
Beyond Cannabidiol: The Contribution of Cannabis sativa Phytocomplex to Skin Anti-Inflammatory Activity in Human Skin Keratinocytes
 , , , , ,
, , , , ,  , and
, and
Abstract
1. Introduction
2. Results
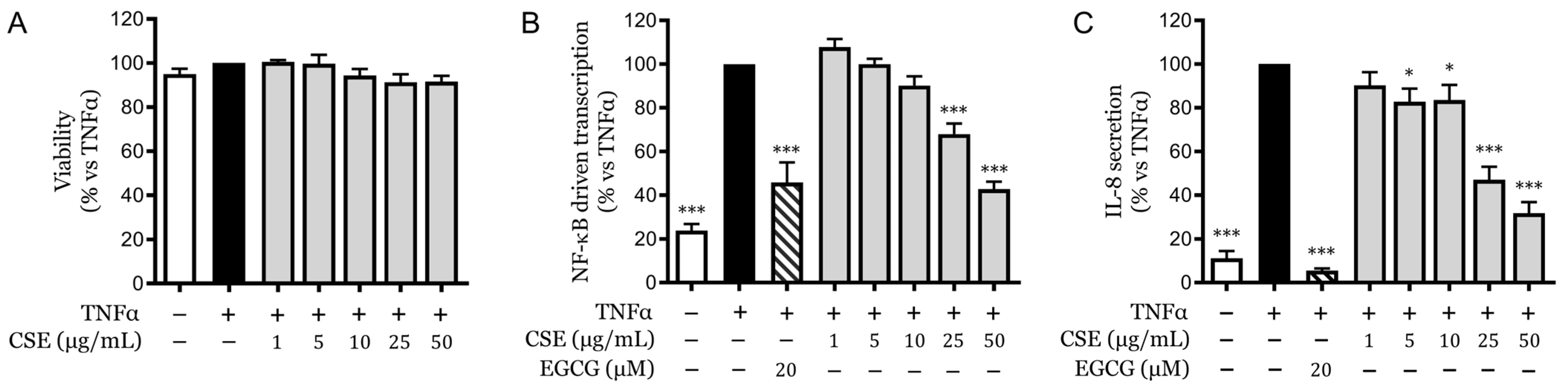
2.1. Cytotoxicity and Anti-Inflammatory Effects of CSE
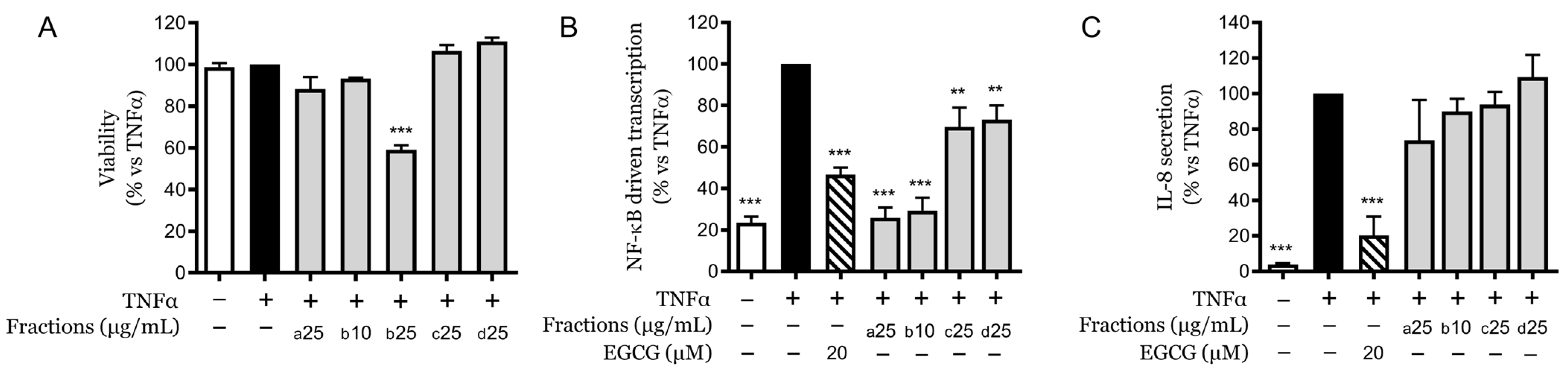
2.2. Fractionation and Differential Cytotoxicity
2.3. Evaluation of Anti-Inflammatory Effects of Fractions at Non-Toxic Concentrations
2.4. Analytical Profiling of Fraction Components
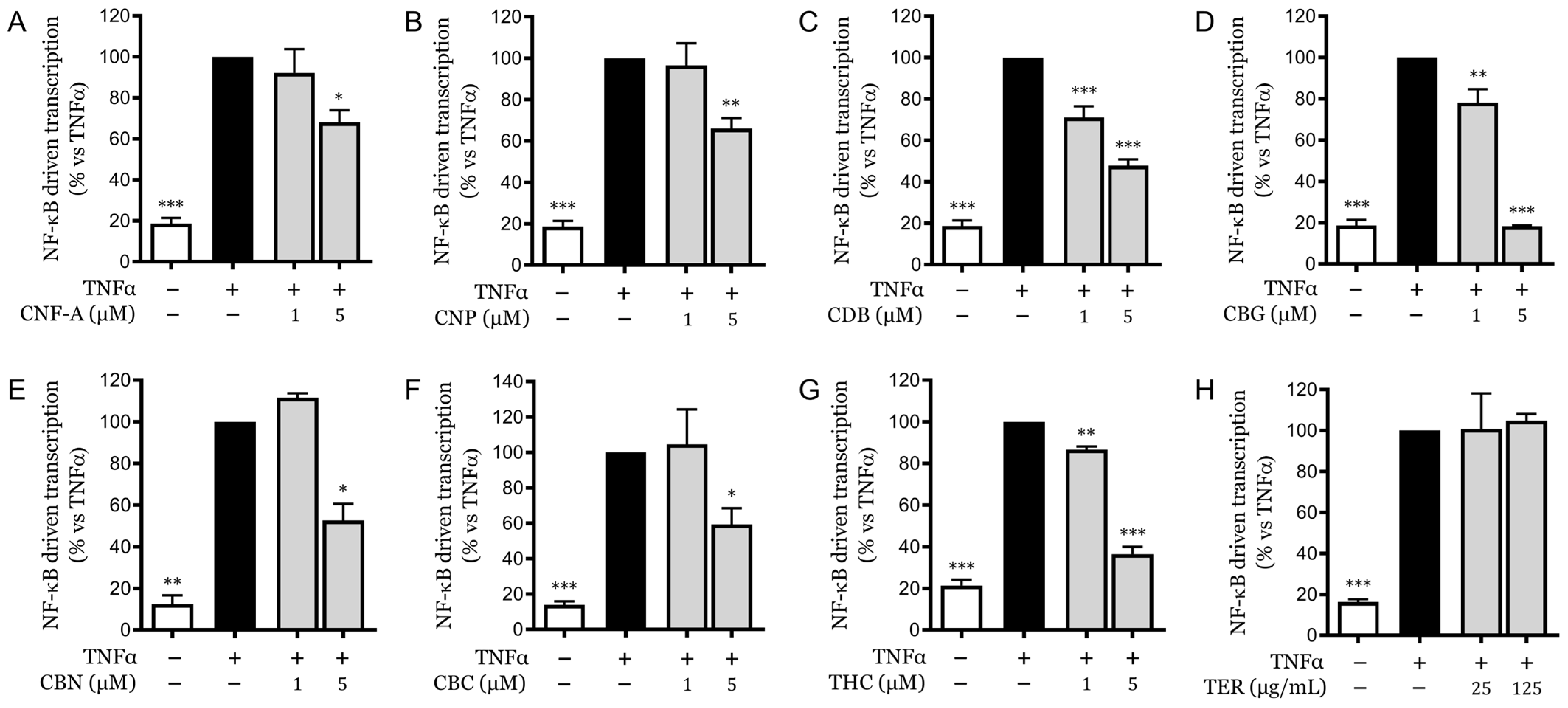
2.5. Analysis of Pure Compounds
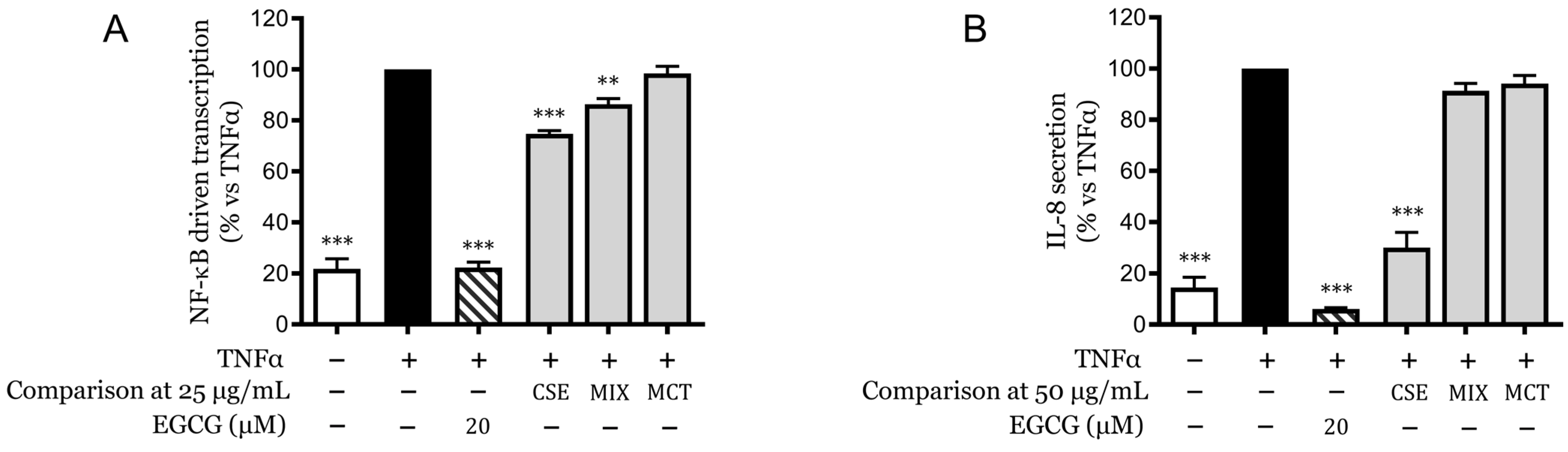
2.6. Synergistic Effects of Cannabinoids and Terpenes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Isolation of Compounds
4.2. Phytochemical Analysis
4.3. Cell Culture Procedures
4.4. Cell Treatment and Assays
4.5. Cytotoxicity Assay
4.6. Measurement of NF-κB Activity
4.7. Measurement of IL-8 Release
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.M.; Gomes, A.L.; Vilas Boas, I.; Marto, J.; Ribeiro, H.M. Cannabis-Based Products for the Treatment of Skin Inflammatory Diseases: A Timely Review. Pharmaceuticals 2022, 15, 210. [Google Scholar] [CrossRef]
- Makhakhe, L. Topical cannabidiol (CBD) in skin pathology—A comprehensive review and prospects for new therapeutic opportunities. S. Afr. Fam. Pract. 2022, 64, e1–e4. [Google Scholar] [CrossRef]
- Martinelli, G.; Magnavacca, A.; Fumagalli, M.; DellʼAgli, M.; Piazza, S.; Sangiovanni, E. Cannabis sativa and Skin Health: Dissecting the Role of Phytocannabinoids. Planta Med. 2022, 88, 492–506. [Google Scholar] [CrossRef] [PubMed]
- McDougall, J.J.; McKenna, M.K. Anti-Inflammatory and Analgesic Properties of the Cannabis Terpene Myrcene in Rat Adjuvant Monoarthritis. Int. J. Mol. Sci. 2022, 23, 7891. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, J.K.; O’Riordan, D.; Downer, E.J. Cannflavin A inhibits TLR4-induced chemokine and cytokine expression in human macrophages. Nat. Prod. Res. 2024, 23, 1–7. [Google Scholar] [CrossRef]
- Li, H.; Deng, N.; Puopolo, T.; Jiang, X.; Seeram, N.P.; Liu, C.; Ma, H. Cannflavins A and B with Anti-Ferroptosis, Anti-Glycation, and Antioxidant Activities Protect Human Keratinocytes in a Cell Death Model with Erastin and Reactive Carbonyl Species. Nutrients 2023, 15, 4565. [Google Scholar] [CrossRef] [PubMed]
- O’Croinin, C.; Guerra, A.G.; Doschak, M.R.; Löbenberg, R.; Davies, N.M. Therapeutic Potential and Predictive Pharmaceutical Modeling of Stilbenes in Cannabis sativa. Pharmaceutics 2023, 15, 26. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Nistico, S.; Paolillo, N.; Minella, D.; Piccirilli, S.; Rispoli, V.; Giardina, E.; Biancolella, M.; Chimenti, S.; Novelli, G.; Nistico, G. Effects of TNF-alpha and IL-1 beta on the activation of genes related to inflammatory, immune responses and cell death in immortalized human HaCat keratinocytes. Int. J. Immunopathol. Pharmacol. 2010, 23, 1057–1072. [Google Scholar] [CrossRef]
- Raingeaud, J.; Pierre, J. Interleukin-4 downregulates TNFα-induced IL-8 production in keratinocytes. FEBS Lett. 2005, 579, 3953–3959. [Google Scholar] [CrossRef] [PubMed]
- Stanescu, A.M.A.; Bejan, G.C.; Balta, M.D.; Andronic, O.; Toma, C.; Busnatu, S. The Perspective of Cannabidiol in Psoriasis Therapy. Psoriasis 2024, 14, 51–61. [Google Scholar] [CrossRef]
- Jurga, M.; Jurga, A.; Jurga, K.; Kazmierczak, B.; Kusmierczyk, K.; Chabowski, M. Cannabis-Based Phytocannabinoids: Overview, Mechanism of Action, Therapeutic Application, Production, and Affecting Environmental Factors. Int. J. Mol. Sci. 2024, 25, 11258. [Google Scholar] [CrossRef] [PubMed]
- Niyangoda, D.; Muayad, M.; Tesfaye, W.; Bushell, M.; Ahmad, D.; Samarawickrema, I.; Sinclair, J.; Kebriti, S.; Maida, V.; Thomas, J. Cannabinoids in Integumentary Wound Care: A Systematic Review of Emerging Preclinical and Clinical Evidence. Pharmaceutics 2024, 16, 1081. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Fumagalli, M.; Pacchetti, B.; Piazza, S.; Magnavacca, A.; Khalilpour, S.; Melzi, G.; Martinelli, G.; Dell’Agli, M. Cannabis sativa, L. Extract and Cannabidiol Inhibit In Vitro Mediators of Skin Inflammation and Wound Injury. Phytother. Res. 2019, 33, 2083–2093. [Google Scholar] [CrossRef]
- Petrosino, S.; Verde, R.; Vaia, M.; Allarà, M.; Iuvone, T.; Di Marzo, V. Anti-inflammatory Properties of Cannabidiol, a Nonpsychotropic Cannabinoid, in Experimental Allergic Contact Dermatitis. J. Pharmacol. Exp. Ther. 2018, 365, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Goldminz, A.M.; Au, S.C.; Kim, N.; Gottlieb, A.B.; Lizzul, P.F. NF-κB: An essential transcription factor in psoriasis. J. Dermatol. Sci. 2013, 69, 89–94. [Google Scholar] [CrossRef]
- Xu, F.W.; Lv, Y.L.; Zhong, Y.F.; Xue, Y.N.; Wang, Y.; Zhang, L.Y.; Hu, X.; Tan, W.Q. Beneficial Effects of Green Tea EGCG on Skin Wound Healing: A Comprehensive Review. Molecules 2021, 26, 6123. [Google Scholar] [CrossRef]
- Tortolani, D.; Di Meo, C.; Standoli, S.; Ciaramellano, F.; Kadhim, S.; Hsu, E.; Rapino, C.; Maccarrone, M. Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 2721. [Google Scholar] [CrossRef]
- Di Meo, C.; Tortolani, D.; Standoli, S.; Angelucci, C.B.; Fanti, F.; Leuti, A.; Sergi, M.; Kadhim, S.; Hsu, E.; Rapino, C.; et al. Effects of Rare Phytocannabinoids on the Endocannabinoid System of Human Keratinocytes. Int. J. Mol. Sci. 2022, 23, 5430. [Google Scholar] [CrossRef]
- Gaffal, E.; Cron, M.; Glodde, N.; Tüting, T. Anti-inflammatory activity of topical THC in DNFB-mediated mouse allergic contact dermatitis independent of CB1 and CB2 receptors. Allergy 2013, 68, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.C.; Jeffery, C.S.; Sandhu, Z.; Brownlee, B.P.; Queimado, L.; Mims, M.M. The effect of cannabinoids on wound healing: A review. Health Sci. Rep. 2024, 7, e1908. [Google Scholar] [CrossRef]
- Simei, J.L.Q.; Souza, J.D.R.; Lisboa, J.R.; Campos, A.C.; Guimaraes, F.S.; Zuardi, A.; Crippa, J.A.S. Does the “Entourage Effect” in Cannabinoids Exist? A Narrative Scoping Review. Cannabis Cannabinoid Res. 2023, 15, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Al-Khazaleh, A.K.; Zhou, X.; Bhuyan, D.J.; Münch, G.W.; Al-Dalabeeh, E.A.; Jaye, K.; Chang, D.N. The Neurotherapeutic Arsenal in Cannabis sativa: Insights into Anti-Neuroinflammatory and Neuroprotective Activity and Potential Entourage Effects. Molecules 2024, 29, 26. [Google Scholar] [CrossRef] [PubMed]
- Toth, K.F.; Adam, D.; Biro, T.; Olah, A. Cannabinoid Signaling in the Skin: Therapeutic Potential of the “C(ut)annabinoid” System. Molecules 2019, 24, 918. [Google Scholar] [CrossRef]
- Filipiuc, S.I.; Neagu, A.N.; Uritu, C.M.; Tamba, B.I.; Filipiuc, L.E.; Tudorancea, I.M.; Boca, A.N.; Hâncu, M.F.; Porumb, V.; Bild, W. The Skin and Natural Cannabinoids-Topical and Transdermal Applications. Pharmaceuticals 2023, 16, 1049. [Google Scholar] [CrossRef] [PubMed]
- Stoeckle, M.Y. Post-transcriptional regulation of groα, β, γ, and IL-8 mRNAs by IL-1β. Nucleic Acids Res. 1991, 19, 917–920. [Google Scholar] [CrossRef]
- Kim, S.; You, D.; Jeong, Y.; Yu, J.; Kim, S.W.; Nam, S.J.; Lee, J.E. Berberine down-regulates IL-8 expression through inhibition of the EGFR/MEK/ERK pathway in triple-negative breast cancer cells. Phytomedicine 2018, 50, 43–49. [Google Scholar] [CrossRef]
- Tanaka, C.; Kamata, H.; Takeshita, H.; Yagisawa, H.; Hirata, H. Redox regulation of lipopolysaccharide (LPS)-induced interleukin-8 (IL-8) gene expression mediated by NF kappa B and AP-1 in human astrocytoma U373 cells. Biochem. Biophys. Res. Commun. 1997, 232, 568–573. [Google Scholar] [CrossRef]
- Chaly, Y.V.; Selvan, R.S.; Fegeding, K.V.; Kolesnikova, T.S.; Voitenok, N.N. Expression of IL-8 gene in human monocytes and lymphocytes: Differential regulation by TNF and IL-1. Cytokine 2000, 12, 636–643. [Google Scholar] [CrossRef]
- Caroli, C.; Baron, G.; Cappellucci, G.; Brighenti, V.; Della Vedova, L.; Fraulini, F.; Oliaro-Bosso, S.; Alessandrini, A.; Zambon, A.; Lusvardi, G.; et al. Extraction, purification and in vitro assessment of the antioxidant and anti-inflammatory activity of policosanols from non-psychoactive Cannabis sativa L. Heliyon 2024, 10, e30291. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival: Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cannabinoids (%) | Canniprene (%) | Terpenes (%) | Cannflavins (%) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Polarity | Fraction | Weight (g) | Yield (%) | CBD | CBDA | CBDV | CBDVA | CBDC4 | THCV | CBG | CBGA | CBC | THC | THCA | Total | Total | Cannflavin A | Cannflavin B |
| low | Fraction a | 19.37 | 6.46 | 28.0 | 0 | 0.3 | 0.1 | 0 | 0 | 0.7 | 0.1 | 1.0 | 1.0 | 0.1 | 0.4 | 1.8 | 0.1 | 0 |
| ↓ | Fraction b | 6.73 | 2.40 | 13.0 | 0 | 0.3 | 0.2 | 0 | 0 | 1.0 | 0.1 | 0.6 | 0 | 0 | 0.3 | 0.2 | 0.4 | 0.3 |
| ↓ | Fraction c | 9.69 | 3.54 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 |
| high | Fraction d | 13.72 | 5.19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.2 | 0 | 0 |
| Cannabinoids (%) | Canniprene (%) | Cannflavins (%) | ||||||||||||
| Batch Number | CBD | CBDA | CBDV | CBDVA | CBDC4 | THCV | CBG | CBGA | CBC | THC | THCA | Total | Cannflavin A | Cannflavin B |
| CM5 74719004 | 4.836 | 0.02 | 0.02 | n.d. | 0.02 | n.d. | 0.14 | n.d. | 0.24 | 0.22 | n.d. | 0.0165 | 0.011 | 0.004 |
| Terpenes (%) | ||||||||||||||
| Batch Number | γ-terpineol | β-caryophyllene | α-humulene | Farnesene isomer 1 | Farnesene isomer 2 | Farnesene isomer 3 | Caryophillene oxide | α-bisabolol | Total | |||||
| CM5 74719004 | 0.031 | 0.050 | 0.012 | 0.010 | 0.007 | 0.015 | 0.015 | 0.011 | 0.151 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fumagalli, M.; Martinelli, G.; Paladino, G.; Rossini, N.; Ciriello, U.; Nicolaci, V.; Maranta, N.; Pozzoli, C.; El Haddad, S.M.; Sonzogni, E.; et al. Beyond Cannabidiol: The Contribution of Cannabis sativa Phytocomplex to Skin Anti-Inflammatory Activity in Human Skin Keratinocytes. Pharmaceuticals 2025, 18, 647. https://doi.org/10.3390/ph18050647
Fumagalli M, Martinelli G, Paladino G, Rossini N, Ciriello U, Nicolaci V, Maranta N, Pozzoli C, El Haddad SM, Sonzogni E, et al. Beyond Cannabidiol: The Contribution of Cannabis sativa Phytocomplex to Skin Anti-Inflammatory Activity in Human Skin Keratinocytes. Pharmaceuticals. 2025; 18(5):647. https://doi.org/10.3390/ph18050647
Chicago/Turabian StyleFumagalli, Marco, Giulia Martinelli, Giuseppe Paladino, Nora Rossini, Umberto Ciriello, Vincenzo Nicolaci, Nicole Maranta, Carola Pozzoli, Safwa Moheb El Haddad, Elisa Sonzogni, and et al. 2025. "Beyond Cannabidiol: The Contribution of Cannabis sativa Phytocomplex to Skin Anti-Inflammatory Activity in Human Skin Keratinocytes" Pharmaceuticals 18, no. 5: 647. https://doi.org/10.3390/ph18050647
APA StyleFumagalli, M., Martinelli, G., Paladino, G., Rossini, N., Ciriello, U., Nicolaci, V., Maranta, N., Pozzoli, C., El Haddad, S. M., Sonzogni, E., Dell’Agli, M., Piazza, S., & Sangiovanni, E. (2025). Beyond Cannabidiol: The Contribution of Cannabis sativa Phytocomplex to Skin Anti-Inflammatory Activity in Human Skin Keratinocytes. Pharmaceuticals, 18(5), 647. https://doi.org/10.3390/ph18050647