
Natural Antidiabetic Agents: Insights into Ericaceae-Derived Phenolics and Their Role in Metabolic and Oxidative Modulation in Diabetes



Abstract
1. Introduction
2. Pathophysiology of Diabetes
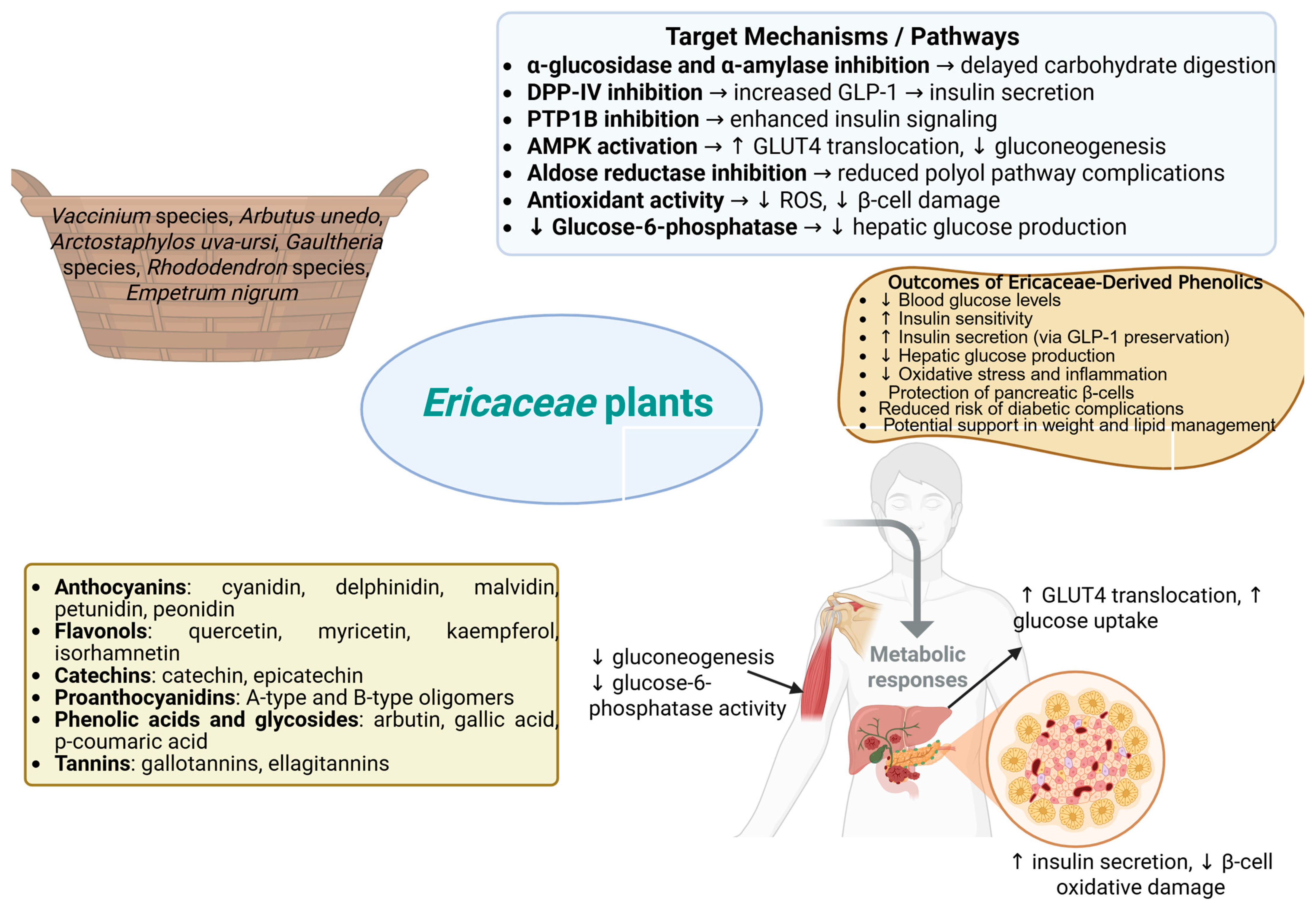
3. Chemical Composition of Ericaceae Plants with Antidiabetic Potential
4. Ericaceae Phenolic Compounds and Enzyme Inhibition in Diabetes
4.1. Inhibition of Digestive Enzymes (α-Glucosidase and α-Amylase)
4.2. DPP-IV Inhibition and Incretin Enhancement
4.3. Aldose Reductase Inhibition and Polyol Pathway Protection
4.4. PTP1B Inhibition and Insulin Signaling Enhancement
4.5. Activation of AMPK and Modulation of Glucose Metabolism
5. In Vitro and In Vivo Antidiabetic Studies for Vaccinium Species
5.1. Bilberry
5.2. Cranberry (V. sect. Oxycoccus)
5.3. Blueberry
5.4. Lingonberry (Vaccinium vitis-idaea L.)
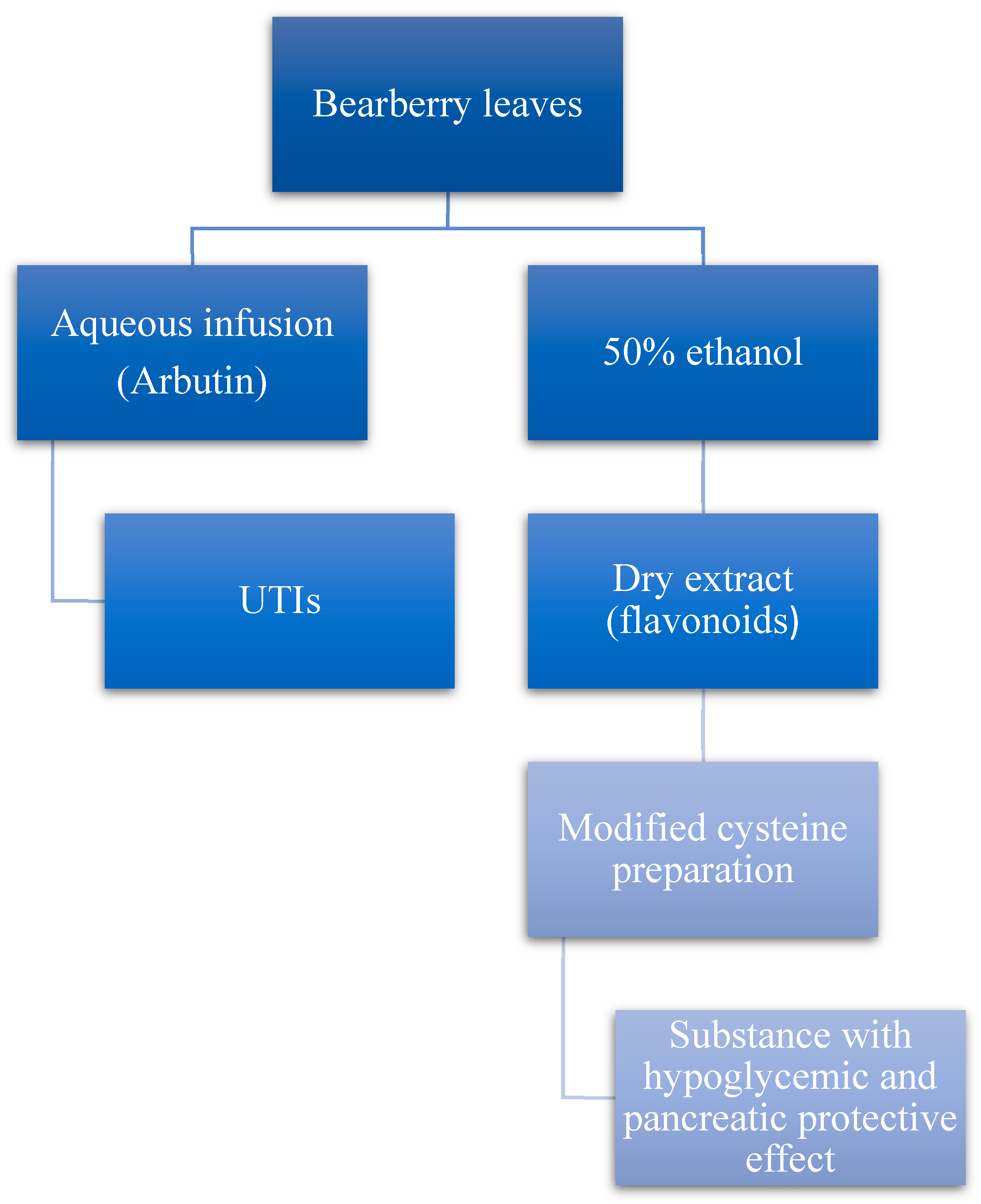
5.5. Bearberry (Arctostaphylos uva-ursi L.)
5.6. Arbutus unedo, (The Strawberry Tree)
5.7. Crowberry
6. Toxicity and Safety Considerations
7. Perspective
8. Methodology
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennici, G.; Almahasheer, H.; Alghrably, M.; Valensin, D.; Kola, A.; Kokotidou, C.; Lachowicz, J.; Jaremko, M. Mitigating Diabetes Associated with Reactive Oxygen Species (ROS) and Protein Aggregation through Pharmacological Interventions. RSC Adv. 2024, 14, 17448–17460. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Professional Practice Committee. Standards of Care in Diabetes-2024. Diabetes Care 2024, 47 (Suppl. 1), S20–S42. [Google Scholar] [CrossRef] [PubMed]
- NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in diabetes prevalence and treatment from 1990 to 2022: A pooled analysis of 1108 population-representative studies with 141 million participants. Lancet 2024, 404, 2077–2093. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Xie, Q.; Pan, X.; Zhang, R.; Zhang, X.; Peng, G.; Zhang, Y.; Shen, S.; Tong, N. Type 2 diabetes mellitus in adults: Pathogenesis, prevention and therapy. Signal Transduct. Target. Ther. 2024, 9, 262. [Google Scholar]
- Strati, M.; Moustaki, M.; Psaltopoulou, T.; Vryonidou, A.; Paschou, S.A. Early onset type 2 diabetes mellitus: An update. Endocrine 2024, 85, 965–978. [Google Scholar] [CrossRef] [PubMed]
- Kaul, N.; Ali, S. Genes, Genetics, and Environment in Type 2 Diabetes: Implication in Personalized Medicine. DNA Cell Biol. 2016, 35, 1–12. [Google Scholar] [CrossRef] [PubMed]
- GBD 2021 Diabetes Collaborators. Global, regional, and national burden of diabetes from 1990 to 2021, with projections of prevalence to 2050: A systematic analysis for the Global Burden of Disease Study 2021. Lancet 2023, 402, 203–234. [Google Scholar] [CrossRef]
- Alicic, R.Z.; Rooney, M.T.; Tuttle, K.R. Diabetic Kidney Disease: Challenges, Progress, and Possibilities. Clin. J. Am. Soc. Nephrol. 2017, 12, 2032–2045. [Google Scholar] [CrossRef]
- Bryl, A.; Mrugacz, M.; Falkowski, M.; Zorena, K. The Effect of Diet and Lifestyle on the Course of Diabetic Retinopathy—A Review of the Literature. Nutrients 2022, 14, 1252. [Google Scholar] [CrossRef]
- Jensen, T.S.; Karlsson, P.; Gylfadottir, S.S.; Andersen, S.T.; Bennett, D.L.; Tankisi, H.; Finnerup, N.B.; Terkelsen, A.J.; Khan, K.; Themistocleous, A.C.; et al. Painful and non-painful diabetic neuropathy, diagnostic challenges and implications for future management. Brain 2021, 144, 1632–1645. [Google Scholar] [CrossRef]
- Perumpail, B.J.; Khan, M.A.; Yoo, E.R.; Cholankeril, G.; Kim, D.; Ahmed, A. Clinical epidemiology and disease burden of nonalcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 8263–8276. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. 8. Pharmacologic Approaches to Glycemic Treatment: Standards of Medical Care in Diabetes-2018. Diabetes Care 2018, 41 (Suppl. S1), S73–S85. [Google Scholar] [CrossRef] [PubMed]
- Gattringer, T.; Posekany, A.; Niederkorn, K.; Knoflach, M.; Poltrum, B.; Mutzenbach, S.; Haring, H.P.; Ferrari, J.; Lang, W.; Willeit, J.; et al. Predicting Early Mortality of Acute Ischemic Stroke. Stroke 2019, 50, 349–356. [Google Scholar] [CrossRef]
- Lee, M.; Saver, J.L.; Hong, K.S.; Song, S.; Chang, K.H.; Ovbiagele, B. Effect of pre-diabetes on future risk of stroke: Meta-analysis. BMJ 2012, 344, e3564. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Hossain, K.S.; Das, S.; Kundu, S.; Adegoke, E.O.; Rahman, M.A.; Hannan, M.A.; Uddin, M.J.; Pang, M.G. Role of Insulin in Health and Disease: An Update. Int. J. Mol. Sci. 2021, 22, 6403. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.B.; Chiang, B.H. A Novel Phenolic Formulation for Treating Hepatic and Peripheral Insulin Resistance by Regulating GLUT4-Mediated Glucose Uptake. J. Tradit. Complement. Med. 2022, 12, 195–205. [Google Scholar] [CrossRef]
- Weinberg Sibony, R.; Segev, O.; Dor, S.; Raz, I. Drug Therapies for Diabetes. Int. J. Mol. Sci. 2023, 24, 17147. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Christopher, A.; Shetty, K. Phenolic Bioactives from Plant-Based Foods for Glycemic Control. Front. Endocrinol. 2022, 12, 727503. [Google Scholar] [CrossRef]
- Chhabria, S.; Mathur, S.; Vadakan, S.; Sahoo, D.K.; Mishra, P.; Paital, B. A Review on Phytochemical and Pharmacological Facets of Tropical Ethnomedicinal Plants as Reformed DPP-IV Inhibitors to Regulate Incretin Activity. Front. Endocrinol. 2022, 13, 1027237. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J. Elpel’s WebWorld Pages, Wildflowers-and-Weeds.com, November 2021. Available online: https://www.wildflowers-and-weeds.com/ (accessed on 25 March 2025).
- Sater, H.M.; Bizzio, L.N.; Tieman, D.M.; Muñoz, P.D. A Review of the Fruit Volatiles Found in Blueberry and Other Vaccinium Species. J. Agric. Food Chem. 2020, 68, 5777–5786. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Lee, H.J. Anti-Inflammatory Activity of Bilberry (Vaccinium myrtillus L.). Curr. Issues Mol. Biol. 2022, 44, 4570–4583. [Google Scholar] [CrossRef] [PubMed]
- Hasanloo, T.; Jafarkhani Kermani, M.; Dalvand, Y.A.; Rezazadeh, S. A Complete Review on the Genus Vaccinium and Iranian Ghareghat. J. Med. Plants 2019, 18, 46–65. [Google Scholar] [CrossRef]
- Niu, C.S.; Li, Y.; Liu, Y.B.; Ma, S.G.; Liu, F.; Li, L.; Xu, S.; Wang, X.J.; Wang, R.B.; Qu, J.; et al. Pierisketolide A and Pierisketones B and C, Three Diterpenes with an Unusual Carbon Skeleton from the Roots of Pieris formosa. Org. Lett. 2017, 19, 906–909. [Google Scholar] [CrossRef]
- Niu, C.-S.; Li, Y.; Liu, Y.-B.; Ma, S.-G.; Li, L.; Qu, J.; Yu, S.-S. Analgesic diterpenoids from the twigs of Pieris formosa. Tetrahedron 2016, 72, 44–49. [Google Scholar] [CrossRef]
- Niu, C.-S.; Li, Y.; Liu, Y.-B.; Ma, S.-G.; Liu, F.; Li, L.; Xu, S.; Wang, X.-J.; Liu, S.; Wang, R.-B.; et al. Biological and chemical guided isolation of 3,4-secograyanane diterpenoids from the roots of Pieris formosa. RSC Adv. 2017, 7, 43921–43932. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, T.; Zhang, H.; Zheng, G.; Qiu, Y.; Deng, M.; Zhang, C.; Yao, G. Anti-inflammatory Grayanane Diterpenoids from the Leaves of Rhododendron molle. J. Nat. Prod. 2018, 81, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.-S.; Li, Y.; Liu, Y.-B.; Ma, S.-G.; Liu, F.; Cui, L.; Yu, H.-B.; Wang, X.-J.; Qu, J.; Yu, S.-S. Grayanane diterpenoids with diverse bioactivities from the roots of Pieris formosa. Tetrahedron 2018, 74, 375–382. [Google Scholar] [CrossRef]
- Sun, N.; Zhu, Y.; Zhou, H.; Zhou, J.; Zhang, H.; Zhang, M.; Zeng, H.; Yao, G. Grayanane Diterpenoid Glucosides from the Leaves of Rhododendron micranthum and Their Bioactivities Evaluation. J. Nat. Prod. 2018, 81, 2673–2681. [Google Scholar] [CrossRef]
- Rabia, R.; Zaitoon, I.; Sajid, A.; Muhammad, N.; Muhammad, Y.K.; Jamshed, I. Identification of Highly Potent and Selective α-Glucosidase Inhibitors with Antiglycation Potential, Isolated from Rhododendron arboreum. Rec. Nat. Prod. 2015, 9, 262–266. [Google Scholar]
- Moyer, R.A.; Hummer, K.E.; Finn, C.E.; Frei, B.; Wrolstad, R.E. Anthocyanins, Phenolics, and Antioxidant Capacity in Diverse Small Fruits: Vaccinium, Rubus, and Ribes. J. Agric. Food Chem. 2002, 50, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Edger, P.P.; Iorizzo, M.; Bassil, N.V.; Benevenuto, J.; Ferrão, L.F.V.; Giongo, L.; Hummer, K.; Lawas, L.M.F.; Leisner, C.P.; Li, C.; et al. There and Back Again: Historical Perspective and Future Directions for Vaccinium Breeding and Research Studies. Hortic. Res. 2022, 9, uhac083. [Google Scholar] [CrossRef] [PubMed]
- Helmstädter, A.; Schuster, N. Vaccinium myrtillus as an Antidiabetic Medicinal Plant—Research through the Ages. Pharmazie 2010, 65, 315–321. [Google Scholar]
- Bujor, O.C.; Tanase, C.; Popa, M.E. Phenolic Antioxidants in Aerial Parts of Wild Vaccinium Species: Towards Pharmaceutical and Biological Properties. Antioxidants 2019, 8, 649. [Google Scholar] [CrossRef] [PubMed]
- Katsarou, A.; Gudbjörnsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, Å. Type 1 diabetes mellitus. Nat. Rev. Dis. Prim. 2017, 3, 1–17. [Google Scholar] [CrossRef]
- Fliegerová, K.O.; Mahayri, T.M.; Sechovcová, H.; Mekadim, C.; Mrázek, J.; Jarošíková, R.; Dubský, M.; Fejfarová, V. Diabetes and gut microbiome. Front. Microbiol. 2025, 15, 1451054. [Google Scholar] [CrossRef]
- Eiselein, L.; Schwartz, H.J.; Rutledge, J.C. The challenge of type 1 diabetes mellitus. ILAR J. 2004, 45, 231–236. [Google Scholar] [CrossRef]
- Bloem, S.J.; Roep, B.O. The elusive role of B lymphocytes and islet autoantibodies in (human) type 1 diabetes. Diabetologia 2017, 60, 1185–1189. [Google Scholar] [CrossRef]
- Campbell-Thompson, M.; Fu, A.; Kaddis, J.S.; Wasserfall, C.; Schatz, D.A.; Pugliese, A.; Atkinson, M.A. Insulitis and β-cell mass in the natural history of type 1 diabetes. Diabetes 2015, 65, 719–731. [Google Scholar] [CrossRef]
- Thingholm, L.B.; Rühlemann, M.C.; Koch, M.; Fuqua, B.; Laucke, G.; Boehm, R.; Bang, C.; Franzosa, E.A.; Hübenthal, M.; Rahnavard, G.; et al. Obese individuals with and without type 2 diabetes show different gut microbial functional capacity and composition. Cell Host Microbe 2019, 26, 252–264.e10. [Google Scholar] [CrossRef]
- Halban, P.A.; Polonsky, K.S.; Bowden, D.W.; Hawkins, M.A.; Ling, C.; Mather, K.J.; Powers, A.C.; Rhodes, C.J.; Sussel, L.; Weir, G.C. Beta-cell failure in type 2 diabetes: Postulated mechanisms and prospects for prevention and treatment. Diabetes Care 2014, 37, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.A.; Gannon, M. The Beta Cell in Type 2 Diabetes. Curr. Diabetes Rep. 2019, 19, 81. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, W.R.; Bone, R.N.; Sohn, P.; Syed, F.; Reissaus, C.A.; Mosley, A.L.; Wijeratne, A.B.; True, J.D.; Tong, X.; Kono, T.; et al. Endoplasmic reticulum stress alters ryanodine receptor function in the murine pancreatic beta cell. J. Biol. Chem. 2019, 294, 168–181. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Hussein, R.A.; El-Anssary, A.A. Plants secondary metabolites: The key drivers of the pharmacological actions of medicinal plants. In Herbal Medicine; Builders, P.F., Ed.; IntechOpen: London, UK, 2019; p. 76139. [Google Scholar]
- Wang, L.; Lan, W.; Chen, D. Blueberry (Vaccinium spp.) Anthocyanins and Their Functions, Stability, Bioavailability, and Applications. Foods 2024, 13, 2851. [Google Scholar] [CrossRef]
- Patanè, G.T.; Putaggio, S.; Tellone, E.; Barreca, D.; Ficarra, S.; Maffei, C.; Calderaro, A.; Laganà, G. Catechins and Proanthocyanidins Involvement in Metabolic Syndrome. Int. J. Mol. Sci. 2023, 24, 9228. [Google Scholar] [CrossRef] [PubMed]
- Boaru, D.L.; Fraile-Martinez, O.; De Leon-Oliva, D.; Garcia-Montero, C.; De Castro-Martinez, P.; Miranda-Gonzalez, A.; Saez, M.A.; Muñon-Zamarron, L.; Castillo-Ruiz, E.; Barrena-Blázquez, S.; et al. Harnessing the Anti-Inflammatory Properties of Polyphenols in the Treatment of Inflammatory Bowel Disease. Int. J. Biol. Sci. 2024, 20, 5608–5672. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, U.; Park, K.G. A Systematic Review of Oxidative Stress and Safety of Antioxidants in Diabetes: Focus on Islets and Their Defense. Diabetes Metab. J. 2013, 37, 106–112. [Google Scholar] [CrossRef]
- Amin, M.M.; Arbid, M.S. Estimation of ellagic acid and/or repaglinide effects on insulin signaling, oxidative stress, and inflammatory mediators of liver, pancreas, adipose tissue, and brain in insulin resistant/type 2 diabetic rats. Appl. Physiol. Nutr. Metab. 2017, 42, 181–192. [Google Scholar] [CrossRef]
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed. Pharmacother. 2017, 96, 305–312. [Google Scholar] [CrossRef]
- Eid, H.M.; Nachar, A.; Thong, F.; Sweeney, G.; Haddad, P.S. The molecular basis of the antidiabetic action of quercetin in cultured skeletal muscle cells and hepatocytes. Pharmacogn. Mag. 2015, 11, 74–81. [Google Scholar] [PubMed]
- Stefanescu, R.; Laczkó-Zöld, E.; O˝sz, B.E.; Vari, C.E. An Updated Systematic Review of Vaccinium myrtillus Leaves: Phytochemistry and Pharmacology. Pharmaceutics 2022, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Martau, G.A.; Bernadette-Emoke, T.; Odocheanu, R.; Soporan, D.A.; Bochis, M.; Simon, E.; Vodnar, D.C. Vaccinium Species (Ericaceae): Phytochemistry and Biological Properties of Medicinal Plants. Molecules 2023, 28, 1533. [Google Scholar] [CrossRef]
- Huang, H.; Luo, Y.; Wang, Q.; Zhang, Y.; Li, Z.; He, R.; Chen, X.; Dong, Z. Vaccinium as Potential Therapy for Diabetes and Microvascular Complications. Nutrients 2023, 15, 2031. [Google Scholar] [CrossRef]
- Shamilova, A.A.; Bubenchikova, V.N.; Chernikov, M.V.; Pozdnyakov, D.I.; Garsiya, E.R.; Larsky, M.V. Bearberry (Arctostaphylos uva-ursi (L.) Spreng.): Chemical Content and Pharmacological Activity. J. Excip. Food Chem. 2021, 12, 3. [Google Scholar]
- Ockun, M.A.; Baranauskaite, J.; Uner, B.; Kan, Y.; Kırmızıbekmez, H. Preparation, characterization and evaluation of liposomal-freeze dried anthocyanin-enriched Vaccinium arctostaphylos L. fruit extract incorporated into fast dissolving oral films. J. Drug Deliv. Sci. Technol. 2022, 72, 103428. [Google Scholar] [CrossRef]
- Alam, F.; Saqib, Q.N.; Ashraf, M. Gaultheria trichophylla (Royle): A Source of Minerals and Biologically Active Molecules, Its Antioxidant and Anti-Lipoxygenase Activities. BMC Complement. Altern. Med. 2017, 17, 3. [Google Scholar] [CrossRef]
- Sharma, M.; Gargi, A.; Borah, A. Rhododendron arboreum and Its Potential Health Benefit: A Review. Pharma Innov. J. 2022, SP-11, 926–933. [Google Scholar]
- Eid, H.M.; Ouchfoun, M.; Saleem, A.; Guerrero-Analco, J.A.; Walshe-Roussel, B.; Musallam, L.; Rapinski, M.; Cuerrier, A.; Martineau, L.C.; Arnason, J.T.; et al. A Combination of (+)-Catechin and (−)-Epicatechin Underlies the In vitro Adipogenic Action of Labrador Tea (Rhododendron groenlandicum), an Antidiabetic Medicinal Plant of the Eastern James Bay Cree Pharmacopeia. J. Ethnopharmacol. 2016, 178, 251–257. [Google Scholar] [CrossRef]
- Bebek Markovinović, A.; Brčić Karačonji, I.; Jurica, K.; Lasić, D.; Skendrović Babojelić, M.; Duralija, B.; Šic Žlabur, J.; Putnik, P.; Bursać Kovačević, D. Strawberry Tree Fruits and Leaves (Arbutus unedo L.) as Raw Material for Sustainable Functional Food Processing: A Review. Horticulturae 2022, 8, 881. [Google Scholar] [CrossRef]
- Jurikova, T.; Mlcek, J.; Skrovankova, S.; Balla, S.; Sochor, J.; Baron, M.; Sumczynski, D. Black Crowberry (Empetrum nigrum L.) Flavonoids and Their Health Promoting Activity. Molecules 2016, 21, 1685. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5280343, Quercetin. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Quercetin (accessed on 25 March 2025).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5280863, Kaempferol. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Kaempferol (accessed on 25 March 2025).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5281672, Myricetin. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Myricetin (accessed on 25 March 2025).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 128861, Cyanidin. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Cyanidin (accessed on 25 March 2025).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 68245, Delphinidin. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Delphinidin (accessed on 25 March 2025).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 159287, CID 159287. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Malvidin (accessed on 25 March 2025).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 73386, Petunidin. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Petunidin (accessed on 25 March 2025).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 67249, Pelargonidin Chloride. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Pelargonidin-Chloride (accessed on 25 March 2025).
- Celik, F.; Bozhuyuk, M.R.; Ercisli, S.; Gundogdu, M. Physicochemical and Bioactive Characteristics of Wild Grown Bilberry (Vaccinium myrtillus L.) Genotypes from Northeastern Turkey. Not. Bot. Horti Agrobot. 2018, 46, 128–133. [Google Scholar] [CrossRef]
- Jurikova, T.; Skrovankova, S.; Mlcek, J.; Balla, S.; Snopek, L. Bioactive Compounds, Antioxidant Activity, and Biological Effects of European Cranberry (Vaccinium oxycoccos). Molecules 2019, 24, 24. [Google Scholar] [CrossRef]
- Urbonaviciene, D.; Bobinaite, R.; Viskelis, P.; Viskelis, J.; Petruskevicius, A.; Puzeryte, V.; Cesoniene, L.; Daubaras, R.; Klavins, L.; Bobinas, C. Nutritional and Physicochemical Properties of Wild Lingonberry (Vaccinium vitis-idaea L.)—Effects of Geographic Origin. Molecules 2023, 28, 4589. [Google Scholar] [CrossRef] [PubMed]
- Sugier, P.; Sęczyk, Ł.; Sugier, D.; Krawczyk, R.; Wójcik, M.; Czarnecka, J.; Okoń, S.; Plak, A. Chemical Characteristics and Antioxidant Activity of Arctostaphylos uva-ursi L. Spreng. at the Southern Border of the Geographical Range of the Species in Europe. Molecules 2021, 26, 7692. [Google Scholar] [CrossRef]
- Özgen, M.; Çelik, H.; Saraçoğlu, O. Less known Vaccinium: Antioxidant and chemical properties of selected Caucasian whortleberry (Vaccinium arctostaphylos L.) fruits native to Black Sea region of Turkey. Acta Sci. Pol. Hortorum Cultus 2014, 13, 59–68. [Google Scholar]
- Bahukhandi, A.; Pandey, A.; Sekar, K.C.; Bhatt, I.D. Polyphenolics, nutrients and antioxidant activity of Gaultheria trichophylla Royle: A high value wild edible plant of Trans Himalaya. Horticult. Int. J. 2017, 1, 39–43. [Google Scholar]
- Barola, A.; Tewari, A.; Kabdwal, N.C. Effect of pre-treatments and drying methods on nutritional quality of Rhododendron arboreum, West Himalaya. J. Adv. Sci. Res. 2022, 13, 121–128. [Google Scholar] [CrossRef]
- Dampc, A.; Luczkiewicz, M. Labrador tea—The aromatic beverage and spice: A review of origin, processing and safety. J. Sci. Food Agric. 2015, 95, 1577–1583. [Google Scholar] [CrossRef]
- Brčić Karačonji, I.; Jurica, K.; Gašić, U.; Dramićanin, A.; Tešić, Ž.; Milojković Opsenica, D. Comparative Study on the Phenolic Fingerprint and Antioxidant Activity of Strawberry Tree (Arbutus unedo L.) Leaves and Fruits. Plants 2021, 11, 25. [Google Scholar] [CrossRef]
- Urbonaviciene, D.; Bobinaite, R.; Viskelis, P.; Bobinas, C.; Petruskevicius, A.; Klavins, L.; Viskelis, J. Geographic Variability of Biologically Active Compounds, Antioxidant Activity and Physico-Chemical Properties in Wild Bilberries (Vaccinium myrtillus L.). Antioxidants 2022, 11, 588. [Google Scholar] [CrossRef] [PubMed]
- Karcheva-Bahchevanska, D.; Nikolova, M.; Iliev, I. Inhibitory Potential of Different Bilberry (Vaccinium myrtillus L.) Extracts on Human Salivary α-Amylase. Molecules 2023, 28, 5820. [Google Scholar] [CrossRef]
- Nemzer, B.V.; Al-Taher, F.; Yashin, A.; Revelsky, I.; Yashin, Y. Cranberry: Chemical Composition, Antioxidant Activity and Impact on Human Health: Overview. Molecules 2022, 27, 1503. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Di Stefano, V.; Lauria, A.; Pitonzo, R.; Gentile, C. Vaccinium macrocarpon (Cranberry)-Based Dietary Supplements: Variation in Mass Uniformity, Proanthocyanidin Dosage and Anthocyanin Profile Demonstrates Quality Control Standard Needed. Nutrients 2020, 12, 992. [Google Scholar] [CrossRef]
- Bernal-Gallardo, J.O.; Mena-Violante, H.G.; Luna-Suárez, S. Study of the Phenolic Compounds and Biological Activities of the Wild Fruits of Vaccinium leucanthum Schltdl. Horticulturae 2024, 10, 1091. [Google Scholar] [CrossRef]
- Wang, S.; Penhallegon, R. Antioxidant Activity in Lingonberries (Vaccinium vitis-idaea L.). HortScience 2005, 40, 1090C. [Google Scholar] [CrossRef]
- Sugier, P.; Sęczyk, Ł.; Sugier, D. Variation in Population and Solvents as Factors Determining the Chemical Composition and Antioxidant Potential of Arctostaphylos uva-ursi (L.) Spreng. Leaf Extracts. Molecules 2022, 27, 2247. [Google Scholar] [CrossRef]
- Barut, B.; Barut, E.N.; Engin, S.; Özel, A.; Sezen, F.S. Investigation of the antioxidant, α-glucosidase inhibitory, anti-inflammatory, and DNA protective properties of Vaccinium arctostaphylos L. Turk. J. Pharm. Sci. 2019, 16, 175–183. [Google Scholar] [CrossRef]
- Grace, M.H.; Esposito, D.; Dunlap, K.L.; Lila, M.A. Comparative Analysis of Phenolic Content and Profile, Antioxidant Capacity, and Anti-inflammatory Bioactivity in Wild Alaskan and Commercial Vaccinium Berries. J. Agric. Food Chem. 2014, 62, 4007–4017. [Google Scholar] [CrossRef]
- Dufour, D.; Pichette, A.; Mshvildadze, V.; Bradette-Hébert, M.E.; Lavoie, S.; Longtin, A.; Laprise, C.; Legault, J. Antioxidant, anti-inflammatory and anticancer activities of methanolic extracts from Ledum groenlandicum Retzius. J. Ethnopharmacol. 2007, 111, 22–28. [Google Scholar] [CrossRef]
- Ogawa, K.; Sakakibara, H.; Iwata, R.; Ishii, T.; Sato, T.; Goda, T.; Shimoi, K.; Kumazawa, S. Anthocyanin Composition and Antioxidant Activity of the Crowberry (Empetrum nigrum) and Other Berries. J. Agric. Food Chem. 2008, 56, 4457–4462. [Google Scholar] [CrossRef] [PubMed]
- Moein, S.; Moein, M.; Javid, H. Inhibition of α-Amylase and α-Glucosidase of Anthocyanin Isolated from Berberis integerrima Bunge Fruits: A Model of Antidiabetic Compounds. Evid. Based Complement. Alternat. Med. 2022, 2022, 6529590. [Google Scholar] [CrossRef] [PubMed]
- Niisato, N.; Marunaka, Y. Therapeutic Potential of Multifunctional Myricetin for Treatment of Type 2 Diabetes Mellitus. Front. Nutr. 2023, 10, 1175660. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, F.; Mahjoub, S.; Pouramir, M.; Khadir, F. Hypoglycemic Activity of Pyrus biossieriana Buhse Leaf Extract and Arbutin: Inhibitory Effects on Alpha-Amylase and Alpha-Glucosidase. Casp. J. Intern. Med. 2013, 4, 763–767. [Google Scholar]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; de Mejia, E.G. Berry and Citrus Phenolic Compounds Inhibit Dipeptidyl Peptidase IV: Implications in Diabetes Management. Evid. Based Complement. Alternat. Med. 2013, 2013, 479505. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Alkhalidy, H.; Liu, D. The Emerging Role of Polyphenols in the Management of Type 2 Diabetes. Molecules 2021, 26, 703. [Google Scholar] [CrossRef]
- Varma, S.D.; Mikuni, I.; Kinoshita, J.H. Flavonoids as Inhibitors of Lens Aldose Reductase. Science 1975, 188, 1215–1216. [Google Scholar] [CrossRef]
- Al Mamun, M.A.; Rakib, A.; Mandal, M.; Kumar, S.; Singla, B.; Singh, U.P. Polyphenols: Role in Modulating Immune Function and Obesity. Biomolecules 2024, 14, 221. [Google Scholar] [CrossRef]
- Kopystecka, A.; Kozioł, I.; Radomska, D.; Bielawski, K.; Bielawska, A.; Wujec, M. Vaccinium uliginosum and Vaccinium myrtillus—Two Species, One Used as a Functional Food. Nutrients 2023, 15, 4119. [Google Scholar] [CrossRef]
- Ansari, P.; Khan, J.T.; Chowdhury, S.; Reberio, A.D.; Kumar, S.; Seidel, V.; Abdel-Wahab, Y.H.A.; Flatt, P.R. Plant-Based Diets and Phytochemicals in the Management of Diabetes Mellitus and Prevention of Its Complications: A Review. Nutrients 2024, 16, 3709. [Google Scholar] [CrossRef]
- Tzeng, T.F.; Liou, S.S.; Liu, I.M. Myricetin Ameliorates Defective Post-Receptor Insulin Signaling via β-Endorphin Signaling in the Skeletal Muscles of Fructose-Fed Rats. Evid. Based Complement. Alternat. Med. 2011, 2011, 150752. [Google Scholar] [CrossRef]
- Li, M.; Ding, L.; Cao, L.; Zhang, Z.; Li, X.; Li, Z.; Xia, Q.; Yin, K.; Song, S.; Wang, Z.; et al. Natural Products Targeting AMPK Signaling Pathway Therapy, Diabetes Mellitus and Its Complications. Front. Pharmacol. 2025, 16, 1534634. [Google Scholar]
- Feng, S.Y.; Wu, S.J.; Chang, Y.C.; Ng, L.T.; Chang, S.J. Stimulation of GLUT4 Glucose Uptake by Anthocyanin-Rich Extract from Black Rice (Oryza sativa L.) via PI3K/Akt and AMPK/p38 MAPK Signaling in C2C12 Cells. Metabolites 2022, 12, 856. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Yan, H.; Fei, X.; Liu, H.; Wu, J. Modulation of Glucose Metabolism by a Natural Compound from Chloranthus japonicus via Activation of AMP-Activated Protein Kinase. Sci. Rep. 2017, 7, 778. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Tenuta, M.C.; Loizzo, M.R.; Bonesi, M.; Finetti, F.; Trabalzini, L.; Deguin, B. Vaccinium species (Ericaceae): From chemical composition to bio-functional activities. Appl. Sci. 2021, 11, 5655. [Google Scholar] [CrossRef]
- Wronska-Pilarek, D.; Szkudlarz, P.; Bocianowski, J. Systematic importance of morphological features of pollen grains of species from Erica (Ericaceae) genus. PLoS ONE 2018, 13, e0204557. [Google Scholar] [CrossRef] [PubMed]
- Gailīte, A.; Gaile, A.; Rungis, D.E. Genetic diversity and structure of wild Vaccinium populations—V. myrtillus, V. vitis-idaea and V. uliginosum in the Baltic states. Silva Fenn. 2020, 54, 10396. [Google Scholar] [CrossRef]
- Kulkarni, K.P.; Vorsa, N.; Natarajan, P.; Elavarthi, S.; Iorizzo, M.; Reddy, U.K.; Melmaiee, K. Admixture analysis using genotyping-by-sequencing reveals genetic relatedness and parental lineage distribution in highbush blueberry genotypes and cross derivatives. Int. J. Mol. Sci. 2021, 22, 163. [Google Scholar] [CrossRef]
- Mitrea, L.; Nemes, S.-A.; Szabo, K.; Teleky, B.-E.; Vodnar, D.-C. Guts Imbalance Imbalances the Brain: A Review of Gut Microbiota Association With Neurological and Psychiatric Disorders. Front. Med. 2022, 9, 813204. [Google Scholar] [CrossRef]
- Aaby, K.; Grimmer, S.; Holtung, L. Extraction of phenolic compounds from bilberry (Vaccinium myrtillus L.) press residue: Effects on phenolic composition and cell proliferation. LWT 2013, 54, 257–264. [Google Scholar] [CrossRef]
- Basu, A.; Feng, D.; Planinic, P.; Ebersole, J.L.; Lyons, T.J.; Alexander, J.M. Dietary Blueberry and Soluble Fiber Supplementation Reduces Risk of Gestational Diabetes in Women with Obesity in a Randomized Controlled Trial. J. Nutr. 2021, 151, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Stote, K.; Corkum, A.; Sweeney, M.; Shakerley, N.; Kean, T.; Gottschall-Pass, K. Postprandial Effects of Blueberry (Vaccinium angustifolium) Consumption on Glucose Metabolism, Gastrointestinal Hormone Response, and Perceived Appetite in Healthy Adults: A Randomized, Placebo-Controlled Crossover Trial. Nutrients 2019, 11, 202. [Google Scholar] [CrossRef]
- Stote, K.S.; Wilson, M.M.; Hallenbeck, D.; Thomas, K.; Rourke, J.M.; Sweeney, M.I.; Gottschall-Pass, K.T.; Gosmanov, A.R. Effect of Blueberry Consumption on Cardiometabolic Health Parameters in Men with Type 2 Diabetes: An 8-Week, Double-Blind, Randomized, Placebo-Controlled Trial. Curr. Dev. Nutr. 2020, 4, nzaa030. [Google Scholar] [CrossRef] [PubMed]
- Mirfeizi, M.; Mehdizadeh Tourzani, Z.; Mirfeizi, S.Z.; Asghari Jafarabadi, M.; Rezvani, H.R.; Afzali, M. Controlling type 2 diabetes mellitus with herbal medicines: A triple-blind randomized clinical trial of efficacy and safety. J. Diabetes 2016, 8, 647–656. [Google Scholar] [CrossRef]
- de Mello, V.D.; Lankinen, M.A.; Lindström, J.; Puupponen-Pimiä, R.; Laaksonen, D.E.; Pihlajamäki, J.; Lehtonen, M.; Uusitupa, M.; Tuomilehto, J.; Kolehmainen, M.; et al. Fasting serum hippuric acid is elevated after bilberry (Vaccinium myrtillus) consumption and associates with improvement of fasting glucose levels and insulin secretion in persons at high risk of developing type 2 diabetes. Mol. Nutr. Food Res. 2017, 61, 1700019. [Google Scholar] [CrossRef]
- Chan, S.W.; Chu, T.T.W.; Choi, S.W.; Benzie, I.F.F.; Tomlinson, B. Impact of short-term bilberry supplementation on glycemic control, cardiovascular disease risk factors, and antioxidant status in Chinese patients with type 2 diabetes. Phytother. Res. 2021, 35, 3236–3245. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ling, W.; Yang, Y.; Chen, Y.; Tian, Z.; Du, Z.; Chen, J.; Xie, Y.; Liu, Z.; Yang, L. Role of Purified Anthocyanins in Improving Cardiometabolic Risk Factors in Chinese Men and Women with Prediabetes or Early Untreated Diabetes—A Randomized Controlled Trial. Nutrients 2017, 9, 1104. [Google Scholar] [CrossRef]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef]
- Schell, J.; Betts, N.M.; Foster, M.; Scofield, R.H.; Basu, A. Cranberries improve postprandial glucose excursions in type 2 diabetes. Food Funct. 2017, 8, 3083–3090. [Google Scholar] [CrossRef] [PubMed]
- Novotny, J.A.; Baer, D.J.; Khoo, C.; Gebauer, S.K.; Charron, C.S. Cranberry juice consumption lowers markers of cardiometabolic risk, including blood pressure and circulating C-reactive protein, triglyceride, and glucose concentrations in adults. J. Nutr. 2015, 145, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Antonella, S.; Barreca, D.; Giuseppina, L.; Ersilia, B.; Domenico, T. Bilberry (Vaccinium myrtyllus L.); Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128124918. [Google Scholar]
- Carvalho, M.; Matos, M.; Carnide, V. Identification of cultivated and wild Vaccinium species grown in Portugal. Span. J. Agric. Res. 2018, 16, e07SC01-01. [Google Scholar] [CrossRef]
- Vaneková, Z.; Rollinger, J.M. Bilberries: Curative and Miraculous—A Review on Bioactive Constituents and Clinical Research. Front. Pharmacol. 2022, 13, 2343. [Google Scholar] [CrossRef] [PubMed]
- Stanoeva, J.P.; Stefova, M.; Andonovska, K.B.; Vankova, A.; Stafilov, T. Phenolics and mineral content in bilberry and bog bilberry from Macedonia. Int. J. Food Prop. 2017, 20, S863–S883. [Google Scholar] [CrossRef]
- Klavins, L.; Maaga, I.; Bertins, M.; Hykkerud, A.L.; Karppinen, K.; Bobinas, C.; Salo, H.M.; Nguyen, N.; Salminen, H.; Stankevica, K.; et al. Trace element concentration and stable isotope ratio analysis in blueberries and bilberries: A tool for quality and authenticity control. Foods 2021, 10, 567. [Google Scholar] [CrossRef] [PubMed]
- Asgary, S.; Rafieian-Kopaei, M.; Sahebkar, A.; Shamsi, F.; Goli-Malekabadi, N. Anti-Hyperglycemic and Anti-Hyperlipidemic Effects of Vaccinium myrtillus Fruit in Experimentally Induced Diabetes (Antidiabetic Effect of Vaccinium myrtillus Fruit). J. Sci. Food Agric. 2016, 96, 764–768. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Laskowski, P.; Oszmiański, J. Evaluation of Sour Cherry (Prunus cerasus L.) Fruits for Their Polyphenol Content, Antioxidant Properties, and Nutritional Components. J. Agric. Food Chem. 2014, 62, 12332–12345. [Google Scholar] [CrossRef] [PubMed]
- Bljajic, K.; Petlevski, R.; Vujic, L.; Cacic, A.; Šoštarić, N.; Jablan, J.; Saraiva De Carvalho, I.; Zovko Končić, M. Chemical Composition, Antioxidant and α-Glucosidase-Inhibiting Activities of the Aqueous and Hydroethanolic Extracts of Vaccinium myrtillus Leaves. Molecules 2017, 22, 703. [Google Scholar] [CrossRef]
- Stefanescu (Braic), R.; Vari, C.; Imre, S.; Huţanu, A.; Fogarasi, E.; Todea, T.; Groşan, A.; Eşianu, S.; Laczkó-Zöld, E.; Dogaru, M. Vaccinium Extracts as Modulators in Experimental Type 1 Diabetes. J. Med. Food 2018, 21, 1106–1112. [Google Scholar] [CrossRef]
- Varut, R.M.; Gîrd, C.E.; Rotaru, L.T.; Varut, M.C.; Pisoschi, C.G. Evaluation of Polyphenol and Flavonoid Profiles and the Antioxidant Effect of Carduus Acanthoides Hydroalcoholic Extract Compared with Vaccinium myrtillus in an Animal Model of Diabetes Mellitus. Pharm. Chem. J. 2018, 51, 1088–1095. [Google Scholar] [CrossRef]
- do Rosario, V.A.; Chang, C.; Spencer, J.; Alahakone, T.; Roodenrys, S.; Francois, M.; Weston-Green, K.; Hölzel, N.; Nichols, D.S.; Kent, K.; et al. Anthocyanins Attenuate Vascular and Inflammatory Responses to a High Fat High Energy Meal Challenge in Overweight Older Adults: A Cross-Over, Randomized, Double-Blind Clinical Trial. Clin. Nutr. 2021, 40, 879–889. [Google Scholar] [CrossRef]
- Grohmann, T.; Walker, A.W.; Russell, W.R.; Hoggard, N.; Zhang, X.; Horgan, G.; de Roos, B. A grape seed and bilberry extract reduces blood pressure in individuals at risk of developing type 2 diabetes: The PRECISE study, a double-blind placebo-controlled cross-over intervention study. Front. Nutr. 2023, 10, 1139880. [Google Scholar] [CrossRef] [PubMed]
- Grussu, D.; Stewart, D.; McDougall, G.J. Berry Polyphenols Inhibit α-Amylase in Vitro: Identifying Active Components in Rowanberry and Raspberry. J. Agric. Food Chem. 2011, 59, 2324–2331. [Google Scholar] [CrossRef]
- Chen, K.; Wei, X.; Zhang, J.; Kortesniemi, M.; Zhang, Y.; Yang, B. Effect of Acylated and Nonacylated Anthocyanins on Urine Metabolic Profile during the Development of Type 2 Diabetes in Zucker Diabetic Fatty Rats. J. Agric. Food Chem. 2022, 70, 15143–15156. [Google Scholar] [CrossRef] [PubMed]
- Avramia, I.; Amariei, S. Formulation of Fast Dissolving β-Glucan/Bilberry Juice Films for Packaging Dry Powdered Pharmaceuticals for Diabetes. Plants 2022, 11, 2040. [Google Scholar] [CrossRef]
- Diaz-Garcia, L.; Schlautman, B.; Covarrubias-Pazaran, G.; Maule, A.; Johnson-Cicalese, J.; Grygleski, E.; Vorsa, N.; Zalapa, J. Massive phenotyping of multiple cranberry populations reveals novel QTLs for fruit anthocyanin content and other important chemical traits. Mol. Genet. Genom. 2018, 293, 1379–1392. [Google Scholar] [CrossRef]
- Bolivar-Medina, J.L.; Villouta, C.; Workmaster, B.A.; Atucha, A. Floral meristem development in cranberry apical buds during winter rest and its implication on yield prediction. J. Am. Soc. Hortic. Sci. 2019, 144, 314–320. [Google Scholar] [CrossRef]
- Feghali, K.; Feldman, M.; La, V.D.; Santos, J.; Grenier, D. Cranberry proanthocyanidins: Natural weapons against periodontal diseases. J. Agric. Food Chem. 2012, 60, 5728–5735. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Bonilla, L.; Williams, K.A.; Rodríguez Bonilla, F.; Matusinec, D.; Maule, A.; Coe, K.; Wiesman, E.; Diaz-Garcia, L.; Zalapa, J. The genetic diversity of cranberry crop wild relatives, Vaccinium macrocarpon Aiton and V. oxycoccos L., in the US, with special emphasis on national forests. Plants 2020, 9, 1446. [Google Scholar] [CrossRef] [PubMed]
- Cesoniene, L.; Daubaras, R. Phytochemical Composition of the Large Cranberry (Vaccinium macrocarpon) and the Small Cranberry (Vaccinium oxycoccos); Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780124081178. [Google Scholar]
- Cermak, R.; Landgraf, S.; Wolffram, S. Quercetin Glucosides Inhibit Glucose Uptake into Brush-Border-Membrane Vesicles of Porcine Jejunum. Br. J. Nutr. 2004, 91, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Strobel, P.; Allard, C.; Perez-Acle, T.; Calderon, R.; Aldunate, R.; Leighton, F. Myricetin, Quercetin and Catechin-Gallate Inhibit Glucose Uptake in Isolated Rat Adipocytes. Biochem. J. 2005, 386, 471–478. [Google Scholar] [CrossRef]
- Rocha, D.M.U.P.; Caldas, A.P.S.; Da Silva, B.P.; Hermsdorff, H.H.M.; Alfenas, R.D.C.G. Effects of Blueberry and Cranberry Consumption on Type 2 Diabetes Glycemic Control: A Systematic Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1816–1828. [Google Scholar] [CrossRef] [PubMed]
- Son, D.H.; Lee, H.S.; Lee, Y.J.; Lee, J.H.; Han, J.H. Comparison of triglyceride-glucose index and HOMA-IR for predicting prevalence and incidence of metabolic syndrome. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, K.M.; Shirinpour, E.; Shirazi, A.M.; Nikniaz, Z. Effect of cranberry supplementation on liver enzymes and cardiometabolic risk factors in patients with NAFLD: A randomized clinical trial. BMC Complement. Med. Ther. 2021, 21, 283. [Google Scholar]
- Hormoznejad, R.; Mansoori, A.; Hosseini, S.A.; Zilaee, M.; Asadi, M.; Fathi, M.; Kiany, F. Effects of cranberry consumption on features of the metabolic syndrome: A systematic review and meta-analysis of randomized control trials. Nutr. Food Sci. 2020, 51, 1006–1016. [Google Scholar] [CrossRef]
- Faheem, S.A.; Saeed, N.M.; El-Naga, R.N.; Ayoub, I.M.; Azab, S.S. Hepatoprotective Effect of Cranberry Nutraceutical Extract in Non-alcoholic Fatty Liver Model in Rats: Impact on Insulin Resistance and Nrf-2 Expression. Front. Pharmacol. 2020, 11, 218. [Google Scholar] [CrossRef]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Khanal, R.C.; Rogers, T.J.; Wilkes, S.E.; Howard, L.R.; Prior, R.L. Effects of dietary consumption of cranberry powder on metabolic parameters in growing rats fed high fructose diets. Food Funct. 2010, 1, 116–123. [Google Scholar] [CrossRef]
- Paquette, M.; Medina Larqué, A.S.; Weisnagel, S.J.; Desjardins, Y.; Marois, J.; Pilon, G.; Dudonné, S.; Marette, A.; Jacques, H. Strawberry and Cranberry Polyphenols Improve Insulin Sensitivity in Insulin-Resistant, Non-Diabetic Adults: A Parallel, Double-Blind, Controlled and Randomised Clinical Trial. Br. J. Nutr. 2017, 117, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, E.; Cameron, D.; Sobhan, R.; Wong, C.; Pontifex, M.G.; Tosi, N.; Mena, P.; Del Rio, D.; Sami, S.; Narbad, A.; et al. Chronic Consumption of Cranberries (Vaccinium macrocarpon) for 12 Weeks Improves Episodic Memory and Regional Brain Perfusion in Healthy Older Adults: A Randomised, Placebo-Controlled, Parallel-Groups Feasibility Study. Front. Nutr. 2022, 9, 849902. [Google Scholar] [CrossRef]
- Koshovyi, O.; Vlasova, I.; Laur, H.; Kravchenko, G.; Krasilnikova, O.; Granica, S.; Piwowarski, J.P.; Heinämäki, J.; Raal, A. Chemical Composition and Insulin-Resistance Activity of Arginine-Loaded American Cranberry (Vaccinium macrocarpon Aiton, Ericaceae) Leaf Extracts. Pharmaceutics 2023, 15, 2528. [Google Scholar] [CrossRef] [PubMed]
- Elhabal, S.F.; Al-Zuhairy, S.A.S.; El-Nabarawi, M.; Elrefai, M.F.M.; Shoela, M.S.; Hababeh, S.; Nelson, J.; Abdel Khalek, M.A.; Fady, M.; Elzohairy, N.A.; et al. Enhancing Photothermal Therapy for Antibiofilm Wound Healing: Insights from Graphene Oxide-Cranberry Nanosheet Loaded Hydrogel in vitro, in silico, and in vivo Evaluation. Int. J. Nanomed. 2024, 19, 12999–13027. [Google Scholar] [CrossRef] [PubMed]
- Rondanelli, M.; Mansueto, F.; Gasparri, C.; Solerte, S.B.; Misiano, P.; Perna, S. Supplementation with Highly Standardized Cranberry Extract Phytosome Achieved the Modulation of Urinary Tract Infection Episodes in Diabetic Postmenopausal Women Taking SGLT-2 Inhibitors: A RCT Study. Nutrients 2024, 16, 2113. [Google Scholar] [CrossRef] [PubMed]
- Girard, K.K.; Sinha, N.K. Cranberry, Blueberry, Currant, and Gooseberry. In Handbook of Fruits and Fruit Processing, 2nd ed.; Sinha, N.K., Sidhu, J.S., Barta, J., Wu, J.S.B., Cano, M.P., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; Chapter 24; pp. 399–417. [Google Scholar]
- Liu, W.; Mao, Y.; Schoenborn, J.; Wang, Z.; Tang, G.; Tang, X. Whole blueberry protects pancreatic beta-cells in diet-induced obese mouse. Nutr. Metab. 2019, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Keirsey, K.I.; Kirkland, R.; Grunewald, Z.I.; Fischer, J.G.; de La Serre, C.B. Blueberry Supplementation Influences the Gut Microbiota, Inflammation, and Insulin Resistance in High-Fat-Diet-Fed Rats. J. Nutr. 2018, 148, 209–219. [Google Scholar] [CrossRef]
- Nunes, S.; Viana, S.D.; Preguiça, I. Blueberry Counteracts Prediabetes in a Hypercaloric Diet-Induced Rat Model and Rescues Hepatic Mitochondrial Bioenergetics. Nutrients 2021, 13, 4192. [Google Scholar] [CrossRef]
- Morissette, A.; Kropp, C.; Songpadith, J.P.; Junges Moreira, R.; Costa, J.; Mariné Casadó, R.; Pilon, G.; Varin, T.V.; Dudonné, S.; Boutekrabt, L.; et al. Blueberry proanthocyanidins and anthocyanins improve metabolic health through a gut microbiota-dependent mechanism in diet-induced obese mice. Am. J. Physiology. Endocrinol. Metab. 2020, 318, E965–E980. [Google Scholar] [CrossRef]
- Mykkänen, O.T.; Huotari, A.; Herzig, K.H.; Dunlop, T.W.; Mykkänen, H.; Kirjavainen, P.V. Wild blueberries (Vaccinium myrtillus) alleviate inflammation and hypertension associated with developing obesity in mice fed with a high-fat diet. PLoS ONE 2014, 9, e114790. [Google Scholar] [CrossRef]
- Vendrame, S.; Zhao, A.; Merrow, T.; Klimis-Zacas, D. The effects of wild blueberry consumption on plasma markers and gene expression related to glucose metabolism in the obese Zucker rat. J. Med. Food 2015, 18, 619–624. [Google Scholar] [CrossRef]
- Seymour, E.M.; Tanone, I.I.; Urcuyo-Llanes, D.E.; Lewis, S.K.; Kirakosyan, A.; Kondoleon, M.G.; Kaufman, P.B.; Bolling, S.F. Blueberry intake alters skeletal muscle and adipose tissue peroxisome proliferator-activated receptor activity and reduces insulin resistance in obese rats. J. Med. Food 2011, 14, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Heyman, L.; Axling, U.; Blanco, N.; Sterner, O.; Holm, C.; Berger, K. Evaluation of Beneficial Metabolic Effects of Berries in High-Fat Fed C57BL/6J Mice. J. Nutr. Metab. 2014, 2014, 403041. [Google Scholar] [CrossRef]
- Khanal, R.C.; Howard, L.R.; Wilkes, S.E.; Rogers, T.J.; Prior, R.L. Effect of dietary blueberry pomace on selected metabolic factors associated with high fructose feeding in growing Sprague-Dawley rats. J. Med. Food 2012, 15, 802–810. [Google Scholar] [CrossRef]
- Brader, L.; Overgaard, A.; Christensen, L.P.; Jeppesen, P.B.; Hermansen, K. Polyphenol-rich bilberry ameliorates total cholesterol and LDL-cholesterol when implemented in the diet of Zucker diabetic fatty rats. Rev. Diabet. Stud. RDS 2013, 10, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Rozanska, D.; Regulska-Ilow, B. The significance of anthocyanins in the prevention and treatment of type 2 diabetes. Adv. Clin. Exp. Med. 2018, 27, 135–142. [Google Scholar] [CrossRef]
- Wedick, N.M.; Pan, A.; Cassidy, A.; Rimm, E.B.; Sampson, L.; Rosner, B.; Willett, W.; Hu, F.B.; Sun, Q.; van Dam, R.M. Dietary flavonoid intakes and risk of type 2 diabetes in US men and women. Am. J. Clin. Nutr. 2012, 95, 925–933. [Google Scholar] [CrossRef]
- Halvorsen, R.E.; Elvestad, M.; Molin, M.; Aune, D. Fruit and vegetable consumption and the risk of type 2 diabetes: A systematic review and dose-response meta-analysis of prospective studies. BMJ Nutr. Prev. Health 2021, 4, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Muraki, I.; Imamura, F.; Manson, J.E.; Hu, F.B.; Willett, W.C.; van Dam, R.M.; Sun, Q. Fruit consumption and risk of type 2 diabetes: Results from three prospective longitudinal cohort studies. BMJ 2013, 347, f5001. [Google Scholar] [CrossRef] [PubMed]
- Stull, A.J.; Cash, K.C.; Johnson, W.D.; Champagne, C.M.; Cefalu, W.T. Bioactives in blueberries improve insulin sensitivity in obese, insulin-resistant men and women. J. Nutr. 2010, 140, 1764–1768. [Google Scholar] [CrossRef] [PubMed]
- Curtis, P.J.; van der Velpen, V.; Berends, L.; Jennings, A.; Feelisch, M.; Umpleby, A.M.; Evans, M.; Fernandez, B.O.; Meiss, M.S.; Minnion, M.; et al. Blueberries improve biomarkers of cardiometabolic function in participants with metabolic syndrome-results from a 6-month, double-blind, randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Stull, A.J.; Cash, K.C.; Champagne, C.M.; Gupta, A.K.; Boston, R.; Beyl, R.A.; Johnson, W.D.; Cefalu, W.T. Blueberries improve endothelial function, but not blood pressure, in adults with metabolic syndrome: A randomized, double-blind, placebo-controlled clinical trial. Nutrients 2015, 7, 4107–4123. [Google Scholar] [CrossRef]
- Krikorian, R.; Skelton, M.R.; Summer, S.S.; Shidler, M.D.; Sullivan, P.G. Blueberry supplementation in midlife for dementia risk reduction. Nutrients 2022, 14, 1619. [Google Scholar] [CrossRef]
- Curtis, P.J.; Berends, L.; van der Velpen, V.; Jennings, A.; Haag, L.; Chandra, P.; Kay, C.D.; Rimm, E.B.; Cassidy, A. Blueberry anthocyanin intake attenuates the postprandial cardiometabolic effect of an energy-dense food challenge: Results from a double blind, randomized controlled trial in metabolic syndrome participants. Clin. Nutr. 2022, 41, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Mane, C.; Loonis, M.; Juhel, C.; Dufour, C.; Malien-Aubert, C. Food Grade Lingonberry Extract: Polyphenolic Composition and In vivo Protective Effect against Oxidative Stress. J. Agric. Food Chem. 2011, 59, 3330–3339. [Google Scholar] [CrossRef] [PubMed]
- Drózdz, P.; Šežiene, V.; Wójcik, J.; Pyrzynska, K. Evaluation of Bioactive Compounds, Minerals and Antioxidant Activity of Lingonberry (Vaccinium vitis-idaea L.) Fruits. Molecules 2017, 23, 53. [Google Scholar] [CrossRef]
- Kowalska, K. Lingonberry (Vaccinium vitis-idaea L.) fruit as a source of bioactive compounds with health-promoting effects-A review. Int. J. Mol. Sci. 2021, 22, 5126. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Li, T.; Li, Y.; Qian, H.; Zhang, H.; Rao, Z.; Wang, L. Vaccinium bracteatum Thunb. as a promising resource of bioactive compounds with health benefits: An updated review. Food Chem. 2021, 356, 129738. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, H.; Yang, H.; Wang, T.; Chang, Y.; Nie, C.; Nie, S.; Fu, Y. Lingonberry (Vaccinium vitis-idaea L.) fruits: Potential characterization of flavor and functional profiles during ripening based on UHPLC-QqQ-MS/MS. J. Food Compos. Anal. 2024, 130, 106143. [Google Scholar] [CrossRef]
- Esposito, D.; Overall, J.; Grace, M.H.; Komarnytsky, S.; Lila, M.A. Alaskan berry extracts promote dermal wound repair throughmodulation of bioenergetics and integrin signaling. Front. Pharmacol. 2019, 10, 1058. [Google Scholar] [CrossRef] [PubMed]
- Ryyti, R.; Hämäläinen, M.; Peltola, R.; Moilanen, E. Beneficial effects of lingonberry (Vaccinium vitis-idaea L.) supplementation on metabolic and inflammatory adverse effects induced by high-fat diet in a mouse model of obesity. PLoS ONE 2020, 15, e0232605.7. [Google Scholar] [CrossRef]
- Marungruang, N.; Kovalenko, T.; Osadchenko, I.; Voss, U.; Huang, F.; Burleigh, S.; Ushakova, G.; Skibo, G.; Nyman, M.; Prykhodko, O.; et al. Lingonberries and their two separated fractions differently alter the gut microbiota, improve metabolic functions, reduce gut inflammatory properties, and improve brain function in ApoE−/− mice fed high-fat diet. Nutr. Neurosci. 2020, 23, 600–612. [Google Scholar] [CrossRef]
- Ho, G.T.T.; Nguyen, T.K.Y.; Kase, E.T.; Tadesse, M.; Barsett, H.; Wangensteen, H. Enhanced Glucose Uptake in Human Liver Cells and Inhibition of Carbohydrate Hydrolyzing Enzymes by Nordic Berry Extracts. Molecules 2017, 22, 1806. [Google Scholar] [CrossRef]
- Torronen, R.; Kolehmainen, M.; Sarkkinen, E.; Mykkanen, H.; Niskanen, L. Postprandial glucose, insulin, and free fatty acid responses to sucrose consumed with blackcurrants and lingonberries in healthy women. Am. J. Clin. Nutr. 2012, 96, 527–533. [Google Scholar] [CrossRef]
- Harbilas, D.; Martineau, L.C.; Harris, C.S.; Adeyiwola-Spoor, D.C.; Saleem, A.; Lambert, J.; Caves, D.; Johns, T.; Prentki, M.; Cuerrier, A.; et al. Evaluation of the antidiabetic potential of selected medicinal plant extracts from the Canadian boreal forest used to treat symptoms of diabetes: Part II. Can. J. Physiol. Pharmacol. 2009, 87, 479–492. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Chang, Q.; Zhou, Z.; Han, R.; Liang, Z. Antioxidant and antidiabetic activity of proanthocyanidins from Fagopyrum dibotrys. Molecules 2021, 26, 2417. [Google Scholar] [CrossRef]
- Eraky, S.M.; Ramadan, N.M.; Abo El-Magd, N.F. Antidiabetic effects of quercetin and liraglutide combination through modulation of TXNIP/IRS-1/PI3K pathway. Cell Biochem. Funct. 2022, 40, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Oyenihi, O.R.; Oyenihi, A.B.; Adeyanju, A.A.; Oguntibeju, O.O. Antidiabetic Effects of Resveratrol: The Way Forward in Its Clinical Utility. J. Diabetes Res. 2016, 2016, 9737483. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Chemposov, V.V.; Chirikova, N.K. Polymeric Compounds of LingonberryWaste: Characterization of Antioxidant and Hypolipidemic Polysaccharides and Polyphenol-Polysaccharide Conjugates from Vaccinium vitis-idaea Press Cake. Foods 2022, 11, 2801. [Google Scholar] [CrossRef] [PubMed]
- Muceniece, R.; Klavins, L.; Kviesis, J.; Jekabsons, K.; Rembergs, R.; Saleniece, K.; Dzirkale, Z.; Saulite, L.; Riekstina, U.; Klavins, M. Antioxidative, hypoglycaemic and hepatoprotective properties of five Vaccinium spp. berry pomace extracts. J. Berry Res. 2019, 9, 267–282. [Google Scholar] [CrossRef]
- Ryyti, R.; Hamalainen, M.; Tolonen, T.; Maki, M.; Jaakkola, M.; Peltola, R.; Moilanen, E. Lingonberry (Vaccinium vitis-idaea L.) Skin Extract PreventsWeight Gain and Hyperglycemia in High-Fat Diet-Induced Model of Obesity in Mice. Nutrients 2024, 16, 2107. [Google Scholar] [CrossRef] [PubMed]
- Bone, K.; Mills, S.C. Principles and Practice of Phytotherapy: Modern Herbal Medicine, 2nd ed.; Churchill Livingstone: Edinburgh, UK, 2013. [Google Scholar]
- Allen, D.; Bilz, M.; Leaman, D.J.; Miller, R.M.; Timoshyna, A.; Window, J. European Red List of Medicinal Plants; Publications Office of the European Union: Luxembourg, 2014. [Google Scholar]
- Stefanescu, B.E.; Szabo, K.; Mocan, A.; Crisan, G. Phenolic compounds from five Ericaceae species leaves and their related bioavailability and health benefits. Molecules 2019, 24, 2046. [Google Scholar] [CrossRef] [PubMed]
- Vranješ, M.; Popovi’c, B.M.; Štajner, D.; Ivetic, V.; Mandic, A.; Vranješ, D. Effects of bearberry, parsley and corn silk extracts on diuresis, electrolytes composition, antioxidant capacity and histopathological features in mice kidneys. J. Funct. Foods 2016, 21, 272–282. [Google Scholar] [CrossRef]
- Wrona, M.; Blasco, S.; Becerril, R.; Nerin, C.; Sales, E.; Asensio, E. Antioxidant and antimicrobial markers by UPLC®–ESIQ-TOF-MSE of a new multilayer active packaging based on Arctostaphylos uva-ursi. Talanta 2019, 196, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Arbutin as a skin depigmenting agent with antimelanogenic and antioxidant properties. Antioxidants 2021, 10, 1129. [Google Scholar] [CrossRef]
- Lin, Y.C. Studies on the hypoglycemic activity of the medicinal herbs. Formos. Med. Assoc. 1964, 63, 400–404. [Google Scholar]
- Kokate, C.K. A Text Book of Pharmacognocy, 35th ed.; Nirali Prakashan: New Delhi, India, 2006; pp. 218–219, 252–253, 268–269, 309–310. [Google Scholar]
- Kravchenko, G.B.; Krasilnikova, O.A.; Mazen, M. The study of the hypoglycemic action of extracts from bearberry leaves under experimental insulin resistance in rats. Clin. Pharm. 2020, 24, 40–46. [Google Scholar] [CrossRef]
- Kravchenko, G.; Matar, M.; Krasilnikova, O. Screening of bearberry leaves extracts hypoglycemic effect and study of acute toxicity. Ukr. Biopharm. J. 2018, 2, 13–16. [Google Scholar] [CrossRef]
- Chaika, N.; Mazen, M.; Koshovyi, O.; Kravchenko, G.; Goryacha, O.; Kireyev, I.; Kovalenko, S.; Darmograi, R. Research in phytochemical composition and hypoglycemic activity screening of the dry extracts from bearberry leaves. Sci. Rise Pharm. Sci. 2021, 3, 42–50. [Google Scholar]
- Kravchenko, G.; Krasilnikova, O.; Raal, A.; Mazen, M.; Chaika, N.; Kireyev, I.; Grytsyk, A.; Koshovyi, O. Arctostaphylos uva-ursi L. leaves extract and its modified cysteine preparation for the management of insulin resistance: Chemical analysis and bioactivity. Nat. Prod. Bioprospect. 2022, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Mazen, M.; Kravchenko, G.; Krasilnikova, O. Study of hepatoprotective effect of bearberry leaves extract under insulin resistance in rats. EUREKA Health Sci. 2021, 6, 48–53. [Google Scholar] [CrossRef]
- Celikel, G.; Demirsoy, L.; Demirsoy, H. The strawberry tree (Arbutus Unedo L.) selection in Turkey. Sci. Hortic. 2008, 118, 115–119. [Google Scholar] [CrossRef]
- Bachar, M.; Zidane, L.; Rochdi, A. Ethno-medicinal and traditional phytotherapy of plants used in bouhachem natural regional park “Rif of Morocco”—Case of tazroute district. J. Mater. Environ. Sci. 2018, 7, 4175–4204. [Google Scholar]
- Rhattas, M.; Douira, A.; Zidane, L. ‘Etude ethnobotanique des plantes m’edicinales dans le Parc National de Talassemtane (Rif Occidental du Maroc). J. Appl. Biosci. 2016, 97, 9187–9211. [Google Scholar] [CrossRef]
- Mrabti, H.N.; Jaradat, N.; Kachmar, M.R. Integrative herbal treatments of diabetes in Beni Mellal region of Morocco. J. Integr. Med. 2019, 17, 93–99. [Google Scholar] [CrossRef]
- Zlabur, J.S.; Bogdanovic, S.; Voca, S.; Babojelic, M.S. Biological potential of fruit and leaves of strawberry tree (Arbutus unedo L.) from Croatia. Molecules 2020, 25, 2521. [Google Scholar] [CrossRef]
- Erkekoglou, I.; Nenadis, N.; Samara, E.; Mantzouridou, F.T. Functional teas from the leaves of Arbutus unedo: Phenolic content, antioxidant activity, and detection of ePcient radical scavengers. Plant Foods Hum. Nutr. 2017, 72, 176–183. [Google Scholar] [CrossRef] [PubMed]
- El Haouari, M.; Mekh, H. Anti-platelet aggregation effects of extracts from Arbutus unedo leaves. Plant Sci. Today 2017, 4, 68–74. [Google Scholar] [CrossRef]
- Bnouham, M.; Merhfour, F.Z.; Ziyyat, A.; Aziz, M.; Legssyer, A.; Mekh, H. Antidiabetic effect of some medicinal plants of Oriental Morocco in neonatal non-insulin-dependent diabetes mellitus rats. Hum. Exp. Toxicol. 2010, 29, 865–871. [Google Scholar] [CrossRef]
- Mrabti, H.N.; Jaradat, N.; Fichtali, I. Separation, identication, and antidiabetic activity of catechin isolated from Arbutus unedo L. Plant roots. Plants 2018, 72, 31. [Google Scholar] [CrossRef]
- Delgado-Pelayo, R.; Gallardo-Guerrero, L.; Hornero-Mendez, D. Carotenoid composition of strawberry tree (Arbutus unedo L.) fruits. Food Chem. 2016, 199, 165–175. [Google Scholar] [CrossRef]
- Lopez, C.J.; Caleja, C.; Prieto, M.A.; Barreiro, M.F.; Barros, L.; Ferreira, I. Optimization and comparison of heat and ultrasound assisted extraction techniques to obtain anthocyanin compounds from Arbutus unedo L. Fruits. Food Chem. 2018, 264, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Bnouham, M.; Merhfour, F.Z.; Legssyer, A.; Mekh, H.; Maallem, S.; Ziyyat, A. Antihyperglycemic activity of Arbutus unedo, Ammoides pusilla and Thymelaea hirsuta. Die Pharm. 2007, 62, 630–632. [Google Scholar]
- Mrabti, H.N.; Sayah, K.; Jaradat, N. Antidiabetic and protective effects of the aqueous extract of Arbutus unedo L. in streptozotocin-nicotinamide-induced diabetic mice. J. Complement. Integr. Med. 2018, 15, 3. [Google Scholar] [CrossRef]
- Mrabti, H.N.; El Abbes Faouzi, M.; Mayuk, F.M. Arbutus unedo L., (Ericaceae) inhibits intestinal glucose absorption and improves glucose tolerance in rodents. J. Ethnopharmacol. 2019, 235, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Touzani, S.; Al-Waili, N.; Imtara, H.; Aboulghazi, A.; Hamma, N.; Falcão, S.; Vilas-Boas, M.; El Arabi, I.; Al-Waili, W.; Lyoussi, B. Arbutus unedo honey and propolis ameliorate acute kidney injury, acute liver injury, and proteinuria via hypoglycemic and antioxidant activity in streptozotocin-treated rats. Cell Physiol. Biochem. 2022, 56, 66–81. [Google Scholar]
- Al-Mijalli, S.H.; Mrabti, H.N.; Ouassou, H.; Flouchi, R.; Abdallah, E.M.; Sheikh, R.A.; Alshahrani, M.M.; Awadh, A.A.A.; Harhar, H.; Omari, N.E.; et al. Chemical Composition, Antioxidant, Anti-Diabetic, Anti-Acetylcholinesterase, Anti-Inflammatory, and Antimicrobial Properties of Arbutus unedo L. and Laurus nobilis L. Essential Oils. Life 2022, 12, 1876. [Google Scholar] [CrossRef] [PubMed]
- Koskela, A.K.; Anttonen, M.J.; Soininen, T.H.; Saviranta, N.M.; Auriola, S.; Julkunen-Tiitto, R.; Karjalainen, R.O. Variation in the anthocyanin concentration of wild populations of crowberries (Empetrum nigrum L subsp. hermaphroditum). J. Agric. Food Chem. 2010, 58, 12286–12291. [Google Scholar]
- Vaisanen, M.; Martz, F.; Kaarlejarvi, E.; Julkunen-Tiitto, R.; Stark, S. Phenolic responses of mountain crowberry (Empetrum nigrum ssp. hermaphroditum) to global climate change are compound specific and depend on grazing by reindeer (Rangifer tarandus). J. Chem. Ecol. 2013, 39, 1390–1399. [Google Scholar]
- Hyun, T.K.; Kim, H.-C.; Ko, Y.-J.; Kim, J.-S. Antioxidant, α-glucosidase inhibitory, and anti-inflammatory effects of aerial parts extract from Korean crowberry (Empetrum nigrum var. japonicum). Saudi J. Biol. Sci. 2016, 23, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Hyun, T.K.; Ra, J.H.; Han, S.H.; Kim, J.S. Antioxidant, antimicrobial, and antidiabetic activities of crowberry fruits. Indian J. Pharm. Sci. 2018, 80, 489–495. [Google Scholar] [CrossRef]
- Kashtoh, H.; Baek, K.-H. Recent Updates on Phytoconstituent Alpha-Glucosidase Inhibitors: An Approach towards the Treatment of Type Two Diabetes. Plants 2022, 11, 2722. [Google Scholar] [CrossRef]
- Törrönen, R.; McDougall, G.J.; Dobson, G.; Stewart, D.; Hellström, J.; Mattila, P.; Pihlava, J.M.; Koskela, A.; Karjalainen, R. Fortification of blackcurrant juice with crowberry: Impact on polyphenol composition, urinary phenolic metabolites, and postprandial glycemic response in healthy subjects. J. Funt. Foods 2012, 4, 746–756. [Google Scholar] [CrossRef]
- Vengrytė, M.; Raudonė, L. Phytochemical Profiling and Biological Activities of Rhododendron Subsect. Ledum: Discovering the Medicinal Potential of Labrador Tea Species in the Northern Hemisphere. Plants 2024, 13, 901. [Google Scholar] [CrossRef]
- Ni, H.; Hayes, H.E.; Stead, D.; Raikos, V. Incorporating salal berry (Gaultheria shallon) and blackcurrant (Ribes nigrum) pomace in yogurt for the development of a beverage with antidiabetic properties. Heliyon 2018, 4, e00875. [Google Scholar] [CrossRef]
- Liu, S.; Sun, L.; Zhang, P.; Niu, C. Recent Advances in Grayanane Diterpenes: Isolation, Structural Diversity, and Bioactivities from Ericaceae Family (2018–2024). Molecules 2024, 29, 1649. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.-C.; et al. Scientific Opinion on the Risks for Human Health Related to the Presence of Grayanotoxins in Certain Honey. EFSA J. 2023, 21, 7866. [Google Scholar]
- Doudach, L.; Mrabti, H.N.; Al-Mijalli, S.H.; Kachmar, M.R.; Benrahou, K.; Assaggaf, H.; Qasem, A.; Abdallah, E.M.; Rajab, B.S.; Harraqui, K.; et al. Phytochemical, Antidiabetic, Antioxidant, Antibacterial, Acute and Sub-Chronic Toxicity of Moroccan Arbutus unedo Leaves. J. Pharmacopunct. 2023, 26, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Cosme, F.; Aires, A.; Pinto, T.; Oliveira, I.; Vilela, A.; Gonçalves, B. A Comprehensive Review of Bioactive Tannins in Foods and Beverages: Functional Properties, Health Benefits, and Sensory Qualities. Molecules 2025, 30, 800. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Species | Content Type | Identified Derivatives | Class |
|---|---|---|---|
| V. myrtillus (bilberry)  | Cyanidin Delphinidin Malvidin Petunidin Myricetin Quercetin Others | cyanidin 3-galactoside, cyanidin 3-glucoside, cyanidin 3-arabinoside, peonidin 3-arabinoside, cyanidin 3-xyloside, cyanidin 5-glucoside, cyanidin 3,5-diglucoside delphinidin 3-galactoside, delphinidin 3-arabinoside, delphinidin 3-glucoside, malvidin 3-galactoside, malvidin 3-arabinoside, malvidin 3-glucoside petunidin 3-arabinoside, petunidin 3-acetylglucoside, petunidin 3-glucoside, myricetin 3-glucoside, myricetin 3-arabinoside, myricetin3-rhamnoside, myricetin-3-xyloside, myricetin 3-galactoside quercetin 3-arabinoside, quercetin 3-rhamnoside, quercetin 3-galactoside, quercetin 3-glucoside, quercetin 3-rutinoside, apigenin, chrysoeriol, isorhamnetin, laricitrin, syringetin, luteolin, kaempferol [54] | ANT ANT ANT ANT ANT F F |
V. macrocarpon (cranberry) | Cyanidin Delphinidin Malvidin Peonidin Pelargonidin Quercetin Kaempferol | cyanidin-3-glucoside, cyanidin-3-galactoside, cyanidin-3-arabinoside delphinidin-3-arabinoside malvidin-3-galactoside, malvidin-3-arabinoside peonidin-3-glucoside, peonidin-3-galactoside, peonidin-3-arabinoside pelargonidin-3-galactoside, pelargonidin-3-arabinoside quercetin-3-galactoside, quercetin-3-arabinoside, quercetin-3-rhamnoside kaempferol-3-glucoside [44] | ANT ANT ANT ANT ANT F F |
| Vaccinium spp. (blueberry)  | Cyanidin Delphinidin Malvidin Petunidin Myricetin Quercetin | cyanidin 3-galactoside, cyanidin 3-glucoside, cyanidin 3-arabinoside delphinidin 3-arabinoside, delphinidin 3-acetylglucoside malvidin 3-galactoside, malvidin 3-glucoside, malvidin 3-arabinoside, malvidin 3-acetylglucoside petunidin 3-galactoside, petunidin 3-glucoside, petunidin 3-arabinoside myricetin 3-galactoside, myricetin 3-glucoside, myricetin 3-rhamnoside quercetin 3-galactoside, quercetin 3-glucoside, quercetin 3-glucosylxyloside, quercetin 3-rutinoside, quercetin 3-xyloside [55] | ANT ANT ANT ANT F F |
| V. vitis idaea (lingonberry)  | Cyanidin Delphinidin Petunidin Peonidin Myricetin Quercetin Kaempferol Isorhamnetin | cyanidin 3-glucoside, cyanidin 3-arabinoside delphinidin 3-glucoside, delphinidin 3-arabinoside, delphinidin 3-galactoside, petunidin 3-galactoside, petunidin 2-glucoside, peonidin 3-arabinoside, peonidin 3-glucoside, peonidin 3-galactoside myricetin 3-glucoside quercetin 3-glucoside, quercetin 3-galactoside, quercetin 3-arabinoside, quercetin 3-xyloside kaempferol 3-rhamnoside, kaempferol 3-glucoside isorhamnetin 3-galactoside, isorhamnetin 3-glucoside [56] | ANT ANT ANT ANT F F F F |
Arctostaphylos uva-ursi L. (Bearberry) | Quercetin Kaempferol Myricetin | Isoquercetin, quercitin-3-gentiobioside, hyperoside, avicularin, rutin Kaempferol-pentoside/hexoside, Myricetin-pentoside/hexoside [57] | F F F |
Vaccinium arctostaphylos (Caucasian whortleberry) | Delphinidin Petunidin Malvidin Cyanidin | delphinidin 3-galactoside, delphinidin 3-arabinoside, delphinidin 3-glucoside petunidin 3-arabinoside, petunidin 3-glucosidemalvidin 3-galactoside, malvidin 3-arabinoside, malvidin 3-glucoside cyanidin-3-O-xyloside [58] | ANT ANT ANT ANT |
Gaultheria trichophylla (Himalayan snowberry) | Quercetin Kaempferol Catechin Epicatechin | Quercetin 3- glycoside, Quercetin 3-O-galactoside, Quercetin 3-rhamnoside, Kaempferol 7- glucoside, Kaempferol 3-glucoside [59] | F |
Rhododendron arboreum (Himalayan rhododendron) | Ursolic acid derivatives Quercetin | quercetin-3-O-galactoside [60] | F |
Rhododendron groenlandicum (Labrador tea) | Catechin Epicatechin [61] | F | |
Arbutus unedo | Myricetin Quercetin Kaempferol | Myricetin 3-glucoside, Myricetin 3-pentosideQuercetin 3- glycoside, Quercetin 3-O-galactoside, Quercetin 3-rhamnoside, Kaempferol 7- glucoside, Kaempferol 3-glucoside [62] | F F F |
Empetrum nigrum (Crowberry) | Cyanidin Delphinidin Petunidin Malvidin Quercetin Kaempferol | cyanidin 3-galactoside, cyanidin 3-glucoside, cyanidin 3-arabinoside delphinidin 3-galactoside, delphinidin 3-arabinoside, delphinidin 3-glucoside petunidin 3-arabinoside, petunidin 3-glucosidemalvidin 3-galactoside, malvidin 3-arabinoside, malvidin 3-glucoside quercetin-3-glucoside, quercetin-3-arabinose, quercetin-3-xyloside, quercetin-3-galactoside, quercetin-3-rhamnoside glucoside and galactoside forms [63] | ANT ANT ANT ANT F F |
| Compound | IUPAC Name | Molecular Formula | 2D Structure |
|---|---|---|---|
| Quercetin [64] | 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one | C15H10O7 |  |
| Kaempferol [65] | 3,5,7-trihydroxy-2-(4-hydroxyphenyl)chromen-4-one | C15H10O6 |  |
| Myricetin [66] | 3,5,7-trihydroxy-2-(3,4,5-trihydroxyphenyl)chromen-4-one | C15H10O8 | 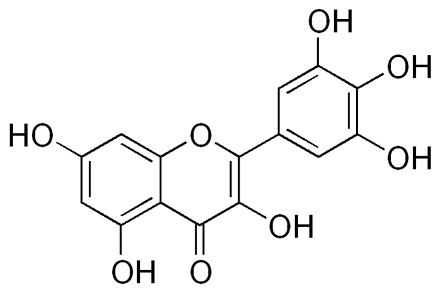 |
| Cyanidin [67] | 2-(3,4-dihydroxyphenyl)chromenylium-3,5,7-triol | C15H11O6 | 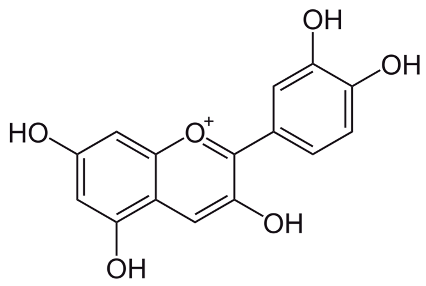 |
| Delphinidin [68] | 2-(3,4,5-trihydroxyphenyl)chromenylium-3,5,7-triol; | C15H11O7 | 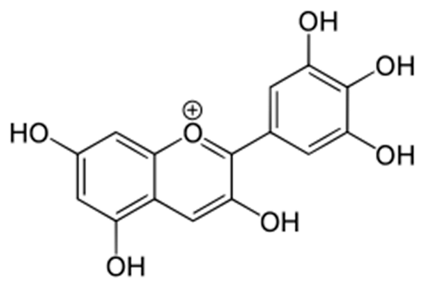 |
| Malvidin [69] | (2S,3R,4S,5S,6R)-2-[5,7-dihydroxy-2-(4-hydroxy-3,5-dimethoxyphenyl)chromenylium-3-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol | C23H25O12 | 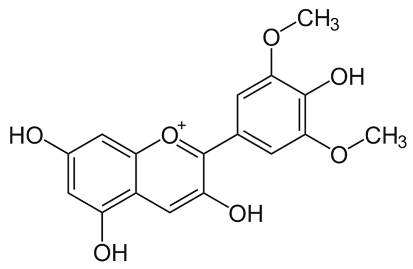 |
| Petunidin [70] | C22H23O12 | (2S,3R,4S,5S,6R)-2-[2-(3,4-dihydroxy-5-methoxyphenyl)-5,7-dihydroxychromenylium-3-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol | 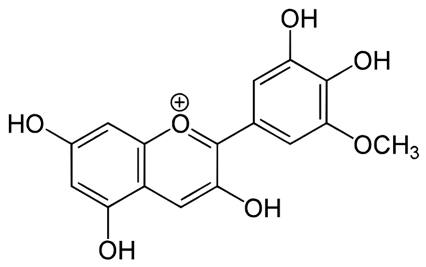 |
| Pelargonidin [71] | C15H11ClO5 | 2-(4-hydroxyphenyl)chromenylium-3,5,7-triol;chloride | 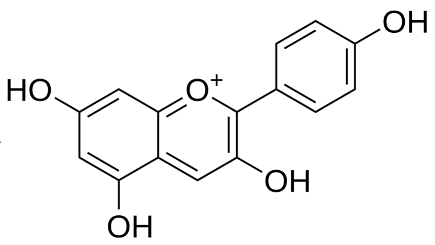 |
| Species | Variety | Total Phenolic Content | Reference |
|---|---|---|---|
| V. myrtillus | Wild bilberry | 492–563 mg GAE/100 g FW | [72] |
| V. macrocarpon | “Early Black” cultivated | ~441 mg GAE/100 g FW | [73] |
| Vaccinium spp. | “Bluecrop” highbush blueberry | ~327 mg GAE/100 g FW | [72] |
| V. vitis-idaea | Wild lingonberry | 468–661 mg GAE/100 g FW | [74] |
| Arctostaphylos uva-ursi L. | Leaf extracts (wild populations) | 238.85–318.28 mg GAE/g DW | [75] |
| Vaccinium arctostaphylos | Wild fruit (Caucasian whortleberry) | 389–578 mg GAE/100 g FW | [76] |
| Gaultheria trichophylla | Wild fruit (Himalayan snowberry) | ~3.71 mg GAE/g FW (≈371 mg/100 g FW) | [77] |
| Rhododendron arboreum | Flower petals (Himalayan rhododendron) | ~4.89 mg GAE/g DW (best processing) | [78] |
| Rhododendron groenlandicum | Leaves (Labrador tea) | 20 g/100 g DW (leaf extract) | [79] |
| Arbutus unedo | Strawberry tree fruit | 16.78–25.86 mg GAE/g DW | [80] |
| Empetrum nigrum | Wild black crowberry (Canada) | ~454 mg GAE/100 g FW (fresh fruit) | [75] |
| Species | Main Compounds Class Responsible for Antioxidant Activity | ABTS | DPPH |
|---|---|---|---|
| V. myrtillus | Anthocyanins (anthocyanidin glycosides)—e.g., delphinidin and cyanidin derivatives | 60.9–106.0 µmol Trolox/g FW (ABTS radical cation scavenging) [81] | 216.5–376.8 µmol Trolox/g FW (DPPH radical scavenging [82] |
| V. macrocarpon | Proanthocyanidins (A-type PAC oligomers) | 189–264 µmol Trolox/g DW (ABTS, cultivar range) [83] | 214–320 µmol Trolox/g DW (DPPH, cultivar range) [84] |
| Vaccinium spp. | Anthocyanins (flavonoid pigments)—malvidin, petunidin, and cyanidin glycosides | ~259.9 µmol Trolox/g DW (ABTS in highbush cv. ‘Biloxi’) [85] | ~214.1 µmol Trolox/g DW (DPPH in ‘Biloxi’ fruit) [85] |
| V. vitis idaea | Anthocyanins (cyanidin glycosides) | ~74.3–104.0 µmol Trolox/g DW (TEAC/ABTS assay) [80] | Potent DPPH scavenging capacity (multiple radicals quenched; ORAC/DPPH assays confirm high activity) [86] |
| Arctostaphylos uva-ursi L. | Phenolic glycosides (arbutin) | 173.5–643.7 mg Trolox/g (ABTS•⁺ scavenging, water vs. EtOH extracts) [87] | 377.4–821.8 mg Trolox/g (DPPH scavenging, water vs. EtOH) [87] |
| Vaccinium arctostaphylos | Anthocyanins (delphinidin, petunidin, and malvidin glycosides) | ~19.5 µmol Trolox/g FW [76] | SC50 = 0.14 mg/mL (ethanol extract) [88,89] |
| Gaultheria trichophylla | Polyphenols (flavonoids and tannins) | 4.35 mM AAE/100 g FW [77] | 2.56 mM AAE/100 g FW [77] |
| Rhododendron arboreum | Anthocyanins and flavonoids (quercetin derivatives) | 21.25–31.87 mM AAE/100 g DW [78] | 22.59–36.61 mM AAE/100 g DW [78] |
| Rhododendron groenlandicum | Flavonol glycosides, catechins (quercetin, myricetin, and catechin) | - | Strong radical scavenging activity (ORAC assay confirms high potency) [90] |
| Arbutus unedo | Flavan-3-ols (catechin and gallocatechin tannins) | 74.3–104.0 µmol Trolox/g DW (antioxidant capacity, TEAC assay) [80] | DPPH radical scavenging EC50 in low mg/mL; activity strongly correlates with total phenolics [80] |
| Empetrum nigrum | Anthocyanins | 107 µmol Trolox/g [91] | ~90 mg Trolox/g [89,91] |
| Plant | Age | Number | Duration | Results | Reference |
|---|---|---|---|---|---|
| Blueberry | 27 ± 5 | 34 | 18 weeks | Lower C-reactive protein and blood glucose levels compared to the control group | [111] |
| 22–65 | 17 | / | Significant increase in pancreatic polypeptide(PP) concentrations in intervention group | [112] | |
| 51–75 | 58 | 8 weeks | Lower hemoglobin A1c, fructosamine, and triglycerides | [113] | |
| Bilberry | 30–65 | 105 | 90 days | Reduced 2 h blood postprandial glucose and homeostasis model assessment of insulin resistance (HOMA-IR) scores | [114] |
| 25–60 | 47 | 8 weeks | Significant increase in fasting serum hippuric acid in intervention group | [115] | |
| 55.8 ± 9.5 | 20 | 4 weeks | Tendency of improved glycemic control in intervention group | [116] | |
| Cranberry | 40–75 | 160 | 12 weeks | Reduced HbA1c, low-density lipoprotein-c, apolipoprotein A-1, apolipoprotein B in intervention group | [117] |
| 56–67 | 58 | 24 weeks | Decreased serum LDL cholesterol, triglycerides, apolipoprotein B, and apo C-III; increased HDL cholesterol | [118] | |
| 56 ± 6 | 25 | / | Lower postprandial increases in glucose at 2 and 4 h in the cranberry group, | [119] | |
| 25–65 | 56 | 8 weeks | Reduced circulating TGs, CRP, glucose, insulin resistance, and diastolic BP in intervention group | [120] | |
| Llingonberry | 25–69 | 20 | / | Improved postprandial glycemic profiles | [74] |
| Plant Part | Extract/Fraction | Model Applied | Effect/Mechanism of Action | Reference |
|---|---|---|---|---|
| Roots | Water | OGTT * IVGTT * | Antihyperglycemic effect Inhibition of jejunal glucose absorption | [216] |
| Roots | Water | OGTT n-str-induced diabetic rats | Hypoglycemic effect Potentiation of the insulin Activity Improved glucose peripheral consumption | [212] |
| Roots | Catechin | α-glucosidase assay | Antidiabetic effect through inhibition of α-glucosidase enzyme | [213] |
| Roots | Water | α–glucosidase and α-amylase assays | Regeneration of pancreatic C-cells | [217] |
| Roots bark | Water | OGTT | Improved oral glucose tolerance | [218] |
| Species | Experimental Model | Observed Activity | Main Compounds |
|---|---|---|---|
| Gaultheria hispidula (Creeping snowberry) [185] | C2C12 myotubes and 3T3-L1 adipocytes (cell assays) | Stimulated glucose uptake; promoted adipogenesis (~50% of rosiglitazone); antioxidant and neuroprotective activity | Phenolic glycosides (e.g., gaultherin and arbutin) and flavonoids |
| Rhododendron tomentosum (Marsh Labrador tea) [227] | C2C12, 3T3-L1 adipocytes, and antioxidant assays | Strong glucose uptake stimulation; PPARγ-like adipogenic activity; potent antioxidant; no insulin secretion | Flavonoids, catechins, and polyphenols |
| Gaultheria shallon (Salal berry) [228] | α-amylase, α-glucosidase, and DPP-IV enzyme assays | Inhibited α-glucosidase, α-amylase (>60%), DPP-IV (~56%); high antioxidant potential | Anthocyanins, procyanidins, and flavonoids |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popescu, M.; Radivojevic, K.; Trasca, D.-M.; Varut, R.M.; Enache, I.; Osman, A. Natural Antidiabetic Agents: Insights into Ericaceae-Derived Phenolics and Their Role in Metabolic and Oxidative Modulation in Diabetes. Pharmaceuticals 2025, 18, 682. https://doi.org/10.3390/ph18050682
Popescu M, Radivojevic K, Trasca D-M, Varut RM, Enache I, Osman A. Natural Antidiabetic Agents: Insights into Ericaceae-Derived Phenolics and Their Role in Metabolic and Oxidative Modulation in Diabetes. Pharmaceuticals. 2025; 18(5):682. https://doi.org/10.3390/ph18050682
Chicago/Turabian StylePopescu, Mihaela, Kristina Radivojevic, Diana-Maria Trasca, Renata Maria Varut, Irina Enache, and Andrei Osman. 2025. "Natural Antidiabetic Agents: Insights into Ericaceae-Derived Phenolics and Their Role in Metabolic and Oxidative Modulation in Diabetes" Pharmaceuticals 18, no. 5: 682. https://doi.org/10.3390/ph18050682
APA StylePopescu, M., Radivojevic, K., Trasca, D.-M., Varut, R. M., Enache, I., & Osman, A. (2025). Natural Antidiabetic Agents: Insights into Ericaceae-Derived Phenolics and Their Role in Metabolic and Oxidative Modulation in Diabetes. Pharmaceuticals, 18(5), 682. https://doi.org/10.3390/ph18050682