Unlocking the Neuroprotective Effect of Quercetin Against Cadmium-Induced Hippocampal Damage in Rats: PPARγ Activation as a Key Mechanism


Abstract
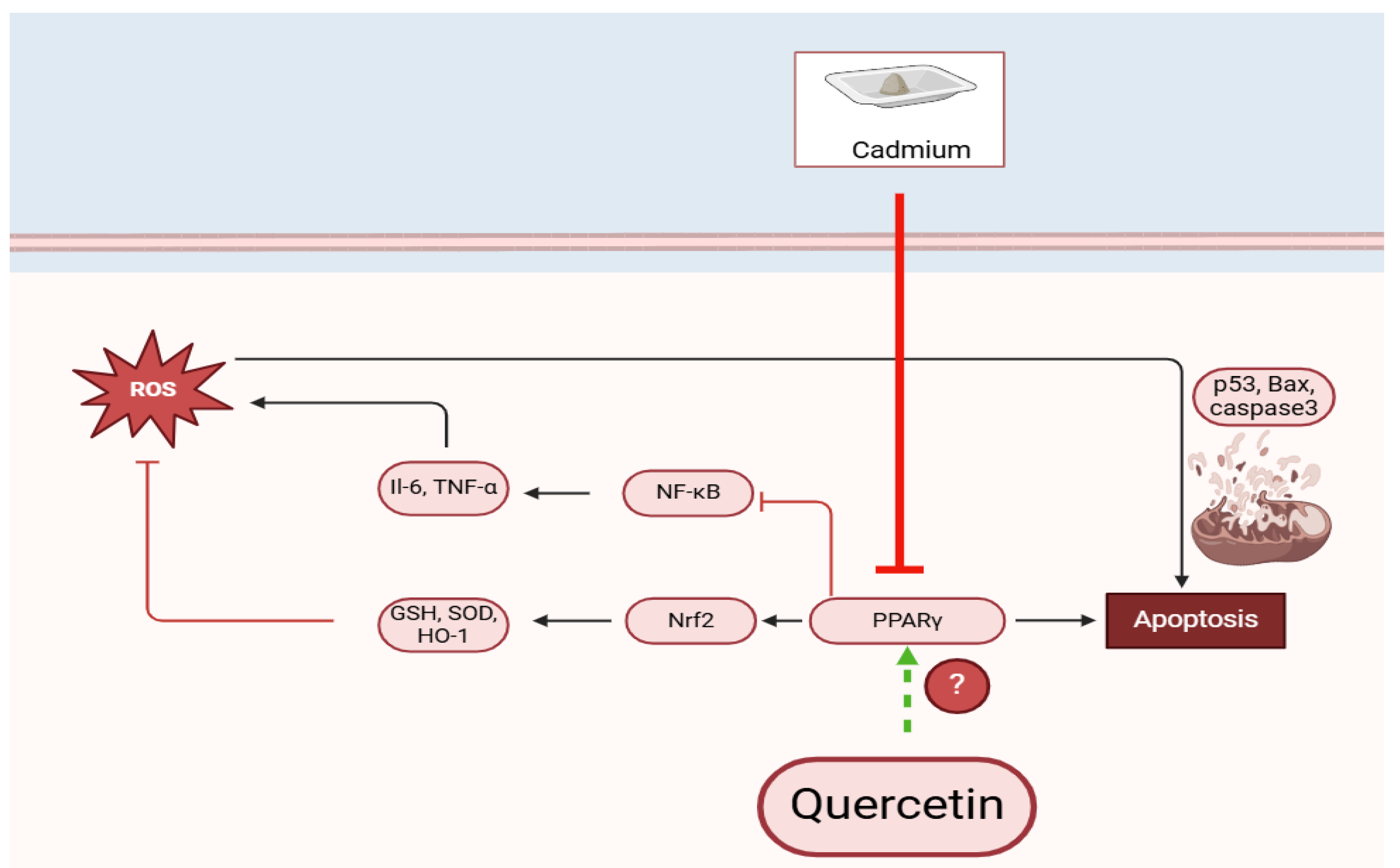
1. Introduction
2. Results
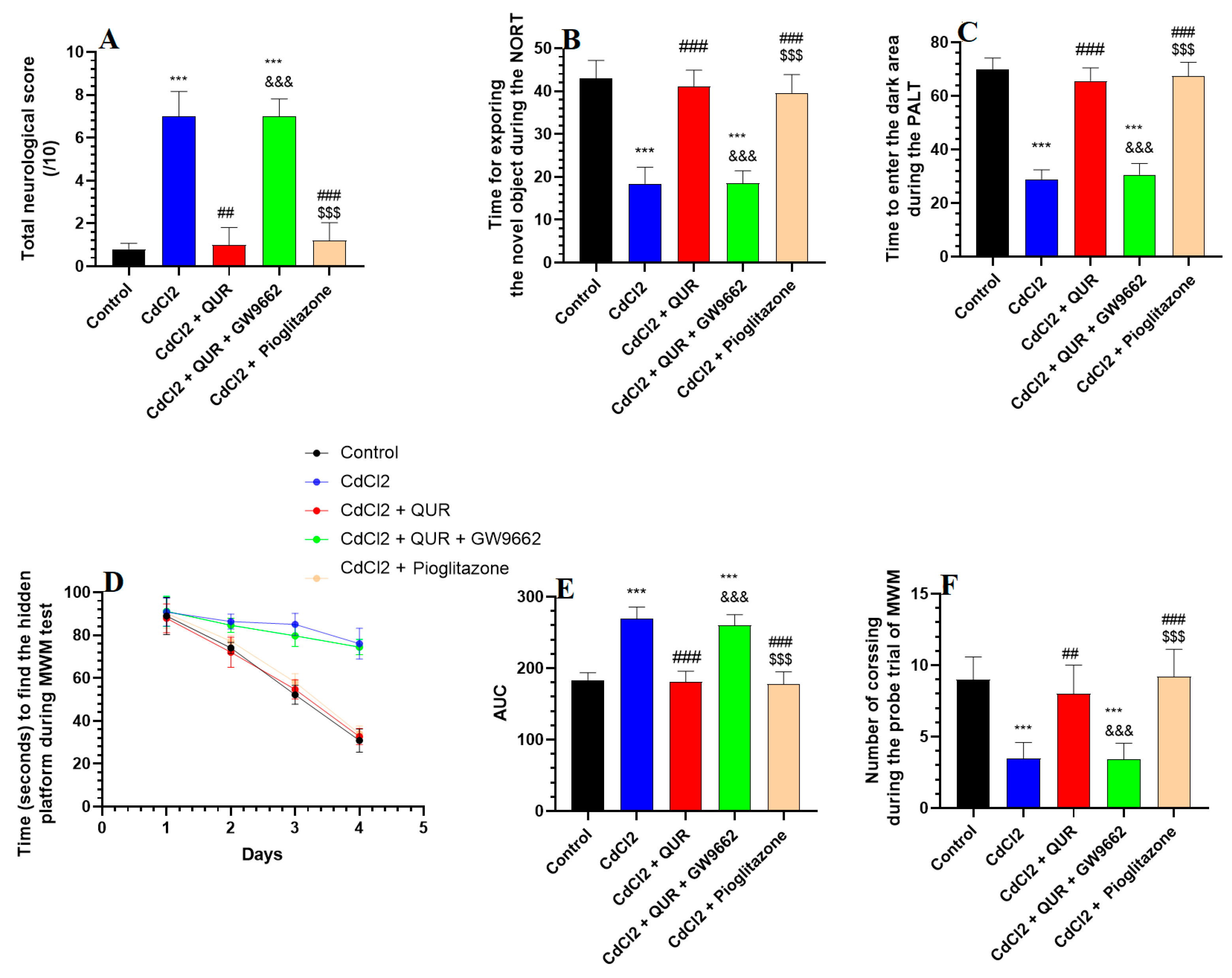
2.1. Effects of QUR and Pioglitazone on Neurological Scores and Memory Function in Rats
2.2. Effect on Hippocampal Levels of Markers of Inflammation and Oxidative Stress
2.3. Effect on Hippocampal Levels of BDNF and Other Cholinergic Markers
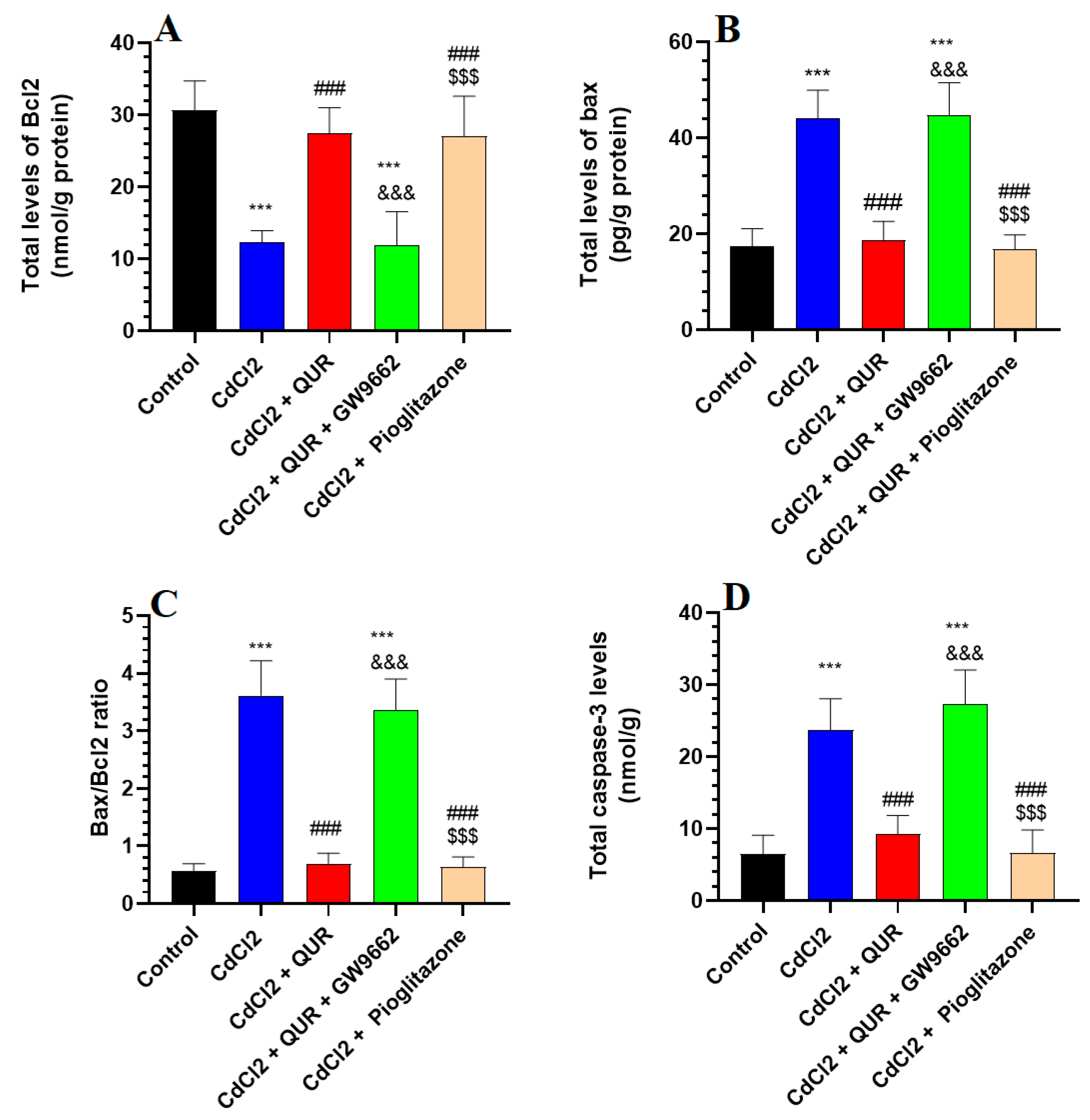
2.4. Effect on Apoptotic and Anti-Apoptotic Markers
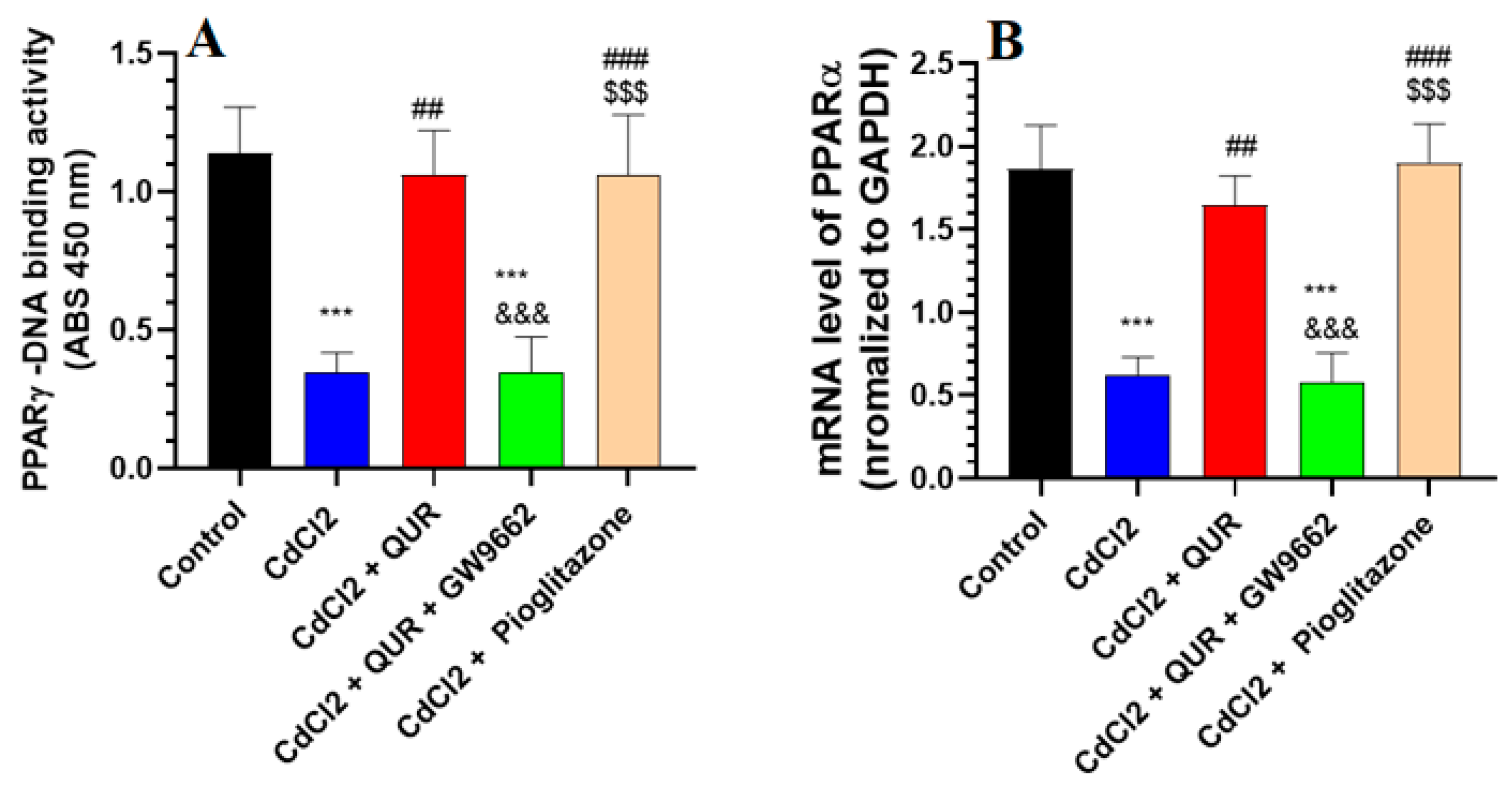
2.5. Effect on the mRNA and Nuclear Activity of PPARα
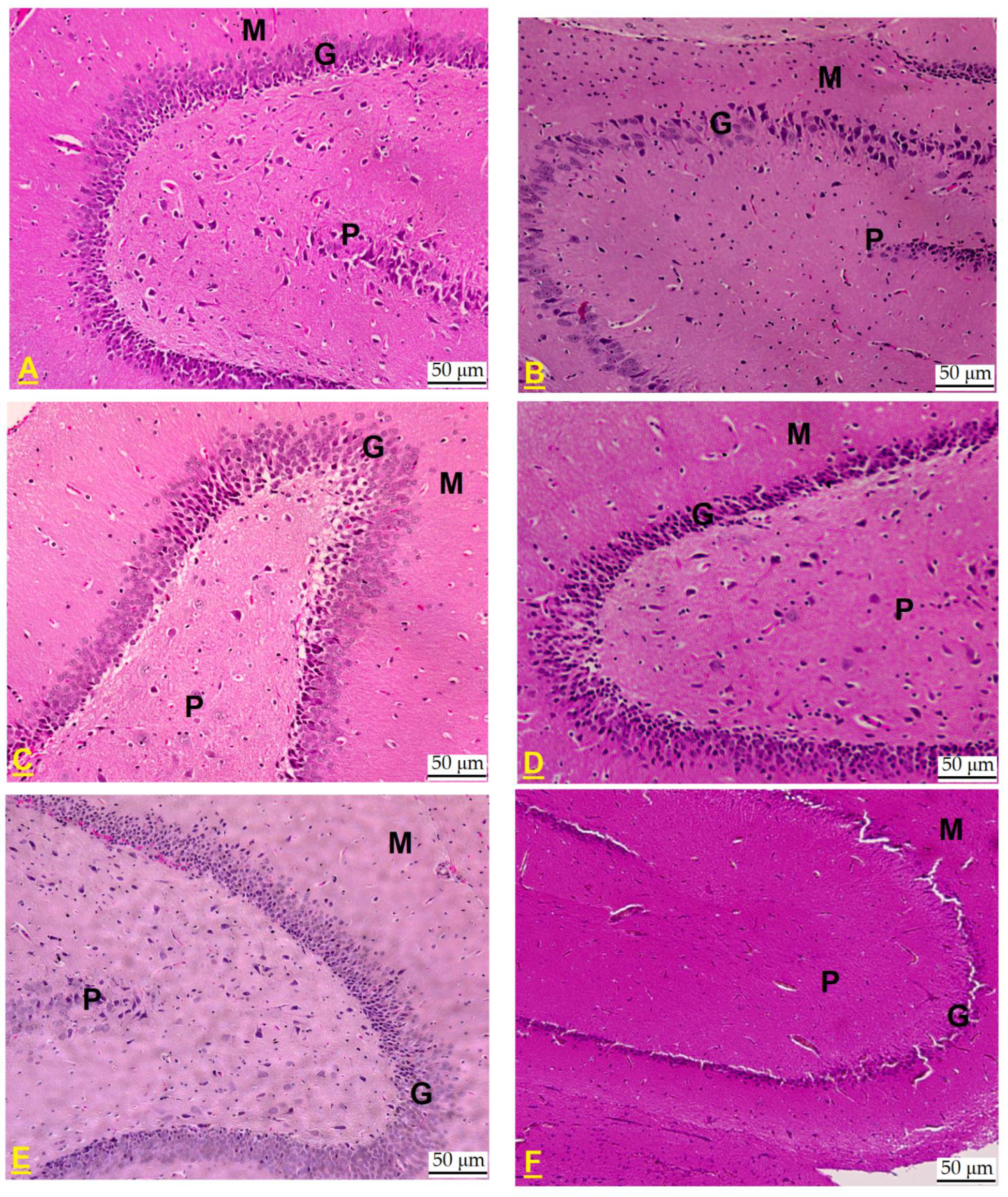
2.6. Effect on Dental Gyrus Histopathology
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Dose Selection and Protocol Regimen
4.4. Assessment of Neurological Score
4.5. Assessment of Memory Function
4.6. The Morris Water Maze (MWM)
4.7. Passive Learning Avoidance Test (PALT)
4.8. The Novel Object Recognition Test (NORT)
4.9. Collection of Brains and Hippocampi
4.10. Enzyme-Linked Immunosorbent Assays (ELISA)
4.11. Real-Time PCR (qPCR)
4.12. Determination of PPARα Nuclear Activity
4.13. Histological Assessment
4.14. Statistical Analysis
5. Conclusions
Study Limitations
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| QUR | Quercetin |
| CdCl2 | Cadmium chloride |
References
- Lamptey, R.N.; Chaulagain, B.; Trivedi, R.; Gothwal, A.; Layek, B.; Singh, J. A review of the common neurodegenerative disorders: Current therapeutic approaches and the potential role of nanotherapeutics. Int. J. Mol. Sci. 2022, 23, 1851. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, F.; Gülöksüz, S. Shedding light on the etiology of neurodegenerative diseases and dementia: The exposome paradigm. npj Ment. Health Res. 2022, 1, 20. [Google Scholar] [CrossRef] [PubMed]
- Arruebarrena, M.A.; Hawe, C.T.; Lee, Y.M.; Branco, R.C. Mechanisms of Cadmium Neurotoxicity. Int. J. Mol. Sci. 2023, 24, 16558. [Google Scholar] [CrossRef] [PubMed]
- Rahimzadeh, M.R.; Rahimzadeh, M.R.; Kazemi, S.; Moghadamnia, A.-A. Cadmium toxicity and treatment: An update. Casp. J. Intern. Med. 2017, 8, 135. [Google Scholar]
- Ji, Y.; Wang, H.; Zhao, X.; Wang, Q.; Zhang, C.; Zhang, Y.; Zhao, M.; Chen, Y.; Meng, X.; Xu, D. Crosstalk between endoplasmic reticulum stress and mitochondrial pathway mediates cadmium-induced germ cell apoptosis in testes. Toxicol. Sci. 2011, 124, 446–459. [Google Scholar] [CrossRef]
- Wang, B.; Du, Y. Cadmium and its neurotoxic effects. Oxidative Med. Cell. Longev. 2013, 2013, 898034. [Google Scholar] [CrossRef]
- Kumar, B.P.; Kumar, A.P.; Jose, J.A.; Prabitha, P.; Yuvaraj, S.; Chipurupalli, S.; Jeyarani, V.; Manisha, C.; Banerjee, S.; Jeyabalan, J.B. Minutes of PPAR-γ agonism and neuroprotection. Neurochem. Int. 2020, 140, 104814. [Google Scholar] [CrossRef]
- Risner, M.; Saunders, A.; Altman, J.; Ormandy, G.; Craft, S.; Foley, I.; Zvartau-Hind, M.; Hosford, D.; Roses, A. Efficacy of rosiglitazone in a genetically defined population with mild-to-moderate Alzheimer’s disease. Pharmacogenom. J. 2006, 6, 246–254. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Yao, L.; Gu, W.; Zhao, S.; Shen, Z.; Lin, Z.; Liu, W.; Yan, T. Pharmacological activity of quercetin: An updated review. Evid. Based Complement. Altern. Med. 2022, 2022, 3997190. [Google Scholar] [CrossRef]
- Mirza, M.A.; Mahmood, S.; Hilles, A.R.; Ali, A.; Khan, M.Z.; Zaidi, S.A.A.; Iqbal, Z.; Ge, Y. Quercetin as a therapeutic product: Evaluation of its pharmacological action and clinical applications—A review. Pharmaceuticals 2023, 16, 1631. [Google Scholar] [CrossRef]
- Abdalla, F.H.; Schmatz, R.; Cardoso, A.M.; Carvalho, F.B.; Baldissarelli, J.; de Oliveira, J.S.; Rosa, M.M.; Nunes, M.A.G.; Rubin, M.A.; da Cruz, I.B. Quercetin protects the impairment of memory and anxiogenic-like behavior in rats exposed to cadmium: Possible involvement of the acetylcholinesterase and Na+, K+-ATPase activities. Physiol. Behav. 2014, 135, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Unsal, C.; Kanter, M.; Aktas, C.; Erboga, M. Role of quercetin in cadmium-induced oxidative stress, neuronal damage, and apoptosis in rats. Toxicol. Ind. Health 2015, 31, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Kanter, M.; Unsal, C.; Aktas, C.; Erboga, M. Neuroprotective effect of quercetin against oxidative damage and neuronal apoptosis caused by cadmium in hippocampus. Toxicol. Ind. Health 2016, 32, 541–550. [Google Scholar] [CrossRef]
- Shati, A.A. Concomitant Administration of Quercetin and α-tocopherol Protects Rats from Cadmium Chloride Induced Neural Apoptosis and Cognitive Dysfunction. J. Eng. Appl. Sci. 2016, 3, 1–10. [Google Scholar] [CrossRef]
- Gupta, R.; Shukla, R.K.; Chandravanshi, L.P.; Srivastava, P.; Dhuriya, Y.K.; Shanker, J.; Singh, M.P.; Pant, A.B.; Khanna, V.K. Protective role of quercetin in cadmium-induced cholinergic dysfunctions in rat brain by modulating mitochondrial integrity and MAP kinase signaling. Mol. Neurobiol. 2017, 54, 4560–4583. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.; Cheng, X.; Qiao, X. The alleviative effects of quercetin on cadmium-induced necroptosis via inhibition ROS/iNOS/NF-κB pathway in the chicken brain. Biol. Trace Elem. Res. 2021, 199, 1584–1594. [Google Scholar] [CrossRef]
- Alshammari, G.M.; Al-Qahtani, W.H.; Alshuniaber, M.A.; Yagoub, A.E.A.; Al-Khalifah, A.S.; Al-Harbi, L.N.; Alhussain, M.H.; AlSedairy, S.A.; Yahya, M.A. Quercetin improves the impairment in memory function and attenuates hippocampal damage in cadmium chloride-intoxicated male rats by suppressing acetylcholinesterase and concomitant activation of SIRT1 signaling. J. Funct. Foods 2021, 86, 104675. [Google Scholar] [CrossRef]
- Larson-Casey, J.L.; Liu, S.; Pyles, J.M.; Lapi, S.E.; Saleem, K.; Antony, V.B.; Gonzalez, M.L.; Crossman, D.K.; Carter, A.B. Impaired PPARγ activation by cadmium exacerbates infection-induced lung injury. JCI Insight 2023, 8, e166608. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, J.; Zhang, Y.; Wang, X.; Li, Y.; Song, J.; Shao, J.; Su, P. Paternal cadmium exposure induces glucolipid metabolic reprogramming in offspring mice via PPAR signaling pathway. Chemosphere 2023, 339, 139592. [Google Scholar] [CrossRef]
- Mori, C.; Lee, J.-Y.; Tokumoto, M.; Satoh, M. Cadmium toxicity is regulated by peroxisome proliferator-activated receptor δ in human proximal tubular cells. Int. J. Mol. Sci. 2022, 23, 8652. [Google Scholar] [CrossRef]
- Reiterer, G.; Toborek, M.; Hennig, B. Quercetin protects against linoleic acid-induced porcine endothelial cell dysfunction. J. Nutr. 2004, 134, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-M.; Moon, J.; Cho, Y.; Chung, J.H.; Shin, M.-J. Quercetin up-regulates expressions of peroxisome proliferator-activated receptor γ, liver X receptor α, and ATP binding cassette transporter A1 genes and increases cholesterol efflux in human macrophage cell line. Nutr. Res. 2013, 33, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Beekmann, K.; Rubió, L.; de Haan, L.H.; Actis-Goretta, L.; van der Burg, B.; van Bladeren, P.J.; Rietjens, I.M. The effect of quercetin and kaempferol aglycones and glucuronides on peroxisome proliferator-activated receptor-gamma (PPAR-γ). Food Funct. 2015, 6, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.-H.; Yeh, C.-L.; Yeh, S.-L.; Lin, E.-S.; Wang, L.-Y.; Wang, Y.-H. Quercetin metabolites inhibit MMP-2 expression in A549 lung cancer cells by PPAR-γ associated mechanisms. J. Nutr. Biochem. 2016, 33, 45–53. [Google Scholar] [CrossRef]
- Castrejón-Tellez, V.; Rodríguez-Pérez, J.M.; Pérez-Torres, I.; Pérez-Hernández, N.; Cruz-Lagunas, A.; Guarner-Lans, V.; Vargas-Alarcón, G.; Rubio-Ruiz, M.E. The effect of resveratrol and quercetin treatment on PPAR mediated uncoupling protein (UCP-) 1, 2, and 3 expression in visceral white adipose tissue from metabolic syndrome rats. Int. J. Mol. Sci. 2016, 17, 1069. [Google Scholar] [CrossRef]
- Jia, Q.; Cao, H.; Shen, D.; Li, S.; Yan, L.; Chen, C.; Xing, S.; Dou, F. Quercetin protects against atherosclerosis by regulating the expression of PCSK9, CD36, PPARγ, LXRα and ABCA1. Int. J. Mol. Med. 2019, 44, 893–902. [Google Scholar] [CrossRef]
- Ballav, S.; Ranjan, A.; Basu, S. Partial Activation of PPAR-γ by Synthesized Quercetin Derivatives Modulates TGF-β1-Induced EMT in Lung Cancer Cells. Adv. Biol. 2023, 7, 2300037. [Google Scholar] [CrossRef]
- Alsaud, M.M.; Alhowail, A.H.; Aldubayan, M.A.; Almami, I.S. The ameliorative effect of pioglitazone against neuroinflammation caused by doxorubicin in rats. Molecules 2023, 28, 4775. [Google Scholar] [CrossRef]
- Branca, J.J.; Fiorillo, C.; Carrino, D.; Paternostro, F.; Taddei, N.; Gulisano, M.; Pacini, A.; Becatti, M. Cadmium-induced oxidative stress: Focus on the central nervous system. Antioxidants 2020, 9, 492. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, Y.; Zhao, S.; Chen, J.; Yang, J.; Wang, T.; Zou, H.; Wang, Y.; Gu, J.; Liu, X. Cadmium-induced apoptosis in neuronal cells is mediated by Fas/FasL-mediated mitochondrial apoptotic signaling pathway. Sci. Rep. 2018, 8, 8837. [Google Scholar] [CrossRef]
- Mimouna, S.B.; Chemek, M.; Boughammoura, S.; Banni, M.; Messaoudi, I. Early-life exposure to cadmium triggers distinct Zn-dependent protein expression patterns and impairs brain development. Biol. Trace Elem. Res. 2018, 184, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Batool, A.; Saleem, S.; Naqvi, F.; Hasan, K.A.; Naqvi, F.; Haider, S. Thymol mitigates cadmium-induced behavioral and cognitive deficits by up-regulating hippocampal BDNF levels in rats. Pak. J. Pharm. Sci. 2022, 35. [Google Scholar]
- El-Kott, A.F.; Bin-Meferij, M.M.; Eleawa, S.M.; Alshehri, M.M. Kaempferol protects against cadmium chloride-induced memory loss and hippocampal apoptosis by increased intracellular glutathione stores and activation of PTEN/AMPK induced inhibition of Akt/mTOR signaling. Neurochem. Res. 2020, 45, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Shati, A.A.; Alfaifi, M.Y. Trans-resveratrol inhibits tau phosphorylation in the brains of control and cadmium chloride-treated rats by activating PP2A and PI3K/Akt induced-inhibition of GSK3β. Neurochem. Res. 2019, 44, 357–373. [Google Scholar] [CrossRef]
- Shagirtha, K.; Muthumani, M.; Prabu, S.M. Melatonin abrogates cadmium induced oxidative stress related neurotoxicity in rats. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1039–1050. [Google Scholar]
- Kim, S.; Cheon, H.-S.; Kim, S.-Y.; Juhnn, Y.-S.; Kim, Y.-Y. Cadmium induces neuronal cell death through reactive oxygen species activated by GADD153. BMC Cell Biol. 2013, 14, 4. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Wang, Y.; Wang, C. Effect of cadmium choride on oxidative damage and expression of NF-κB in brain tissue of mice. Wei Sheng Yan Jiu J. Hyg. Res. 2017, 46, 807–812. [Google Scholar]
- Winiarska-Mieczan, A. Protective effect of tea against lead and cadmium-induced oxidative stress—A review. Biometals 2018, 31, 909–926. [Google Scholar] [CrossRef]
- Tang, K.-K.; Liu, X.-Y.; Wang, Z.-Y.; Qu, K.-C.; Fan, R.-F. Trehalose alleviates cadmium-induced brain damage by ameliorating oxidative stress, autophagy inhibition, and apoptosis. Metallomics 2019, 11, 2043–2051. [Google Scholar] [CrossRef]
- Shati, A.A.; El-Kott, A.F. Resolvin D1 protects against cadmium chloride-induced memory loss and hippocampal damage in rats: A comparison with docosahexaenoic acid. Hum. Exp. Toxicol. 2021, 40, S215–S232. [Google Scholar] [CrossRef]
- Arab, H.H.; Eid, A.H.; Alsufyani, S.E.; Ashour, A.M.; El-Sheikh, A.A.; Darwish, H.W.; Georgy, G.S. Neuroprotective impact of linagliptin against cadmium-induced cognitive impairment and neuropathological aberrations: Targeting SIRT1/Nrf2 axis, apoptosis, and autophagy. Pharmaceuticals 2023, 16, 1065. [Google Scholar] [CrossRef] [PubMed]
- Corona, J.C.; Duchen, M.R. PPARγ and PGC-1α as therapeutic targets in Parkinson’s. Neurochem. Res. 2015, 40, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Nicolakakis, N.; Hamel, E. The nuclear receptor PPARγ as a therapeutic target for cerebrovascular and brain dysfunction in Alzheimer’s disease. Front. Aging Neurosci. 2010, 2, 1389. [Google Scholar] [CrossRef]
- D’Angelo, M.; Castelli, V.; Catanesi, M.; Antonosante, A.; Dominguez-Benot, R.; Ippoliti, R.; Benedetti, E.; Cimini, A. PPARγ and cognitive performance. Int. J. Mol. Sci. 2019, 20, 5068. [Google Scholar] [CrossRef]
- Chiang, M.-C.; Chen, C.-M.; Lee, M.-R.; Chen, H.-W.; Chen, H.-M.; Wu, Y.-S.; Hung, C.-H.; Kang, J.-J.; Chang, C.-P.; Chang, C. Modulation of energy deficiency in Huntington’s disease via activation of the peroxisome proliferator-activated receptor gamma. Hum. Mol. Genet. 2010, 19, 4043–4058. [Google Scholar] [CrossRef]
- Chiang, M.-C.; Tsai, T.-Y.; Wang, C.-J. The potential benefits of quercetin for brain health: A review of anti-inflammatory and neuroprotective mechanisms. Int. J. Mol. Sci. 2023, 24, 6328. [Google Scholar] [CrossRef]
- Costa, L.G.; Garrick, J.M.; Roquè, P.J.; Pellacani, C. Mechanisms of neuroprotection by quercetin: Counteracting oxidative stress and more. Oxidative Med. Cell. Longev. 2016, 2016, 2986796. [Google Scholar] [CrossRef]
- Babaei, F.; Mirzababaei, M.; Nassiri-Asl, M. Quercetin in food: Possible mechanisms of its effect on memory. J. Food Sci. 2018, 83, 2280–2287. [Google Scholar] [CrossRef]
- Spencer, J.P. The interactions of flavonoids within neuronal signalling pathways. Genes Nutr. 2007, 2, 257–273. [Google Scholar] [CrossRef]
- Jiang, W.; Luo, T.; Li, S.; Zhou, Y.; Shen, X.-Y.; He, F.; Xu, J.; Wang, H.-Q. Quercetin protects against okadaic acid-induced injury via MAPK and PI3K/Akt/GSK3β signaling pathways in HT22 hippocampal neurons. PLoS ONE 2016, 11, e0152371. [Google Scholar] [CrossRef]
- Zubčić, K.; Radovanović, V.; Vlainić, J.; Hof, P.R.; Oršolić, N.; Šimić, G.; Jembrek, M.J. PI3K/Akt and ERK1/2 Signalling Are Involved in Quercetin-Mediated Neuroprotection against Copper-Induced Injury. Oxidative Med. Cell. Longev. 2020, 2020, 9834742. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shoorei, H.; Sasi, A.K.; Taheri, M.; Ayatollahi, S.A. The impact of the phytotherapeutic agent quercetin on expression of genes and activity of signaling pathways. Biomed. Pharmacother. 2021, 141, 111847. [Google Scholar] [CrossRef] [PubMed]
- Falcone, R.; Florio, T.M.; Giacomo, E.D.; Benedetti, E.; Cristiano, L.; Antonosante, A.; Fidoamore, A.; Massimi, M.; Alecci, M.; Ippoliti, R. PPARβ/δ and γ in a rat model of Parkinson’s disease: Possible involvement in PD symptoms. J. Cell. Biochem. 2015, 116, 844–855. [Google Scholar] [CrossRef]
- Gao, F.; Zang, L.; Wu, D.; Li, Y.; Zhang, Q.; Wang, H.; Tian, G.; Mu, Y. Pioglitazone improves the ability of learning and memory via activating ERK1/2 signaling pathway in the hippocampus of T2DM rats. Neurosci. Lett. 2017, 651, 165–170. [Google Scholar] [CrossRef]
- Beheshti, F.; Hosseini, M.; Hashemzehi, M.; Soukhtanloo, M.; Khazaei, M.; Shafei, M.N. The effects of PPAR-γ agonist pioglitazone on hippocampal cytokines, brain-derived neurotrophic factor, memory impairment, and oxidative stress status in lipopolysaccharide-treated rats. Iran. J. Basic Med. Sci. 2019, 22, 940. [Google Scholar]
- Huang, R.; Zhang, C.; Wang, X.; Hu, H. PPARγ in ischemia-reperfusion injury: Overview of the biology and therapy. Front. Pharmacol. 2021, 12, 600618. [Google Scholar] [CrossRef]
- Alhowail, A.; Alsikhan, R.; Alsaud, M.; Aldubayan, M.; Rabbani, S.I. Protective effects of pioglitazone on cognitive impairment and the underlying mechanisms: A review of literature. Drug Des. Dev. Ther. 2022, 16, 2919–2931. [Google Scholar] [CrossRef]
- Fernandez-Martos, C.M.; Atkinson, R.A.; Chuah, M.I.; King, A.E.; Vickers, J.C. Combination treatment with leptin and pioglitazone in a mouse model of Alzheimer’s disease. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2017, 3, 92–106. [Google Scholar] [CrossRef]
- Seok, H.; Lee, M.; Shin, E.; Yun, M.R.; Lee, Y.-H.; Moon, J.H.; Kim, E.; Lee, P.H.; Lee, B.-W.; Kang, E.S. Low-dose pioglitazone can ameliorate learning and memory impairment in a mouse model of dementia by increasing LRP1 expression in the hippocampus. Sci. Rep. 2019, 9, 4414. [Google Scholar] [CrossRef]
- Victor, N.; Wanderi, E.; Gamboa, J.; Zhao, X.; Aronowski, J.; Deininger, K.; Lust, W.; Landreth, G.; Sundararajan, S. Altered PPARγ expression and activation after transient focal ischemia in rats. Eur. J. Neurosci. 2006, 24, 1653–1663. [Google Scholar] [CrossRef]
- Mandrekar-Colucci, S.; Sauerbeck, A.; Popovich, P.G.; McTigue, D.M. PPAR agonists as therapeutics for CNS trauma and neurological diseases. ASN Neuro 2013, 5, AN20130030. [Google Scholar] [CrossRef] [PubMed]
- Sundararajan, S.; Gamboa, J.; Victor, N.; Wanderi, E.; Lust, W.; Landreth, G. Peroxisome proliferator-activated receptor-γ ligands reduce inflammation and infarction size in transient focal ischemia. Neuroscience 2005, 130, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Godoy, J.A.; Quintanilla, R.A.; Koenig, C.S.; Bronfman, M. Peroxisome proliferator-activated receptor γ is expressed in hippocampal neurons and its activation prevents β-amyloid neurodegeneration: Role of Wnt signaling. Exp. Cell Res. 2005, 304, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Wu, J.-S.; Tsai, H.-D.; Huang, C.-Y.; Chen, J.-J.; Sun, G.Y.; Lin, T.-N. Peroxisome proliferator-activated receptor gamma (PPAR-γ) and neurodegenerative disorders. Mol. Neurobiol. 2012, 46, 114–124. [Google Scholar] [CrossRef]
- Zolezzi, J.M.; Silva-Alvarez, C.; Ordenes, D.; Godoy, J.A.; Carvajal, F.J.; Santos, M.J.; Inestrosa, N.C. Peroxisome proliferator-activated receptor (PPAR) γ and PPARα agonists modulate mitochondrial fusion-fission dynamics: Relevance to reactive oxygen species (ROS)-related neurodegenerative disorders? PLoS ONE 2013, 8, e64019. [Google Scholar] [CrossRef]
- Cai, W.; Yang, T.; Liu, H.; Han, L.; Zhang, K.; Hu, X.; Zhang, X.; Yin, K.-J.; Gao, Y.; Bennett, M.V. Peroxisome proliferator-activated receptor γ (PPARγ): A master gatekeeper in CNS injury and repair. Prog. Neurobiol. 2018, 163, 27–58. [Google Scholar] [CrossRef]
- Wang, L.; Botchway, B.O.; Liu, X. The repression of the HMGB1-TLR4-NF-κB signaling pathway by safflower yellow may improve spinal cord injury. Front. Neurosci. 2021, 15, 803885. [Google Scholar] [CrossRef]
- Duan, C.; Jiao, D.; Wang, H.; Wu, Q.; Men, W.; Yan, H.; Li, C. Activation of the PPARγ prevents ferroptosis-induced neuronal loss in response to intracerebral hemorrhage through synergistic actions with the Nrf2. Front. Pharmacol. 2022, 13, 869300. [Google Scholar] [CrossRef]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; de Oliveira, R.M.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-γ. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef]
- Han, L.; Zhou, R.; Niu, J.; McNutt, M.A.; Wang, P.; Tong, T. SIRT1 is regulated by a PPARγ–SIRT1 negative feedback loop associated with senescence. Nucleic Acids Res. 2010, 38, 7458–7471. [Google Scholar] [CrossRef]
- Ruderman, N.B.; Xu, X.J.; Nelson, L.; Cacicedo, J.M.; Saha, A.K.; Lan, F.; Ido, Y. AMPK and SIRT1: A long-standing partnership? Am. J. Physiol. Endocrinol. Metab. 2010, 298, E751–E760. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.; Ying, N.; Lai, J.; Feng, L.; Zheng, S.; Jiang, F.; Song, Q.; Chai, H.; Dou, X. Activation of the AMPK-SIRT1 pathway contributes to protective effects of Salvianolic acid A against lipotoxicity in hepatocytes and NAFLD in mice. Front. Pharmacol. 2020, 11, 560905. [Google Scholar]
- Fryer, L.G.; Parbu-Patel, A.; Carling, D. The anti-diabetic drugs rosiglitazone and metformin stimulate AMP-activated protein kinase through distinct signaling pathways. J. Biol. Chem. 2002, 277, 25226–25232. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.K.; Avilucea, P.R.; Ye, J.-M.; Assifi, M.M.; Kraegen, E.W.; Ruderman, N.B. Pioglitazone treatment activates AMP-activated protein kinase in rat liver and adipose tissue in vivo. Biochem. Biophys. Res. Commun. 2004, 314, 580–585. [Google Scholar] [CrossRef]
- Lan, L.F.; Zheng, L.; Yang, X.; Ji, X.T.; Fan, Y.H.; Zeng, J.S. Peroxisome Proliferator-activated Receptor-γ Agonist Pioglitazone Ameliorates White Matter Lesion and Cognitive Impairment in Hypertensive Rats. CNS Neurosci. Ther. 2015, 21, 410–416. [Google Scholar] [CrossRef]
- Sun, L.; Yuan, Q.; Xu, T.; Yao, L.; Feng, J.; Ma, J.; Wang, L.; Lu, C.; Wang, D. Pioglitazone; a peroxisome proliferator-activated receptor γ agonist, ameliorates chronic kidney disease by enhancing antioxidative capacity and attenuating angiogenesis in the kidney of a 5/6 nephrectomized rat model. Cell. Physiol. Biochem. 2016, 38, 1831–1840. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Q.; Dai, W.; Hua, B.; Li, H.; Li, W. Pioglitazone downregulates Twist-1 expression in the kidney and protects renal function of Zucker diabetic fatty rats. Biomed. Pharmacother. 2019, 118, 109346. [Google Scholar] [CrossRef]
- Liu, M.; Bachstetter, A.D.; Cass, W.A.; Lifshitz, J.; Bing, G. Pioglitazone attenuates neuroinflammation and promotes dopaminergic neuronal survival in the nigrostriatal system of rats after diffuse brain injury. J. Neurotrauma 2017, 34, 414–422. [Google Scholar] [CrossRef]
- Sun, M.-H.; Chen, K.-J.; Sun, C.-C.; Tsai, R.-K. Protective Effect of Pioglitazone on Retinal Ganglion Cells in an Experimental Mouse Model of Ischemic Optic Neuropathy. Int. J. Mol. Sci. 2022, 24, 411. [Google Scholar] [CrossRef]
- Vázquez-González, D.; Corona, J.C. Pioglitazone enhances brain mitochondrial biogenesis and phase II detoxification capacity in neonatal rats with 6-OHDA-induced unilateral striatal lesions. Front. Neurosci. 2023, 17, 1186520. [Google Scholar] [CrossRef]
- Baumann, A.; Burger, K.; Brandt, A.; Staltner, R.; Jung, F.; Rajcic, D.; Pisarello, M.J.L.; Bergheim, I. GW9662, a peroxisome proliferator-activated receptor gamma antagonist, attenuates the development of non-alcoholic fatty liver disease. Metabolism 2022, 133, 155233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-S.; Li, H.; Zhang, D.-D.; Yan, H.-Y.; Zhang, Z.-H.; Zhou, C.-H.; Ye, Z.-N.; Chen, Q.; Jiang, T.-W.; Liu, J.-P. Inhibition of myeloid differentiation factor 88 (MyD88) by ST2825 provides neuroprotection after experimental traumatic brain injury in mice. Brain Res. 2016, 1643, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Alfaris, N.; Alshammari, G.; Altamimi, J.; Aljabryn, D.; Alagal, R.; Aldera, H.; Alkhateeb, M.; Yahya, M. Ellagic acid prevents streptozotocin-induced hippocampal damage and memory loss in rats by stimulating Nrf2 and nuclear factor-κB, and activating insulin receptor substrate/PI3K/Akt axis. J. Physiol. Pharmacol. 2021, 72, 503–515. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | CdCl2 | CdCl2 + QUR | CdCl2 + QUR + GW9662 | CdCl2 + Pioglitazone |
|---|---|---|---|---|---|
| Total Levels | |||||
| MDA (nmol/g protein) | 0.65 ± 0.08 | 2.88 ± 0.39 *** | 0.89 ± 0.06 *,### | 2.62 ± 0.27 ***,&&& | 0.71 ± 0.09 ###,$$$ |
| GSH (µg/g protein) | 76.5 ± 6.8 | 27.8 ± 2.4 *** | 79.4 ± 7.1 ### | 31.4 ± 3.3 ***,&&& | 71.4 ± 8.3 ###,$$$ |
| SOD (U/g protein) | 29.7 ± 2.9 | 11.3 ± 1.6 *** | 26.7 ± 2.1 ### | 13.5 ± 1.8 ***,&&& | 31.2 ± 3.7 ###,$$$ |
| HO-1 (U/g protein) | 24.5 ± 2.7 | 8.7 ± 0.92 *** | 22.4 ± 3.2 ### | 9.4 ± 0.82 ***,&&& | 26.5 ± 2.1 ###,$$$ |
| 8-OHdG (pg/g protein) | 320.2 ± 27.7 | 738.2 ± 66.4 *** | 305 ± 37.3 ### | 822.5 ± 73.4 ***,&&& | 352 ± 44.7 ###,$$$ |
| RAGE (ng/g protein) | 7.6 ± 0.64 | 25.7 ± 3.1 *** | 9.4 ± 0.83 *,### | 27.5 ± 2.6 ***,&&& | 8.5 ± 0.67 ###,$$$ |
| TNF-α (pg/g protein) | 10.6 ± 0.97 | 46.5 ± 5.2 *** | 17.5 ± 1.8 **,### | 51.2 ± 6.4 ***,&&& | 16.8 ± 1.4 ###,$$$ |
| IL-6 (pg/g protein) | 2.4 ± 2.4 | 37.6 ± 3.1 *** | 6.7 ± 0.72 ***,### | 41.2 ± 5.7 ***,&&& | 8.5 ± 0.91 ###,$$$ |
| Nuclear levels | |||||
| Nrf2 (pg/g protein) | 325.5 ± 24.5 | 122.5 ± 11.9 *** | 211.3 ± 22.8 ### | 143.6 ± 15.3 ***,&&& | 318.7 ± 28.4 ###,$$$ |
| NF-κβ p65 (pg/g protein) | 219.2 ± 18.9 | 513.1 ± 48.4 *** | 245.8 ± 22.1 *,### | 489.7 ± 43.2 ***,&&& | 239.9 ± 26.6 ###,$$$ |
| Parameter | Control | CdCl2 | CdCl2 + QUR | CdCl2 + QUR + GW9662 | CdCl2 + Pioglitazone |
|---|---|---|---|---|---|
| BDNF (ng/g protein) | 104.2 ± 12.4 | 43.2 ± 4.6 *** | 87.9 ± 6.8 **,### | 38.5 ± 3.1 ***,&& | 91.9 ± 7.7 ###,$$$ |
| Ach (μmol/g protein) | 4.56 ± 0.27 | 1.63 ± 0.26 *** | 3.92 ± 0.41 *,### | 1.29 ± 0.22 ***,#,&&& | 4.34 ± 0.62 ###,$$$ |
| AchE (pg/g protein) | 18.5 ± 1.7 | 46.5 ± 6.4 *** | 22.5 ± 1.9 *,### | 51.8 ± 5.8 ***,#,&&& | 19.3 ± 1.3 ###,$$$ |
| ChAT (mU/gprotein) | 11.42 ± 1.3 | 4.52 ± 0.44 *** | 10.21 ± 1.37 ### | 3.42 ± 0.45 ***,#,&&& | 9.88 ± 0.86 ###,$$$ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Nouri, D.M. Unlocking the Neuroprotective Effect of Quercetin Against Cadmium-Induced Hippocampal Damage in Rats: PPARγ Activation as a Key Mechanism. Pharmaceuticals 2025, 18, 657. https://doi.org/10.3390/ph18050657
Al-Nouri DM. Unlocking the Neuroprotective Effect of Quercetin Against Cadmium-Induced Hippocampal Damage in Rats: PPARγ Activation as a Key Mechanism. Pharmaceuticals. 2025; 18(5):657. https://doi.org/10.3390/ph18050657
Chicago/Turabian StyleAl-Nouri, Doha M. 2025. "Unlocking the Neuroprotective Effect of Quercetin Against Cadmium-Induced Hippocampal Damage in Rats: PPARγ Activation as a Key Mechanism" Pharmaceuticals 18, no. 5: 657. https://doi.org/10.3390/ph18050657
APA StyleAl-Nouri, D. M. (2025). Unlocking the Neuroprotective Effect of Quercetin Against Cadmium-Induced Hippocampal Damage in Rats: PPARγ Activation as a Key Mechanism. Pharmaceuticals, 18(5), 657. https://doi.org/10.3390/ph18050657