


Antibacterial Activity and Molecular Docking of Lignans Isolated from Artemisia cina Against Multidrug-Resistant Bacteria

 ,
,  , , , , , ,
, , , , , ,  and
and 
Abstract

1. Introduction
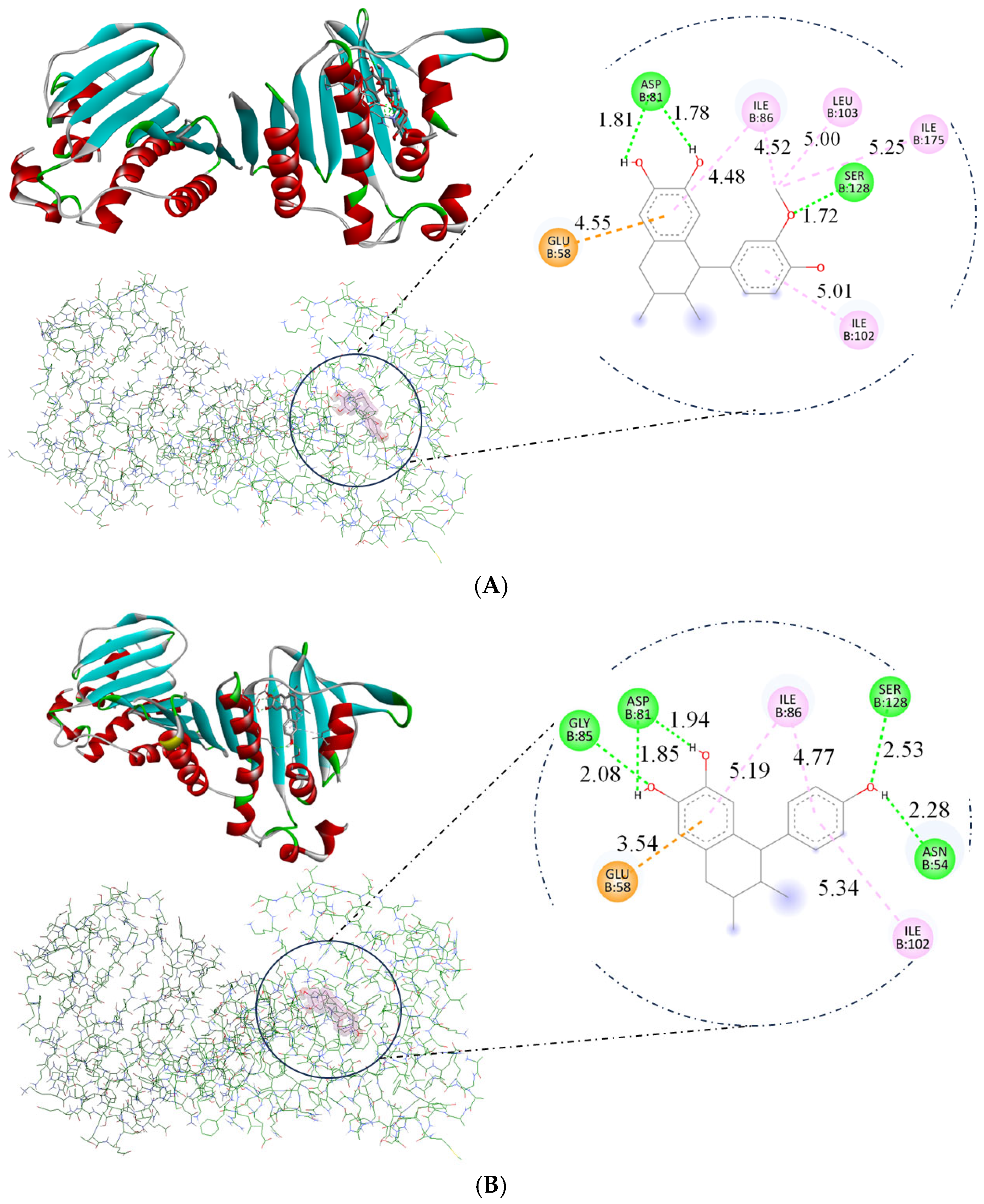
2. Results
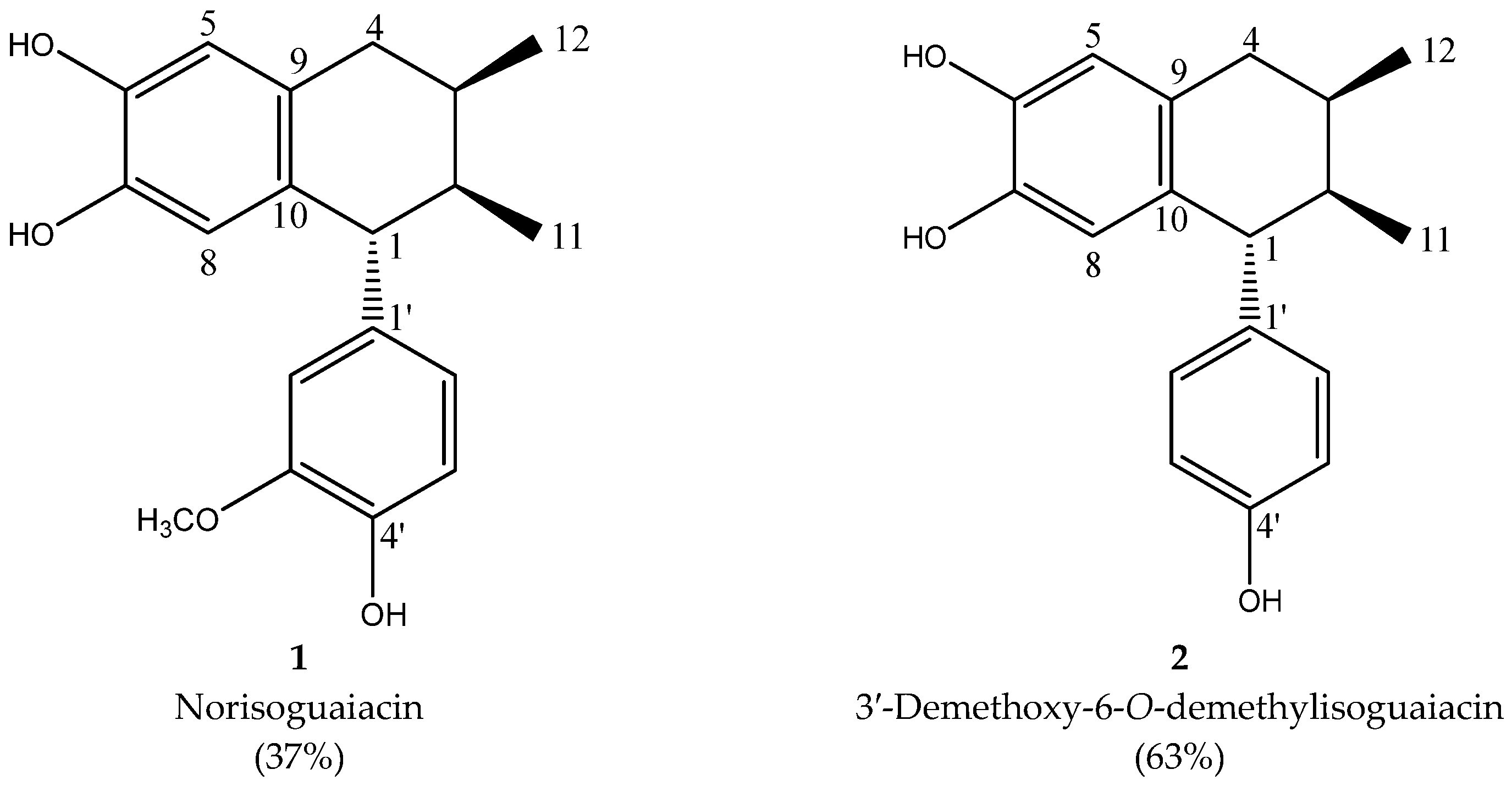
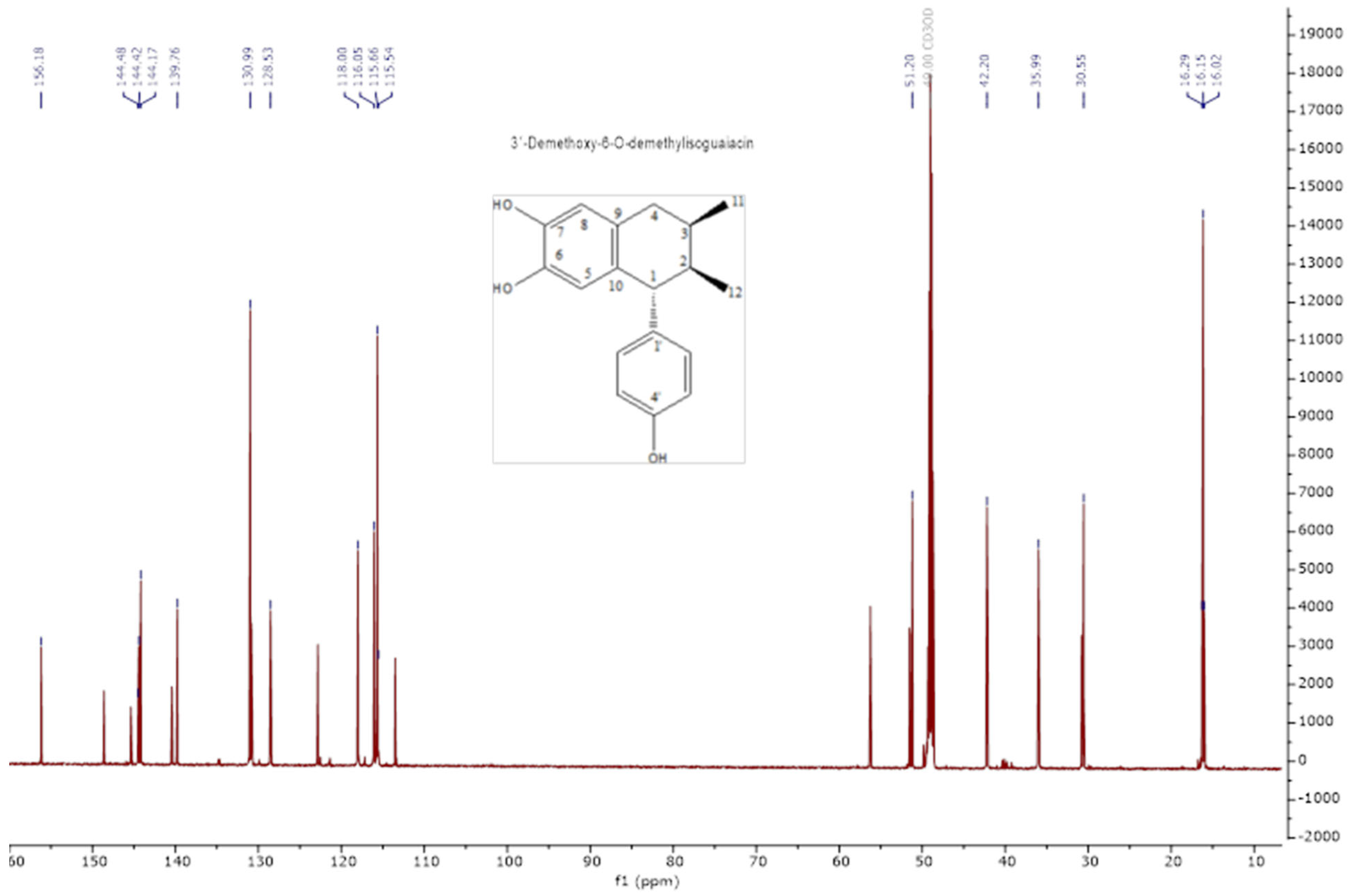
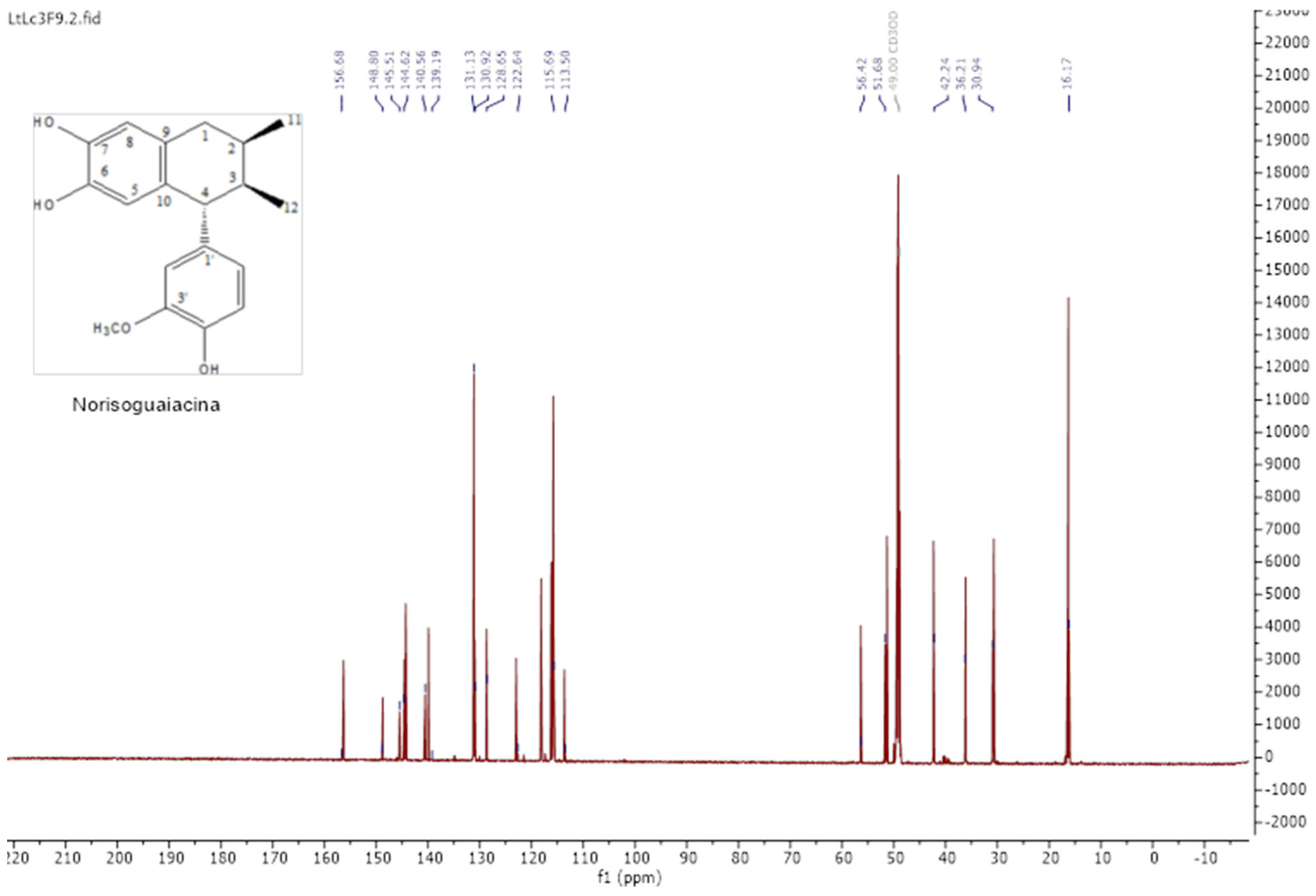
2.1. Extraction and Lignan Isolation of Artemisia cina
2.2. Antibacterial Activity
Minimum Inhibitory Concentration (MIC)
2.3. Minimum Bactericidal Concentration (MBC)
2.4. Time-Kill Kinetics Assay
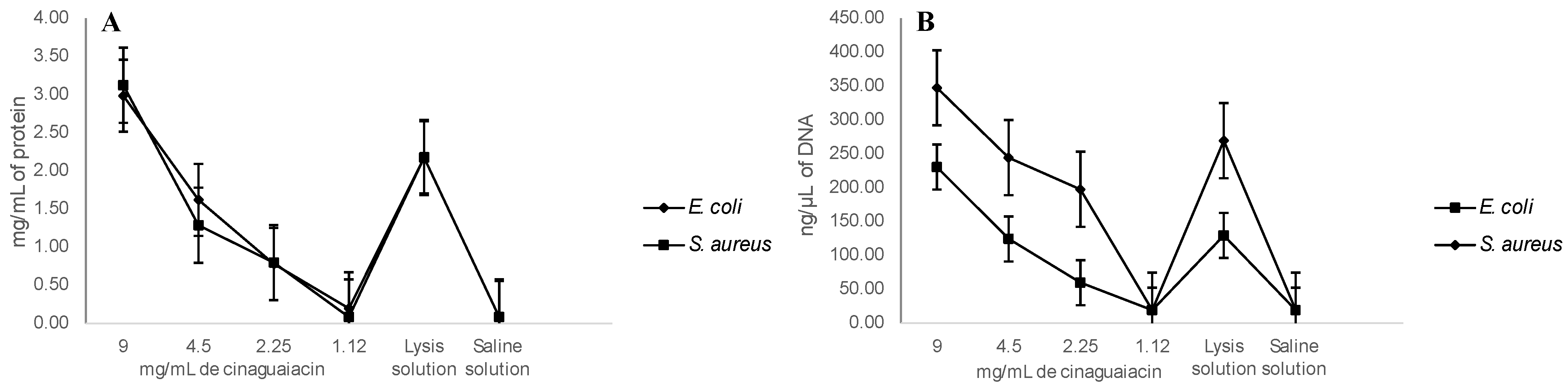
2.5. Cell Membrane Integrity and DNA Release
2.6. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction and Lignan Isolation of Artemisia cina
4.3. Bacterial Strains
4.4. Antibacterial Activity
4.4.1. Sterility Test
4.4.2. Reactivation of Bacterial Strains
4.4.3. Preparation of Inoculum
4.4.4. Minimum Inhibitory Concentration (MIC)
4.4.5. Minimum Bactericidal Concentration (MBC)
4.5. Time-Kill Kinetics Assay
4.6. Cell Membrane Integrity and DNA Release
4.7. Statistical Analysis
4.8. Quantum Chemical Calculations
4.9. DNA Gyrase B Preparation
4.10. Docking Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fair, R.J.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st Century. Perspect. Med. Chem. 2014, 6, S14459. [Google Scholar] [CrossRef] [PubMed]
- Marchioro, M.; Blank, M.d.F.A.; Mourão, R.H.V.; Antoniolli, Â.R. Anti-Nociceptive Activity of the Aqueous Extract of Erythrina velutina Leaves. Fitoterapia 2005, 76, 637–642. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Odey, T.O.J.; Tanimowo, W.O.; Afolabi, K.O.; Jahid, I.K.; Reuben, R.C. Antimicrobial Use and Resistance in Food Animal Production: Food Safety and Associated Concerns in Sub-Saharan Africa. Int. Microbiol. 2023, 27, 1–23. [Google Scholar] [CrossRef] [PubMed]
- de Lima, M.R.F.; Ximenes, E.C.A.; Luna, J.S.; Sant’Ana, A.E.G. The Antibiotic Activity of Some Brazilian Medicinal Plants. Rev. Bras. Farmacogn. 2006, 16, 300–306. [Google Scholar] [CrossRef]
- Fik-Jaskółka, M.; Mittova, V.; Motsonelidze, C.; Vakhania, M.; Vicidomini, C.; Roviello, G.N. Antimicrobial Metabolites of Caucasian Medicinal Plants as Alternatives to Antibiotics. Antibiotics 2024, 13, 487. [Google Scholar] [CrossRef] [PubMed]
- Alwathnani, H.A. Antibacterial Activity of Aqueous Extracts of Artemisia Species against Some Pathogenic Bacteria. Biosci. Biotechnol. Res. Asia 2017, 14, 621–624. [Google Scholar] [CrossRef]
- Morales-Ubaldo, A.L.; Gonzalez-Cortazar, M.; Zaragoza-Bastida, A.; Meza-Nieto, M.A.; Valladares-Carranza, B.; Alsayegh, A.A.; Batiha, G.E.-S.; Rivero-Perez, N. Nor 3′-Demethoxyisoguaiacin from Larrea Tridentata Is a Potential Alternative against Multidrug-Resistant Bacteria Associated with Bovine Mastitis. Molecules 2022, 27, 3620. [Google Scholar] [CrossRef]
- Bordean, M.-E.; Ungur, R.A.; Toc, D.A.; Borda, I.M.; Marțiș, G.S.; Pop, C.R.; Filip, M.; Vlassa, M.; Nasui, B.A.; Pop, A.; et al. Antibacterial and Phytochemical Screening of Artemisia Species. Antioxidants 2023, 12, 596. [Google Scholar] [CrossRef]
- Higuera-Piedrahita, R.I.; Dolores-Hernández, M.; de la Cruz-Cruz, H.A.; López-Arellano, R.; Gives, P.M.-D.; Olmedo-Juárez, A.; Cuéllar-Ordaz, J.A.; González-Cortazar, M.; Ble-González, E.A.; López-Arellano, M.E.; et al. 3′-Demethoxy-6-O-Demethylisoguaiacin and Norisoguaiacin Nematocidal Lignans from Artemisia cina against Haemonchus contortus Infective Larvae. Plants 2023, 12, 820. [Google Scholar] [CrossRef]
- Gnabre, J.N.; Ito, Y.; Ma, Y.; Huang, R.C. Isolation of Anti-HIV-1 Lignans from Larrea Tridentata by Counter-Current Chromatography. J. Chromatogr. A 1996, 719, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Gnabre, J.N. Extracts of Larrea tridentata Having Antiviral Activity and Their Use for Treating Viral Infections. Información de la Patente US No. 5989555, 23 November 1999. [Google Scholar]
- Garza-González, E. Use of 3’demethoxy-6-o-Demethyl Isoguaiacin as an Antibacterial Agent 2015. Mexican Patent Appling. Información de la patente MX: 2013009248A, 18 February 2015. [Google Scholar]
- Pardini, R.S.; Kim, C.H.; Blagini, R.; Morms, R.J.; Fletcher, D.C. Inhibition of Mitochondrial Electron-Transport Systems by nor-Isoguaiacin. Biochem. Pharmacol. 1973, 22, 1921–1925. [Google Scholar] [CrossRef] [PubMed]
- Gisvold, O.; Thaker, E. Lignans from Larrea divaricata. J. Pharm. Sci. 1974, 63, 1905–1907. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, S.H.; Amer, H.H. Synthesis, spectroscopic and molecular docking studies on new Schiff bases, nucleosides and α-aminophosphonate derivatives as antibacterial agents. Saudi J. Biol. Sci. 2020, 27, 3481–3488. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Patel, S.; Sharma, N.; Soisson, S.M.; Kishii, R.; Takei, M.; Fukuda, Y.; Lumb, K.J.; Singh, S.B. Structures of Kibdelomycin Bound to Staphylococcus aureus GyrB and ParE Showed a Novel U-Shaped Binding Mode. ACS Chem. Biol. 2014, 9, 2023–2031. [Google Scholar] [CrossRef]
- Dogra, S.; Koul, B.; Singh, J.; Mishra, M.; Yadav, D. Phytochemical Analysis, Antimicrobial Screening and In Vitro Pharmacological Activity of Artemisia vestita Leaf Extract. Molecules 2024, 29, 1829. [Google Scholar] [CrossRef]
- Molokoane, T.L.; Kemboi, D.; Siwe-Noundou, X.; Famuyide, I.M.; McGaw, L.J.; Tembu, V.J. Extractives from Artemisia Afra with Anti-Bacterial and Anti-Fungal Properties. Plants 2023, 12, 3369. [Google Scholar] [CrossRef]
- Phan, M.A.T.; Paterson, J.; Bucknall, M.; Arcot, J. Interactions between Phytochemicals from Fruits and Vegetables: Effects on Bioactivities and Bioavailability. Crit. Rev. Food Sci. Nutr. 2018, 58, 1310–1329. [Google Scholar] [CrossRef]
- Benamar-Aissa, B.; Gourine, N.; Ouinten, M.; Yousfi, M. Synergistic Effects of Essential Oils and Phenolic Extracts on Antimicrobial Activities Using Blends of Artemisia campestris, Artemisia herba alba, and Citrus aurantium. Biomol. Concepts 2024, 15, 20220040. [Google Scholar] [CrossRef]
- Buzgaia, N.; Awin, T.; Elabbar, F.; Abdusalam, K.; Lee, S.Y.; Rukayadi, Y.; Abas, F.; Shaari, K. Antibacterial Activity of Arbutus Pavarii Pamp against Methicillin-Resistant Staphylococcus aureus (MRSA) and UHPLC-MS/MS Profile of the Bioactive Fraction. Plants 2020, 9, 1539. [Google Scholar] [CrossRef]
- Haran, P.; Shanmugam, R.; Deenadayalan, P. Free Radical Scavenging, Anti-Inflammatory and Antibacterial Activity of Acorus calamus Leaves Extract Against Pseudomonas aeruginosa and Staphylococcus aureus. Cureus 2024, 16, e55987. [Google Scholar] [CrossRef] [PubMed]
- Umam, K.; Feng, C.-S.; Yang, G.; Tu, P.-C.; Lin, C.-Y.; Yang, M.-T.; Kuo, T.-F.; Yang, W.-C.; Minh, H.T.N. Phytochemistry, Pharmacology and Mode of Action of the Anti-Bacterial Artemisia Plants. Bioengineering 2023, 10, 633. [Google Scholar] [CrossRef] [PubMed]
- Favela-Hernández, J.M.J.; Clemente-Soto, A.F.; Balderas-Rentería, I.; Garza-González, E.; Camacho-Corona, M.D.R. Potential Mechanism of Action of 3′-Demethoxy-6-O-demethyl-isoguaiacin on Methicillin Resistant Staphylococcus aureus. Molecules 2015, 20, 12450–12458. [Google Scholar] [CrossRef]
- Luna-Vázquez, F.; Ibarra-Alvarado, C.; Camacho-Corona, M.; Rojas-Molina, A.; Rojas-Molina, J.; García, A.; Bah, M. Vasodilator Activity of Compounds Isolated from Plants Used in Mexican Traditional Medicine. Molecules 2018, 23, 1474. [Google Scholar] [CrossRef]
- Torres, R.; Urbina, F.; Morales, C.; Modak, B.; Monache, F.D. Antioxidant properties of lignans and ferulic acid from the resinous exudate of Larrea nitida. J. Chil. Chem. Soc. 2003, 48, 61–63. [Google Scholar] [CrossRef]
- Schmidt, T.J.; Rzeppa, S.; Kaiser, M.; Brun, R. Larrea Tridentata—Absolute Configuration of Its Epoxylignans and Investigations on Its Antiprotozoal Activity. Phytochem. Lett. 2012, 5, 632–638. [Google Scholar] [CrossRef]
- CLSI document M7-A5; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically (Approved Standards) CLSI. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012.
- Zaragoza-Bastida, A.; Flores-Aguilar, S.C.; Aguilar-Castro, L.M.; Morales-Ubaldo, A.L.; Valladares-Carranza, B.; Rangel-López, L.; Olmedo-Juarez, A.; Rosenfeld-Miranda, C.E.; Rivero-Perez, N. Antibacterial and Hemolytic Activity of Crotalus triseriatus and Crotalus ravus Venom. Animals 2020, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Olmedo-Juárez, A.; Briones-Robles, T.I.; Zaragoza-Bastida, A.; Zamilpa, A.; Ojeda-Ramírez, D.; Mendoza de Gives, P.; Olivares-Pérez, J.; Rivero-Perez, N. Antibacterial activity of compounds isolated from Caesalpinia coriaria (Jacq) Willd against important bacteria in public health. Microb. Pathog. 2019, 136, 103660. [Google Scholar] [CrossRef]
- Al-Mijalli, S.H.; El Hachlafi, N.; Abdallah, E.M.; Jeddi, M.; Assaggaf, H.; Qasem, A.; Alnasser, S.M.; Attar, A.; Naem, M.A.; Lee, L.-H.; et al. Exploring the Antibacterial Mechanisms of Chemically Characterized Essential Oils from Leaves and Buds of Syzygium aromaticum (L.) Merr. E. coli against Staphylococcus aureus and Pseudomonas aeruginosa. Ind. Crops Prod. 2023, 205, 117561. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | n-Hexane Extract (mg/mL) | Cinaguaiacin (mg/mL) | Kanamycin A (µg/mL) | |
|---|---|---|---|---|
| Reference ATCC | E. coli | NA | 1.12 bC | 1aB |
| S. typhi | 42.50 cC | 1.12 bC | 2 aC | |
| S. aureus | 21.25 cB | 1.12 bC | 2 aC | |
| L. monocytogenes | 21.25 cB | 0.28 bA | 0.5 aA | |
| Multidrug-resistant | B. cereus | 5.31 cA | 0.56 bB | 0.5 aA |
| K. pneumoniae | 42.50 cC | 1.12 bC | 2 aC | |
| E. coli01 | 42.50 cC | 1.12 bC | 2 aC | |
| S. aureus02 | 21.25 cB | 2.25 bC | 4 aD | |
| p-Value | 0.001 | 0.001 | 0.001 |
| Bacteria | n-Hexane Extract (mg/mL) | Cinaguaiacin (mg/mL) | Kanamicin A (µg/mL) | |
|---|---|---|---|---|
| Reference ATCC | E. coli | NA | NA | 2 aB |
| S. Typhi | NA | 2.25bB | 4 aC | |
| S. aureus | NA | 4.50 bC | 2 aB | |
| L. monocytogenes | NA | NA | 4 aC | |
| Multidrug-resistant | B. cereus | 0.62 bA | 1.12 CA | 1 aA |
| K. pneumoniae | NA | NA | 4 aC | |
| E. coli01 | NA | NA | 4 aC | |
| S. aureus02 | 85.0cB | 4.50 bC | 8 aD | |
| p-Value | 0.001 | 0.001 | 0.001 |
| Compound | ΔG (kcal/mol) | Amino Acids Interact with GyrB |
|---|---|---|
| Kanamycin | −9.6 | ASN54, ASP57, GLU58, ASP81, SER128 |
| 1 | −7.14 | GLU58, ASP81, ILE86, ILE102, LEU103, SER128, ILE175 |
| 2 | −7.12 | ASN54, GLU58, ASP81, GLY85, ILE86, ILE102, SER128 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García Hernández, L.C.; Higuera-Piedrahita, R.I.; Rivero-Perez, N.; Morales-Ubaldo, A.L.; Valladares-Carranza, B.; de la Cruz-Cruz, H.A.; Cuéllar-Ordaz, J.A.; González-Ruiz, C.; Nicolás-Vázquez, M.I.; Zaragoza-Bastida, A. Antibacterial Activity and Molecular Docking of Lignans Isolated from Artemisia cina Against Multidrug-Resistant Bacteria. Pharmaceuticals 2025, 18, 781. https://doi.org/10.3390/ph18060781
García Hernández LC, Higuera-Piedrahita RI, Rivero-Perez N, Morales-Ubaldo AL, Valladares-Carranza B, de la Cruz-Cruz HA, Cuéllar-Ordaz JA, González-Ruiz C, Nicolás-Vázquez MI, Zaragoza-Bastida A. Antibacterial Activity and Molecular Docking of Lignans Isolated from Artemisia cina Against Multidrug-Resistant Bacteria. Pharmaceuticals. 2025; 18(6):781. https://doi.org/10.3390/ph18060781
Chicago/Turabian StyleGarcía Hernández, Leslie Cynthia, Rosa Isabel Higuera-Piedrahita, Nallely Rivero-Perez, Ana Lizet Morales-Ubaldo, Benjamín Valladares-Carranza, Héctor Alejandro de la Cruz-Cruz, Jorge Alfredo Cuéllar-Ordaz, Cynthia González-Ruiz, María Inés Nicolás-Vázquez, and Adrian Zaragoza-Bastida. 2025. "Antibacterial Activity and Molecular Docking of Lignans Isolated from Artemisia cina Against Multidrug-Resistant Bacteria" Pharmaceuticals 18, no. 6: 781. https://doi.org/10.3390/ph18060781
APA StyleGarcía Hernández, L. C., Higuera-Piedrahita, R. I., Rivero-Perez, N., Morales-Ubaldo, A. L., Valladares-Carranza, B., de la Cruz-Cruz, H. A., Cuéllar-Ordaz, J. A., González-Ruiz, C., Nicolás-Vázquez, M. I., & Zaragoza-Bastida, A. (2025). Antibacterial Activity and Molecular Docking of Lignans Isolated from Artemisia cina Against Multidrug-Resistant Bacteria. Pharmaceuticals, 18(6), 781. https://doi.org/10.3390/ph18060781