Mechanistic and Molecular Insights into Empagliflozin’s Role in Ferroptosis and Inflammation Trajectories in Acetaminophen-Induced Hepatotoxicity

 , , and
, , and
Abstract
1. Introduction
2. Results
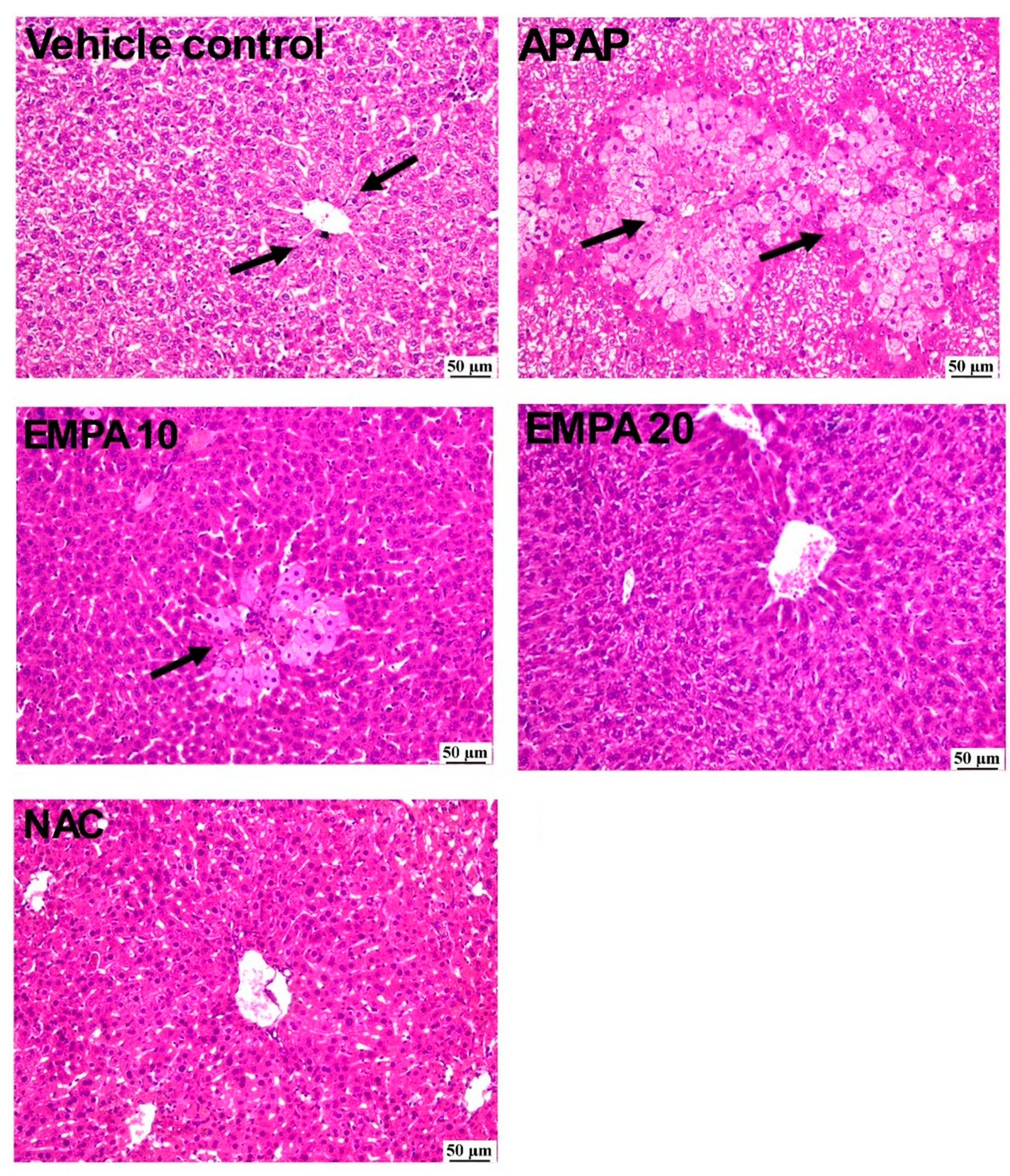
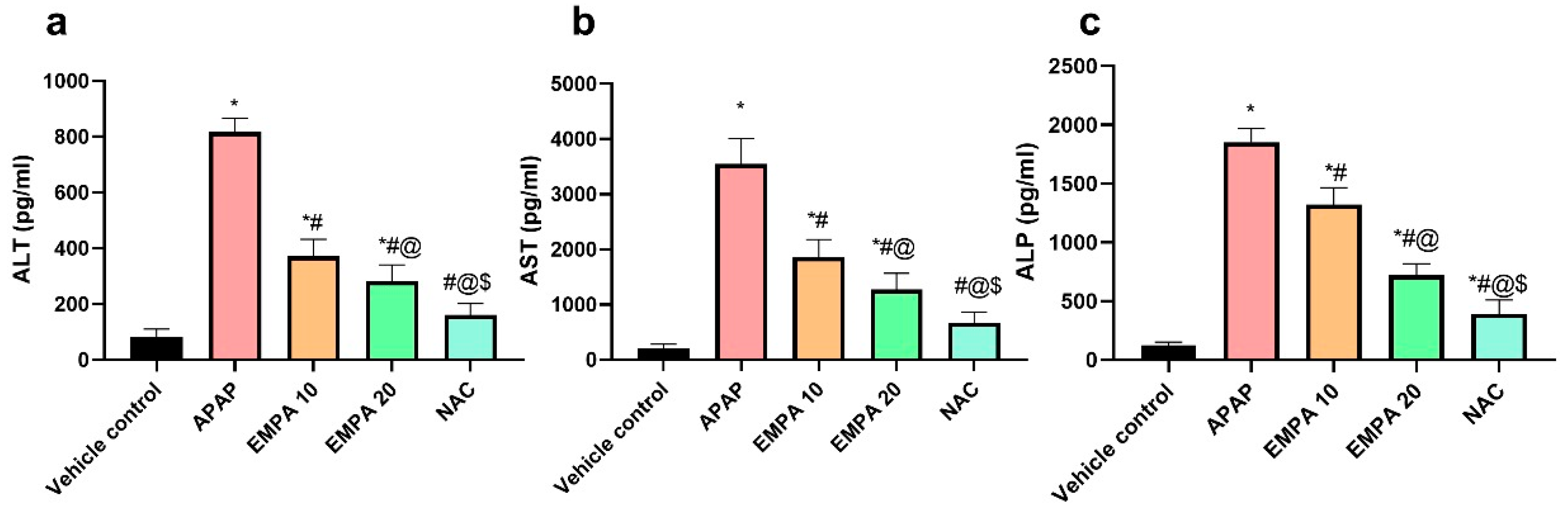
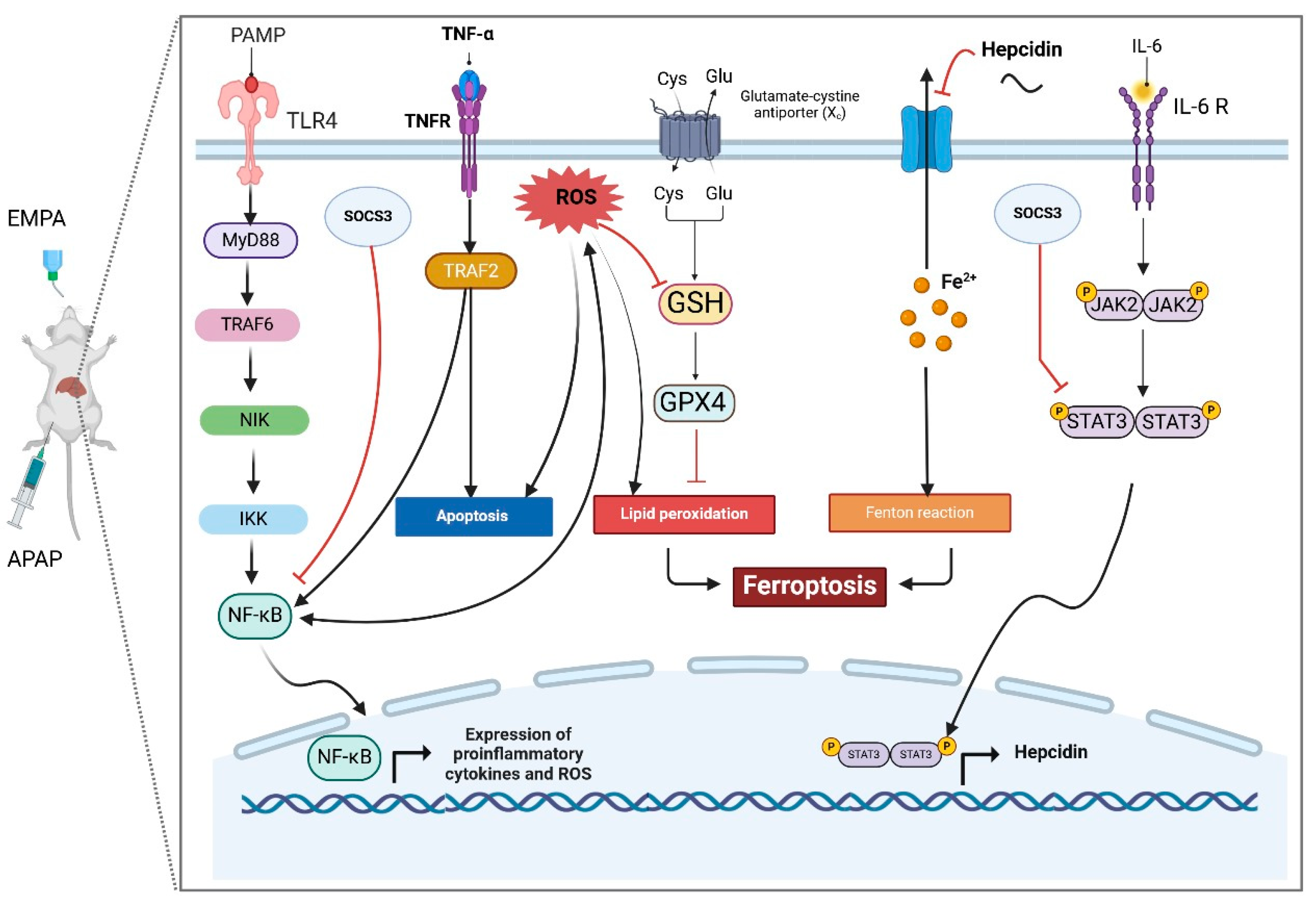
2.1. EMPA Pretreatment Alleviated Liver Injury Induced by APAP
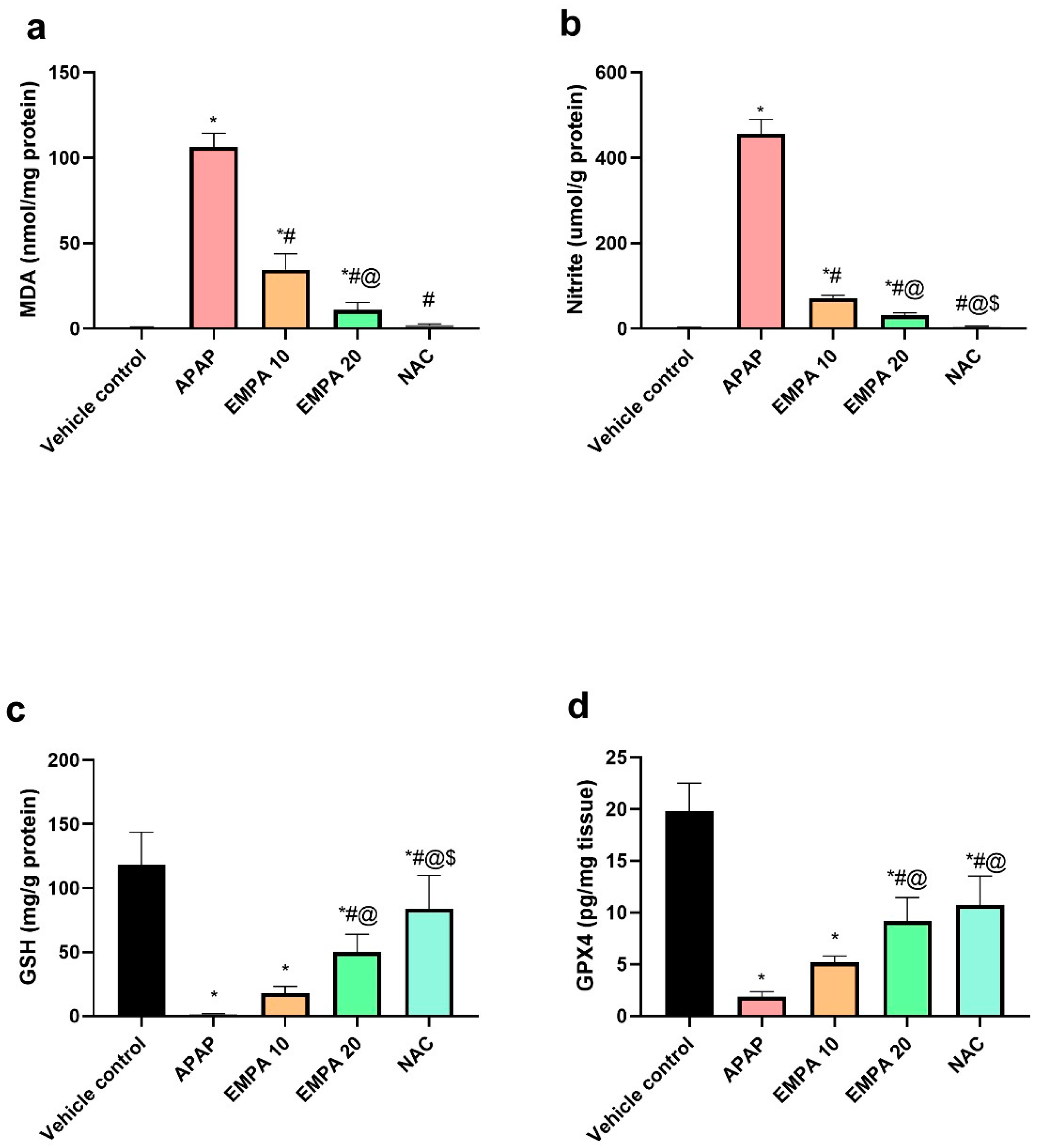
2.2. EMPA Pretreatment Restored the Imbalanced Redox State Induced by APAP
2.3. EMPA Pretreatment Counteracted Hepatic Inflammation Induced by APAP
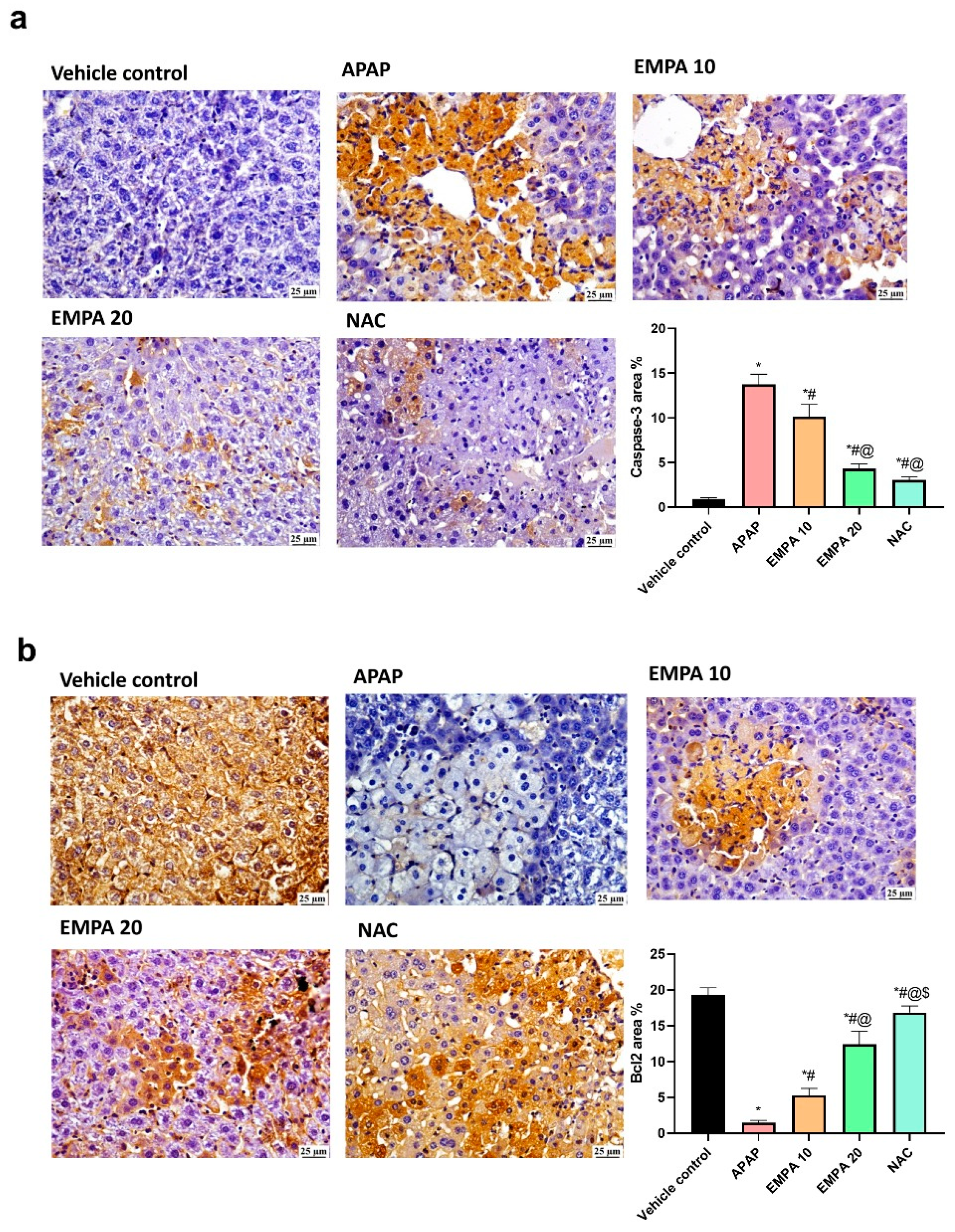
2.4. EMPA Pretreatment Reduced Apoptotic Changes in Liver Induced by APAP
2.5. EMPA Pretreatment Diminished Liver Iron Deposition-Evoked Ferroptosis Induced by APAP
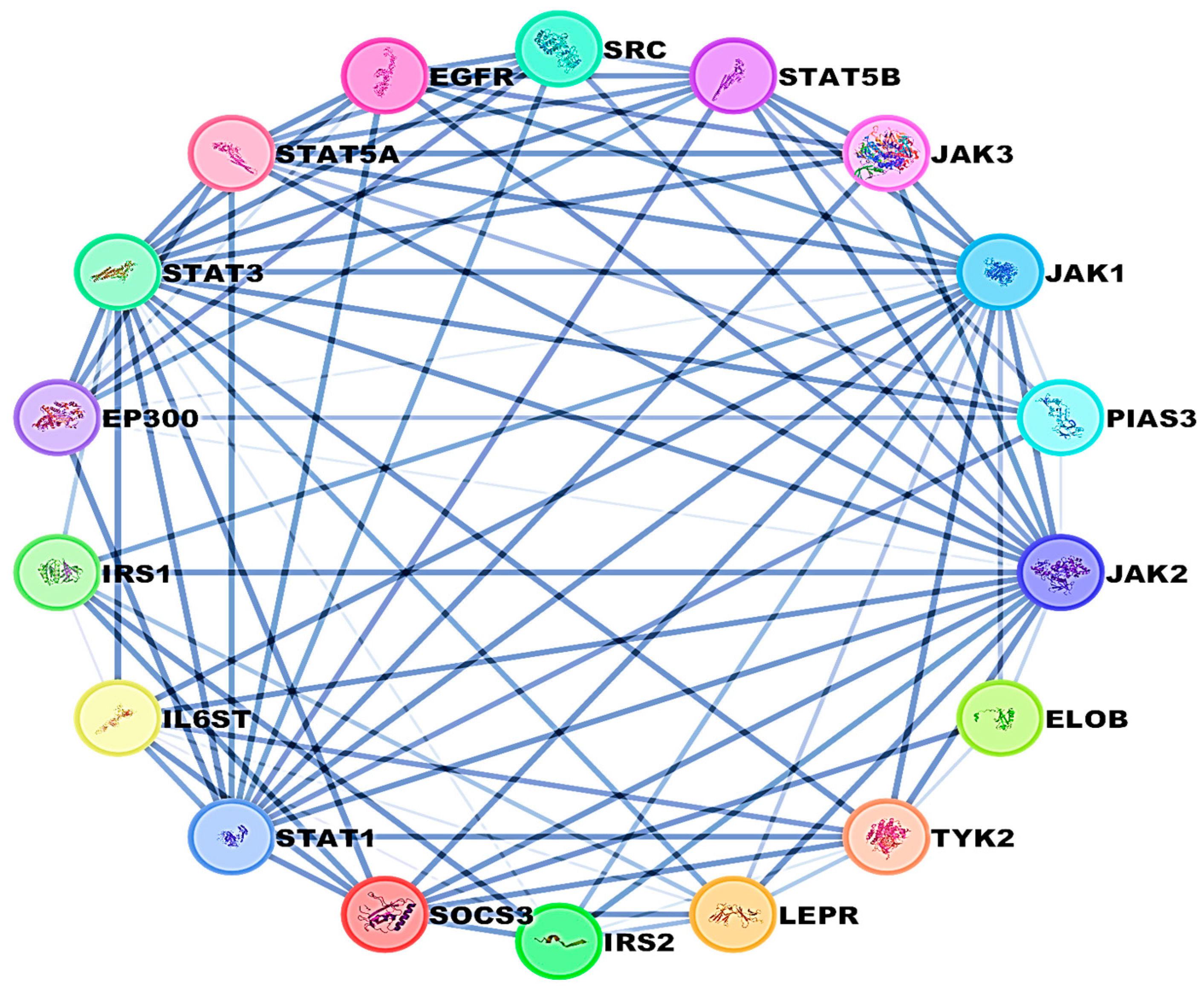
2.6. Protein–Protein Interaction (PPI) Network
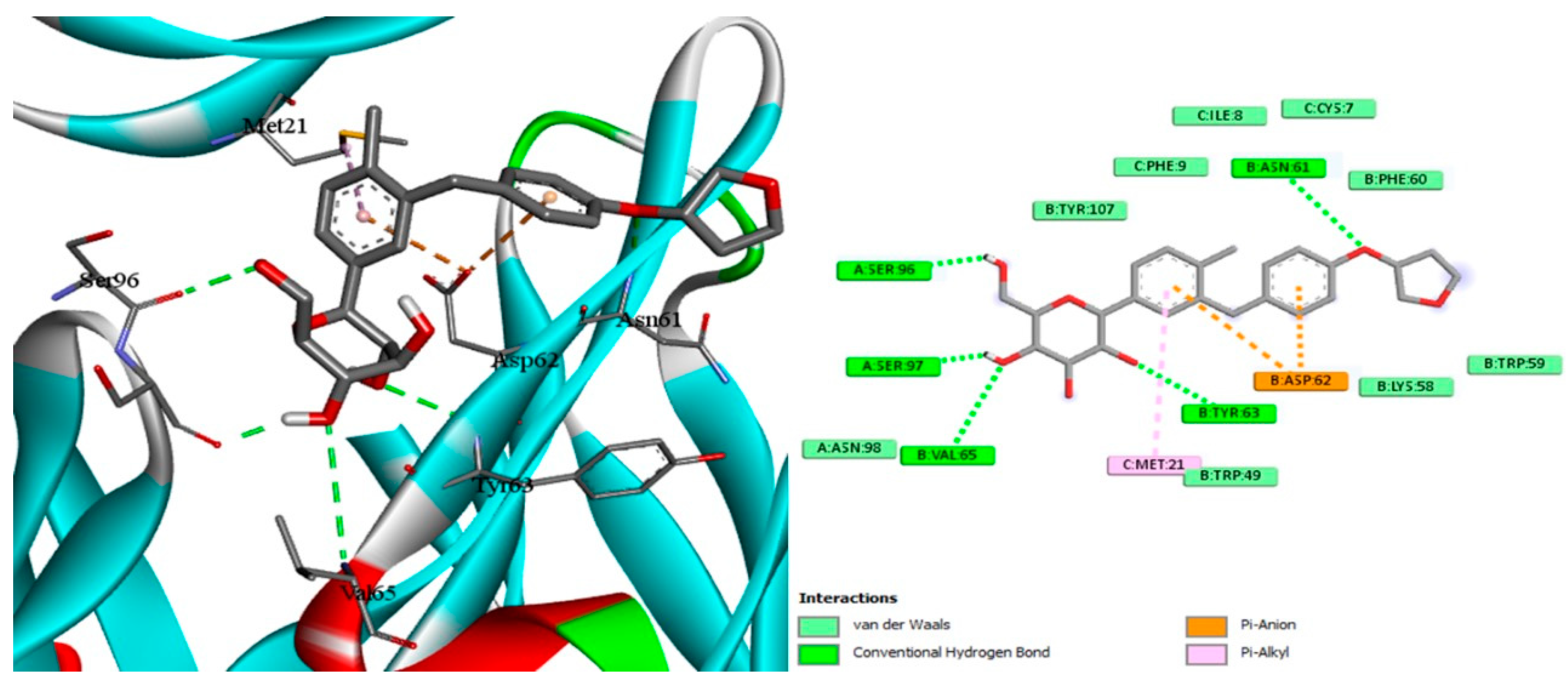
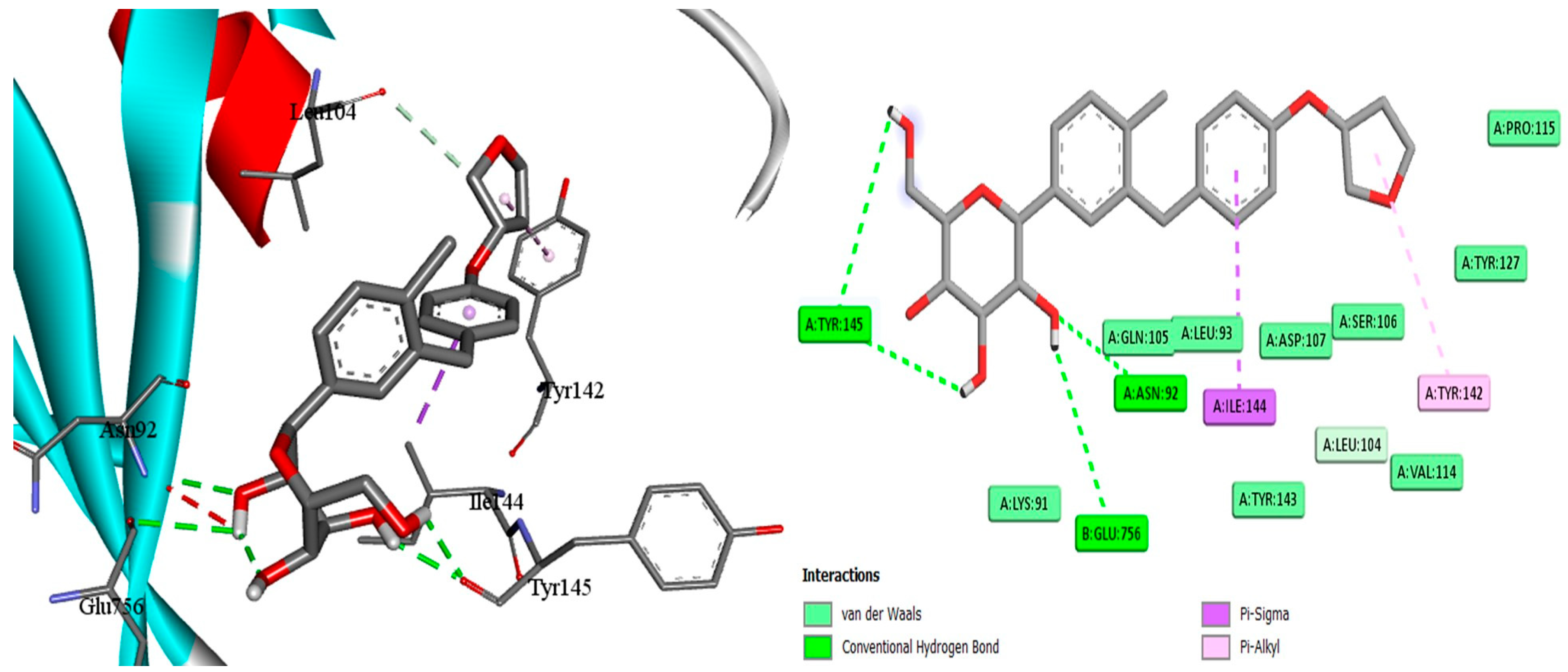
2.7. Docking of EMPA
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Animals
4.3. Study Design
4.4. Collection of the Samples
4.5. Histopathology and Lesion Score
4.6. Immunohistochemistry
4.7. Determination of Liver Toxicity Parameters
4.8. Determination of Oxidative Stress Markers
4.9. Determination of Inflammatory Markers
4.10. Determination of STAT3 by Western Blot Assay
4.11. Determination of Ferroptosis-Related Biomarkers
4.12. PPI Network Analysis
4.13. Experimental Docking
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernal, W.; Auzinger, G.; Dhawan, A.; Wendon, J. Acute Liver Failure. Lancet 2010, 376, 190–201. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R.; Jaeschke, H. Metabolism and Disposition of Acetaminophen: Recent Advances in Relation to Hepatotoxicity and Diagnosis. Pharm. Res. 2013, 30, 2174–2187. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, J.; Kang, R.; Klionsky, D.J.; Tang, D. Ferroptosis: Machinery and Regulation. Autophagy 2021, 17, 2054–2081. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yu, C.; Kang, R.; Tang, D. Iron Metabolism in Ferroptosis. Front. Cell Dev. Biol. 2020, 8, 590226. [Google Scholar] [CrossRef]
- Rochette, L.; Dogon, G.; Rigal, E.; Zeller, M.; Cottin, Y.; Vergely, C. Lipid Peroxidation and Iron Metabolism: Two Corner Stones in the Homeostasis Control of Ferroptosis. Int. J. Mol. Sci. 2022, 24, 449. [Google Scholar] [CrossRef]
- Zhang, H.; Ostrowski, R.; Jiang, D.; Zhao, Q.; Liang, Y.; Che, X.; Zhao, J.; Xiang, X.; Qin, W.; He, Z. Hepcidin Promoted Ferroptosis through Iron Metabolism Which Is Associated with DMT1 Signaling Activation in Early Brain Injury Following Subarachnoid Hemorrhage. Oxid. Med. Cell. Longev. 2021, 2021, 9800794. [Google Scholar] [CrossRef]
- Yang, L.; Wang, H.; Yang, X.; Wu, Q.; An, P.; Jin, X.; Liu, W.; Huang, X.; Li, Y.; Yan, S. Auranofin Mitigates Systemic Iron Overload and Induces Ferroptosis via Distinct Mechanisms. Signal Transduct. Target. Ther. 2020, 5, 138. [Google Scholar] [CrossRef]
- Wu, J.; Xue, R.; Wu, M.; Yin, X.; Xie, B.; Meng, Q. Nrf2-Mediated Ferroptosis Inhibition Exerts a Protective Effect on Acute-on-Chronic Liver Failure. Oxid. Med. Cell. Longev. 2022, 2022, 4505513. [Google Scholar] [CrossRef]
- Ye, J.; Peng, J.; Liu, K.; Zhang, T.; Huang, W. MCTR1 Inhibits Ferroptosis by Promoting NRF2 Expression to Attenuate Hepatic Ischemia-Reperfusion Injury. Am. J. Physiol.-Gastrointest. Liver Physiol. 2022, 323, G283–G293. [Google Scholar] [CrossRef]
- Li, H.; Weng, Q.; Gong, S.; Zhang, W.; Wang, J.; Huang, Y.; Li, Y.; Guo, J.; Lan, T. Kaempferol Prevents Acetaminophen-Induced Liver Injury by Suppressing Hepatocyte Ferroptosis via Nrf2 Pathway Activation. Food Funct. 2023, 14, 1884–1896. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Karasawa, T.; Kimura, H.; Watanabe, S.; Komada, T.; Kamata, R.; Sampilvanjil, A.; Ito, J.; Nakagawa, K.; Kuwata, H. Ferroptosis Driven by Radical Oxidation of N-6 Polyunsaturated Fatty Acids Mediates Acetaminophen-Induced Acute Liver Failure. Cell Death Dis. 2020, 11, 144. [Google Scholar] [CrossRef]
- Tao, J.; Xue, C.; Wang, X.; Chen, H.; Liu, Q.; Jiang, C.; Zhang, W. GAS1 Promotes Ferroptosis of Liver Cells in Acetaminophen-Induced Acute Liver Failure. Int. J. Med. Sci. 2023, 20, 1616. [Google Scholar] [CrossRef] [PubMed]
- Novikov, A.; Vallon, V. Sodium Glucose Cotransporter 2 Inhibition in the Diabetic Kidney: An Update. Curr. Opin. Nephrol. Hypertens. 2016, 25, 50–58. [Google Scholar] [CrossRef]
- Ndefo, U.A.; Anidiobi, N.O.; Basheer, E.; Eaton, A.T. Empagliflozin (Jardiance): A Novel SGLT2 Inhibitor for the Treatment of Type-2 Diabetes. Pharm. Ther. 2015, 40, 364. [Google Scholar]
- Lahnwong, C.; Chattipakorn, S.C.; Chattipakorn, N. Potential Mechanisms Responsible for Cardioprotective Effects of Sodium–Glucose Co-Transporter 2 Inhibitors. Cardiovasc. Diabetol. 2018, 17, 101. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Butler, A.E.; Atkin, S.L.; Katsiki, N.; Sahebkar, A. Sodium–Glucose Cotransporter 2 Inhibitors and Inflammation in Chronic Kidney Disease: Possible Molecular Pathways. J. Cell. Physiol. 2019, 234, 223–230. [Google Scholar] [CrossRef]
- Perrone-Filardi, P.; Avogaro, A.; Bonora, E.; Colivicchi, F.; Fioretto, P.; Maggioni, A.P.; Sesti, G.; Ferrannini, E. Mechanisms Linking Empagliflozin to Cardiovascular and Renal Protection. Int. J. Cardiol. 2017, 241, 450–456. [Google Scholar] [CrossRef]
- Taheri, H.; Malek, M.; Ismail-Beigi, F.; Zamani, F.; Sohrabi, M.; Reza Babaei, M.; Khamseh, M.E. Effect of Empagliflozin on Liver Steatosis and Fibrosis in Patients with Non-Alcoholic Fatty Liver Disease without Diabetes: A Randomized, Double-Blind, Placebo-Controlled Trial. Adv. Ther. 2020, 37, 4697–4708. [Google Scholar] [CrossRef]
- ElBaset, M.A.; Salem, R.S.; Ayman, F.; Ayman, N.; Shaban, N.; Afifi, S.M.; Esatbeyoglu, T.; Abdelaziz, M.; Elalfy, Z.S. Effect of Empagliflozin on Thioacetamide-Induced Liver Injury in Rats: Role of AMPK/SIRT-1/HIF-1α Pathway in Halting Liver Fibrosis. Antioxidants 2022, 11, 2152. [Google Scholar] [CrossRef]
- Abdelhamid, A.M.; Elsheakh, A.R.; Abdelaziz, R.R.; Suddek, G.M. Empagliflozin Ameliorates Ethanol-Induced Liver Injury by Modulating NF-ΚB/Nrf-2/PPAR-γ Interplay in Mice. Life Sci. 2020, 256, 117908. [Google Scholar] [CrossRef] [PubMed]
- Kheradpezhouh, E.; Panjehshahin, M.-R.; Miri, R.; Javidnia, K.; Noorafshan, A.; Monabati, A.; Dehpour, A.-R. Curcumin Protects Rats against Acetaminophen-Induced Hepatorenal Damages and Shows Synergistic Activity with N-Acetyl Cysteine. Eur. J. Pharmacol. 2010, 628, 274–281. [Google Scholar] [CrossRef]
- Mohamed Kamel, G.A.; Harahsheh, E.; Hussein, S. Diacerein Ameliorates Acetaminophen Hepatotoxicity in Rats via Inhibiting HMGB1/TLR4/NF-ΚB and Upregulating PPAR-γ Signal. Mol. Biol. Rep. 2022, 49, 5863–5874. [Google Scholar] [CrossRef]
- Kalantari, E.; Zolbanin, N.M.; Ghasemnejad-Berenji, M. Protective Effects of Empagliflozin on Methotrexate Induced Hepatotoxicity in Rats. Biomed. Pharmacother. 2024, 170, 115953. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Wu, A.; Bing, Y.; Liu, Y.; Luo, J.; Yan, H.; Zheng, P.; Yu, J.; Chen, D. Retinoic Acid Protects against Lipopolysaccharide-Induced Ferroptotic Liver Injury and Iron Disorders by Regulating Nrf2/HO-1 and RARβ Signaling. Free Radic. Biol. Med. 2023, 205, 202–213. [Google Scholar] [CrossRef]
- Tong, J.; Lan, X.; Zhang, Z.; Liu, Y.; Sun, D.; Wang, X.; Ou-Yang, S.; Zhuang, C.; Shen, F.; Wang, P. Ferroptosis Inhibitor Liproxstatin-1 Alleviates Metabolic Dysfunction-Associated Fatty Liver Disease in Mice: Potential Involvement of PANoptosis. Acta Pharmacol. Sin. 2023, 44, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M.; Abdel-Halim, N.H.M.; El-Shenbaby, I.; Helmy, M.A.; Hammad, M.O.; Habotta, O.A.; El Nashar, E.M.; Alghamdi, M.A.; Aldahhan, R.A.; Al-Khater, K.M. Phytic Acid Attenuates Acetaminophen-Induced Hepatotoxicity via Modulating Iron-Mediated Oxidative Stress and SIRT-1 Expression in Mice. Front. Pharmacol. 2024, 15, 1384834. [Google Scholar] [CrossRef]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, Biology and Role in Disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Liao, Z.; Tong, B.; Ou, Z.; Wei, J.; Lei, M.; Yang, C. The Role of Extracellular Vesicles in Iron Homeostasis and Ferroptosis: Focus on Musculoskeletal Diseases. Traffic 2023, 24, 384–396. [Google Scholar] [CrossRef]
- Stockwell, B.R. Ferroptosis Turns 10: Emerging Mechanisms, Physiological Functions, and Therapeutic Applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef]
- Bu, T.; Wang, C.; Meng, Q.; Huo, X.; Sun, H.; Sun, P.; Zheng, S.; Ma, X.; Liu, Z.; Liu, K. Hepatoprotective Effect of Rhein against Methotrexate-Induced Liver Toxicity. Eur. J. Pharmacol. 2018, 834, 266–273. [Google Scholar] [CrossRef]
- Ahmad, M.M.; Rezk, N.A.; Fawzy, A.; Sabry, M. Protective Effects of Curcumin and Silymarin against Paracetamol Induced Hepatotoxicity in Adult Male Albino Rats. Gene 2019, 712, 143966. [Google Scholar] [CrossRef] [PubMed]
- Iannantuoni, F.; de Marañon, A.M.; Diaz-Morales, N.; Falcon, R.; Bañuls, C.; Abad-Jimenez, Z.; Victor, V.M.; Hernandez-Mijares, A.; Rovira-Llopis, S. The SGLT2 Inhibitor Empagliflozin Ameliorates the Inflammatory Profile in Type 2 Diabetic Patients and Promotes an Antioxidant Response in Leukocytes. J. Clin. Med. 2019, 8, 1814. [Google Scholar] [PubMed]
- Wang, Y.; Ding, Y.; Sun, P.; Zhang, W.; Xin, Q.; Wang, N.; Niu, Y.; Chen, Y.; Luo, J.; Lu, J. Empagliflozin-Enhanced Antioxidant Defense Attenuates Lipotoxicity and Protects Hepatocytes by Promoting FoxO3a-and Nrf2-Mediated Nuclear Translocation via the CAMKK2/AMPK Pathway. Antioxidants 2022, 11, 799. [Google Scholar] [CrossRef]
- Li, X.; Wang, M.; Kalina, J.-O.; Preckel, B.; Hollmann, M.W.; Albrecht, M.; Zuurbier, C.J.; Weber, N.C. Empagliflozin Prevents Oxidative Stress in Human Coronary Artery Endothelial Cells via the NHE/PKC/NOX Axis. Redox Biol. 2024, 69, 102979. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Yang, L.; Xiao, J.-J.; Liu, Q.; Ni, L.; Hu, J.-W.; Yu, H.; Wu, X.; Zhang, B.-F. Empagliflozin Attenuates the Renal Tubular Ferroptosis in Diabetic Kidney Disease through AMPK/NRF2 Pathway. Free Radic. Biol. Med. 2023, 195, 89–102. [Google Scholar] [CrossRef]
- Du, Z.; Ma, Z.; Lai, S.; Ding, Q.; Hu, Z.; Yang, W.; Qian, Q.; Zhu, L.; Dou, X.; Li, S. Atractylenolide I Ameliorates Acetaminophen-Induced Acute Liver Injury via the TLR4/MAPKs/NF-ΚB Signaling Pathways. Front. Pharmacol. 2022, 13, 797499. [Google Scholar] [CrossRef]
- Du, Y.-C.; Lai, L.; Zhang, H.; Zhong, F.-R.; Cheng, H.-L.; Qian, B.-L.; Tan, P.; Xia, X.-M.; Fu, W.-G. Kaempferol from Penthorum Chinense Pursh Suppresses HMGB1/TLR4/NF-ΚB Signaling and NLRP3 Inflammasome Activation in Acetaminophen-Induced Hepatotoxicity. Food Funct. 2020, 11, 7925–7934. [Google Scholar] [CrossRef]
- Saad, M.A.; El-Sahhar, A.E.; Arab, H.H.; Al-Shorbagy, M.Y. Nicorandil Abates Arthritic Perturbations Induced by Complete Freund’s Adjuvant in Rats via Conquering TLR4-MyD88-TRAF6 Signaling Pathway. Life Sci. 2019, 218, 284–291. [Google Scholar] [CrossRef]
- Du, H.; Tong, S.; Kuang, G.; Gong, X.; Jiang, N.; Yang, X.; Liu, H.; Li, N.; Xie, Y.; Xiang, Y. Sesamin Protects against APAP-Induced Acute Liver Injury by Inhibiting Oxidative Stress and Inflammatory Response via Deactivation of HMGB1/TLR4/NFκB Signal in Mice. J. Immunol. Res. 2023, 2023, 1116841. [Google Scholar] [CrossRef]
- Lingappan, K. NF-ΚB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Zhang, Z.; Shen, H.; Xiong, Y.; Shah, Y.M.; Liu, Y.; Fan, X.; Rui, L. Hepatic NF-κB-Inducing Kinase and Inhibitor of NF-κB Kinase Subunit α Promote Liver Oxidative Stress, Ferroptosis, and Liver Injury. Hepatol. Commun. 2021, 5, 1704–1720. [Google Scholar] [CrossRef] [PubMed]
- Salama, S.A.; Abdel-Bakky, M.S.; Mohamed, A.A. Upregulation of Nrf2 Signaling and Suppression of Ferroptosis and NF-ΚB Pathway by Leonurine Attenuate Iron Overload-Induced Hepatotoxicity. Chem. Biol. Interact. 2022, 356, 109875. [Google Scholar] [CrossRef]
- Chen, Y.; Fang, Z.M.; Yi, X.; Wei, X.; Jiang, D.S. The Interaction between Ferroptosis and Inflammatory Signaling Pathways. Cell Death Dis. 2023, 14, 205. [Google Scholar] [CrossRef]
- Zaghloul, M.S.; Elshal, M.; Abdelmageed, M.E. Preventive Empagliflozin Activity on Acute Acetic Acid-Induced Ulcerative Colitis in Rats via Modulation of SIRT-1/PI3K/AKT Pathway and Improving Colon Barrier. Environ. Toxicol. Pharmacol. 2022, 91, 103833. [Google Scholar] [CrossRef]
- Shao, Q.; Meng, L.; Lee, S.; Tse, G.; Gong, M.; Zhang, Z.; Zhao, J.; Zhao, Y.; Li, G.; Liu, T. Empagliflozin, a Sodium Glucose Co-Transporter-2 Inhibitor, Alleviates Atrial Remodeling and Improves Mitochondrial Function in High-Fat Diet/Streptozotocin-Induced Diabetic Rats. Cardiovasc. Diabetol. 2019, 18, 165. [Google Scholar] [CrossRef]
- Rofaeil, R.R.; Welson, N.N.; Fawzy, M.A.; Ahmed, A.F.; Atta, M.; Bahaa El-deen, M.A.; Abdelzaher, W.Y. The IL-6/HO-1/STAT3 Signaling Pathway Is Implicated in the Amelioration of Acetaminophen-Induced Hepatic Toxicity: A Neonatal Rat Model. Hum. Exp. Toxicol. 2023, 42, 09603271231151376. [Google Scholar] [CrossRef]
- Zai, W.; Chen, W.; Luan, J.; Fan, J.; Zhang, X.; Wu, Z.; Ding, T.; Ju, D.; Liu, H. Dihydroquercetin Ameliorated Acetaminophen-Induced Hepatic Cytotoxicity via Activating JAK2/STAT3 Pathway and Autophagy. Appl. Microbiol. Biotechnol. 2018, 102, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Kowdley, K.V.; Gochanour, E.M.; Sundaram, V.; Shah, R.A.; Handa, P. Hepcidin Signaling in Health and Disease: Ironing out the Details. Hepatol. Commun. 2021, 5, 723–735. [Google Scholar] [CrossRef]
- Poli, M.; Girelli, D.; Campostrini, N.; Maccarinelli, F.; Finazzi, D.; Luscieti, S.; Nai, A.; Arosio, P. Heparin: A Potent Inhibitor of Hepcidin Expression in Vitro and in Vivo. Blood J. Am. Soc. Hematol. 2011, 117, 997–1004. [Google Scholar] [CrossRef]
- Meynard, D.; Babitt, J.L.; Lin, H.Y. The Liver: Conductor of Systemic Iron Balance. Blood J. Am. Soc. Hematol. 2014, 123, 168–176. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, J.; Ma, H.; Han, Y.; Xu, W.; Wang, J.; Cai, Y.; Jia, X.; Jia, Q.; Yang, Q. High Hepcidin Levels Promote Abnormal Iron Metabolism and Ferroptosis in Chronic Atrophic Gastritis. Biomedicines 2023, 11, 2338. [Google Scholar] [CrossRef]
- Bao, W.-D.; Fan, Y.; Deng, Y.-Z.; Long, L.-Y.; Wang, J.; Guan, D.-X.; Qian, Z.-Y.; An, P.; Feng, Y.-Y.; He, Z.-Y. Iron Overload in Hereditary Tyrosinemia Type 1 Induces Liver Injury through the Sp1/Tfr2/Hepcidin Axis. J. Hepatol. 2016, 65, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Shi, Y.; Li, X.; Jin, N.; Zhang, M.; Liu, Z.; Liang, Y.; Xie, J. Human Umbilical Cord Mesenchymal Stem Cells Protect against Ferroptosis in Acute Liver Failure through the IGF1-Hepcidin-FPN1 Axis and Inhibiting Iron Loading: Mesenchymal Stem Cells Protect against Ferroptosis in Acute Liver Failure. Acta Biochim. Biophys. Sin. 2024, 56, 280. [Google Scholar]
- Bloomer, S.A.; Brown, K.E. Hepcidin and Iron Metabolism in Experimental Liver Injury. Am. J. Pathol. 2021, 191, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Yang, Y.; Wu, K.; Zhao, T.; Shi, Y.; Song, M.; Li, J. The Effects of Dandelion Polysaccharides on Iron Metabolism by Regulating Hepcidin via JAK/STAT Signaling Pathway. Oxid. Med. Cell. Longev. 2021, 2021, 7184760. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, Y.; Li, B.; Xie, J.; Ma, Z.; Thirupathi, A.; Yu, P.; Gao, G.; Shi, M.; Zhou, C. Propofol Prevents Oxidative Stress and Apoptosis by Regulating Iron Homeostasis and Targeting JAK/STAT3 Signaling in SH-SY5Y Cells. Brain Res. Bull. 2019, 153, 191–201. [Google Scholar] [CrossRef]
- Lee, N.; Heo, Y.J.; Choi, S.-E.; Jeon, J.Y.; Han, S.J.; Kim, D.J.; Kang, Y.; Lee, K.W.; Kim, H.J. Anti-inflammatory Effects of Empagliflozin and Gemigliptin on LPS-stimulated Macrophage via the IKK/NF-κB, MKK7/JNK, and JAK2/STAT1 Signalling Pathways. J. Immunol. Res. 2021, 2021, 9944880. [Google Scholar] [CrossRef]
- Sheng, W.; Yu, J.; Zhang, H.; Zhang, J. Empagliflozin Attenuates Inflammation Levels in Autoimmune Myocarditis through the STAT3 Pathway and Macrophage Phenotype Transformation. Mol. Immunol. 2024, 167, 43–52. [Google Scholar] [CrossRef]
- Ali, R.B.; Ahmed, M.H.; Ibrahim, H.K.; Mahmood, H.S. Tracking Hepcidin Level in Induced Type 2 Diabetic Rats and How Empagliflozin Affects Its Level. J. Popul. Ther. Clin. Pharmacol. 2022, 29, 158–166. [Google Scholar]
- Ibrahim, S.M.; Al-Shorbagy, M.Y.; Abdallah, D.M.; El-Abhar, H.S. Activation of A7 Nicotinic Acetylcholine Receptor Ameliorates Zymosan-Induced Acute Kidney Injury in BALB/c Mice. Sci. Rep. 2018, 8, 16814. [Google Scholar] [CrossRef]
- Santana, F.P.R.; Tomari, S.F.; de Pontes Miranda, C.J.C.; Pinheiro, N.M.; Caperuto, L.C.; de Fátima Lopes Calvo Tibério, I.; Prado, M.A.M.; de Arruda Martins, M.; Prado, V.F.; Prado, C.M. Nicotinic Alpha-7 Receptor Stimulation (A7nAChR) Inhibited NF-KB/STAT3/SOCS3 Pathways in a Murine Model of Asthma. Eur. Respir. J. 2017, 50 (Suppl. 61), OA282. [Google Scholar]
- Shafiey, S.I.; Ahmed, K.A.; Abo-Saif, A.A.; Abo-Youssef, A.M.; Mohamed, W.R. Galantamine Mitigates Testicular Injury and Disturbed Spermatogenesis in Adjuvant Arthritic Rats via Modulating Apoptosis, Inflammatory Signals, and IL-6/JAK/STAT3/SOCS3 Signaling. Inflammopharmacology 2024, 32, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Xu, L.; Han, X.; Qi, Y.; Xu, Y.; Yin, L.; Liu, K.; Peng, J. Effects of the Total Saponins from Rosa Laevigata Michx Fruit against Acetaminophen-Induced Liver Damage in Mice via Induction of Autophagy and Suppression of Inflammation and Apoptosis. Molecules 2014, 19, 7189–7206. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Dai, T.; Chi, Z.; Zhao, Z.; Wu, G.; Yang, S.; Dong, D. Naringin Alleviates a Cetaminophen-induced Acute Liver Injury by Activating Nrf2 via CHAC2 Upregulation. Environ. Toxicol. 2022, 37, 1332–1342. [Google Scholar] [CrossRef]
- Wang, J.; Shi, Q.; Zhou, Q.; Zhang, L.; Qiu, Y.; Lou, D.; Zhou, L.; Yang, B.; He, Q.; Weng, Q. Sapidolide A Alleviates Acetaminophen-Induced Acute Liver Injury by Inhibiting NLRP3 Inflammasome Activation in Macrophages. Acta Pharmacol. Sin. 2022, 43, 2016–2025. [Google Scholar] [CrossRef]
- Samra, Y.A.; Hamed, M.F.; El-Sheakh, A.R. Hepatoprotective Effect of Allicin against Acetaminophen-induced Liver Injury: Role of Inflammasome Pathway, Apoptosis, and Liver Regeneration. J. Biochem. Mol. Toxicol. 2020, 34, e22470. [Google Scholar] [CrossRef]
- Liu, X.; Liu, R.; Wang, Y. Indole-3-Carboxaldehyde Alleviates Acetaminophen-Induced Liver Injury via Inhibition of Oxidative Stress and Apoptosis. Biochem. Biophys. Res. Commun. 2024, 710, 149880. [Google Scholar] [CrossRef]
- Munshi, C.; Paul, T.; Jas, K.; Bose, M.; Bhattacharya, S. Mitochondrial Involvement in Ferroptotic Cell Death. Glob. Transl. Med. 2024, 3, 2208. [Google Scholar] [CrossRef]
- Tian, G.; Yu, Y.; Deng, H.; Yang, L.; Shi, X.; Yu, B. Empagliflozin Alleviates Ethanol-induced Cardiomyocyte Injury through Inhibition of Mitochondrial Apoptosis via a SIRT1/PTEN/Akt Pathway. Clin. Exp. Pharmacol. Physiol. 2021, 48, 837–845. [Google Scholar] [CrossRef]
- Ye, T.; Zhang, J.; Wu, D.; Shi, J.; Kuang, Z.; Ma, Y.; Xu, Q.; Chen, B.; Kan, C.; Sun, X. Empagliflozin Attenuates Obesity-Related Kidney Dysfunction and NLRP3 Inflammasome Activity Through the HO-1–Adiponectin Axis. Front. Endocrinol. 2022, 13, 907984. [Google Scholar] [CrossRef]
- Pan, Y.; Cao, M.; You, D.; Qin, G.; Liu, Z. Research Progress on the Animal Models of Drug-induced Liver Injury: Current Status and Further Perspectives. Biomed. Res. Int. 2019, 2019, 1283824. [Google Scholar] [CrossRef] [PubMed]
- Botros, S.R.; Matouk, A.I.; Anter, A.; Khalifa, M.M.A.; Heeba, G.H. Protective Effect of Empagliflozin on Gentamicin-Induced Acute Renal Injury via Regulation of SIRT1/NF-ΚB Signaling Pathway. Environ. Toxicol. Pharmacol. 2022, 94, 103907. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008; ISBN 0443102791. [Google Scholar]
- Blazka, M.E.; Elwell, M.R.; Holladay, S.D.; Wilson, R.E.; Luster, M.I. Histopathology of Acetaminophen-Induced Liver Changes: Role of Interleukin 1α and Tumor Necrosis Factor α. Toxicol. Pathol. 1996, 24, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Groner, B.; Müller, C.W. Three-Dimensional Structure of the Stat3β Homodimer Bound to DNA. Nature 1998, 394, 145–151. [Google Scholar] [CrossRef]
- Ahmed, A.H.; Thompson, M.D.; Fenwick, M.K.; Romero, B.; Loh, A.P.; Jane, D.E.; Sondermann, H.; Oswald, R.E. Mechanisms of Antagonism of the GluR2 AMPA Receptor: Structure and Dynamics of the Complex of Two Willardiine Antagonists with the Glutamate Binding Domain. Biochemistry 2009, 48, 3894–3903. [Google Scholar] [CrossRef]
- Bergamin, E.; Wu, J.; Hubbard, S.R. Structural Basis for Phosphotyrosine Recognition by Suppressor of Cytokine Signaling-3. Structure 2006, 14, 1285–1292. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Sanner, M.F. Python: A Programming Language for Software Integration and Development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Gasteiger, J.; Marsili, M. Iterative Partial Equalization of Orbital Electronegativity—A Rapid Access to Atomic Charges. Tetrahedron 1980, 36, 3219–3228. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Median | Min | Max |
|---|---|---|---|
| Vehicle control | 0 | 0 | 0 |
| APAP | 3 * | 2 | 3 |
| EMPA 10 | 2 * | 1 | 2 |
| EMPA 20 | 1 # | 0 | 1 |
| NAC | 0 #, @ | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhaddad, A.; Mosalam, E.M.; AboShabaan, H.S.; Sallam, A.S.; Mahfouz, M.M.; Elhosary, E.; Mohammed, A.A.; Metwally, E.M.; Shaldam, M.A.; Ghoneim, M.E.-S. Mechanistic and Molecular Insights into Empagliflozin’s Role in Ferroptosis and Inflammation Trajectories in Acetaminophen-Induced Hepatotoxicity. Pharmaceuticals 2025, 18, 405. https://doi.org/10.3390/ph18030405
Alhaddad A, Mosalam EM, AboShabaan HS, Sallam AS, Mahfouz MM, Elhosary E, Mohammed AA, Metwally EM, Shaldam MA, Ghoneim ME-S. Mechanistic and Molecular Insights into Empagliflozin’s Role in Ferroptosis and Inflammation Trajectories in Acetaminophen-Induced Hepatotoxicity. Pharmaceuticals. 2025; 18(3):405. https://doi.org/10.3390/ph18030405
Chicago/Turabian StyleAlhaddad, Aisha, Esraa M. Mosalam, Hind S. AboShabaan, Amany Said Sallam, Marwa M. Mahfouz, Enas Elhosary, Asmaa A. Mohammed, Ebtehal M. Metwally, Moataz A. Shaldam, and Mai El-Sayed Ghoneim. 2025. "Mechanistic and Molecular Insights into Empagliflozin’s Role in Ferroptosis and Inflammation Trajectories in Acetaminophen-Induced Hepatotoxicity" Pharmaceuticals 18, no. 3: 405. https://doi.org/10.3390/ph18030405
APA StyleAlhaddad, A., Mosalam, E. M., AboShabaan, H. S., Sallam, A. S., Mahfouz, M. M., Elhosary, E., Mohammed, A. A., Metwally, E. M., Shaldam, M. A., & Ghoneim, M. E.-S. (2025). Mechanistic and Molecular Insights into Empagliflozin’s Role in Ferroptosis and Inflammation Trajectories in Acetaminophen-Induced Hepatotoxicity. Pharmaceuticals, 18(3), 405. https://doi.org/10.3390/ph18030405