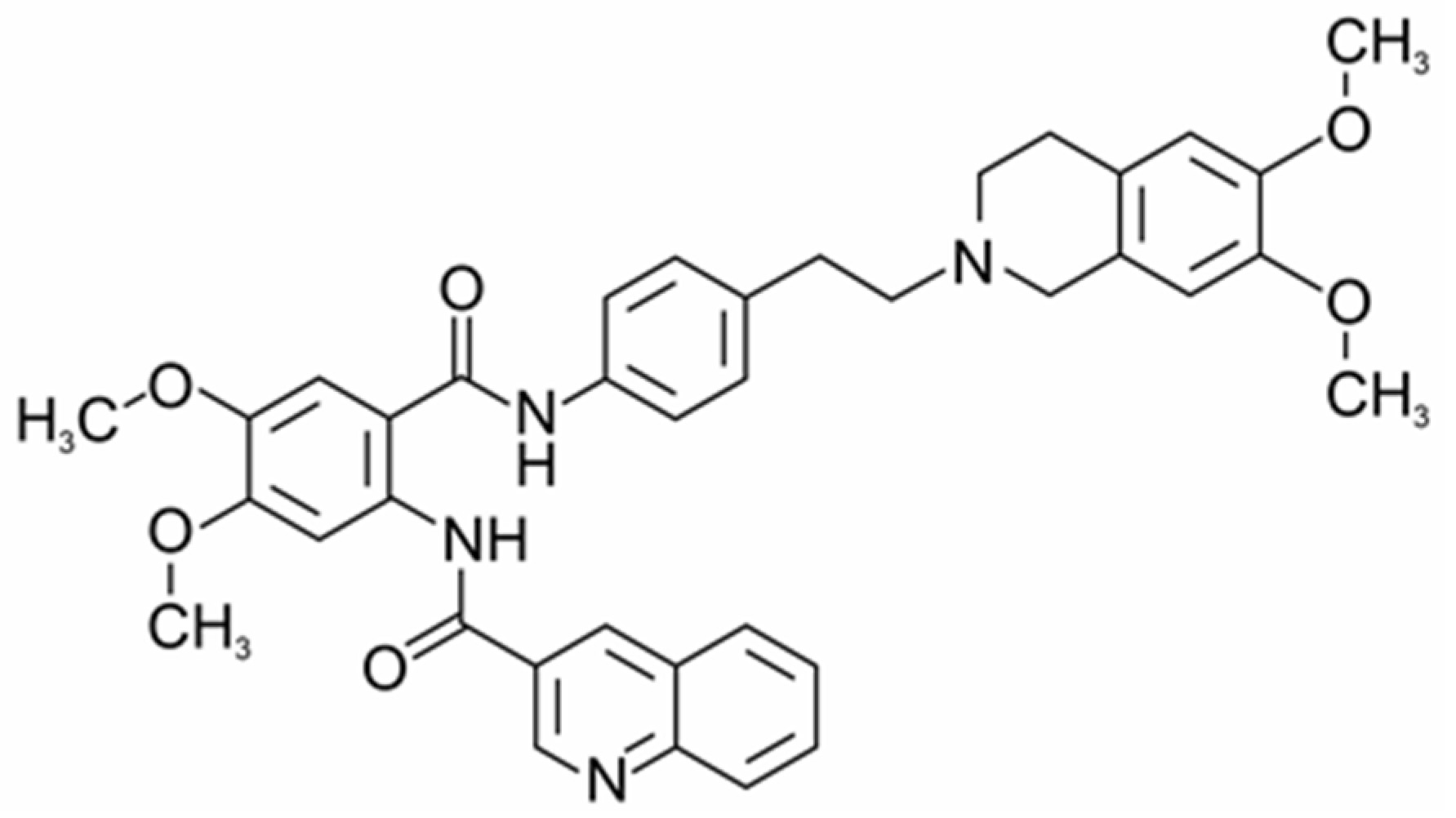
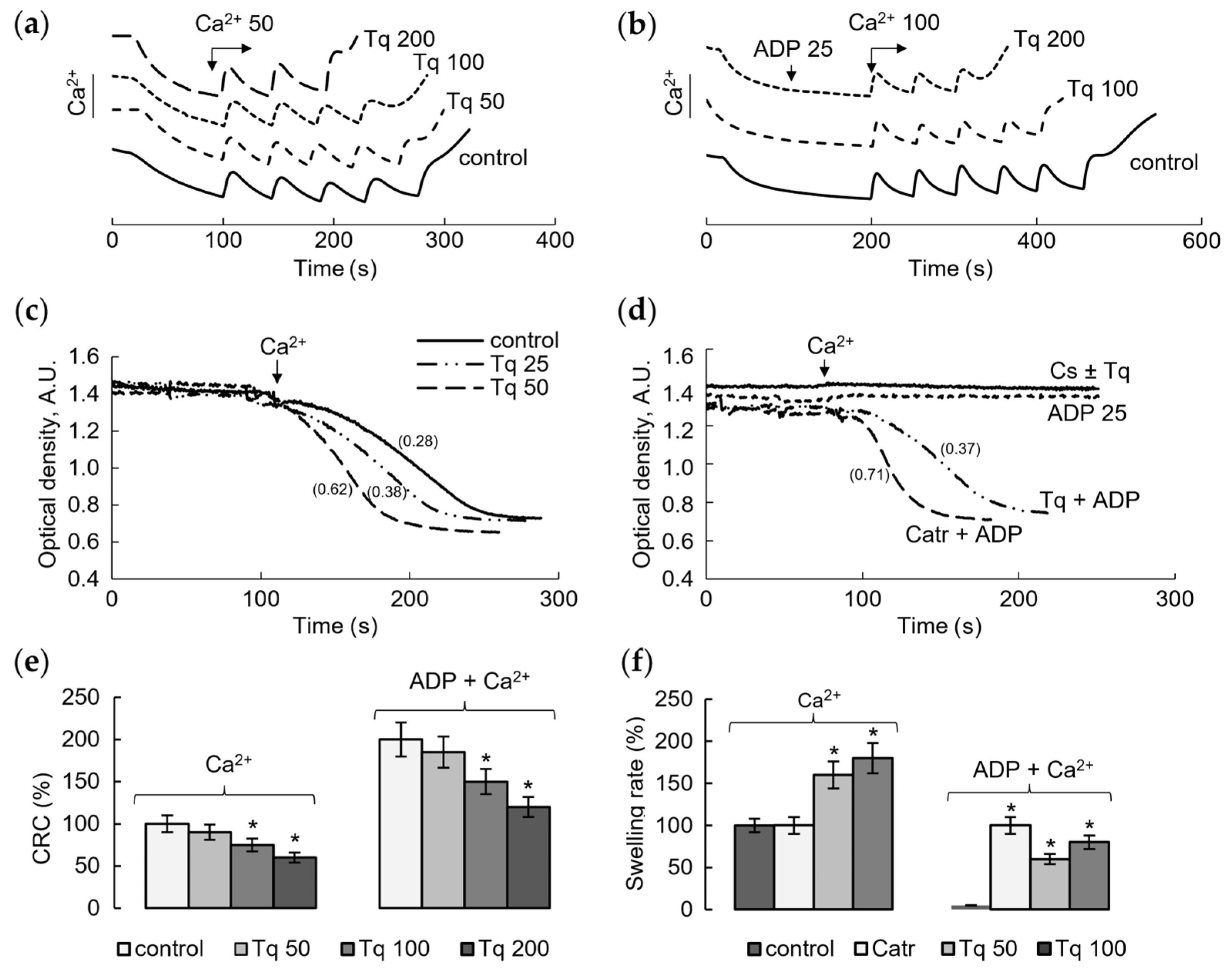
2.1. Influence of Tq on Resistance to Calcium Load and Ca2+-Induced mPTP Opening
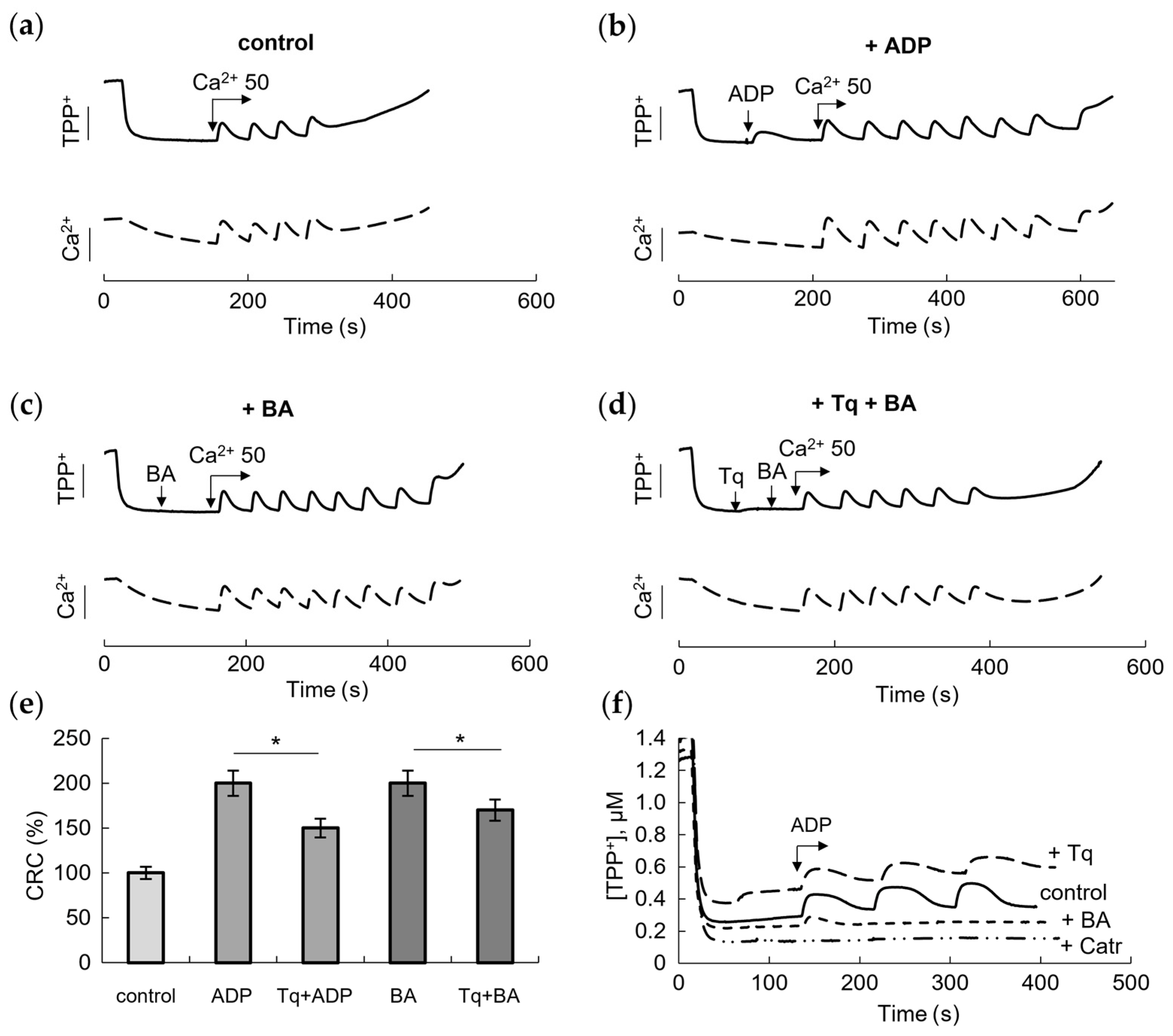
2.2. Influence of Tq on the Membrane Potential and Resistance to the Calcium Load and ANT Inhibitors
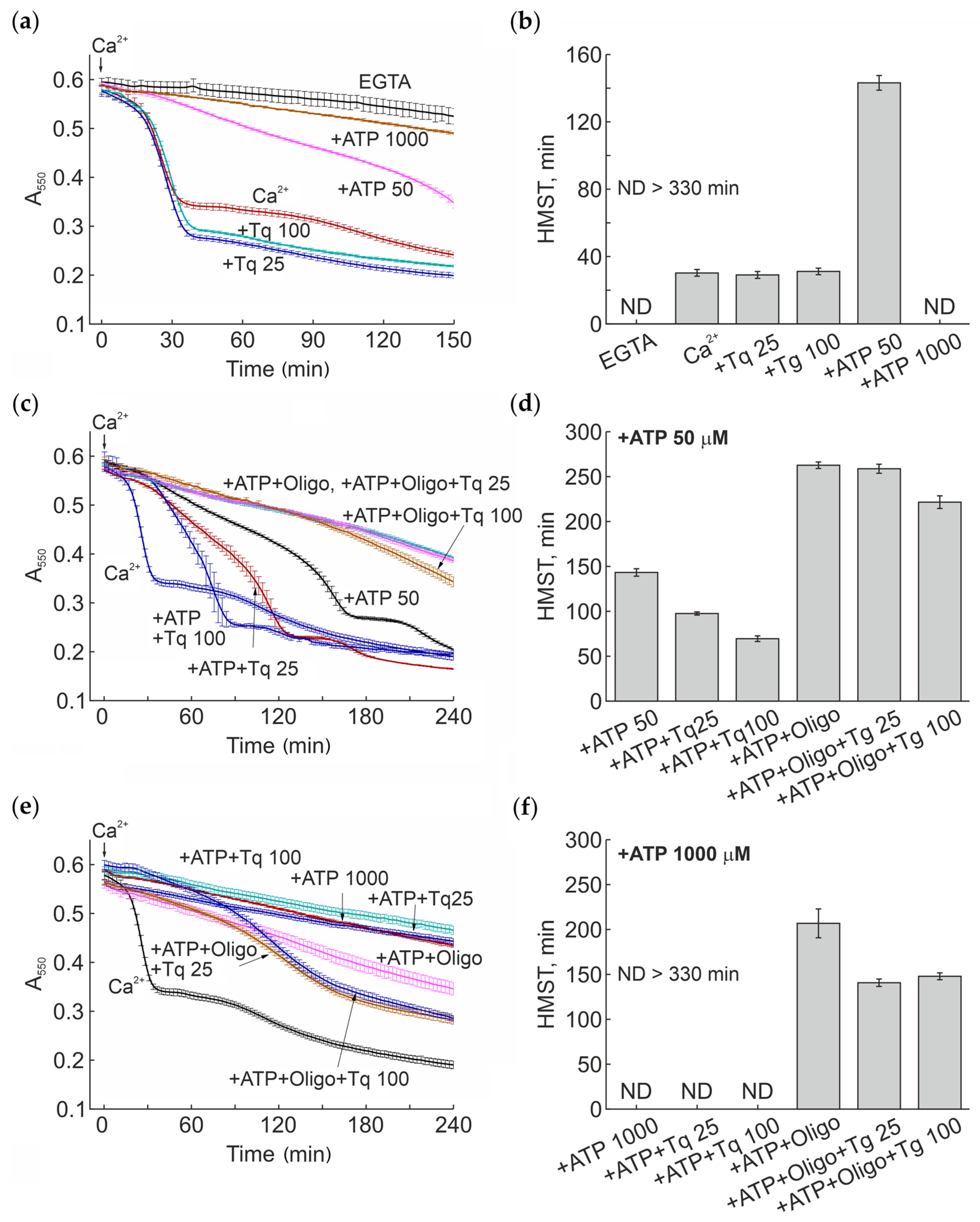
2.3. Modulation of the Ca2+-Induced mPTP Opening by Tq at Different Concentrations of Adenine Nucleotides
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Preparation of Rat Liver Mitochondria
4.3. Determination of Calcium Retention Capacity
4.4. Assay of Swelling of Mitochondria
4.5. Determination of the Mitochondrial Membrane Potential
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABC | ATP binding cassette |
| ANT | adenine nucleotide translocase |
| BA | bongkrekic acid |
| Catr | Carboxyatractylozide |
| CsA | cyclosporine A |
| HMST | half-maximum swelling time |
| mPTP | mitochondrial permeability transition pore |
| MDR | multidrug resistance |
| Tq | Tariquidar |
| TPP+ | tetraphenylphosphonium ion |
References
- Goebel, J.; Chmielewski, J.; Hrycyna, C.A. The Roles of the Human ATP-Binding Cassette Transporters P-Glycoprotein and ABCG2 in Multidrug Resistance in Cancer and at Endogenous Sites: Future Opportunities for Structure-Based Drug Design of Inhibitors. Cancer Drug Resist. 2021, 4, 784–804. [Google Scholar] [CrossRef] [PubMed]
- Seeger, M.A.; Van Veen, H.W. Molecular Basis of Multidrug Transport by ABC Transporters. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2009, 1794, 725–737. [Google Scholar] [CrossRef]
- Xiao, H.; Zheng, Y.; Ma, L.; Tian, L.; Sun, Q. Clinically-Relevant ABC Transporter for Anti-Cancer Drug Resistance. Front. Pharmacol. 2021, 12, 648407. [Google Scholar] [CrossRef]
- Mollazadeh, S.; Sahebkar, A.; Hadizadeh, F.; Behravan, J.; Arabzadeh, S. Structural and Functional Aspects of P-Glycoprotein and Its Inhibitors. Life Sci. 2018, 214, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Di, J.; Luo, D.; Vaishnav, Y.; Kamal; Nuralieva, N.; Verma, D.; Verma, P.; Verma, S. Recent developments of P-glycoprotein inhibitors and its structure-activity relationship (SAR) studies. Bioorg. Chem. 2024, 143, 106997. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, A.; Sousa, E.; Vasconcelos, M.H.; Pinto, M.M. Three Decades of P-Gp Inhibitors: Skimming Through Several Generations and Scaffolds. Curr. Med. Chem. 2012, 19, 1946–2025. [Google Scholar] [CrossRef]
- Tian, Y.; Lei, Y.; Wang, Y.; Lai, J.; Wang, J.; Xia, F. Mechanism of Multidrug Resistance to Chemotherapy Mediated by P-glycoprotein (Review). Int. J. Oncol. 2023, 63, 119. [Google Scholar] [CrossRef]
- Moesgaard, L.; Pedersen, M.L.; Uhd Nielsen, C.; Kongsted, J. Structure-Based Discovery of Novel P-Glycoprotein Inhibitors Targeting the Nucleotide Binding Domains. Sci. Rep. 2023, 13, 21217. [Google Scholar] [CrossRef]
- Teodori, E.; Braconi, L.; Manetti, D.; Romanelli, M.N.; Dei, S. The Tetrahydroisoquinoline Scaffold in ABC Transporter Inhibitors that Act as Multidrug Resistance (MDR) Reversers. Curr. Top. Med. Chem. 2022, 22, 2535–2569. [Google Scholar] [CrossRef]
- Joshi, P.; Vishwakarma, R.A.; Bharate, S.B. Natural Alkaloids as P-Gp Inhibitors for Multidrug Resistance Reversal in Cancer. Eur. J. Med. Chem. 2017, 138, 273–292. [Google Scholar] [CrossRef]
- Chufan, E.E.; Kapoor, K.; Ambudkar, S.V. Drug–Protein Hydrogen Bonds Govern the Inhibition of the ATP Hydrolysis of the Multidrug Transporter P-Glycoprotein. Biochem. Pharmacol. 2016, 101, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Inxight Drugs. Available online: https://drugs.ncats.io/substance/J58862DTVD (accessed on 8 June 2025).
- Yu, C.P.; Lin, S.W.; Tsai, J.C.; Shyong, Y.J. Long acting tariquidar loaded stearic acid-modified hydroxyapatite enhances brain penetration and antitumor effect of temozolomide. Eur. J. Pharm. Biopharm. 2024, 197, 114231. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.J.; Draper, D.; Chen, C.C.; Robey, R.W.; Figg, W.D.; Piekarz, R.L.; Chen, X.; Gardner, E.R.; Balis, F.M.; Venkatesan, A.M.; et al. A pharmacodynamic study of docetaxel in combination with the P-glycoprotein antagonist tariquidar (XR9576) in patients with lung, ovarian, and cervical cancer. Clin. Cancer Res. 2011, 17, 569–580. [Google Scholar] [CrossRef]
- Ilyas-Feldmann, M.; Langer, O.; Bauer, M.; Asselin, M.C.; Hendrikse, N.H.; Sisodiya, S.M.; Duncan, J.S.; Löscher, W.; Koepp, M. Tolerability of tariquidar—A third generation P-gp inhibitor as add-on medication to antiseizure medications in drug-resistant epilepsy. Seizure 2024, 119, 44–51. [Google Scholar] [CrossRef]
- Thompson, E.M.; Cheng, L.; Spasojevic, I. Enhanced plasma and brain concentrations and medulloblastoma cytotoxicity of asciminib and nilotinib by P-glycoprotein inhibition with tariquidar. Anticancer Drugs 2025. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.; Back, H.; Lee, J.B.; Oh, S.; Guo, T.; Girgis, S.; Park, C.; Haroutounian, S.; Kagan, L. Pharmacokinetic Modeling of the Effect of Tariquidar on Ondansetron Disposition into the Central Nervous System. Pharm. Res. 2024, 41, 1401–1411. [Google Scholar] [CrossRef]
- Genovese, I.; Carinci, M.; Modesti, L.; Aguiari, G.; Pinton, P.; Giorgi, C. Mitochondria: Insights into Crucial Features to Overcome Cancer Chemoresistance. Int. J. Mol. Sci. 2021, 22, 4770. [Google Scholar] [CrossRef]
- Bokil, A.; Sancho, P. Mitochondrial Determinants of Chemoresistance. Cancer Drug Resist. 2019, 2, 634–646. [Google Scholar] [CrossRef]
- Jangholi, E.; Sharifi, Z.N.; Hoseinian, M.; Zarrindast, M.-R.; Rahimi, H.R.; Mowla, A.; Aryan, H.; Javidi, M.A.; Parsa, Y.; Ghaffarpasand, F.; et al. Verapamil Inhibits Mitochondria-Induced Reactive Oxygen Species and Dependent Apoptosis Pathways in Cerebral Transient Global Ischemia/Reperfusion. Oxidative Med. Cell. Longev. 2020, 2020, 5872645. [Google Scholar] [CrossRef]
- Malik, C.; Ghosh, S. Quinidine Partially Blocks Mitochondrial Voltage-Dependent Anion Channel (VDAC). Eur. Biophys. J. 2020, 49, 193–205. [Google Scholar] [CrossRef]
- Fedotcheva, T.; Shimanovsky, N.; Fedotcheva, N. Involvement of Multidrug Resistance Modulators in the Regulation of the Mitochondrial Permeability Transition Pore. Membranes 2022, 12, 890. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Bhatti, G.K.; Chhabra, R.; Reddy, P.H.; Bhatti, J.S. Targeting Mitochondria as a Potential Therapeutic Strategy against Chemoresistance in Cancer. Biomed. Pharmacother. 2023, 160, 114398. [Google Scholar] [CrossRef]
- Borbolis, F.; Ploumi, C.; Palikaras, K. Calcium-mediated regulation of mitophagy: Implications in neurodegenerative diseases. NPJ Metab. Health Dis. 2025, 3, 4. [Google Scholar] [CrossRef]
- Endlicher, R.; Drahota, Z.; Štefková, K.; Červinková, Z.; Kučera, O. The Mitochondrial Permeability Transition Pore-Current Knowledge of Its Structure, Function, and Regulation, and Optimized Methods for Evaluating Its Functional State. Cells 2023, 12, 1273. [Google Scholar] [CrossRef]
- Halestrap, A.; Brenner, C. The Adenine Nucleotide Translocase: A Central Component of the Mitochondrial Permeability Transition Pore and Key Player in Cell Death. Curr. Med. Chem. 2003, 10, 1507–1525. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, L.; Liu, J.; Ma, L.; Zhang, W. Adenine Nucleotide Translocase: Current Knowledge in Post-translational Modifications, Regulations and Pathological Implications for Human Diseases. FASEB J. 2023, 37, e22953. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Deng, X.; Li, Y.; Hu, J.; Xie, L.; Shi, F.; Tang, M.; Bode, A.M.; Zhang, X.; Liao, W.; et al. Conformational Change of Adenine Nucleotide Translocase-1 Mediates Cisplatin Resistance Induced by EBV-LMP1. EMBO Mol. Med. 2021, 13, e14072. [Google Scholar] [CrossRef]
- Hu, K.; Lai, Y.; Zhou, J.; Hu, C.; Guo, S.; Zhang, H.; Wang, G.; Zhang, Q.; Gao, X.; Wang, Z.; et al. Aberrant Activation of Adenine Nucleotide Translocase 3 Promotes Progression and Chemoresistance in Multiple Myeloma Dependent on PINK1 Transport. Int. J. Biol. Sci. 2025, 21, 233–250. [Google Scholar] [CrossRef]
- Zhao, L.; Tang, M.; Bode, A.M.; Liao, W.; Cao, Y. ANTs and Cancer: Emerging Pathogenesis, Mechanisms, and Perspectives. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2021, 1875, 188485. [Google Scholar] [CrossRef]
- Lytovchenko, O.; Kunji, E.R.S. Expression and Putative Role of Mitochondrial Transport Proteins in Cancer. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 641–654. [Google Scholar] [CrossRef]
- Jang, J.-Y.; Choi, Y.; Jeon, Y.-K.; Aung, K.C.Y.; Kim, C.-W. Over-Expression of Adenine Nucleotide Translocase 1 (ANT1) Induces Apoptosis and Tumor Regression in Vivo. BMC Cancer 2008, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Gallerne, C.; Touat, Z.; Chen, Z.X.; Martel, C.; Mayola, E.; Sharaf El Dein, O.; Buron, N.; Le Bras, M.; Jacotot, E.; Borgne-Sanchez, A. The Fourth Isoform of the Adenine Nucleotide Translocator Inhibits Mitochondrial Apoptosis in Cancer Cells. Int. J. Biochem. Cell Biol. 2010, 42, 623–629. [Google Scholar] [CrossRef]
- Teodori, E.; Dei, S.; Bartolucci, G.; Perrone, M.G.; Manetti, D.; Romanelli, M.N.; Contino, M.; Colabufo, N.A. Structure–Activity Relationship Studies on 6,7-Dimethoxy-2-phenethyl-1,2,3,4-tetrahydroisoquinoline Derivatives as Multidrug Resistance Reversers. ChemMedChem 2017, 12, 1369–1379. [Google Scholar] [CrossRef]
- Hwang, S.-H.; Yang, Y.; Jung, J.-H.; Kim, Y. Heterogeneous Response of Cancer-Associated Fibroblasts to the Glucose Deprivation through Mitochondrial Calcium Uniporter. Exp. Cell Res. 2021, 406, 112778. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Luo, X.; Zhang, B.; Luo, D.; Huang, L.; Long, Q. Unveiling OSCP as the Potential Therapeutic Target for Mitochondrial Dysfunction-Related Diseases. Life Sci. 2024, 336, 122293. [Google Scholar] [CrossRef]
- Pan, T.; Yang, B.; Yao, S.; Wang, R.; Zhu, Y. Exploring the Multifaceted Role of Adenosine Nucleotide Translocase 2 in Cellular and Disease Processes: A Comprehensive Review. Life Sci. 2024, 351, 122802. [Google Scholar] [CrossRef]
- Gerle, C.; Jiko, C.; Nakano, A.; Yokoyama, K.; Gopalasingam, C.C.; Shigematsu, H.; Abe, K. Human F-ATP Synthase as a Drug Target. Pharmacol. Res. 2024, 209, 107423. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Sharma, K.; Jena, L.; Kaur, P.; Singh, S.; Munshi, A. Mitochondrial Bioenergetics of Breast Cancer. Mitochondrion 2024, 79, 101951. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, C.; Luo, L.; Tang, Y.; Yu, Y.; Li, Y.; Xing, J.; Pan, X. Membrane-Assisted Tariquidar Access and Binding Mechanisms of Human ATP-Binding Cassette Transporter P-Glycoprotein. Front. Mol. Biosci. 2024, 11, 1364494. [Google Scholar] [CrossRef]
- Ferté, J. Analysis of the Tangled Relationships between P-glycoprotein-mediated Multidrug Resistance and the Lipid Phase of the Cell Membrane. Eur. J. Biochem. 2000, 267, 277–294. [Google Scholar] [CrossRef]
- Fedotcheva, T.A.; Beloborodova, N.V.; Fedotcheva, N.I. Common Mitochondrial Targets of Curcumin and Cinnamic Acid, the Membrane-Active Natural Phenolic Compounds. Pharmaceutics 2024, 16, 1272. [Google Scholar] [CrossRef] [PubMed]
- Beloborodova, N.V.; Fedotcheva, N.I. Influence of the Microbial Metabolite Acetyl Phosphate on Mitochondrial Functions Under Conditions of Exogenous Acetylation and Alkalization. Metabolites 2024, 14, 703. [Google Scholar] [CrossRef] [PubMed]
- Kharechkina, E.S.; Nikiforova, A.B.; Kruglov, A.G. Regulation of Mitochondrial Permeability Transition Pore Opening by Monovalent Cations in Liver Mitochondria. Int. J. Mol. Sci. 2023, 24, 9237. [Google Scholar] [CrossRef]
- Kharechkina, E.S.; Nikiforova, A.B.; Teplova, V.V.; Odinokova, I.V.; Krestinina, O.V.; Baburina, Y.L.; Kruglova, S.A.; Kruglov, A.G. Regulation of Permeability Transition Pore Opening in Mitochondria by External NAD(H). Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 771–783. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fedotcheva, T.A.; Kruglov, A.G.; Fedotcheva, N.I. Influence of Tariquidar, an ABC Transporter Inhibitor, on the Ca2+-Dependent Mitochondrial Permeability Transition Pore. Pharmaceuticals 2025, 18, 924. https://doi.org/10.3390/ph18060924
Fedotcheva TA, Kruglov AG, Fedotcheva NI. Influence of Tariquidar, an ABC Transporter Inhibitor, on the Ca2+-Dependent Mitochondrial Permeability Transition Pore. Pharmaceuticals. 2025; 18(6):924. https://doi.org/10.3390/ph18060924
Chicago/Turabian StyleFedotcheva, Tatiana A., Alexey G. Kruglov, and Nadezhda I. Fedotcheva. 2025. "Influence of Tariquidar, an ABC Transporter Inhibitor, on the Ca2+-Dependent Mitochondrial Permeability Transition Pore" Pharmaceuticals 18, no. 6: 924. https://doi.org/10.3390/ph18060924
APA StyleFedotcheva, T. A., Kruglov, A. G., & Fedotcheva, N. I. (2025). Influence of Tariquidar, an ABC Transporter Inhibitor, on the Ca2+-Dependent Mitochondrial Permeability Transition Pore. Pharmaceuticals, 18(6), 924. https://doi.org/10.3390/ph18060924