
Chemical Composition and Biological Activities of Lagopsis supina Extract: Antioxidant, Adipogenic, and Ani-Inflammatory Effects

 ,
,  , and
, and
Abstract
1. Introduction
2. Results
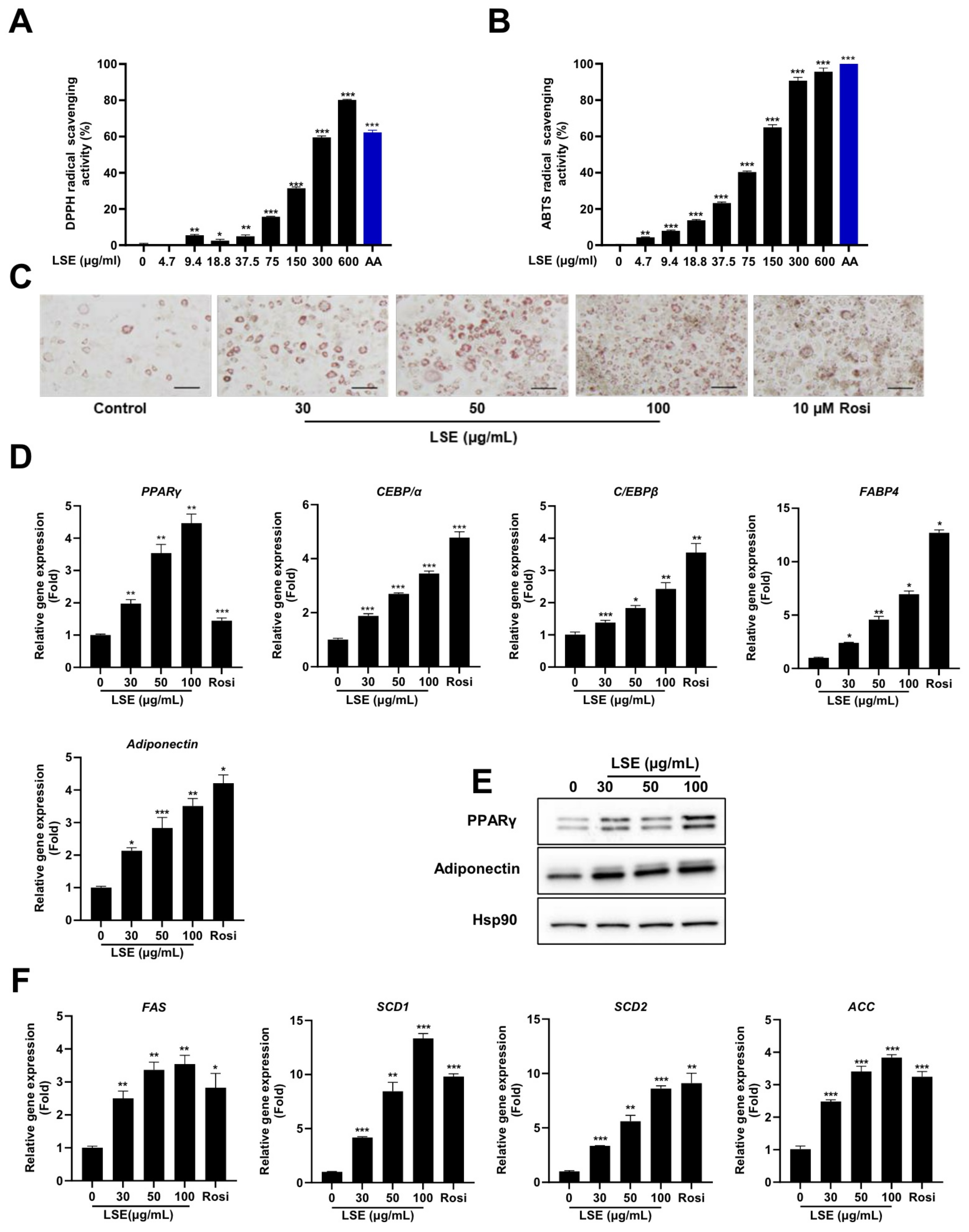
2.1. Antioxidative Capacity of LSE Through Radical Scavenging Activity
2.2. LSE Stimulates 3T3-L1 Adipocyte Differentiation Without Cytotoxicity
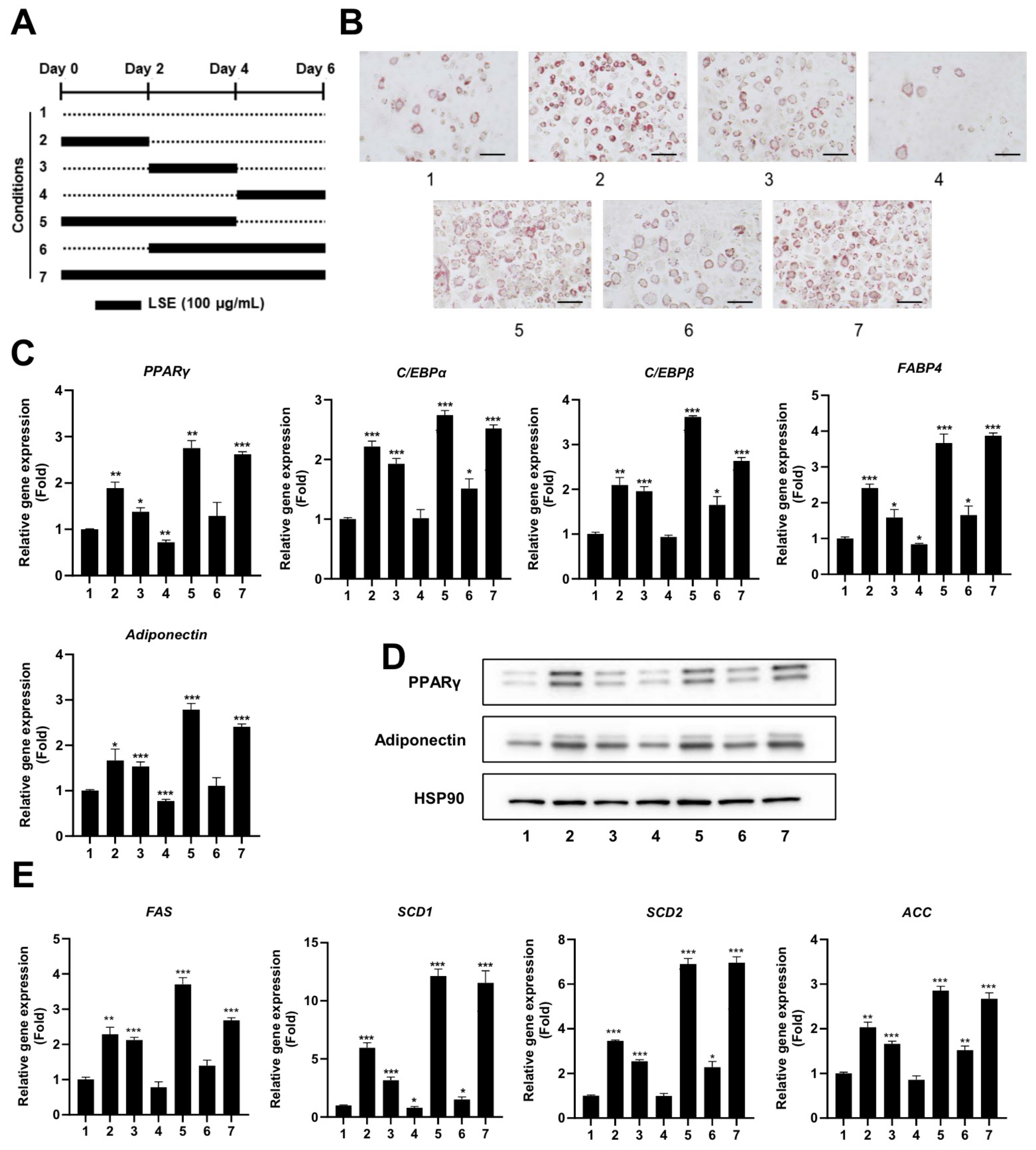
2.3. LSE Exhibits Enhanced Effects on Adipocyte Differentiation During Early and Intermediate Stages
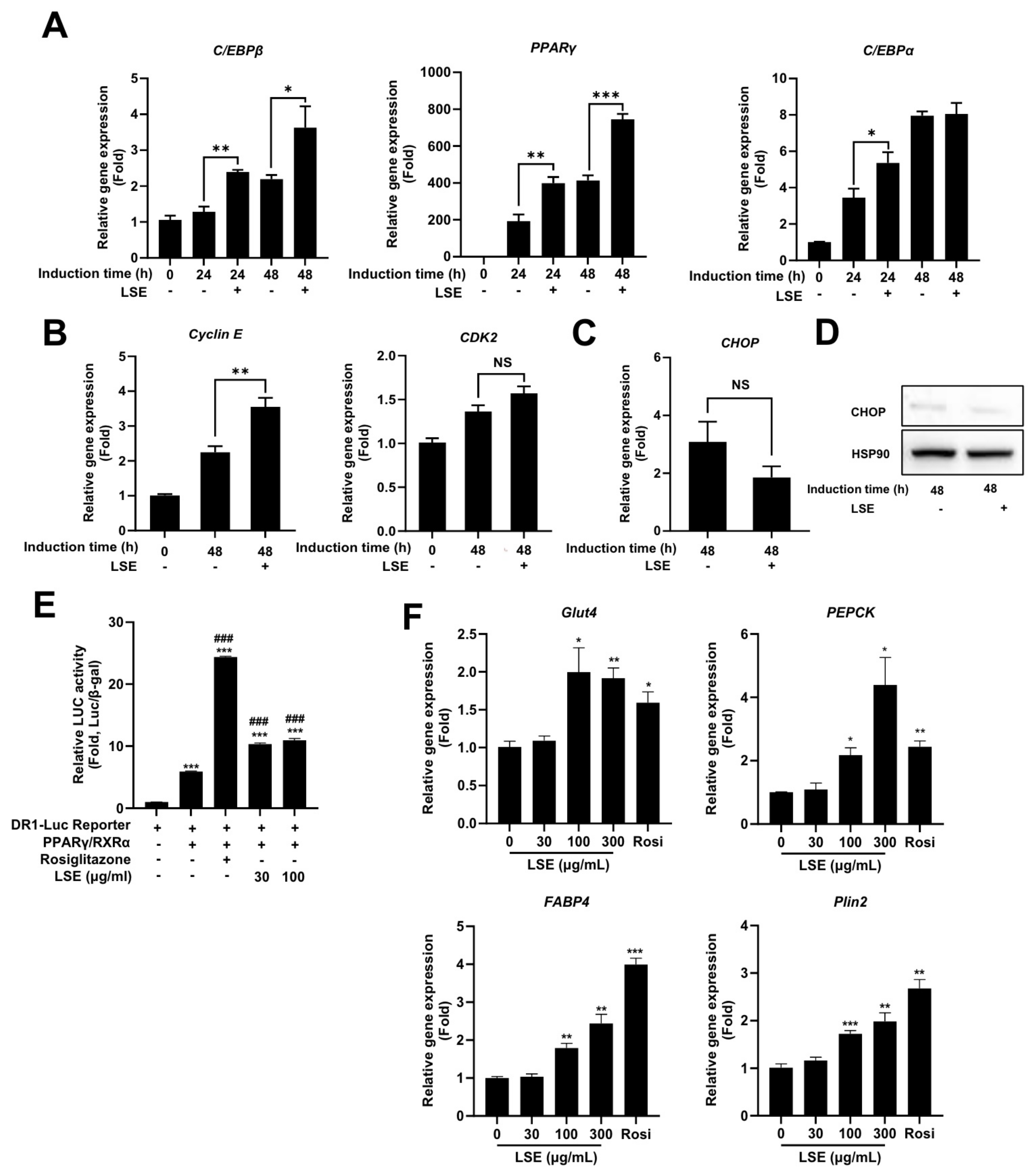
2.4. LSE Promotes Adipocyte Differentiation Through Enhanced Expression of Key Regulatory Genes
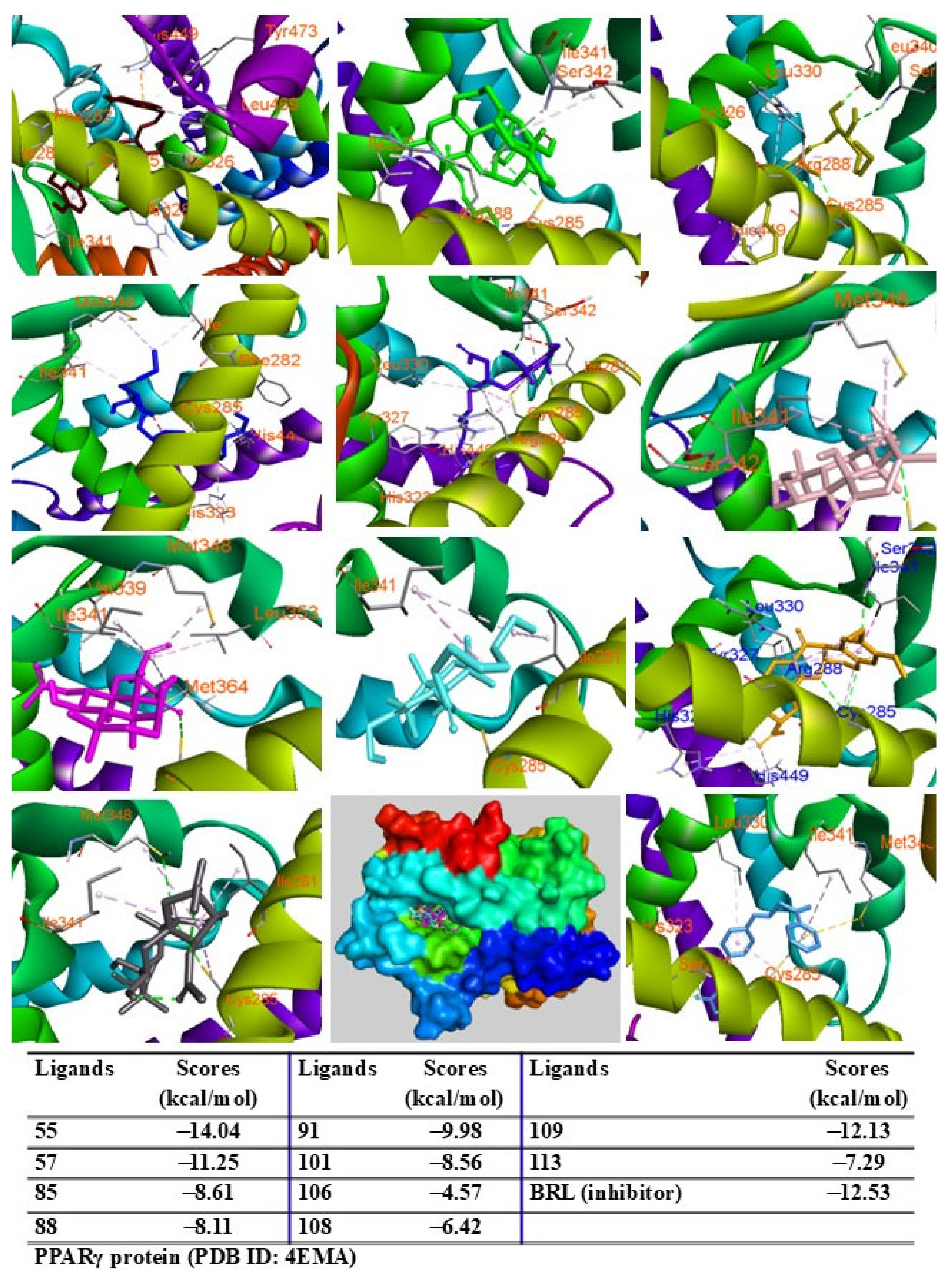
2.5. LSE Stimulates PPARγ Activity and Upregulates PPARγ Target Genes
2.6. Cytotoxicity and Nitric Oxide (NO) Inhibitory Activity of LSE in RAW264.7 Macrophages
2.7. Molecular Mechanisms Underlying the Anti-Inflammatory Effect of LSE
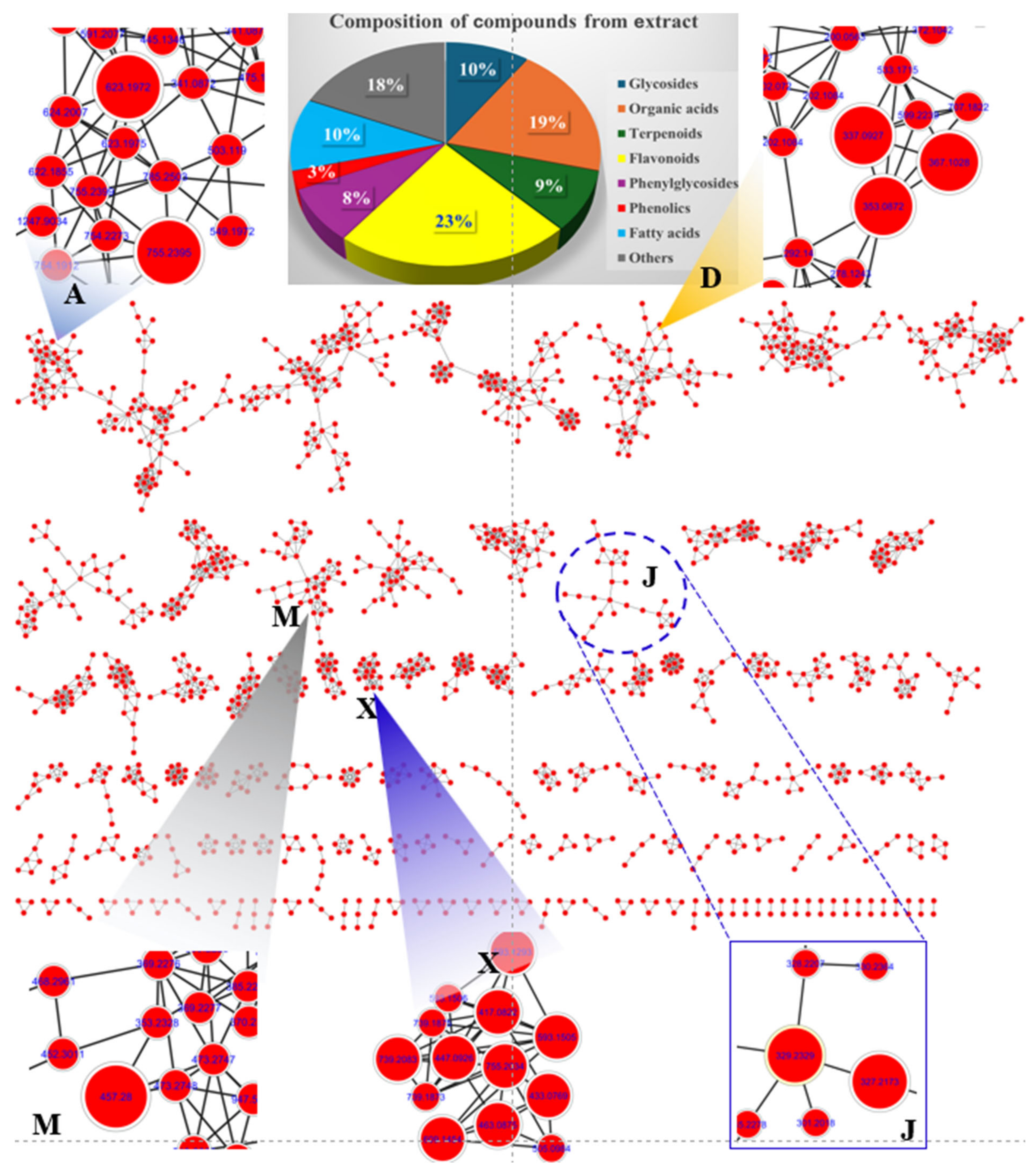
2.8. Identification of Metabolites from LSE
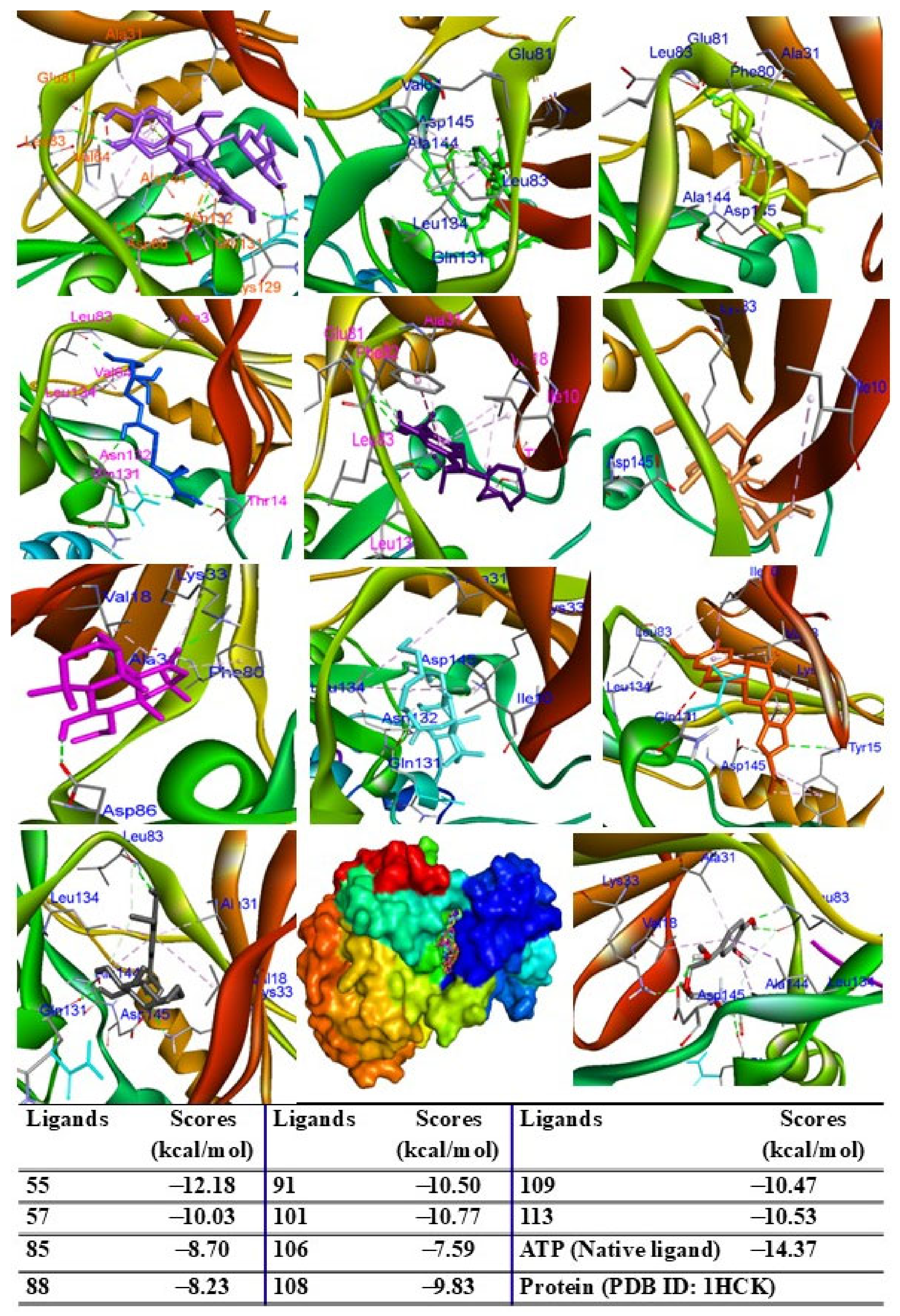
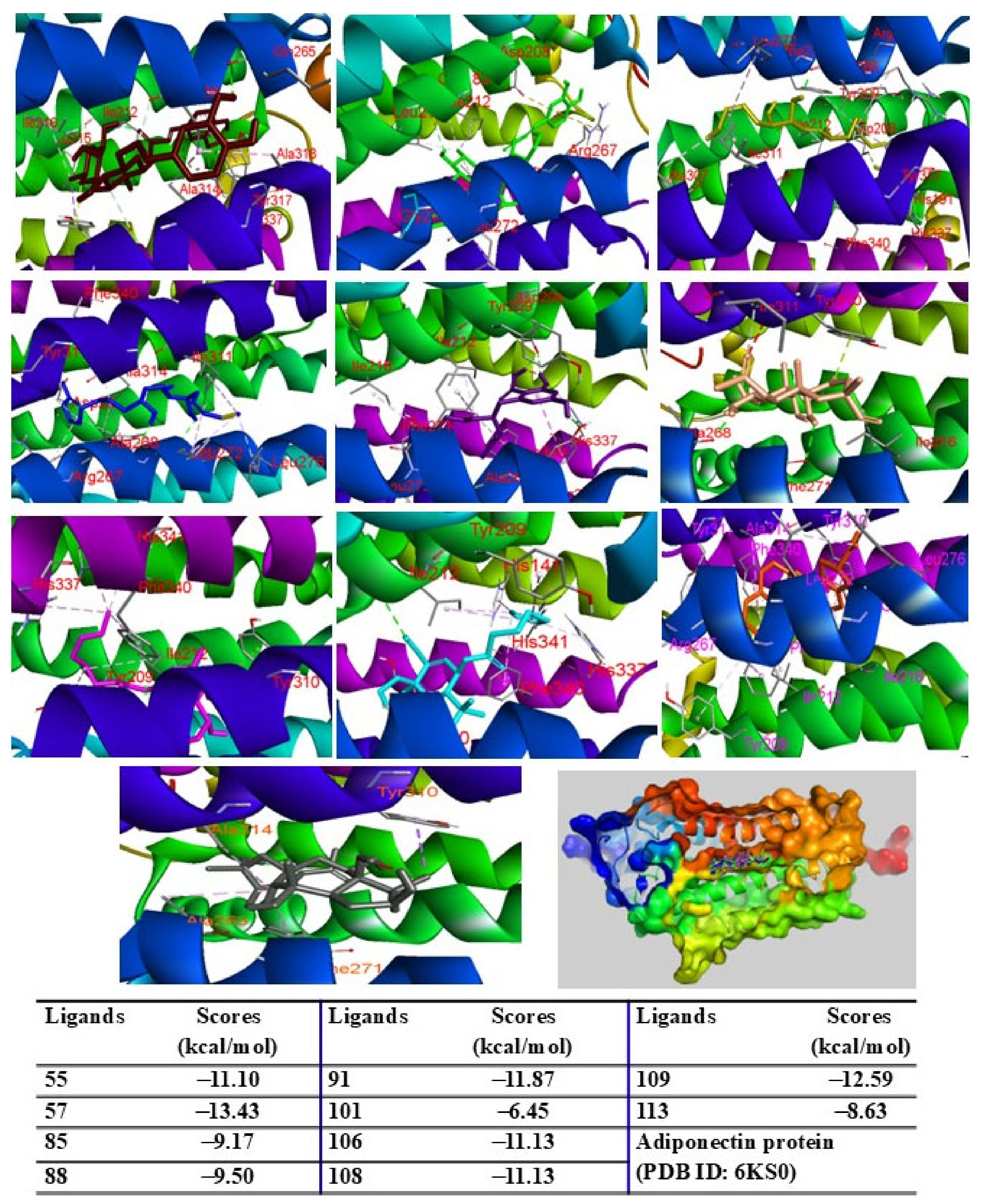
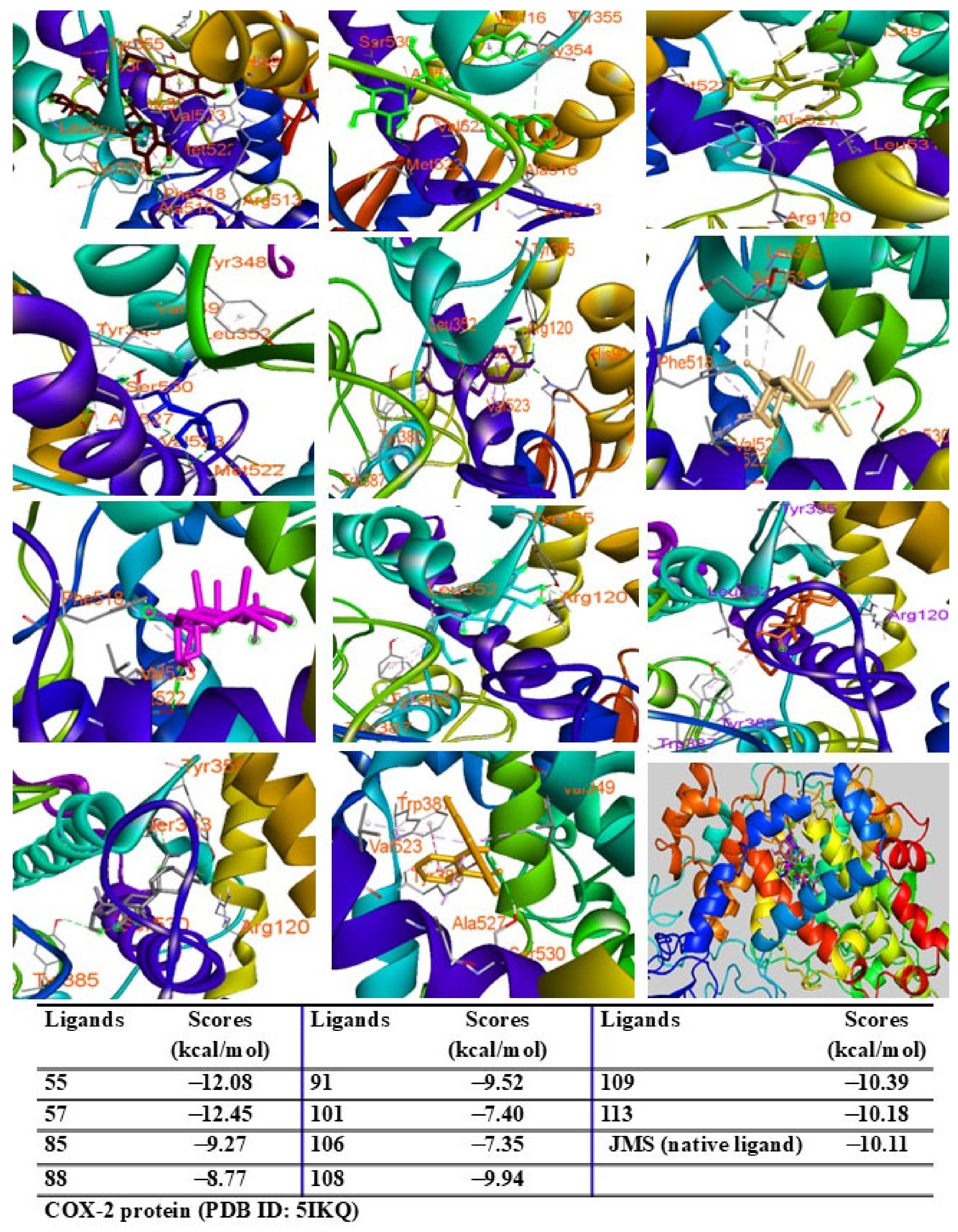
2.9. Molecular Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Methanol Extract of Lagopsis Supina (Stephan Ex Willd.) Ikonn.-Gal
4.2. DPPH and ABTS Assays
4.3. Cell Culture and Adipocyte Differentiation
4.4. Cell Viability Assay
4.5. NO Production Assay in RAW 264.7 Cells
4.6. Oil Red O Staining for Lipid Droplets
4.7. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR) Analysis
4.8. Western Blot Analysis
4.9. Transfection and Luciferase and β-galactosidase Assay
4.10. UPLC-ESI-Orbitrap MS/MS
4.11. Molecular Networking
4.12. LC-MS Annotation
4.13. Molecular Docking Assay
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Parvanova, A.; Reseghetti, E.; Abbate, M.; Ruggenenti, P. Mechanisms and treatment of obesity-related hypertension—Part 1: Mechanisms. Clin. Kidney J. 2024, 17, sfad282. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Shen, Z.; Gu, W.; Lyu, Z.; Qi, X.; Mu, Y.; Ning, Y. Prevalence of obesity and associated complications in China: A cross-sectional, real-world study in 15.8 million adults. Diabetes Obes. Metab. 2023, 25, 3390–3399. [Google Scholar] [CrossRef]
- Pi-Sunyer, X. The medical risks of obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef]
- Shimobayashi, M.; Albert, V.; Woelnerhanssen, B.; Frei, I.C.; Weissenberger, D.; Meyer-Gerspach, A.C.; Clement, N.; Moes, S.; Colombi, M.; Meier, J.A. Insulin resistance causes inflammation in adipose tissue. J. Clin. Investig. 2018, 128, 1538–1550. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic inflammation and insulin resistance in obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like receptors and the control of immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Laroux, F.; Pavlick, K.; Hines, I.; Kawachi, S.; Harada, H.; Bharwani, S.; Hoffman, J.; Grisham, M. Role of nitric oxide in inflammation. Acta Physiol. Scand. 2001, 173, 113–118. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in inflammatory disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Z.; Huo, H.-X.; Li, Y.-T.; Pang, D.-R.; Zheng, J.; Zhang, Q.; Zhao, Y.-F.; Tu, P.-F.; Li, J. Chemical constituents from Lagopsis supina (steph.) Ik.-Gal. Ex knorr. Biochem. Syst. Ecol. 2015, 61, 424–428. [Google Scholar] [CrossRef]
- Li, H.; Li, M.-M.; Su, X.-Q.; Sun, J.; Gu, Y.-F.; Zeng, K.-W.; Zhang, Q.; Zhao, Y.-F.; Ferreira, D.; Zjawiony, J.K. Anti-inflammatory labdane diterpenoids from Lagopsis supina. J. Nat. Prod. 2014, 77, 1047–1053. [Google Scholar] [CrossRef]
- Yang, L.; Zeng, H.; Xia, X.; Wang, H.; Zhao, B.; He, J. Natural phenylethanoid glycosides diuretics derived from Lagopsis supina: Biological activity, mechanism, molecular docking, and structure-activity relationship. Bioorg. Chem. 2022, 129, 106165. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Liu, Y.-Y.; Li, Y.-L.; Shi, W.-Y.; Li, K.-X.; Lu, L.-W.; Zhou, L.; Lin, B.; Huang, X.-X.; Song, S.-J. Meroterpenoids and sesquiterpene dimers from Sarcandra glabra with anti-neuroinflammatory activity. Ind. Crops Prod. 2022, 183, 114983. [Google Scholar] [CrossRef]
- He, J.; Zeng, L.; Wei, R.; Zhong, G.; Zhu, Y.; Xu, T.; Yang, L. Lagopsis supina exerts its diuretic effect via inhibition of aquaporin-1, 2 and 3 expression in a rat model of traumatic blood stasis. J. Ethnopharmacol. 2019, 231, 446–452. [Google Scholar] [CrossRef]
- He, J.; Zhang, Q.; Xia, X.; Yang, L. Lagopsis supina ameliorates myocardial ischemia injury by regulating angiogenesis, thrombosis, inflammation, and energy metabolism through VEGF, ROS and HMGB1 signaling pathways in rats. Phytomedicine 2023, 120, 155050. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, L.; Li, L.; Wei, R.; Luo, X.; Xu, T.; Huang, Y.; Mu, Z.; He, J. Diuretic and antidiuretic activities of ethanol extract and fractions of Lagopsis supina in normal rats. Biomed. Res. Int. 2019, 2019, 6927374. [Google Scholar] [CrossRef]
- Zhang, J.; Pang, D.; Huang, Z.; Huo, H.; Li, Y.; Zheng, J.; Zhang, Q.; Zhao, Y.; Tu, P.; Li, J. Flavonoids from whole plants of Lagopsis supina. Zhongguo Zhong Yao Za Zhi 2015, 40, 3224–3228. [Google Scholar] [PubMed]
- Yang, L.; He, Z.-W.; He, J.-W. The chemical profiling of aqueous soluble fraction from Lagopsis supina and its diuretic effects via suppression of AQP and RAAS pathways in saline-loaded rats. J. Ethnopharmacol. 2021, 272, 113951. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, J.; Sharma, R.P. Inhibition of p38 and ERK MAP kinases blocks endotoxin-induced nitric oxide production and differentially modulates cytokine expression. Pharmacol. Res. 2004, 49, 433–439. [Google Scholar] [CrossRef]
- Wang, K.; Shi, J.-T.; Zhang, Y.; Ma, L.-M.; Wang, J.; Yang, J.-L. Labdane diterpenoids from Lagopsis supina and their anti-neuroinflammatory activity. Fitoterapia 2023, 169, 105611. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; He, J. Lagopsis supina extract and its fractions exert prophylactic effects against blood stasis in rats via anti-coagulation, anti-platelet activation and anti-fibrinolysis and chemical characterization by UHPLC-qTOF-MS/MS. Biomed. Pharmacother. 2020, 132, 110899. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; He, X.; Ma, C. Metabolism schemes of chlorogenic, quinic and caffeic acid derivatives in rats deduced by molecular fragments detected with optimized UPLC–APCI-MS/MS conditions. Int. J. Mass Spectrom. 2013, 337, 34–42. [Google Scholar] [CrossRef]
- Pikulski, M.; Brodbelt, J.S. Differentiation of flavonoid glycoside isomers by using metal complexation and electrospray ionization mass spectrometry. J. Am. Soc. Mass Spectrom. 2003, 14, 1437–1453. [Google Scholar] [CrossRef]
- Pascale, R.; Acquavia, M.A.; Cataldi, T.R.I.; Onzo, A.; Coviello, D.; Bufo, S.A.; Scrano, L.; Ciriello, R.; Guerrieri, A.; Bianco, G. Profiling of quercetin glycosides and acyl glycosides in sun-dried peperoni di Senise peppers (Capsicum annuum L.) by a combination of LC-ESI(-)-MS/MS and polarity prediction in reversed-phase separations. Anal. Bioanal. Chem. 2020, 412, 3005–3015. [Google Scholar] [CrossRef]
- Haththotuwa, R.N.; Wijeyaratne, C.N.; Senarath, U. Worldwide epidemic of obesity. In Obesity and Obstetrics; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–8. [Google Scholar] [CrossRef]
- Scully, T.; Ettela, A.; LeRoith, D.; Gallagher, E.J. Obesity, type 2 diabetes, and cancer risk. Front. Oncol. 2021, 10, 615375. [Google Scholar] [CrossRef]
- Mohajan, D.; Mohajan, H.K. Obesity and its related diseases: A new escalating alarming in global health. J. Innovat. Med. Res. 2023, 2, 12–23. [Google Scholar] [CrossRef]
- Wadden, T.A.; Tronieri, J.S.; Butryn, M.L. Lifestyle modification approaches for the treatment of obesity in adults. Am. Psychol. 2020, 75, 235. [Google Scholar] [CrossRef]
- Barrea, L.; Salzano, C.; Pugliese, G.; Laudisio, D.; Frias-Toral, E.; Savastano, S.; Colao, A.; Muscogiuri, G. The challenge of weight loss maintenance in obesity: A review of the evidence on the best strategies available. Int. J. Food Sci. Nutr. 2022, 73, 1030–1046. [Google Scholar] [CrossRef]
- Williams, D.M.; Nawaz, A.; Evans, M. Drug therapy in obesity: A review of current and emerging treatments. Diabetes Ther. 2020, 11, 1199–1216. [Google Scholar] [CrossRef]
- Gadde, K.M.; Atkins, K.D. The limits and challenges of antiobesity pharmacotherapy. Expert Opin. Pharmacother. 2020, 21, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, Z.; Sun, Y.; Yan, M.; Wang, Y.; Li, Y.; Zhang, Y.; Zhu, M. Supplementation of chlorogenic acid alleviates the effects of H2O2-induced oxidative stress on laying performance, egg quality, antioxidant capacity, hepatic inflammation, mitochondrial dysfunction, and lipid accumulation in laying hens. Antioxidants 2024, 13, 1303. [Google Scholar] [CrossRef] [PubMed]
- Jang, E. Hyperoside as a potential natural product targeting oxidative stress in liver diseases. Antioxidants 2022, 11, 1437. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Liu, Q.; Salter, A.M.; Lomax, M.A. Synergism between cAMP and PPARγ signalling in the initiation of UCP1 gene expression in HIB1B brown adipocytes. PPAR Res. 2013, 2013, 476049. [Google Scholar] [CrossRef]
- Shrestha, D.; Kim, E.; Shrestha, K.K.; Suh, S.-S.; Kim, S.-H.; Seo, J.B. Methanol extract of Elsholtzia fruticosa promotes 3T3-L1 preadipocyte differentiation. J. Anim. Sci. Technol. 2024, 66, 204–218. [Google Scholar] [CrossRef]
- Bakkar, N.-M.Z.; Dwaib, H.S.; Fares, S.; Eid, A.H.; Al-Dhaheri, Y.; El-Yazbi, A.F. Cardiac autonomic neuropathy: A progressive consequence of chronic low-grade inflammation in type 2 diabetes and related metabolic disorders. Int. J. Mol. Sci. 2020, 21, 9005. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as cytokine modulators: A possible therapy for inflammation-related diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef]
- Ambriz-Pérez, D.L.; Leyva-López, N.; Gutierrez-Grijalva, E.P.; Heredia, J.B. Phenolic compounds: Natural alternative in inflammation treatment. A Review. Cogent Food Agric. 2016, 2, 1131412. [Google Scholar] [CrossRef]
- Grilli, E.; Tugnoli, B.; Passey, J.L.; Stahl, C.H.; Piva, A.; Moeser, A.J. Impact of dietary organic acids and botanicals on intestinal integrity and inflammation in weaned pigs. BMC Vet. Res. 2015, 11, 96. [Google Scholar] [CrossRef]
- Huang, J.; Xie, M.; He, L.; Song, X.; Cao, T. Chlorogenic acid: A review on its mechanisms of anti-inflammation, disease treatment, and related delivery systems. Front. Pharmacol. 2023, 14, 1218015. [Google Scholar] [CrossRef]
- Xue, J.-C.; Yuan, S.; Meng, H.; Hou, X.-T.; Li, J.; Zhang, H.-M.; Chen, L.-L.; Zhang, C.-H.; Zhang, Q.-G. The role and mechanism of flavonoid herbal natural products in ulcerative colitis. Biomed. Pharmacother. 2023, 158, 114086. [Google Scholar] [CrossRef] [PubMed]
- Adeyi, O.E.; Somade, O.T.; Ajayi, B.O.; James, A.S.; Adeboye, T.R.; Olufemi, D.A.; Oyinlola, E.V.; Sanyaolu, E.T.; Mufutau, I.O. The anti-inflammatory effect of ferulic acid is via the modulation of NFκB-TNF-α-IL-6 and STAT1-PIAS1 signaling pathways in 2-methoxyethanol-induced testicular inflammation in rats. Phytomed. Plus 2023, 3, 100464. [Google Scholar] [CrossRef]
- Lee, J.; Choi, J.W.; Sohng, J.K.; Pandey, R.P.; Park, Y.I. The immunostimulating activity of quercetin 3-O-xyloside in murine macrophages via activation of the ASK1/MAPK/NF-κB signaling pathway. Int. Immunopharmacol. 2016, 31, 88–97. [Google Scholar] [CrossRef]
- Krishnamoorthy, K.; Senguttuvan, J. Evaluation of anti-inflammatory and antioxidant properties of crude extract and forskolin from Solena amplexicaulis Leaf. Indian J. Pharm. Sci. 2016, 78, 377–387. [Google Scholar] [CrossRef]
- Bombicino, S.S.; Iglesias, D.E.; Zaobornyj, T.; Boveris, A.; Valdez, L.B. Mitochondrial nitric oxide production supported by reverse electron transfer. Arch. Biochem. Biophys. 2016, 607, 8–19. [Google Scholar] [CrossRef]
- Peng, S.-g.; Pang, Y.-l.; Zhu, Q.; Kang, J.-h.; Liu, M.-x.; Wang, Z. Chlorogenic acid functions as a novel agonist of PPARγ2 during the differentiation of mouse 3T3-L1 preadipocytes. BioMed. Res. Int. 2018, 2018, 8594767. [Google Scholar] [CrossRef]
- Park, J.-E.; Han, J.-S. Improving the effect of ferulic acid on inflammation and insulin resistance by regulating the JNK/ERK and NF-κB pathways in TNF-α-treated 3T3-L1 adipocytes. Nutrients 2024, 16, 294. [Google Scholar] [CrossRef]
- Yeon, J.; Kim, E.; Bazarragchaa, B.; Kim, S.-Y.; Huh, J.Y.; Park, H.; Suh, S.-S.; Seo, J.B. Stellera chamaejasme L. extract inhibits adipocyte differentiation through activation of the extracellular signal-regulated kinase pathway. PLoS ONE 2024, 19, e0300520. [Google Scholar] [CrossRef]
- Le, D.D.; Kim, E.; Dang, T.; Lee, J.; Shin, C.H.; Park, J.W.; Lee, S.-g.; Seo, J.B.; Lee, M. Chemical investigation and regulation of adipogenic differentiation of cultivated Moringa oleifera. Pharmaceuticals 2024, 17, 1310. [Google Scholar] [CrossRef]
- Le, D.D.; Kim, Y.-R.; Yu, S.; Dang, T.; Lee, K.-T.; Lee, M. Phytochemical and bioactivities of promising flavonoid glycosides and their content from unmatured fruits of Vicia bungei with their bioactivity. Food Chem. 2024, 460, 140541. [Google Scholar] [CrossRef]
- Le, D.D.; Jang, Y.S.; Truong, V.; Yu, S.; Dinh, T.; Lee, M. Bioactivities of quinic acids from Vitex rotundifolia obtained by supercritical fluid extraction. Antioxidants 2024, 13, 1235. [Google Scholar] [CrossRef] [PubMed]


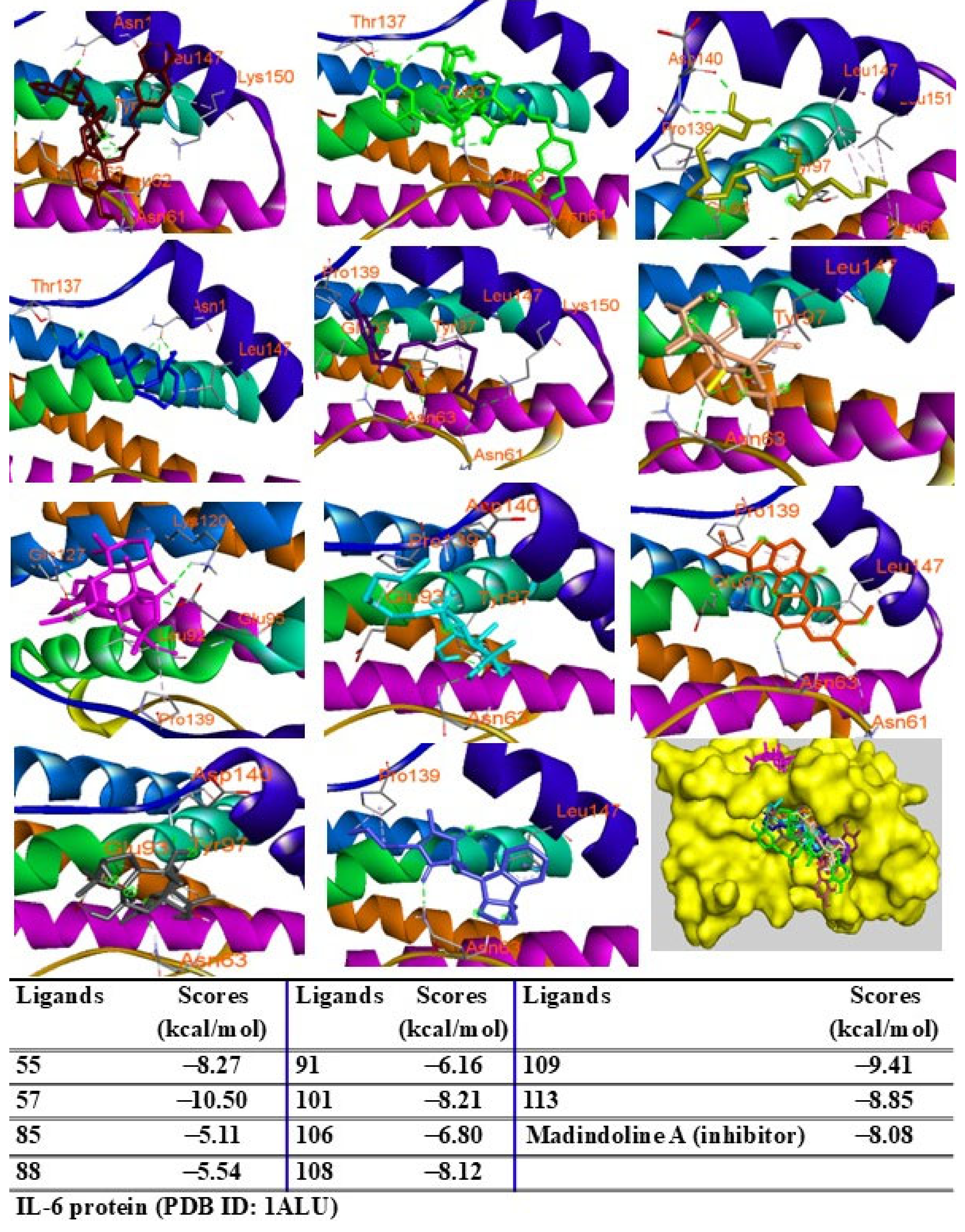
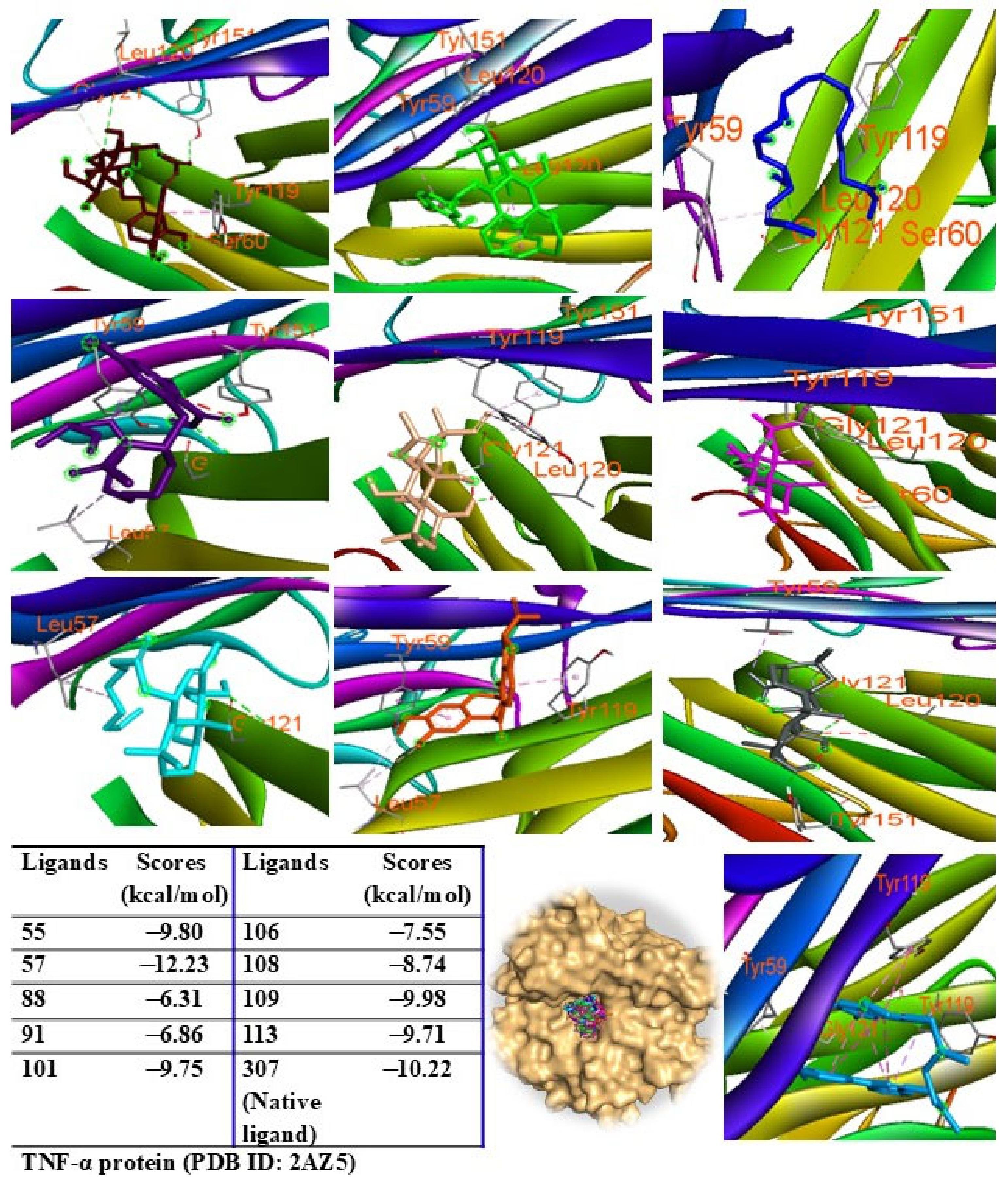
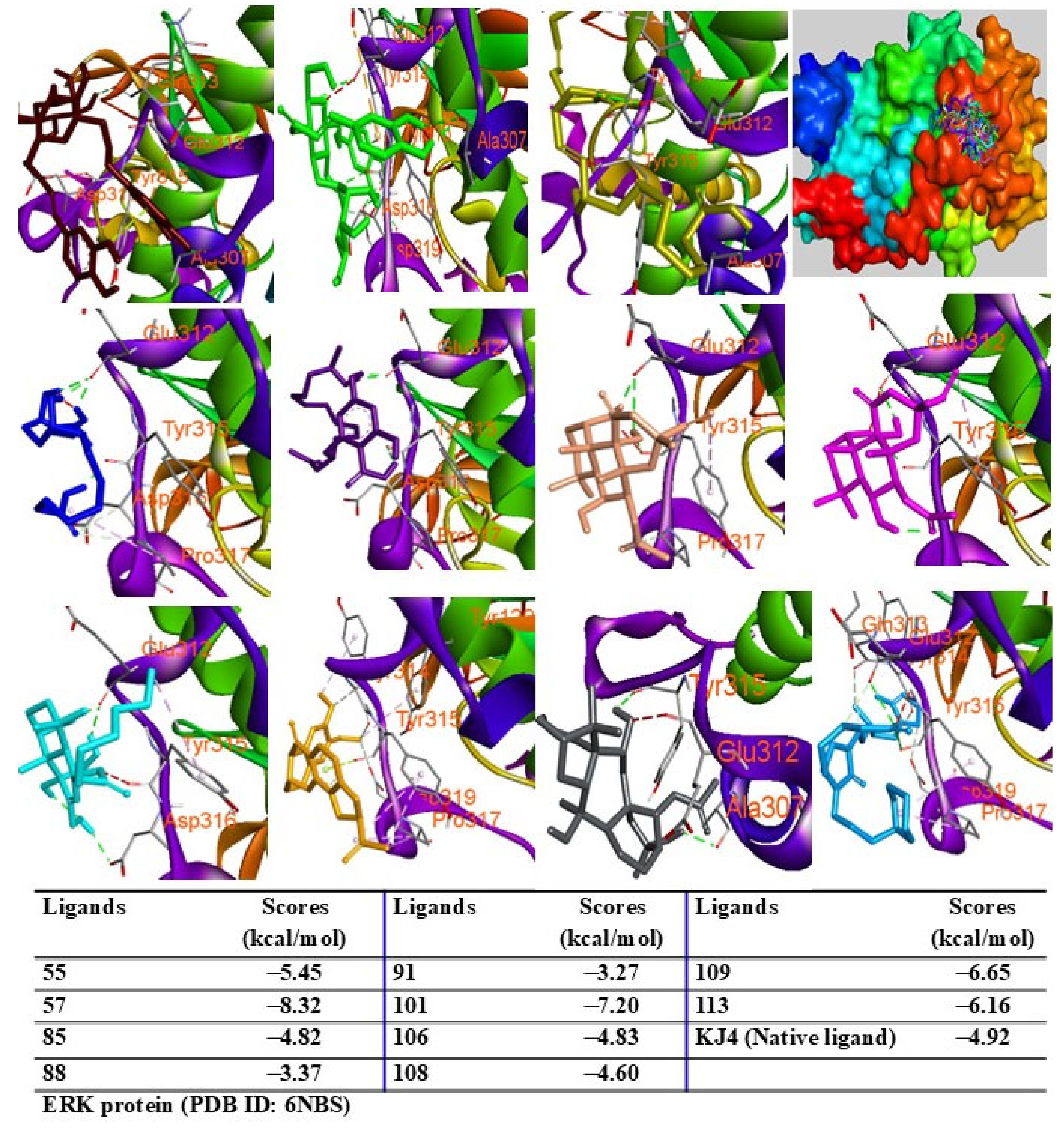

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | RT (min) | Formula | Adduct | m/z (Da) | Error (mDa) | Class |
|---|---|---|---|---|---|---|---|
| 1 | Unknown | 1.080 | - | [M-H]− | 386.9382 | - | - |
| 2 | Unknown | 1.090 | - | [M-H]− | 272.9587 | - | - |
| 3 | Unknown | 1.090 | - | [M-H]− | 158.9782 | - | - |
| 4 | Geshoidin | 1.273 | C18H18O9 | [M-H]− | 377.0853 | 2.496 | Glycosides |
| 5 | Quinic acid † | 1.304 | C7H12O6 | [M-H]− | 191.0559 | 0.172 | Organic acids |
| 6 | Unknown | 1.310 | - | [M-H]− | 404.1040 | - | - |
| 7 | (S)-malate | 1.342 | C4H6O5 | [M-H]− | 133.0140 | 0.168 | Organic acids |
| 8 | N-Fructosyl pyroglutamate | 1.349 | C11H17NO8 | [M-H]− | 290.0879 | 0.782 | Organic acids |
| 9 | Sucrose | 1.379 | C12H22O11 | [M-H]− | 341.1083 | 0.651 | Glycosides |
| 10 | 5-Oxo-D-proline | 1.902 | C5H7NO3 | [M-H]− | 128.0352 | 0.103 | Organic acids |
| 11 | Citrate | 1.902 | C6H8O7 | [M-H]− | 191.0194 | 0.258 | Organic acids |
| 12 | Succinate | 2.187 | C4H6O4 | [M-H]− | 117.0192 | 0.061 | Organic acids |
| 13 | 5,9-dihydroxy-5,7,7-trimethyl-4,5a,6,8,8a,9-hexahydro-1H-azuleno[5,6-c]furan-3-one | 2.221 | C15H22O4 | [M-H]− | 265.0928 | 1.156 | Sesquiterpenes |
| 14 | 1-O-Caffeoylglucose | 2.221 | C15H18O9 | [M-H]− | 341.0873 | 0.295 | Glycosides |
| 15 | (1S,6S,13S)-13-Hydroxy-16,17-dimethoxy-6-prop-1-en-2-yl-2,7,20-trioxapentacyclo[11.8.0.03,11.04,8.014,19]henicosa-3(11),4(8),9,14,16,18-hexaen-12-one | 2.327 | C23H22O7 | [M-H]− | 409.1349 | 4.917 | Isoflavonoids |
| 16 | 3-(4-Hydroxyphenyl)lactate | 3.155 | C9H10O4 | [M-H]− | 181.0506 | 0.032 | Organic acids |
| 17 | Unknown | 3.155 | - | [M-H]− | 361.1137 | - | - |
| 18 | 6a,12a-Didehydroamorphigenin | 3.155 | C23H20O7 | [M-H]− | 407.1183 | 4.768 | Isoflavonoids |
| 19 | 7-methyl-1-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-1,5,6,7a-tetrahydrocyclopenta[c]pyran-4a,5,7-triol | 4.053 | C15H24O10 | [M-H]− | 363.1290 | 0.588 | Glycosides |
| 20 | Benzoic acid † | 6.132 | C13H16O8 | [M-H]− | 137.0243 | 0.206 | Organic acids |
| 21 | Neochlorogenic acid † | 6.154 | C16H18O9 | [M-H]− | 353.0873 | 1.699 | Organic acids |
| 22 | [(2R,3S,4S,5R,6S)-6-(3,4-dihydroxybenzoyl)oxy-3,4,5-trihydroxyoxan-2-yl]methyl 3,4-dihydroxybenzoate | 6.189 | C20H20O12 | [M-H]− | 451.2175 | 4.395 | Phenylglycosides |
| 23 | Chlorogenic acid † | 6.652 | C167H18O9 | [M-H]− | 353.0872 | 0.824 | Phenolics |
| 24 | 2-[(4-Hydroxy-3,5-dimethoxyphenyl)methoxy]-6-(hydroxymethyl)oxane-3,4,5-triol | 6.936 | C15H22O9 | [M-H]− | 345.1186 | 0.459 | Glycosides |
| 25 | 7-methyl-1-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-1,4a,5,6,7,7a-hexahydrocyclopenta[c]pyran-4-carboxylic acid | 6.936 | C16H24O9 | [M-H]− | 405.1397 | 0.480 | Terpenoids |
| 26 | 9,13-Dihydroxy-4-megastigmen-3-one 9-glucoside | 7.152 | C19H32O8 | [M+HCOO]− | 433.2071 | 1.882 | Terpenoids |
| 27 | 4-(β-D-Glucopyranosyloxy)benzyl 2,3-dihydroxy-3-methylbutanoate | 7.220 | C18H26O10 | [M+HCOO]− | 447.1503 | 0.436 | Organic acids |
| 28 | 7-Epi-12-hydroxyjasmonic acid glucoside | 7.371 | C18H28O9 | [M-H]− | 387.1655 | 0.551 | Glycosides |
| 29 | Epigallocatechin † | 7.395 | C15H14O7 | [M-H]− | 305.0699 | 3.288 | Flavanols |
| 30 | 5-hydroxy-2,2,6,6-tetramethyl-4-[2-methyl-1-[2,4,6-trihydroxy-3-(2-methylpropanoyl)phenyl]propyl]cyclohex-4-ene-1,3-dione | 7.406 | C24H32O7 | [M-H]− | 431.1915 | 4.226 | Fatty acids |
| 31 | Methyl 2,4,10-triacetyloxy-5,9-dihydroxy-1,4a-dimethyl-7-propan-2-yl-2,3,4,9,10,10a-hexahydrophenanthrene-1-carboxylate | 7.406 | C27H36O10 | [M-H]− | 519.2437 | 0.771 | Sesquiterpenes |
| 32 | Aesculetin | 7.486 | C9H6O4 | [M-H]− | 177.0190 | 0.244 | Coumarins |
| 33 | Oregonoyl A | 7.509 | C33H36O12 | [M-H]− | 623.2188 | 4.846 | Phenylglycosides |
| 34 | Chrysophanol | 7.531 | C15H10O4 | [M-H]− | 253.0713 | 2.260 | Quinones |
| 35 | Phenylacetic acid | 7.676 | C8H8O2 | [M-H]− | 135.0451 | 0.048 | Organic acids |
| 36 | 3-(4-Hydroxyphenyl)pyruvic acid | 7.676 | C9H8O4 | [M-H]− | 179.0347 | 0.249 | Organic acids |
| 37 | Unknown | 7.709 | - | [M-H]− | 601.2348 | - | - |
| 38 | Bonanniol A | 7.742 | C25H28O6 | [M-H]− | 423.1866 | 4.618 | Dihydroflavonols |
| 39 | (2E)-4-Hydroxy-3,4-dimethyl-2-penten-1-yl 6-O-[(2S,3R,4R)-3,4-dihydroxy-4-(hydroxymethyl)tetrahydro-2-furanyl]-β-D-glucopyranoside | 7.742 | C18H32O11 | [M+HCOO]− | 469.1921 | 0.527 | Glycosides |
| 40 | 1H-3,9a-Methanocyclopent[c]oxocin-4-carboxylic acid, 3,4,5,6,6a,9-hexahydro-4-hydroxy-7-(1-hydroxy-1-methylethyl)- | 7.753 | C15H22O5 | [M-H]− | 281.1389 | 0.477 | Organic acids |
| 41 | Flavovilloside | 7.786 | C33H40O20 | [M-H]− | 755.2026 | 1.337 | Flavonoid glycosides |
| 42 | Strobide B | 7.831 | C11H10O7 | [M-H]− | 253.0351 | 0.233 | Phenolics |
| 43 | Manghaslin | 8.213 | C33H40O20 | [M-H]− | 755.2034 | 0.549 | Flavonoid glycosides |
| 44 | 2,3-dihydroxy-4-methoxy-4′-ethoxybenzophenone | 8.214 | C16H16O5 | [M-H]− | 287.1500 | 3.949 | Fatty acids |
| 45 | 3-O-Feruloylquinic acid † | 8.378 | C33H40O20 | [M-H]− | 367.1028 | 0.183 | Organic acids |
| 46 | Caffeic acid † | 8.410 | C9H8O4 | [M-H2O-H]− | 179.0347 | 0.170 | Organic acids |
| 47 | Unknown | 8.410 | - | [M-H]− | 591.0980 | - | - |
| 48 | Vanillylmandelic acid | 8.419 | C9H10O5 | [M-H]− | 197.0454 | 0.100 | Organic acids |
| 49 | [(2S,3S)-2-[(E,3R,4S)-3,4-dihydroxypent-1-enyl]-6-oxo-2,3-dihydropyran-3-yl] (E)-2-methylbut-2-enoate | 8.419 | C15H20O6 | [M-H]− | 295.1184 | 0.242 | Monoterpenoids |
| 50 | (-)-12-hydroxyjasmonic acid | 8.584 | C12H18O4 | [M-H]− | 225.1133 | 0.414 | Fatty acids |
| 51 | Coumaroyl quinic acid † | 8.752 | C16H18O8 | [M-H]− | 337.0927 | 0.702 | Organic acids |
| 52 | 3-[4,5-dihydroxy-3-[(2R,3R,4R,5R,6S)-3,4,5-trihydroxy-6-methyloxan-2-yl]oxy-6-[[(2R,3R,4R,5R,6S)-3,4,5-trihydroxy-6-methyloxan-2-yl]oxymethyl]oxan-2-yl]oxy-5,7-dihydroxy-2-(4-hydroxyphenyl)chromen-4-one | 8.808 | C33H40O19 | [M-H]− | 739.2083 | 0.671 | Flavonoid glycosides |
| 53 | Quercetin 3-O-α-rhamnopyranosyl-(1-2)-β-galactopyranoside | 8.899 | C27H30O16 | [M-H]− | 609.1454 | 0.860 | Flavonoid glycosides |
| 54 | Rutin † | 9.405 | C27H30O16 | [M-H]− | 609.1454 | 0.321 | Flavonoid glycosides |
| 55 | [(2R,3R,4R,5R,6R)-2-[[(2R,3R,4R)-3,4-dihydroxy-4-(hydroxymethyl)oxolan-2-yl]oxymethyl]-4-[(2S,3R,4R,5R,6S)-4,5-dihydroxy-6-methyl-3-[(2S,3R,4S,5S)-3,4,5-trihydroxyoxan-2-yl]oxyoxan-2-yl]oxy-6-[2-(3,4-dihydroxyphenyl)ethoxy]-5-hydroxyoxan-3-yl] (E)-3-(3,4-dihydroxyphenyl)prop-2-enoate | 9.415 | C34H44O19 | [M-H]− | 755.2395 | 1.025 | Phenylglycosides |
| 56 | Kaempferol 3-O-neohesperidoside † | 9.596 | C30H26O13 | [M-H]− | 593.1505 | 0.488 | Flavonoid glycosides |
| 57 | Acteoside | 9.705 | C29H36O15 | [M-H]− | 623.1972 | 1.668 | Phenylglycosides |
| 58 | Quercetin-3-O-galactoside | 9.896 | C21H20O12 | [M-H]− | 463.0875 | 1.265 | Flavonoid glycosides |
| 59 | Ferulic acid † | 10.343 | C10H10O4 | [M-H]− | 193.0504 | 0.224 | Phenolics |
| 60 | Quercetin-3-O-pentosyl(1-2)acetylpentoside | 10.386 | C27H28O16 | [M-H]− | 607.1293 | 1.106 | Flavonoid glycosides |
| 61 | (6R,9S)-3-oxo-α-ionol glucoside | 10.517 | C19H30O7 | [M+HCOO]− | 415.1970 | 0.371 | Terpenoids |
| 62 | [6-[2-(3,4-dihydroxyphenyl)ethoxy]-2-(hydroxymethyl)-4-(3,4,5-trihydroxy-6-methyloxan-2-yl)oxy-5-(3,4,5-trihydroxyoxan-2-yl)oxyoxan-3-yl] (E)-3-(4-hydroxy-3-methoxyphenyl)prop-2-enoate | 10.529 | C35H46O19 | [M-H]− | 769.2544 | 1.693 | Phenylglycosides |
| 63 | Unknown | 10.716 | - | [M-H]− | 629.2650 | - | - |
| 64 | Quercetin-3-O-xyloside | 10.774 | C20H18O11 | [M-H]− | 433.0769 | 1.099 | Flavonoid glycosides |
| 65 | 3-[(2S,3R,4R,5S)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]oxy-2-(3,4-dihydroxyphenyl)-5,7-dihydroxychromen-4-one | 10.791 | C20H18O11 | [M-H]− | 433.07714 | 0.501 | Flavonoid glycosides |
| 66 | Kaempferol-3-O-glucoside † | 10.825 | C21H20O11 | [M-H]− | 447.09260 | 1.587 | Flavonoid glycosides |
| 67 | [(2R,3S,4S,5R,6S)-6-(3,4-dihydroxybenzoyl)oxy-3,4,5-trihydroxyoxan-2-yl]methyl 3,4-dihydroxybenzoate | 11.142 | C20H20O12 | [M-H]− | 451.2175 | 4.396 | Phenylglycosides |
| 68 | Salidroside | 11.185 | C14H20O7 | [M-H]− | 299.1138 | 0.188 | Glycosides |
| 69 | 6″-O-(3-Hydroxy-3-methylglutaroyl)astragalin | 11.426 | C27H28O15 | [M-H]− | 591.1347 | 0.491 | Glycosides |
| 70 | Azelaic acid † | 11.545 | C9H16O4 | [M-H]− | 187.0974 | 0.026 | Organic acids |
| 71 | 6-[(6,8-dihydroxy-7-methoxy-3-methyl-1-oxo-3,4-dihydroisochromen-4-yl)oxy]-4,8-dihydroxy-7-methoxy-3-methyl-3,4-dihydroisochromen-1-one | 11.741 | C22H22O11 | [M-H]− | 461.1085 | 0.392 | Quinones |
| 72 | Kaempferol-3-O-arabinoside † | 11.842 | C20H18O10 | [M-H]− | 417.0822 | 0.794 | Flavonoid glycosides |
| 73 | β-D-Glucopyranoside, 2-(3,4-dimethoxyphenyl)ethyl 3-O-(6-deoxy-alpha-L-mannopyranosyl)-4-O-[(2E)-3-(3,4-dihydroxyphenyl)-1-oxo-2-propen-1-yl]-6-O-[(2S,3R,4R)-tetrahydro-3,4-dihydroxy-4-(hydroxymethyl)-2-furanyl]- | 11.886 | C36H48O19 | [M-H]− | 783.2706 | 1.131 | Phenylglycosides |
| 74 | (5E)-3,4,9-trihydroxy-2-propyl-2,3,4,7,8,9-hexahydrooxecin-10-one | 11.986 | C12H20O5 | [M-H]− | 243.1237 | 0.047 | Polyketides |
| 75 | Tricin 5-glucoside | 12.020 | C23H24O12 | [M-H]− | 491.1197 | 0.233 | Flavonoid glycosides |
| 76 | Corylifol A | 12.032 | C25H26O4 | [M-H]− | 389.1808 | 4.979 | Flavonoids |
| 77 | 2-Hydroxyanthraquinone | 12.050 | C14H8O3 | [M-H]− | 223.0371 | 2.994 | Phenols |
| 78 | Phyllanthin † | 12.153 | C24H34O6 | [M-H]− | 417.2485 | 3.298 | Phenylglycosides |
| 79 | [5-hydroxy-6-[2-(4-hydroxy-3-methoxyphenyl)ethoxy]-2-(hydroxymethyl)-4-(3,4,5-trihydroxy-6-methyloxan-2-yl)oxyoxan-3-yl] (E)-3-(4-hydroxy-3-methoxyphenyl)prop-2-enoate | 12.377 | C31H40O15 | [M-H]− | 651.2297 | 0.266 | Phenylglycosides |
| 80 | [6-[2-(3,4-dihydroxyphenyl)-8-hydroxy-4-oxochromen-7-yl]oxy-3,4,5-trihydroxyoxan-2-yl]methyl (E)-3-(4-hydroxyphenyl)prop-2-enoate | 13.741 | C30H26O13 | [M-H]− | 593.1293 | 0.451 | Flavonoid glycosides |
| 81 | Tribuloside | 14.111 | C30H26O13 | [M-H]− | 593.1293 | 0.732 | Flavonoid glycosides |
| 82 | Unknown | 14.289 | - | [M-H]− | 507.2697 | - | - |
| 83 | Apigenin-7-neohesperidoside | 14.559 | C27H30O14 | [M-H]− | 577.1638 | 3.592 | Flavonoid glycosides |
| 84 | Procyanidin B2 | 14.943 | C30H26O12 | [M-H]− | 577.1346 | 0.451 | Proanthocyanidin |
| 85 | (10E,15Z)-9,12,13-trihydroxyoctadeca-10,15-dienoic acid | 15.402 | C18H32O5 | [M-H]− | 327.2173 | 0.880 | Fatty acids |
| 86 | [(2R,3S,4S,5R,6S)-6-[2,3-dihydroxy-4-[(E)-3-(4-hydroxyphenyl)prop-2-enoyl]phenoxy]-3,4,5-trihydroxyoxan-2-yl]methyl (E)-3-(4-hydroxyphenyl)prop-2-enoate | 15.452 | C30H28O12 | [M-H]− | 579.1497 | 0.571 | Chalcones |
| 87 | 1-(3,5-Dihydroxyphenyl)-12-hydroxytridecan-2-one | 15.531 | C19H30O4 | [M+HCOO]− | 367.2118 | 0.806 | Fatty acids |
| 88 | (Z)-9,12,13-trihydroxyoctadec-15-enoic acid | 16.462 | C18H34O5 | [M-H]− | 329.2329 | 0.984 | Fatty acids |
| 89 | Caperatic acid | 16.512 | C21H38O7 | [M-H]− | 803.5136 | 2.373 | Organic acids |
| 90 | Unknown | 16.824 | - | [M-H]− | 473.2751 | - | - |
| 91 | 2-(2-Acetyloxy-12-hydroxytridecyl)-4,6-dihydroxybenzoic acid | 17.308 | C22H34O7 | [M-H]− | 409.2223 | 1.304 | Organic acids |
| 92 | Unknown | 17.308 | - | [M-H]− | 819.4519 | - | - |
| 93 | 2-(2-Hydroxybut-3-en-2-yl)-3a,6,6,9a-tetramethyl-2,4,5,5a,7,8,9,9b-octahydro-1H-benzo[e][1]benzofuran-4,5-diol | 17.523 | C20H34O4 | [M-H]− | 337.2383 | 0.076 | Organic acids |
| 94 | [4,5-Dihydroxy-3,4-bis(hydroxymethyl)-4a,8,8-trimethyl-5,6,7,8a-tetrahydro-1H-naphthalen-1-yl] hexanoate | 17.523 | C21H36O6 | [M-H]− | 383.2437 | 0.180 | Fatty acids |
| 95 | Unknown | 18.063 | - | [M-H]− | 441.2488 | - | - |
| 96 | Aceroside VIII | 18.131 | C30H42O12 | [M-H]− | 593.2626 | 2.264 | Glycosides |
| 97 | Unknown | 18.295 | - | [M-H]− | 473.2752 | - | - |
| 98 | 9,10-Epoxy-13-oxo-11-octadecenoic acid | 18.343 | C18H30O4 | [M-H]− | 309.2070 | 1.042 | Fatty acids |
| 99 | Unknown | 18.372 | - | [M-H]− | 491.2750 | - | - |
| 100 | 2-(3-(8-Hydroxyoctyl)phenoxy)-2-methylpropanoic acid | 19.466 | C18H28O4 | [M-H]− | 307.1910 | 0.409 | Fatty acids |
| 101 | Forskolin | 19.687 | C22H34O7 | [M-H]− | 409.2223 | 0.763 | Diterpenes |
| 102 | 3″,4″-Di-O-p-coumaroylafzelin | 19.870 | C39H32O14 | [M-H]− | 723.1699 | 1.943 | Flavonoids |
| 103 | Unknown | 20.081 | - | [M-H]− | 489.2597 | - | - |
| 104 | 3-[(2S,3R,4S,5S,6R)-3-[(2S,3R,4R)-3,4-dihydroxy-4-(hydroxymethyl)oxolan-2-yl]oxy-4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-5-hydroxy-2-(4-hydroxyphenyl)-7-[(2S,3R,4R,5R,6S)-3,4,5-trihydroxy-6-methyloxan-2-yl]oxychromen-4-one | 20.148 | C32H38O19 | [M-H]− | 725.1878 | 5.683 | Flavonoid glycosides |
| 105 | Biochanin A | 20.437 | C16H12O5 | [M-H]− | 283.0607 | 0.407 | Isoflavone |
| 106 | 1-Epi coleonol | 20.475 | C22H34O7 | [M-H]− | 409.2224 | 0.793 | Terpenoids |
| 107 | Amorfrutin | 20.581 | C18H26O4 | [M-H]− | 305.1754 | 0.378 | Terpenoids |
| 108 | [4,5-dihydroxy-3,4-bis(hydroxymethyl)-4a,8,8-trimethyl-5,6,7,8a-tetrahydro-1H-naphthalen-1-yl] octanoate | 21.023 | C23H40O6 | [M-H]− | 457.2798 | 0.867 | Fatty acids |
| 109 | Rotenone | 21.292 | C23H22O6 | [M-H]− | 393.2271 | 0.143 | Isoflavone |
| 110 | Nandrolone | 22.280 | C18H26O2 | [M-H2O-H]− | 275.2015 | 1.519 | Steroids |
| 111 | 15,16-Epoxy-9,12-octadecadienoic acid | 22.290 | C18H30O3 | [M-H]− | 293.2120 | 0.271 | Fatty acids |
| 112 | 2-Hydroxy-4,5′,8a′-trimethyl-1′-oxo-4-vinyloctahydro-1′H-spiro[cyclopentane-1,2′-naphthalene]-5′-carboxylic acid | 22.588 | C20H30O4 | [M-H]− | 333.2071 | 0.005 | Organic acids |
| 113 | Grayanotoxin I | 22.830 | C22H36O7 | [M-H2O-H]− | 393.2276 | 0.176 | Diterpenes |
| 114 | 9-Hydroxy-10E,12Z-octadecadienoic acid | 22.962 | C18H32O3 | [M-H]− | 295.2272 | 0.636 | Fatty acids |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Le, D.D.; Roh, N.; Lee, J.; Shrestha, D.; Dinh, T.; Truong, V.; Bazarragchaa, B.; Kim, S.-Y.; Suh, S.-S.; et al. Chemical Composition and Biological Activities of Lagopsis supina Extract: Antioxidant, Adipogenic, and Ani-Inflammatory Effects. Pharmaceuticals 2025, 18, 150. https://doi.org/10.3390/ph18020150
Choi J, Le DD, Roh N, Lee J, Shrestha D, Dinh T, Truong V, Bazarragchaa B, Kim S-Y, Suh S-S, et al. Chemical Composition and Biological Activities of Lagopsis supina Extract: Antioxidant, Adipogenic, and Ani-Inflammatory Effects. Pharmaceuticals. 2025; 18(2):150. https://doi.org/10.3390/ph18020150
Chicago/Turabian StyleChoi, Juhyun, Duc Dat Le, Nayoung Roh, Jiseok Lee, Deumaya Shrestha, Thientam Dinh, Vinhquang Truong, Badamtsetseg Bazarragchaa, Soo-Yong Kim, Sung-Suk Suh, and et al. 2025. "Chemical Composition and Biological Activities of Lagopsis supina Extract: Antioxidant, Adipogenic, and Ani-Inflammatory Effects" Pharmaceuticals 18, no. 2: 150. https://doi.org/10.3390/ph18020150
APA StyleChoi, J., Le, D. D., Roh, N., Lee, J., Shrestha, D., Dinh, T., Truong, V., Bazarragchaa, B., Kim, S.-Y., Suh, S.-S., Lee, M., & Seo, J. B. (2025). Chemical Composition and Biological Activities of Lagopsis supina Extract: Antioxidant, Adipogenic, and Ani-Inflammatory Effects. Pharmaceuticals, 18(2), 150. https://doi.org/10.3390/ph18020150