
In Vitro and In Silico Assessments of Curcuminoids and Turmerones from Curcuma longa as Novel Inhibitors of Leishmania infantum Arginase

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results and Discussion
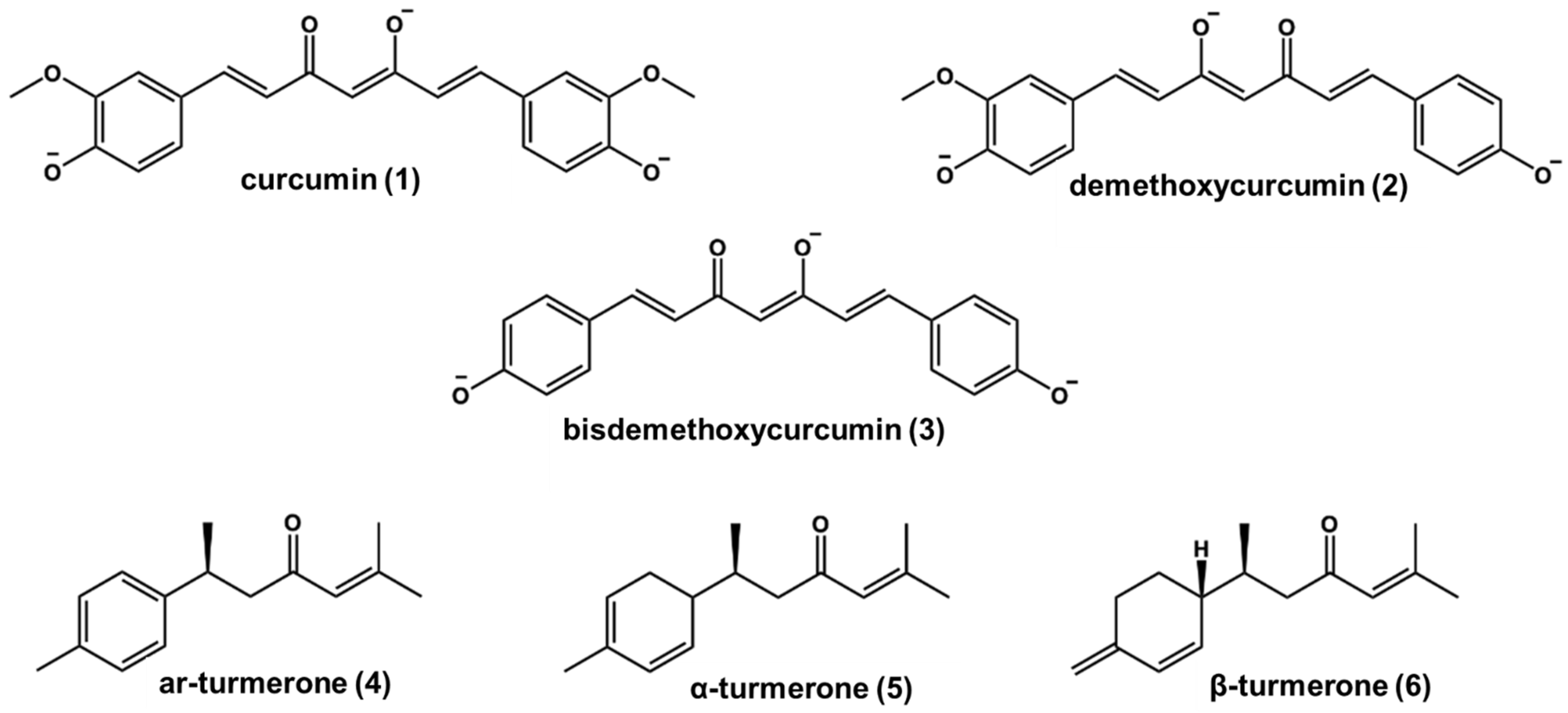
2.1. Phytochemical Profile of C. longa
2.2. LiARG Inhibition In Vitro
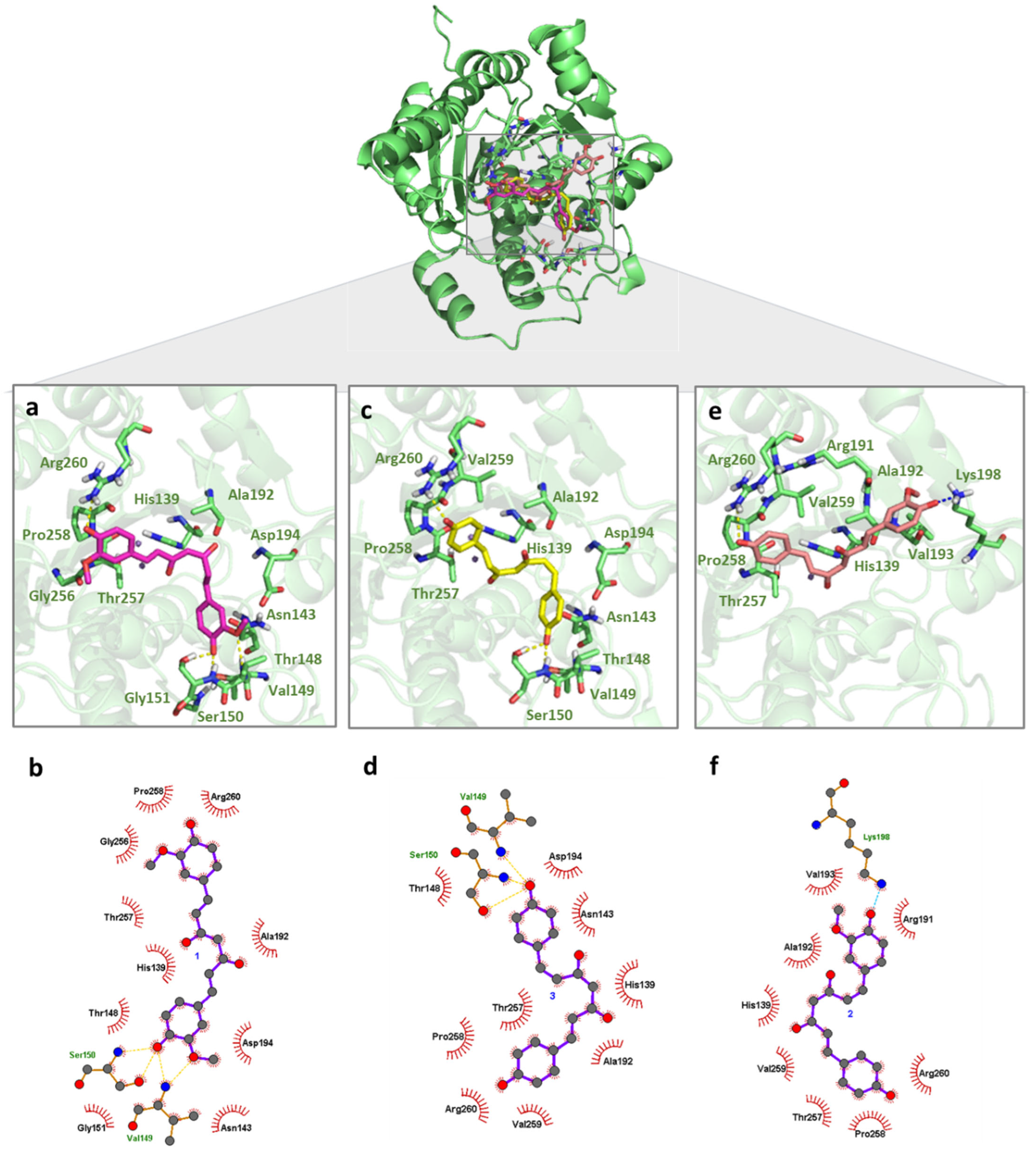
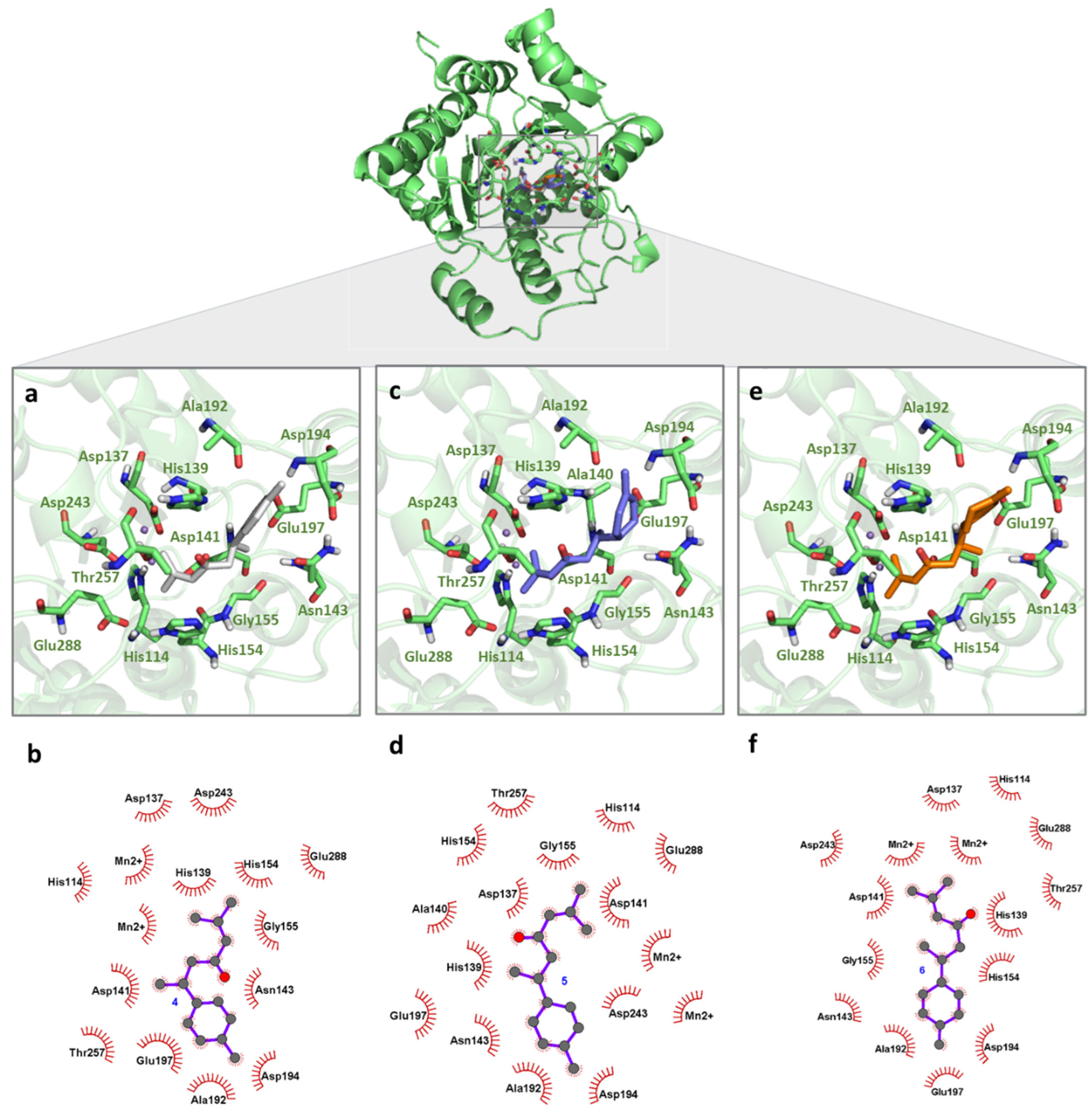
2.3. LiARG In Silico Binding Mode Analysis
2.4. In Silico ADMET Profile
2.5. Cytotoxic and Hemolytic Effects of C. longa Extracts and Curcumin
2.6. Antileishmanial Activity
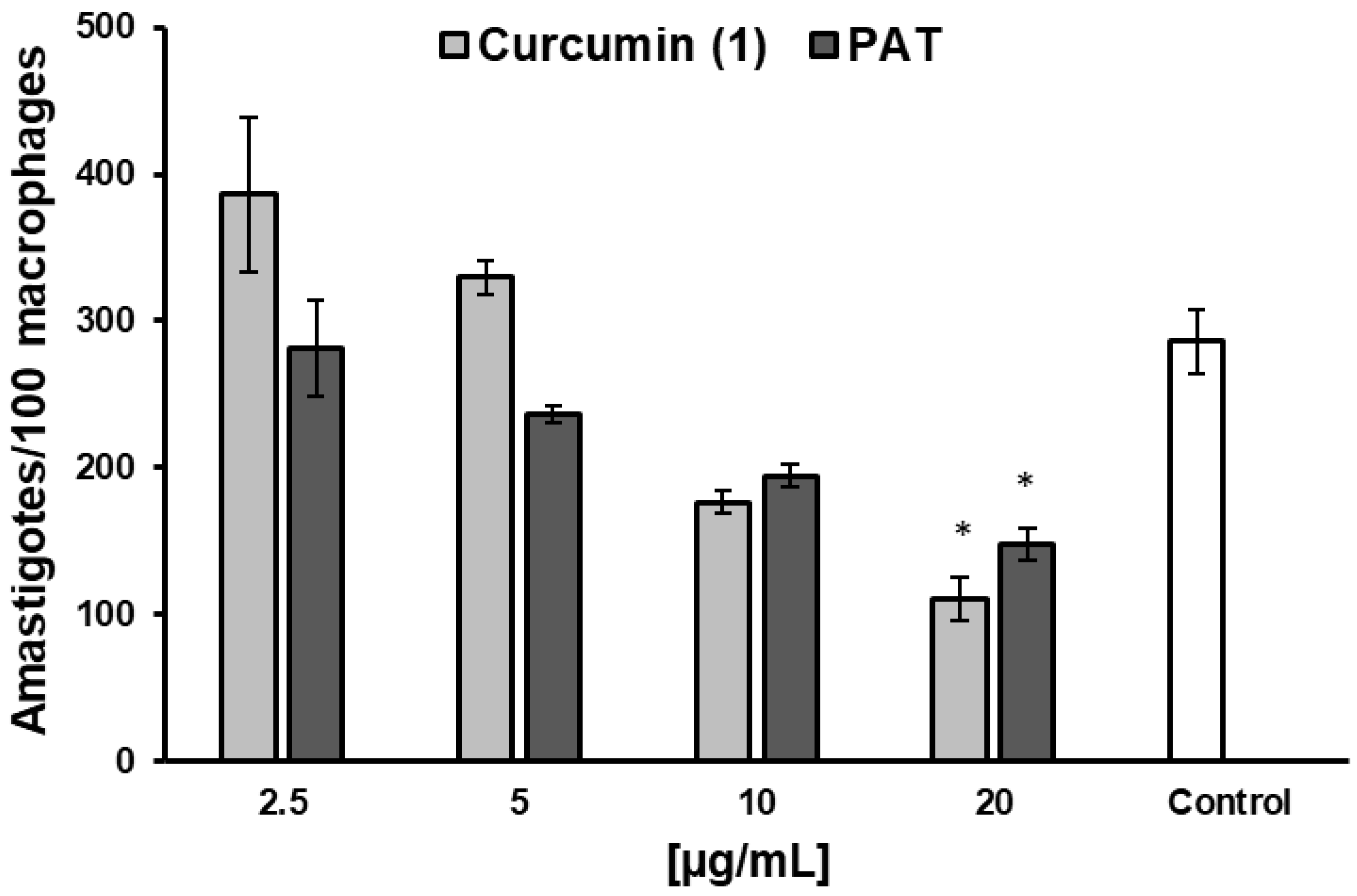
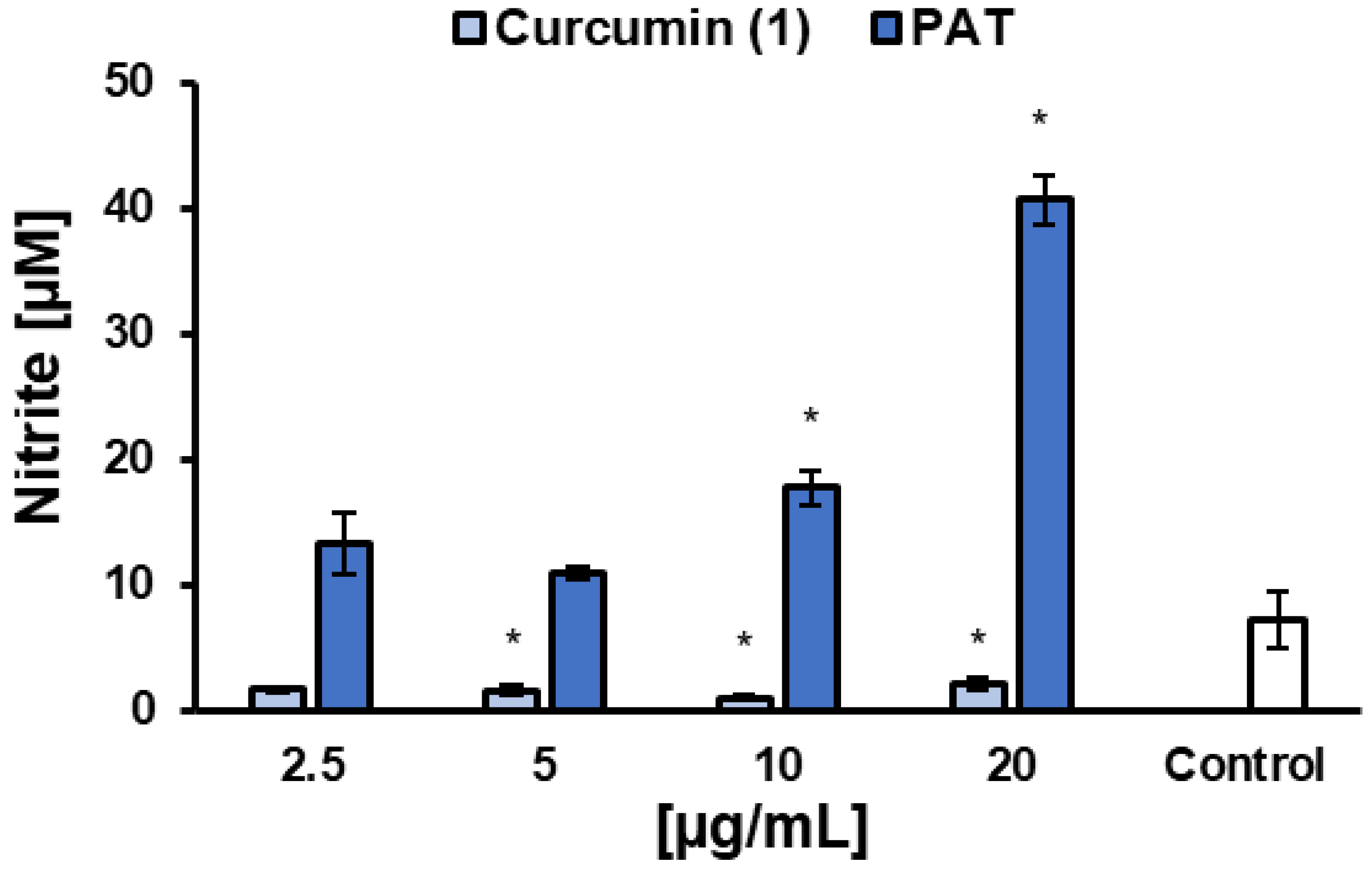
2.7. Effect of Curcumin on NO Production of L. infantum-Infected Macrophages
3. Materials and Methods
3.1. Chemicals, Reagents, and Culture Media
3.2. Plant Material and Extraction Procedures
3.3. Volatile Organic Compounds
3.3.1. Sample Preparation and Extraction SPME
3.3.2. Volatile Compounds Analysis by GC–MS
3.4. Identification of the Compounds by LC-QTOF-ESI-HRMS
3.5. LiARG Inhibition Assay
3.6. Molecular Docking for Binding Mode Analysis
3.6.1. Preparation of Structure
3.6.2. Grid Parameters
3.6.3. Docking Parameters and Analysis
3.7. Computational ADMET Analysis
3.8. Cytotoxicity Assay
3.9. Hemolysis Assay
3.10. In Vitro Anti-Leishmania spp. Activity
3.11. In Vitro Infection and Inhibition of Intracellular Amastigote Activity
3.12. Determination of NO
3.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO, World Health Organization. Leishmaniasis. 2025. Available online: https://www.who.int/leishmaniasis/en/> (accessed on 26 February 2025).
- Roatt, B.M.; Cardoso, J.M.O.; De Brito, R.C.F.; Coura-Vital, W.; Aguiar-Soares, R.D.O.; Reis, A.B. Recent advances and new strategies on leishmaniasis treatment. Appl. Microbiol. Biotechnol. 2020, 104, 8965–8977. [Google Scholar] [CrossRef] [PubMed]
- Ponte-Sucre, A.; Gamarro, F.; Dujardin, J.C.; Barrett, M.P.; López-Vélez, R.; García-Hernández, R.; Pountain, A.W.; Mwenechanya, R.; Papadopoulou, B. Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Negl. Trop. Dis. 2017, 11, e0006052. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Neto, V.V.; Cunha-Junior, E.F.; Dos Santos, V.F.; Pereira, T.M.; Silva, R.L.; Leon, L.L.; Torres-Santos, E.C. Leishmaniasis treatment: Update of possibilities for drug repurposing. Front. Biosci. 2018, 23, 967–996. [Google Scholar] [CrossRef]
- Ilari, A.; Fiorillo, A.; Genovese, I.; Colotti, G. Polyamine-trypanothione pathway: An update. Future Med. Chem. 2017, 9, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Iniesta, V.; Gomez-Nieto, L.C.; Corraliza, I. The inhibition of arginase by N(ômega)-hydroxy-l-arginine controls the growth of Leishmania inside macrophages. J. Exp. Med. 2001, 193, 777–784. [Google Scholar] [CrossRef]
- Da Silva, E.R.; Maquiaveli Cdo, C.; Magalhaes, P.P. The Leishmanicidal Flavonols Quercetin and Quercitrin Target Leishmania (Leishmania) amazonensis Arginase. Exp. Parasitol. 2012, 130, 183–188. [Google Scholar] [CrossRef]
- Cruz, E.M.; da Silva, E.R.; Maquiaveli, C.C.; Alves, E.S.; Lucon, J.F., Jr.; dos Reis, M.B.; de Toledo, C.E.; Cruz, F.G.; Vannier-Santos, M.A. Leishmanicidal activity of Cecropia pachystachya flavonoids: Arginase inhibition and altered mitochondrial DNA arrangement. Phytochemistry 2013, 89, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, M.B.; Manjolin, L.C.; Maquiaveli Cdo, C.; Santos-Filho, O.A.; Da Silva, E.R. Inhibition of Leishmania (Leishmania) amazonensis and Rat Arginases by Green Tea Egcg, (+)-Catechin and (−)-Epicatechin: A Comparative Structural Analysis of Enzyme-Inhibitor Interactions. PLoS ONE 2013, 8, e78387. [Google Scholar] [CrossRef]
- Garcia, A.R.; Oliveira, D.M.P.; Amaral, A.C.F.; Jesus, J.B.; Sodero, A.C.R.; Souza, A.M.T.; Supuran, C.T.; Vermelho, A.B.; Rodrigues, I.A.; Pinheiro, A.S. Leishmania infantum arginase: Biochemical characterization and inhibition by naturally occurring phenolic substances. J. Enzyme Inhib. Med. Chem. 2019, 34, 1100–1109. [Google Scholar] [CrossRef]
- Da Silva, E.R.; Brogi, S.; Grillo, A.; Campiani, G.; Gemma, S.; Vieira, P.C.; Maquiaveli, C.D.C. Cinnamic acids derived compounds with antileishmanial activity target Leishmania amazonensis arginase. Chem. Biol. Drug Des. 2019, 93, 139–146. [Google Scholar] [CrossRef]
- Garcia, A.R.; Oliveira, D.M.P.; Jesus, J.B.; Souza, A.M.T.; Sodero, A.C.R.; Vermelho, A.B.; Leal, I.C.R.; Souza, R.O.M.A.; Miranda, L.S.M.; Pinheiro, A.S.; et al. Identification of chalcone derivatives as inhibitors of Leishmania infantum arginase and promising antileishmanial agents. Front. Chem. 2021, 8, 624678. [Google Scholar] [CrossRef] [PubMed]
- De Lima, C.E.; Castelo-Branco, F.S.; Maquiaveli, C.C.; Farias, A.B.; Rennó, M.N.; Boechat, N.; Silva, E.R. Phenylhydrazides as inhibitors of Leishmania amazonensis arginase and antileishmanial activity. Bioorg. Med. Chem. 2019, 27, 3853–3859. [Google Scholar] [CrossRef]
- Motoshima, R.A.; Rosa, T.D.F.; Mendes, L.D.C.; Silva, E.V.D.; Viana, S.R.F.; Amaral, B.S.D.; de Souza, D.H.F.; Lião, L.M.; da Silva, M.L.C.; de Sousa, L.R.F.; et al. Inhibition of Leishmania amazonensis arginase by fucogalactan isolated from Agrocybe aegerita mushroom. Carbohydr. Polym. 2018, 201, 532–538. [Google Scholar] [CrossRef]
- Betancourt-Conde, I.; Avitia-Domínguez, C.; Hernández-Campos, A.; Castillo, R.; Yépez-Mulia, L.; Oria-Hernández, J.; Méndez, S.T.; Sierra-Campos, E.; Valdez-Solana, M.; Martínez-Caballero, S.; et al. Benzimidazole Derivatives as New and Selective Inhibitors of Arginase from Leishmania mexicana with Biological Activity against Promastigotes and Amastigotes. Int. J. Mol. Sci. 2021, 22, 3613. [Google Scholar] [CrossRef] [PubMed]
- Araújo, C.A.C.; Leon, L.L. Biological Activities of Curcuma longa L. Mem. Inst. Oswaldo Cruz 2021, 96, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, V.S. Turmeric—Chemistry, technology, and quality. Crit. Rev. Food Sci. Nutr. 1980, 12, 199–301. [Google Scholar] [CrossRef]
- Chauhan, I.S.; Rao, G.S.; Shankar, J.; Chauhan, L.K.S.; Kapadia, G.J.; Singh, N. Chemoprevention of Leishmaniasis: In-vitro antiparasitic activity of dibenzalacetone, a synthetic curcumin analog leads to apoptotic cell death in Leishmania donovani. Parasitol. Int. 2018, 67, 627–636. [Google Scholar] [CrossRef]
- Changtam, C.; de Koning, H.P.; Ibrahim, H.; Sajid, M.S.; Gould, M.K.; Suksamrarn, A. Curcuminoid analogs with potent activity against Trypanosoma and Leishmania species. Eur. J. Med. Chem. 2010, 45, 941–956. [Google Scholar] [CrossRef]
- Amaral, A.C.F.; Gomes, L.A.; Silva, J.R.A.; Ferreira, J.L.P.; Ramos, A.S.; Rosa, M.S.S.; Vermelho, A.B.; Rodrigues, I.A. Liposomal formulation of turmerone-rich hexane fractions from Curcuma longa enhances their antileishmanial activity. Biomed. Res. Int. 2014, 2014, 94934. [Google Scholar] [CrossRef]
- Dourado, D.; Silva, T.M.; do Nascimento, E.A.; Matos, E.S.; Formiga, F.R. Curcumin-loaded nanostructured systems for treatment of leishmaniasis: A review. Beilstein J. Nanotechnol. 2024, 15, 37–50. [Google Scholar] [CrossRef]
- Aqeeli, G.; Shayan, P.; Ebrahimzade, E.A.; Mohebali, M. Evaluation of curcumin and CM11 peptide alone and in combination against amastigote form of Iranian strain of L. major (MRHO/IR75/ER) in vitro. Exp. Parasitol. 2021, 229, 108151. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.; Pahuja, R.; Kumar, P.; Rath, S.K.; Gupta, K.C.; Goyal, N. Nanotized Curcumin and Miltefosine, a Potential Combination for Treatment of Experimental Visceral Leishmaniasis. Antimicrob. Agents Chemother. 2017, 61, e01169-16. [Google Scholar] [CrossRef]
- Pinto, J.G.; Fontana, L.C.; de Oliveira, M.A.; Kurachi, C.; Raniero, L.J.; Ferreira-Strixino, J. In vitro evaluation of photodynamic therapy using curcumin on Leishmania major and Leishmania braziliensis. Lasers Med. Sci. 2016, 31, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Marcolino, L.M.C.; Ambrosio, J.A.; Pinto, J.G.; Ferreira, I.; Simioni, A.R.; Ferreira-Strixino, J. Photodynamic therapy of cationic and anionic BSA-curcumin nanoparticles on amastigotes of Leishmania braziliensis and Leishmania major and Leishmania amazonensis. Photodiagnosis Photodyn. Ther. 2024, 46, 104001. [Google Scholar] [CrossRef]
- Das, R.; Roy, A.; Dutta, N.; Majumder, H.K. Reactive oxygen species and imbalance of calcium homeostasis contributes to curcumin induced programmed cell death in Leishmania donovani. Apoptosis 2008, 13, 867–882. [Google Scholar] [CrossRef]
- Ullah, W.; Wu, W.F.; Malak, N.; Nasreen, N.; Swelum, A.A.; Marcelino, L.A.; Niaz, S.; Khan, A.; Said, B.M.; Chen, C.C. Computational investigation of turmeric phytochemicals targeting PTR1 enzyme of Leishmania species. Heliyon 2024, 10, e27907. [Google Scholar] [CrossRef]
- Bounaama, A.; Djerdjouri, B.; Laroche-Clary, A.; Le Morvan, V.; Robert, J. Short curcumin treatment modulates oxidative stress, arginase activity, aberrant crypt foci, and TGF-β1 and HES-1 transcripts in 1,2-dimethylhydrazine-colon carcinogenesis in mice. Toxicology 2012, 302, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Elamin, M.; Al-Olayan, E.; Abdel-Gaber, R.; Yehia, R.S. Anti-proliferative and apoptosis induction activities of curcumin on Leishmania major. Rev. Argent. Microbiol. 2021, 53, 240–247. [Google Scholar] [CrossRef]
- Alonso, L.; Dorta, M.L.; Alonso, A. Ivermectin and curcumin cause plasma membrane rigidity in Leishmania amazonensis due to oxidative stress. Biochim. Biophys. Acta Biomembr. 2022, 1864, 183977. [Google Scholar] [CrossRef]
- Hoang, N.N.; Kodama, T.; Nakashima, Y.; Do, K.M.; Hnin, S.Y.Y.; Lee, Y.E.; Prema; Ikumi, N.; Morita, H. Arginase inhibitory activities of guaiane sesquiterpenoids from Curcuma comosa rhizomes. J. Nat. Med. 2023, 77, 891–897. [Google Scholar] [CrossRef]
- Mohseni, F.; Sharifi, I.; Oliaee, R.T.; Babaei, Z.; Mostafavi, M.; Almani, P.G.N.; Keyhani, A.; Salarkia, E.; Sharifi, F.; Nave, H.H.; et al. Antiproliferative properties of Turmerone on Leishmania major: Modes of action confirmed by antioxidative and immunomodulatory roles. Comp. Immunol. Microbiol. Infect. Dis. 2022, 84, 101797. [Google Scholar] [CrossRef]
- Teles, A.M.; Rosa, T.D.D.S.; Mouchrek, A.N.; Abreu-Silva, A.L.; Calabrese, K.D.S.; Almeida-Souza, F. Cinnamomum zeylanicum, Origanum vulgare, and Curcuma longa Essential Oils: Chemical Composition, Antimicrobial and Antileishmanial Activity. Evid. Based Complement. Alternat Med. 2019, 2019, 2421695. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Kar, S.K. Curcuminoids: The Novel Molecules of Nature. In Herbs and Species; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Camargo, P.G.; Bortoleti, B.T.S.; Fabris, M.; Gonçalves, M.D.; Tomiotto-Pellissier, F.; Costa, I.N.; Conchon-Costa, I.; Lima, C.H.S.; Pavanelli, W.R.; Bispo, M.L.F.; et al. Thiohydantoins as anti-leishmanial agents: In vitro biological evaluation and multi-target investigation by molecular docking studies. J. Biomol. Struct. Dyn. 2022, 40, 3213–3222. [Google Scholar] [CrossRef] [PubMed]
- Come, J.A.A.S.S.; Zhuang, Y.; Li, T.; Brogi, S.; Gemma, S.; Liu, T.; Silva, E.R. In vitro and in silico analyses of new cinnamid and rosmarinic acid-derived compounds biosynthesized in Escherichia coli as Leishmania amazonensis arginase inhibitors. Pathogens 2020, 11, 1020. [Google Scholar] [CrossRef] [PubMed]
- D’Antonio, E.L.; Ullman, B.; Roberts, S.C.; Dixit, U.G.; Wilson, M.E.; Hai, Y.; Christianson, D.W. Crystal structure of arginase from Leishmania Mexicana and implications for the inhibiton of polyamine biosynthesis in parasitic infections. Arch. Biochem. Biophys. 2013, 535, 163–176. [Google Scholar] [CrossRef]
- Santiago-Silva, K.M.; Bortoleti, B.T.S.; Brito, T.O.; Costa, I.C.; Lima, C.H.S.; Macedo Jr, F.; Miranda-Sapla, M.M.; Pavanelli, W.R.; Bispo, M.L.F. Exploring the antileishmanial activity of N1,N2-disubstituted-benzoylguanidines: Synthesis and molecular modeling studies. J. Biomol. Struct. Dyn. 2021, 40, 11495–11510. [Google Scholar] [CrossRef]
- Silva, E.R.; Come, J.A.A.S.S.; Brogi, S.; Calderone, V.; Chemi, G.; Campiani, G.; Oliveira, T.M.F.S.; Pham, T.N.; Pudlo, M.; Girard, C.; et al. Cinnamides target Leishmania amazonensis arginase selectively. Molecules 2020, 25, 5271. [Google Scholar] [CrossRef]
- Bohnert, T.; Gan, L.-S. Plasma protein binding: From discovery to development. J. Pharm. Sci. 2013, 102, 2953–2994. [Google Scholar] [CrossRef]
- Leung, M.H.M.; Kee, T.W. Effective stabilization of curcumin by association to plasma proteins: Human serum albumin and fibrinogen. Langmuir 2009, 25, 5773–5777. [Google Scholar] [CrossRef]
- Kar, T.; Basak, P.; Sen, S.; Ghosh, R.K.; Bhattacharyya, M. Analysis of curcumin interaction with human serum albumin using spectroscopic studies with molecular simulation. Front. Biol. 2017, 12, 199–209. [Google Scholar] [CrossRef]
- Seyfinejad, B.; Ozkan, S.A.; Jouyban, A. Recent advances in the determination of unbound concentration and plasma protein binding of drugs: Analytical methods. Talanta 2021, 225, 122052. [Google Scholar] [CrossRef]
- Chearwae, W.; Anuchapreeda, S.; Nandigama, K.; Ambudkar, S.V.; Limtrakul, P. Biochemical mechanism of modulation of human P-glycoprotein (ABCB1) by curcumin I, II, and III purified from Turmeric powder. Biochem. Pharmacol. 2004, 68, 2043–2052. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.G.; Cheng, S.W.; Yu, H.; Xu, Z.S.; Lee, J.K.; Hon, P.M.; Lee, M.Y.; Kennelly, E.J.; Deng, G.; Yeung, S.K.; et al. The role of turmerones on curcumin transportation and P-glycoprotein activities in intestinal Caco-2 cells. J. Med. Food 2012, 15, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Virmani, T.; Rachamalla, M.; Farasani, A.; Chigurupati, S.; Alsubayiel, A.M.; et al. In Vitro Phytochemical Screening, Cytotoxicity Studies of Curcuma longa Extracts with Isolation and Characterisation of Their Isolated Compounds. Molecules. 2021, 26, 7509. [Google Scholar] [CrossRef] [PubMed]
- Rahbardar, M.G.; Hosseinzadeh, H. The ameliorative effect of turmeric (Curcuma longa Linn) extract and its major constituent, curcumin, and its analogs on ethanol toxicity. Phytother. Res. 2024, 38, 2165–2181. [Google Scholar] [CrossRef]
- Cai, Y.; Huang, C.; Zhou, M.; Xu, S.; Xie, Y.; Gao, S.; Yang, Y.; Deng, Z.; Zhang, L.; Shu, J.; et al. Role of curcumin in the treatment of acute kidney injury: Research challenges and opportunities. Phytomedicine 2022, 104, 154306. [Google Scholar] [CrossRef]
- Soleimani, V.; Sahebkar, A.; Hosseinzadeh, H. Turmeric (Curcuma longa) and its major constituent (curcumin) as nontoxic and safe substances: Review. Phytother. Res. 2018, 32, 985–995. [Google Scholar] [CrossRef]
- Amin, K.; Dannenfelser, R.M. In vitro hemolysis: Guidance for the pharmaceutical scientist. J. Pharm. Sci. 2006, 95, 1173–1176. [Google Scholar] [CrossRef]
- Sandur, S.K.; Pandey, M.K.; Sung, B.; Ahn, K.S.; Murakami, A.; Sethi, G.; Limtrakul, P.; Badmaev, V.; Aggarwal, B.B. Curcumin, demethoxycurcumin, bisdemethoxycurcumin, tetrahydrocurcumin and turmerones differentially regulate anti-inflammatory and anti-proliferative responses through a ROS-independent mechanism. Carcinogenesis 2007, 28, 1765–1773. [Google Scholar] [CrossRef]
- Rasmussen, H.B.; Christensen, S.B.; Kvist, L.P.; Karazmi, A. A simple and efficient separation of the curcumins, the antiprotozoal constituents of Curcuma longa. Planta Med. 2000, 66, 396–398. [Google Scholar] [CrossRef]
- Chakravarty, J.; Sundar, S. Current and emerging medications for the treatment of leishmaniasis. Expert Opin. Pharmacother. 2019, 20, 1251–1265. [Google Scholar] [CrossRef] [PubMed]
- Araujo, C.A.; Alegrio, L.V.; Gomes, D.C.; Lima, M.E.; Gomes-Cardoso, L.; Leon, L.L. Studies on the effectiveness of diarylheptanoids derivatives against Leishmania amazonensis. Mem. Inst. Oswaldo Cruz 1999, 94, 791–794. [Google Scholar] [CrossRef] [PubMed]
- Saleheen, D.; Ali, S.A.; Ashfaq, K.; Siddiqui, A.A.; Agha, A.; Yasinzai, M.M. Latent activity of curcumin against leishmaniasis in vitro. Biol. Pharm. Bull. 2002, 25, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Olekhnovitch, R.; Bousso, P. Induction, Propagation, and Activity of Host Nitric Oxide: Lessons from Leishmania Infection. Trends Parasitol. 2015, 31, 653–664. [Google Scholar] [CrossRef]
- Chang, K.P. Leishmanicidal mechanisms of human polymorphonuclear phagocytes. Am. J. Trop. Med. Hyg. 1981, 30, 322–333. [Google Scholar] [CrossRef]
- Zhang, X.N.; Zhao, N.; Guo, F.F.; Wang, Y.R.; Liu, S.X.; Zeng, T. Diallyl disulfide suppresses the lipopolysaccharide-driven inflammatory response of macrophages by activating the Nrf2 pathway. Food Chem. Toxicol. 2022, 59, 112760. [Google Scholar] [CrossRef]
- Ayub, H.; Islam, M.; Saeed, M.; Ahmad, H.; Al-Asmari, F.; Ramadan, M.F.; Alissa, M.; Arif, M.A.; Rana, M.U.J.; Subtain, M.; et al. On the health effects of curcumin and its derivatives. Food Sci. Nutr. 2024, 12, 8623–8650. [Google Scholar] [CrossRef]
- Adapala, N.; Chan, M.M. Long-term use of an antiinflammatory, curcumin, suppressed type 1 immunity and exacerbated visceral leishmaniasis in a chronic experimental model. Lab. Investig. 2008, 88, 1329–1339. [Google Scholar] [CrossRef]
- Chan, M.M.; Adapala, N.S.; Fong, D. Curcumin overcomes the inhibitory effect of nitric oxide on Leishmania. Parasitol. Res. 2005, 96, 49–56. [Google Scholar] [CrossRef]
- Alinejad, S.; Khademvatan, S.; Amani, S.; Asadi, N.; Tappeh, K.H.; Yousefi, E.; Miandoabi, T. The Effect of Curcumin on the Expression of INFγ, TNF-α, and iNOS Genes in PBMCs Infected with Leishmania major [MRHO/IR/75/ER]. Infect. Disord. Drug Targets 2022, 22, e040422203031. [Google Scholar] [CrossRef]
- Clemente, C.M.; Murillo, J.; Garro, A.G.; Arbeláez, N.; Pineda, T.; Robledo, S.M.; Ravetti, S. Piperine, quercetin, and curcumin identified as promising natural products for topical treatment of cutaneous leishmaniasis. Parasitol. Res. 2024, 123, 185. [Google Scholar] [CrossRef]
- Sahebi, K.; Shahsavani, F.; Mehravar, F.; Hatam, G.; Alimi, R.; Radfar, A.; Bahreini, M.S.; Pouryousef, A.; Teimouri, A. In vitro and in vivo anti-parasitic activity of curcumin nanoemulsion on Leishmania major (MRHO/IR/75/ER). BMC Complement. Med. Ther. 2024, 24, 238. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rosell, G.; Giorgino, T.; Fabritiis, G.D. PlayMolecule ProteinPrepare: A web application for protein preparation for molecular dynamics simulations. J. Chem. Inf. Model. 2017, 57, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef]
- Benfenati, E.; Manganaro, A.; Gini, G. VEJA-QSAR:AI inside a platform for predictive toxicology. In Proceedings of the CEUR Workshop Proceedings, Popularize Artificial Intelligence, Turin, Italy, 5 December 2013; p. 1107. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 16, 55–63. [Google Scholar] [CrossRef]
- Corral, M.J.; González, E.; Cuquerella, M.; Alunda, J.M. Improvement of 96-well microplate assay for estimation of cell growth and inhibition of Leishmania with Alamar Blue. J. Microbiol. Methods 2013, 94, 111–116. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LiARG IC50 ± SE [µg/mL] | |
|---|---|
| HEXCURC | 14.4 ± 2.4 a |
| DCCURC | 10.04 ± 1.5 a |
| ETOHCURC | 52.64 ± 0.3 b |
| Curcumin (1) | 17.55 ± 0.4 a |
| Quercetin (7) * | <6.25 |
| Pharmacokinetic and Toxicological Endpoints | Compounds | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| Lipinski’s rule of 5 | 0 | 0 | 0 | 0 | 0 | 0 |
| Unbound (%) to plasma protein | 6.7 | 6.2 | 7.7 | 8.9 | 12.1 | 11.2 |
| P-glycoprotein interaction | Yes | Yes | Yes | No | Yes | Yes |
| Mutagenicity (Ames test) | No | No | No | No | No | No |
| Carcinogenicity (rats) | No | No | No | No | No | No |
| Acute toxicity (rats) | No | No | No | No | No | No |
| Cardiotoxicity (hERG) | No | No | No | No | No | No |
| Hepatotoxicity | No | No | No | No | Yes | Yes |
| RAW 264.7 | VERO | RBC | |
|---|---|---|---|
| HEXCURC | 47.87 ± 6.9 a,b | 60 ± 3 a | 119.65 ± 3.4 a |
| DCCURC | 55.08 ± 5.4 a,b | 58 ± 3.7 a | 101.85 ± 9.6 a |
| ETOHCURC | 70.16 ± 3.3 a | 89.3 ± 7.9 a | 144.15 ± 13.35 a |
| Curcumin (1) | 69.75 ± 2.8 a | n.d. | >200 |
| PAT | 30.8 ± 0.73 b | 23.9 ± 1.5 b | >100 |
| L. amazonensis | L. infantum | |||
|---|---|---|---|---|
| IC50 ± SE [µg/mL] | SI | IC50 ± SE [µg/mL] | SI | |
| HEXCURC | 102.3 ± 7.6 a | 0.47 | 69.6 ± 0.2 a | 0.7 |
| DCCURC | 59.6 ± 10.3 a,b | 0.9 | 28.2 ± 1.5 b | 1.95 |
| ETOHCURC | 53.3 ± 9.9 b,c | 1.3 | 74.7 ± 1.4 a | 0.94 |
| Curcumin (1) | 5.5 ± 0.45 d | 12.7 | 15.5 ± 1.7 c | 4.5 |
| PAT | 11.9 ± 3.1 c,d | 2.6 | <6.25 | >4.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spíndola, F.F.S.; Pinheiro, A.S.; Mpalantinos, M.A.; Silva, J.R.A.; Neto, W.S.M.F.; Conceição, R.A.; Barreto, E.M.; Abrahim-Vieira, B.A.; Rodrigues, C.R.; Souza, A.M.T.; et al. In Vitro and In Silico Assessments of Curcuminoids and Turmerones from Curcuma longa as Novel Inhibitors of Leishmania infantum Arginase. Pharmaceuticals 2025, 18, 851. https://doi.org/10.3390/ph18060851
Spíndola FFS, Pinheiro AS, Mpalantinos MA, Silva JRA, Neto WSMF, Conceição RA, Barreto EM, Abrahim-Vieira BA, Rodrigues CR, Souza AMT, et al. In Vitro and In Silico Assessments of Curcuminoids and Turmerones from Curcuma longa as Novel Inhibitors of Leishmania infantum Arginase. Pharmaceuticals. 2025; 18(6):851. https://doi.org/10.3390/ph18060851
Chicago/Turabian StyleSpíndola, Flora F. S., Anderson S. Pinheiro, Maria Athana Mpalantinos, Jefferson R. A. Silva, Walter S. M. F. Neto, Raissa A. Conceição, Eduarda M. Barreto, Barbara A. Abrahim-Vieira, Carlos R. Rodrigues, Alessandra M. T. Souza, and et al. 2025. "In Vitro and In Silico Assessments of Curcuminoids and Turmerones from Curcuma longa as Novel Inhibitors of Leishmania infantum Arginase" Pharmaceuticals 18, no. 6: 851. https://doi.org/10.3390/ph18060851
APA StyleSpíndola, F. F. S., Pinheiro, A. S., Mpalantinos, M. A., Silva, J. R. A., Neto, W. S. M. F., Conceição, R. A., Barreto, E. M., Abrahim-Vieira, B. A., Rodrigues, C. R., Souza, A. M. T., Nico, D., Amaral, A. C. F., Garcia, A. R., & Rodrigues, I. A. (2025). In Vitro and In Silico Assessments of Curcuminoids and Turmerones from Curcuma longa as Novel Inhibitors of Leishmania infantum Arginase. Pharmaceuticals, 18(6), 851. https://doi.org/10.3390/ph18060851