
TRPV1 Activation Antagonizes High-Fat Diet-Induced Obesity at Thermoneutrality and Enhances UCP-1 Transcription via PRDM-16

Abstract

1. Introduction
2. Results
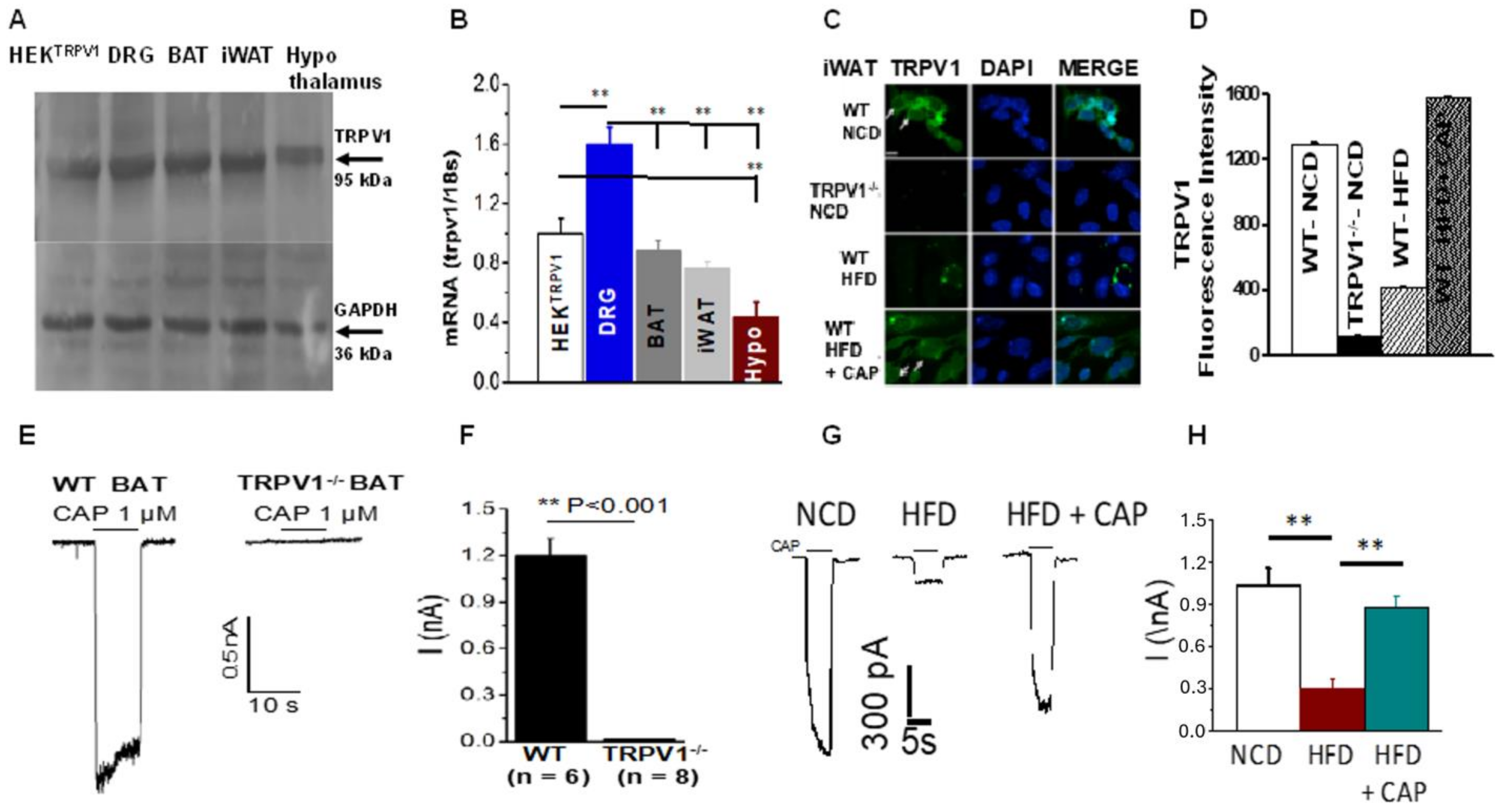
2.1. TRPV1 Expression and Activity in BAT and iWAT
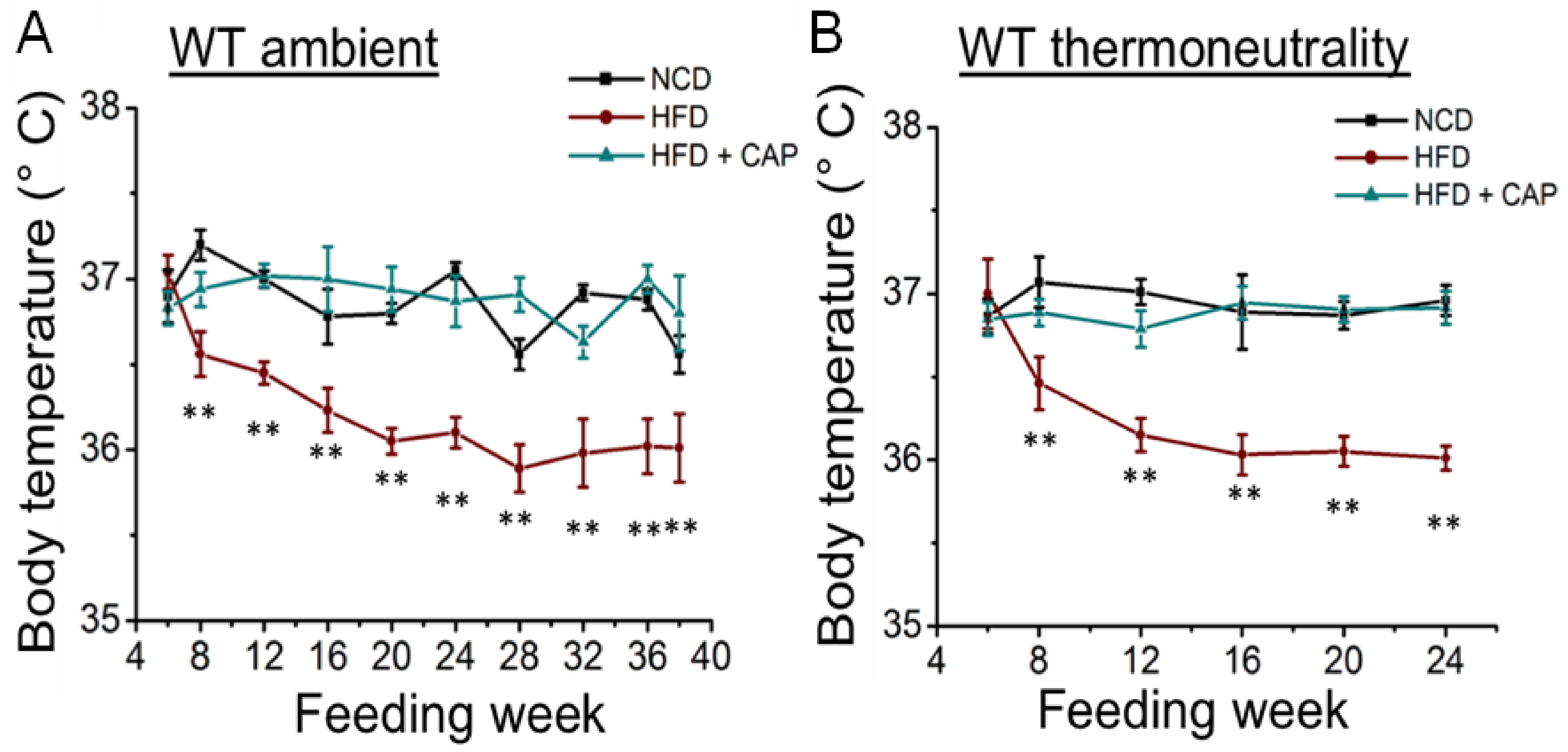
2.2. Decreased Body Temperature in Diet-Induced Obese WT Mice at Ambient Temperature and in the Thermoneutral Zone
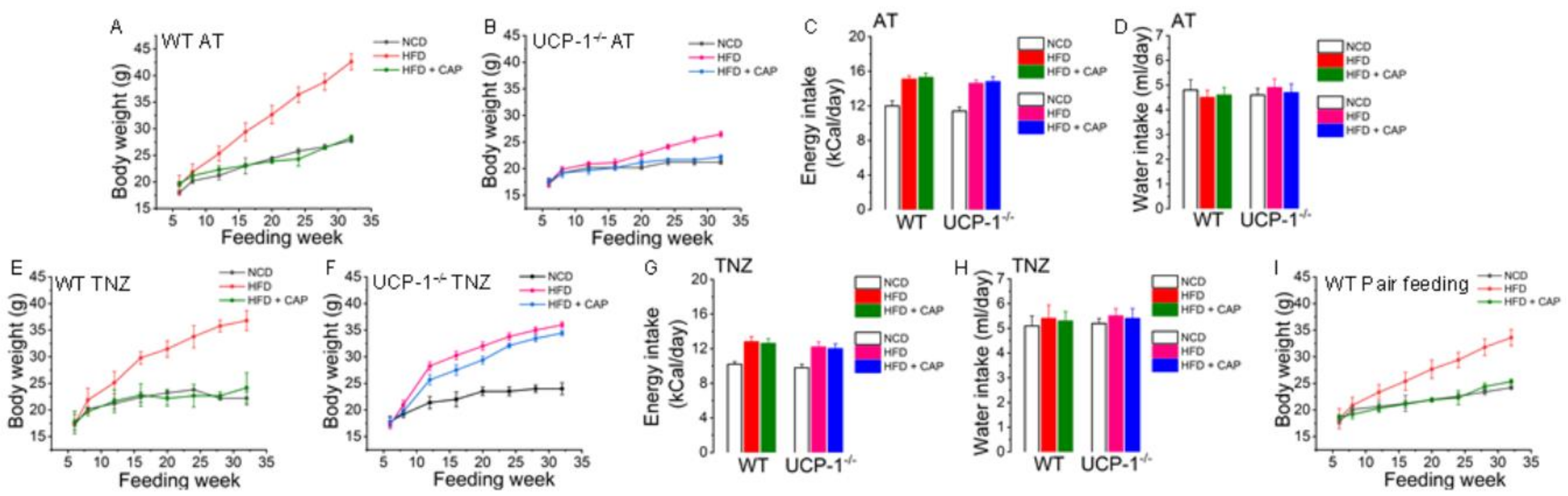
2.3. CAP Inhibited Weight Gain in WT and UCP1−/− Mice at Ambient Temperature but Inhibited HFD-Induced Weight Gain Only in WT Mice and Not in UCP−/− Mice in the TNZ
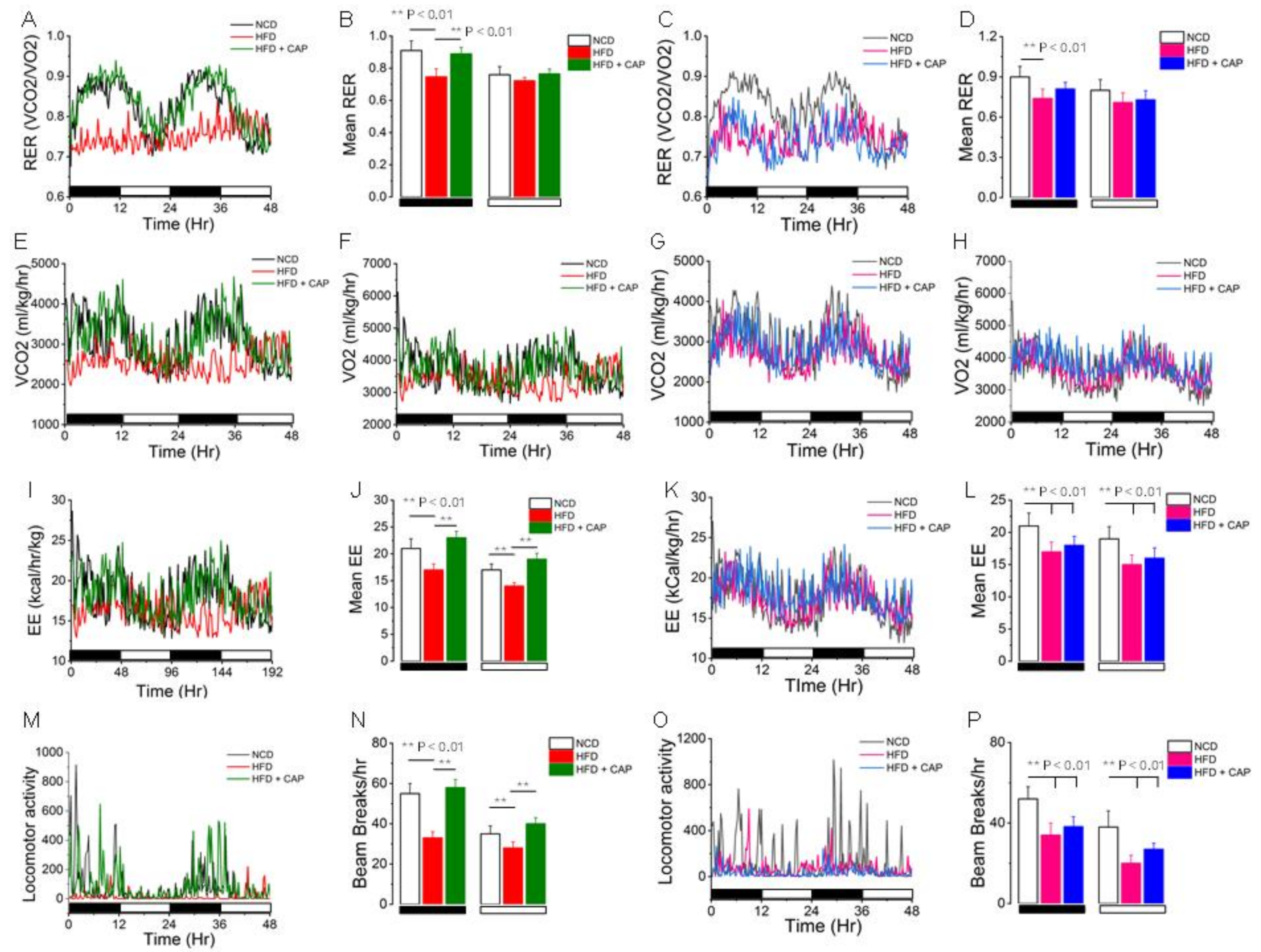
2.4. Metabolic Activity
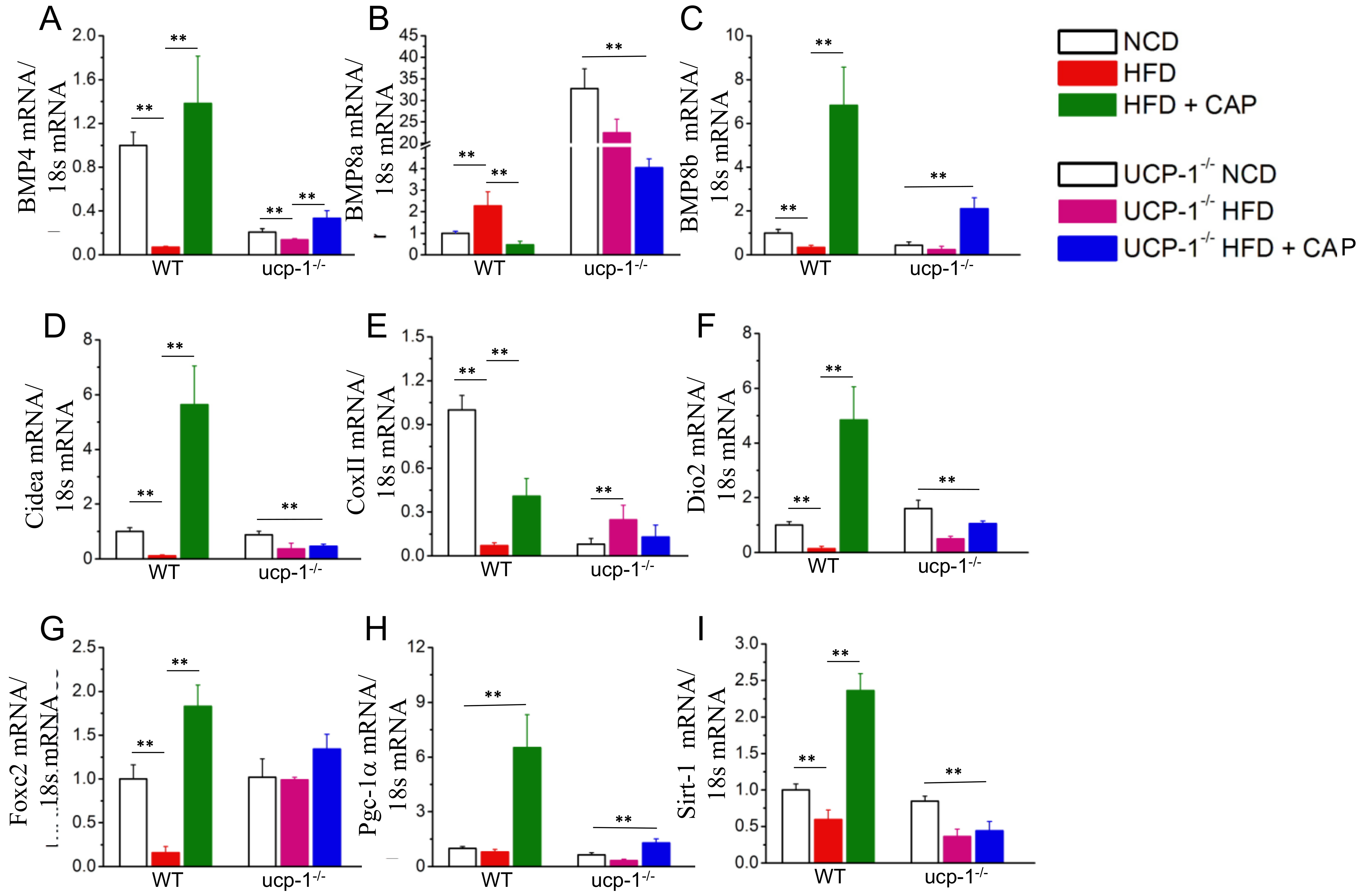
2.5. White Adipogenic and Thermogenic Gene Expression
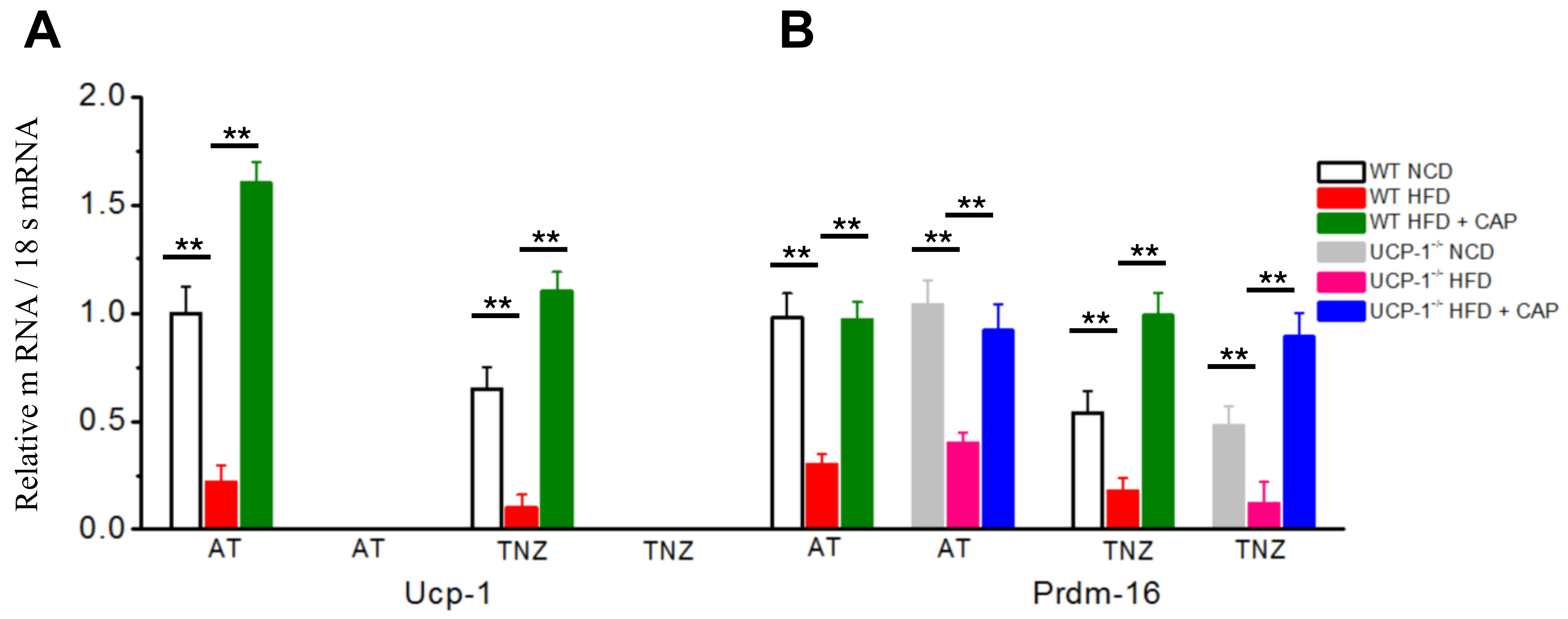
2.6. UCP-1 and PRDM-16 Gene Expression
2.7. CAP Fails to Counter HFD-Induced Weight Gain in Adipose Tissue-Specific PRDM16−/− Mice
2.8. PRDM-16 Overexpression Upregulates UCP-1 Transcription in HEK TRPV1 Cells
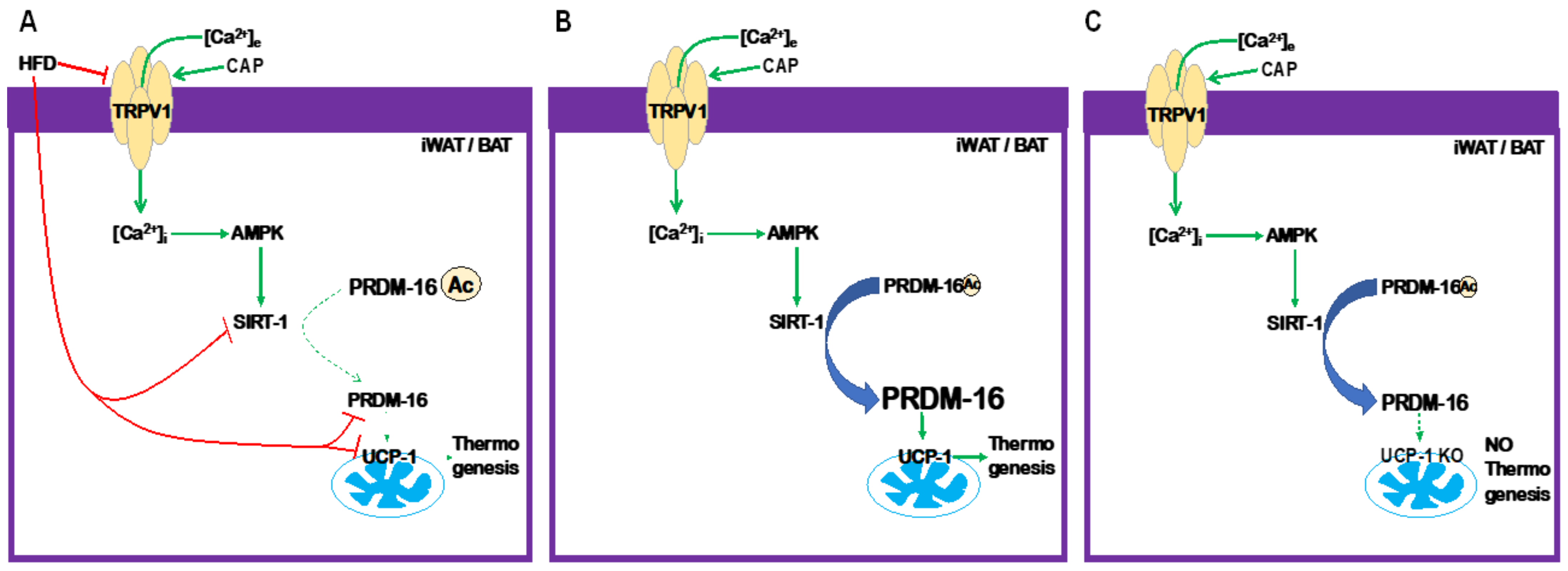
3. Discussion
4. Materials and Methods
4.1. Mouse Model of High-Fat Diet-Induced Obesity
4.2. Adipose Tissue-Specific PRDM-16 Knockout Mice
4.3. Measurement of Rectal Temperature in Mice
4.4. Metabolic Activity Measurement
4.5. Fat Tissue Isolation [43]
4.6. Isolation and Primary Culture of Brown Adipose Tissue Preadipocytes
4.7. Immunohistochemistry
4.8. Whole-Cell Patch Clamp of Primary Brown Adipocytes
4.9. Quantitative RT-PCR Measurements (qRT-PCR)
4.10. Immunoblotting
4.11. PRDM16 and GFP-UCP1 Overexpression and Stimulation in the HEK TRPV1 Cell Line
4.12. Drugs and Chemicals
4.13. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Troynikov, O.; Watson, C.G.; Nawaz, N. Sleep environments and sleep physiology: A review. J. Therm. Biol. 2018, 78, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Someren, E.J.W.V. Age-Related Changes in Thermoreception and Thermoregulation. Available online: https://www.sciencedirect.com/science/article/abs/pii/B9780123786388000221 (accessed on 9 August 2024).
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. The Importance of Energy Balance. Eur. Endocrinol. 2013, 9, 111–115. [Google Scholar]
- Manini, T.M. Energy expenditure and aging. Ageing Res. Rev. 2010, 9, 1. [Google Scholar]
- Tremblay, A.; Royer, M.M.; Chaput, J.P.; Doucet, E. Adaptive thermogenesis can make a difference in the ability of obese individuals to lose body weight. Int. J. Obes. (Lond.) 2013, 37, 759–764. [Google Scholar] [PubMed]
- Ravussin, Y.; LeDuc, C.A.; Watanabe, K.; Leibel, R.L. Effects of ambient temperature on adaptive thermogenesis during maintenance of reduced body weight in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R438–R448. [Google Scholar] [PubMed]
- Klingenspor, M.; Bast, A.; Bolze, F.; Li, Y.; Maurer, S.; Schweizer, S.; Willershäuser, M.; Fromme, T. Brown Adipose Tissue. 2017. Available online: https://portal.fis.tum.de/en/publications/brown-adipose-tissue-2 (accessed on 9 August 2024).
- Fedorenko, A.; Lishko, P.V.; Kirichok, Y. Mechanism of fatty-acid-dependent UCP1 uncoupling in brown fat mitochondria. Cell 2012, 151, 400–413. [Google Scholar] [CrossRef]
- Razzoli, M.; Emmett, M.J.; Lazar, M.A.; Bartolomucci, A. β-Adrenergic receptors control brown adipose UCP-1 tone and cold response without affecting its circadian rhythmicity. FASEB J. 2018, 32, 5640–5646. [Google Scholar] [CrossRef]
- John, L.M.; Petersen, N.; Gerstenberg, M.K.; Torz, L.; Pedersen, K.; Christoffersen, B.; Kuhre, R.E. Housing-temperature reveals energy intake counter-balances energy expenditure in normal-weight, but not diet-induced obese, male mice. Commun. Biol. 2022, 5, 946. [Google Scholar] [CrossRef]
- Cui, X.; Nguyen, N.L.; Zarebidaki, E.; Cao, Q.; Li, F.; Zha, L.; Bartness, T.; Shi, H.; Xue, B. Thermoneutrality decreases thermogenic program and promotes adiposity in high-fat diet-fed mice. Physiol. Rep. 2016, 4, e12799. [Google Scholar] [CrossRef]
- Dieckmann, S.; Strohmeyer, A.; Willershäuser, M.; Maurer, S.F.; Wurst, W.; Marschall, S.; de Angelis, M.H.; Kühn, R.; Worthmann, A.; Fuh, M.M.; et al. Susceptibility to diet-induced obesity at thermoneutral conditions is independent of UCP1. Am. J. Physiol. Endocrinol. Metab. 2022, 322, e85–e100. [Google Scholar] [CrossRef]
- Baskaran, P.; Krishnan, V.; Fettel, K.; Gao, P.; Zhu, Z.; Ren, J.; Thyagarajan, B. TRPV1 activation counters diet-induced obesity through sirtuin-1 activation and PRDM-16 deacetylation in brown adipose tissue. Int. J. Obes. (Lond.) 2017, 41, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, P.; Krishnan, V.; Ren, J.; Thyagarajan, B. Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br. J. Pharmacol. 2016, 173, 2369–2389. [Google Scholar] [CrossRef] [PubMed]
- Kingma, B.R.; Frijns, A.J.; Schellen, L.; van Marken Lichtenbelt, W.D. Beyond the classic thermoneutral zone: Including thermal comfort. Temperature 2014, 1, 142–149. [Google Scholar] [CrossRef]
- Kingma, B.; Frijns, A.; van Marken Lichtenbelt, W. The thermoneutral zone: Implications for metabolic studies. Front. Biosci. 2012, 4, 1975–1985. [Google Scholar] [CrossRef]
- Lehr, L.; Canola, K.; Asensio, C.; Jimenez, M.; Kuehne, F.; Giacobino, J.P.; Muzzin, P. The control of UCP1 is dissociated from that of PGC-1alpha or of mitochondriogenesis as revealed by a study using beta-less mouse brown adipocytes in culture. FEBS Lett. 2006, 580, 4661–4666. [Google Scholar] [CrossRef]
- Teruel, T.; Clapham, J.C.; Smith, S.A. PPARalpha activation by Wy 14643 induces transactivation of the rat UCP-1 promoter without increasing UCP-1 mRNA levels and attenuates PPARgamma-mediated increases in UCP-1 mRNA levels induced by rosiglitazone in fetal rat brown adipocytes. Biochem. Biophys. Res. Commun. 1999, 264, 311–315. [Google Scholar] [CrossRef]
- Barbera, M.J.; Schluter, A.; Pedraza, N.; Iglesias, R.; Villarroya, F.; Giralt, M. Peroxisome proliferator-activated receptor alpha activates transcription of the brown fat uncoupling protein-1 gene. A link between regulation of the thermogenic and lipid oxidation pathways in the brown fat cell. J. Biol. Chem. 2001, 276, 1486–1493. [Google Scholar] [CrossRef]
- Iida, S.; Chen, W.; Nakadai, T.; Ohkuma, Y.; Roeder, R.G. PRDM16 enhances nuclear receptor-dependent transcription of the brown fat-specific Ucp1 gene through interactions with Mediator subunit MED1. Genes. Dev. 2015, 29, 308–321. [Google Scholar] [CrossRef]
- Baskaran, P.; Markert, L.; Bennis, J.; Zimmerman, L.; Fox, J.; Thyagarajan, B. Assessment of Pharmacology, Safety, and Metabolic activity of Capsaicin Feeding in Mice. Sci. Rep. 2019, 9, 8588. [Google Scholar] [CrossRef]
- Redinger, R.N. The pathophysiology of obesity and its clinical manifestations. Gastroenterol. Hepatol. (NY) 2007, 3, 856–863. [Google Scholar]
- NIDDK. Overweight & Obesity Statistics. Available online: https://www.niddk.nih.gov/health-information/health-statistics/overweight-obesity (accessed on 9 August 2024).
- Betz, M.J.; Enerbäck, S. Human Brown Adipose Tissue: What We Have Learned So Far. Diabetes 2015, 64, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Abete, I. Genetic Regulation of Energy Homeostasis: Obesity Implications. Available online: https://www.biorxiv.org/content/10.1101/2020.07.27.223529v1.full (accessed on 9 August 2024).
- Chouchani, E.T.; Kazak, L.; Spiegelman, B.M. New Advances in Adaptive Thermogenesis: UCP1 and Beyond. Cell Metab. 2019, 29, 27–37. [Google Scholar] [CrossRef]
- Kim, K.; Wann, J.; Kim, H.G.; So, J.; Rosen, E.D.; Roh, H.C. Uncoupling protein 1-driven Cre (Ucp1-Cre) is expressed in the epithelial cells of mammary glands and various non-adipose tissues. bioRxiv 2023. [Google Scholar] [CrossRef]
- Enerbäck, S.; Jacobsson, A.; Simpson, E.M.; Guerra, C.; Yamashita, H.; Harper, M.E.; Kozak, L.P. Mice lacking mitochondrial uncoupling protein are cold-sensitive but not obese. Nature 1997, 387, 90–94. [Google Scholar] [CrossRef]
- Liu, X.; Rossmeisl, M.; McClaine, J.; Riachi, M.; Harper, M.E.; Kozak, L.P. Paradoxical resistance to diet-induced obesity in UCP1-deficient mice. J. Clin. Investig. 2003, 111, 399–407. [Google Scholar] [CrossRef]
- Li, Y.; Fromme, T. Uncoupling Protein 1 Does Not Produce Heat without Activation. Int. J. Mol. Sci. 2022, 23, 2406. [Google Scholar] [CrossRef]
- Qian, S.W.; Tang, Y.; Li, X.; Liu, Y.; Zhang, Y.Y.; Huang, H.Y.; Xue, R.D.; Yu, H.Y.; Guo, L.; Gao, H.D.; et al. BMP4-mediated brown fat-like changes in white adipose tissue alter glucose and energy homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, E798–E807. [Google Scholar] [CrossRef]
- Whittle, A.J.; Carobbio, S.; Martins, L.; Slawik, M.; Hondares, E.; Vázquez, M.J.; Morgan, D.; Csikasz, R.I.; Gallego, R.; Rodriguez-Cuenca, S.; et al. BMP8B increases brown adipose tissue thermogenesis through both central and peripheral actions. Cell 2012, 149, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Jash, S.; Banerjee, S.; Lee, M.J.; Farmer, S.R.; Puri, V. CIDEA Transcriptionally Regulates UCP1 for Britening and Thermogenesis in Human Fat Cells. iScience 2019, 20, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Banhos Danneskiold-Samsøe, N.; Sonne, S.B.; Larsen, J.M.; Hansen, A.N.; Fjære, E.; Isidor, M.S.; Petersen, S.; Henningsen, J.; Severi, I.; Sartini, L.; et al. Overexpression of cyclooxygenase-2 in adipocytes reduces fat accumulation in inguinal white adipose tissue and hepatic steatosis in high-fat fed mice. Sci. Rep. 2019, 9, 8979. [Google Scholar] [CrossRef]
- Tabuchi, C.; Sul, H.S. Signaling Pathways Regulating Thermogenesis. Front. Endocrinol. (Lausanne) 2021, 12, 595020. [Google Scholar] [CrossRef]
- Gan, L.; Liu, Z.; Chen, Y.; Dan, L.; Feng, F.; Liu, G.; Sun, C. α-MSH and Foxc2 promote fatty acid oxidation through C/EBPβ negative transcription in mice adipose tissue. Sci. Rep. 2016, 6, 36661. [Google Scholar] [CrossRef]
- Uldry, M.; Yang, W.; St-Pierre, J.; Lin, J.; Seale, P.; Spiegelman, B.M. Complementary action of the PGC-1 coactivators in mitochondrial biogenesis and brown fat differentiation. Cell Metab. 2006, 3, 333–341. [Google Scholar] [CrossRef]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown remodeling of white adipose tissue by SirT1-dependent deacetylation of Pparγ. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.J.; Ishibashi, J.; Wang, W.; Lim, H.W.; Goyama, S.; Sato, T.; Kurokawa, M.; Won, K.J.; Seale, P. Prdm16 is required for the maintenance of brown adipocyte identity and function in adult mice. Cell Metab. 2014, 19, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Levy, J.D.; Zhang, Y.; Frontini, A.; Kolodin, D.P.; Svensson, K.J.; Lo, J.C.; Zeng, X.; Ye, L.; Khandekar, M.J.; et al. Ablation of PRDM16 and beige adipose causes metabolic dysfunction and a subcutaneous to visceral fat switch. Cell 2014, 156, 304–316. [Google Scholar]
- Turdi, S.; Kandadi, M.R.; Zhao, J.; Huff, A.F.; Du, M.; Ren, J. Deficiency in AMP-activated protein kinase exaggerates high fat diet-induced cardiac hypertrophy and contractile dysfunction. J. Mol. Cell Cardiol. 2011, 50, 712–722. [Google Scholar] [CrossRef]
- Ren, J. Leptin and hyperleptinemia—From friend to foe for cardiovascular function. J. Endocrinol. 2004, 181, 1–10. [Google Scholar]
- Bagchi, D.P.; MacDougald, O.A. Identification and Dissection of Diverse Mouse Adipose Depots. J. Vis. Exp. 2019. [Google Scholar] [CrossRef]
- Thyagarajan, B.; Lukacs, V.; Rohacs, T. Hydrolysis of phosphatidylinositol 4,5-bisphosphate mediates calcium-induced inactivation of TRPV6 channels. J. Biol. Chem. 2008, 283, 14980–14987. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession Number) | Forward Primer | Reverse Primer |
|---|---|---|
| 18s (X00686) | accgcagctaggaataatgga | gcctcagttccgaaaacca |
| Gapdh (NM_001411844) | cgtgccgcctggagaaacc | tggaagagtgggagttgctgttg |
| mtrpv1 (NM_001001445) | caacaagaaggggcttacacc | tctggagaatgtaggccaagac |
| sirt-1 (NM_019812) | tcgtggagacatttttaatcagg | gcttcatgatggcaagtgg |
| prdm-16 (XM_03616442) | cagcacggtgaagccattc | gcgtgcatccgcttgtg |
| ucp-1 (NM_009463) | cgactcagtccaagagtacttctcttc | gccggctgagatcttgtttc |
| pgc-1α (NR_175325) | agagaggcagaagcagaaagcaat | attctgtccgcgttgtgtcagg |
| Cidea (NM_007702) | atcacaactggcctggttacg | tactacccggtgtccatttct |
| cox2 (AF378830) | ataaccgagtcgttctgccaat | tttcagagcattggccatagaa |
| foxc2 (NM_013519) | gcaacccaacagcaaactttc | gacggcgtagctcgatagg |
| dio2 (NM_010050) | gttgcttctgagccgctc | gctctgcactggcaaagtc |
| bmp4(NM_007554) | ctccagtctggggaggag | gatgaggtgcccaggcac |
| bmp8a (NM_007558) | aaccatgccatcttgcagtct | cagaggtggcactcagtttgg |
| bmp8b (NM_007559) | tccaccaaccacgccactat | cagtaggcacacagcacacct |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baskaran, P.; Gustafson, N.; Chavez, N. TRPV1 Activation Antagonizes High-Fat Diet-Induced Obesity at Thermoneutrality and Enhances UCP-1 Transcription via PRDM-16. Pharmaceuticals 2024, 17, 1098. https://doi.org/10.3390/ph17081098
Baskaran P, Gustafson N, Chavez N. TRPV1 Activation Antagonizes High-Fat Diet-Induced Obesity at Thermoneutrality and Enhances UCP-1 Transcription via PRDM-16. Pharmaceuticals. 2024; 17(8):1098. https://doi.org/10.3390/ph17081098
Chicago/Turabian StyleBaskaran, Padmamalini, Noah Gustafson, and Nicolas Chavez. 2024. "TRPV1 Activation Antagonizes High-Fat Diet-Induced Obesity at Thermoneutrality and Enhances UCP-1 Transcription via PRDM-16" Pharmaceuticals 17, no. 8: 1098. https://doi.org/10.3390/ph17081098
APA StyleBaskaran, P., Gustafson, N., & Chavez, N. (2024). TRPV1 Activation Antagonizes High-Fat Diet-Induced Obesity at Thermoneutrality and Enhances UCP-1 Transcription via PRDM-16. Pharmaceuticals, 17(8), 1098. https://doi.org/10.3390/ph17081098