



Cancer Cell Secreted Legumain Promotes Gastric Cancer Resistance to Anti-PD-1 Immunotherapy by Enhancing Macrophage M2 Polarization
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
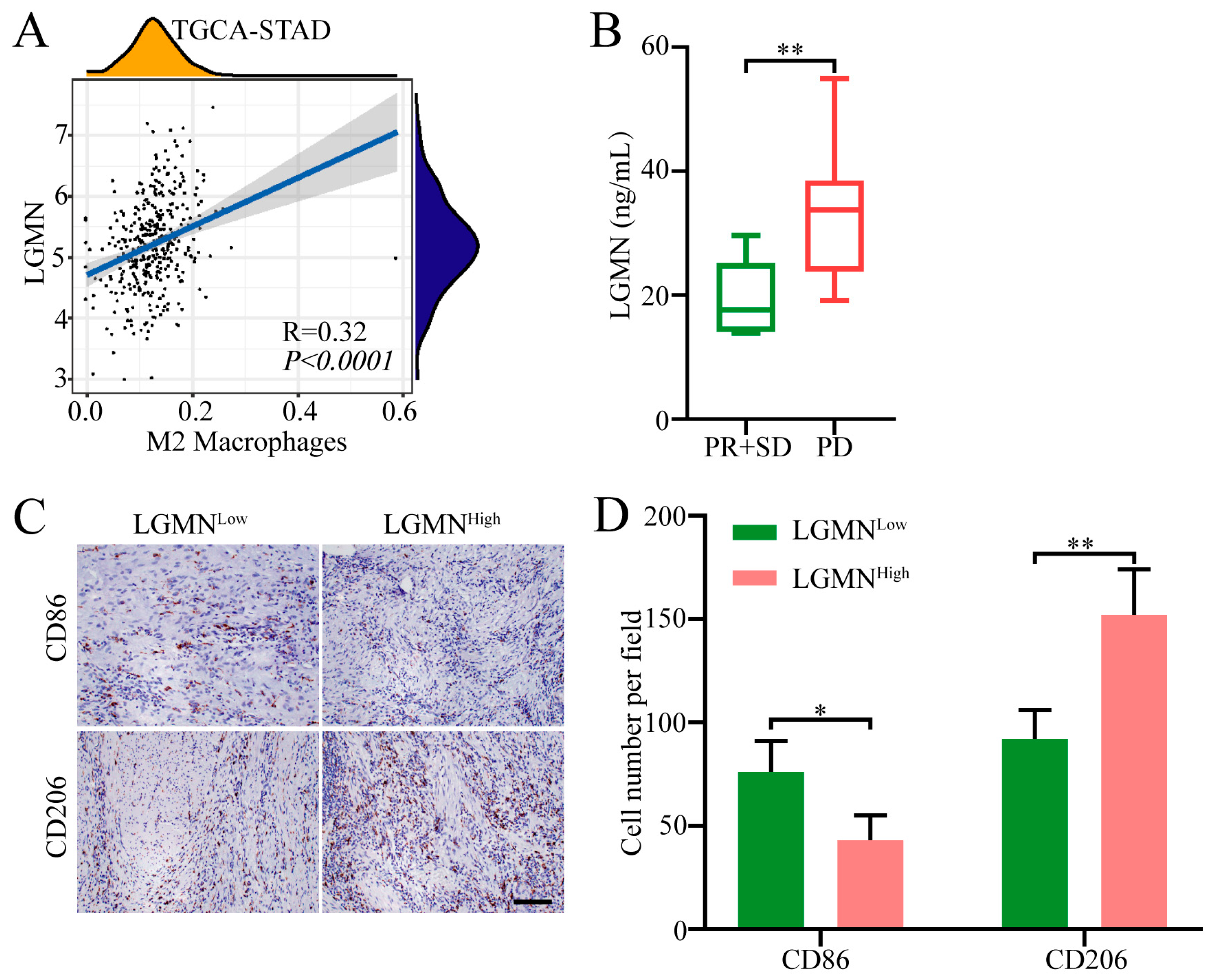
2.1. Forced Serum LGMN Expression Is Associated with Increased M2 Macrophages in GC Tissues
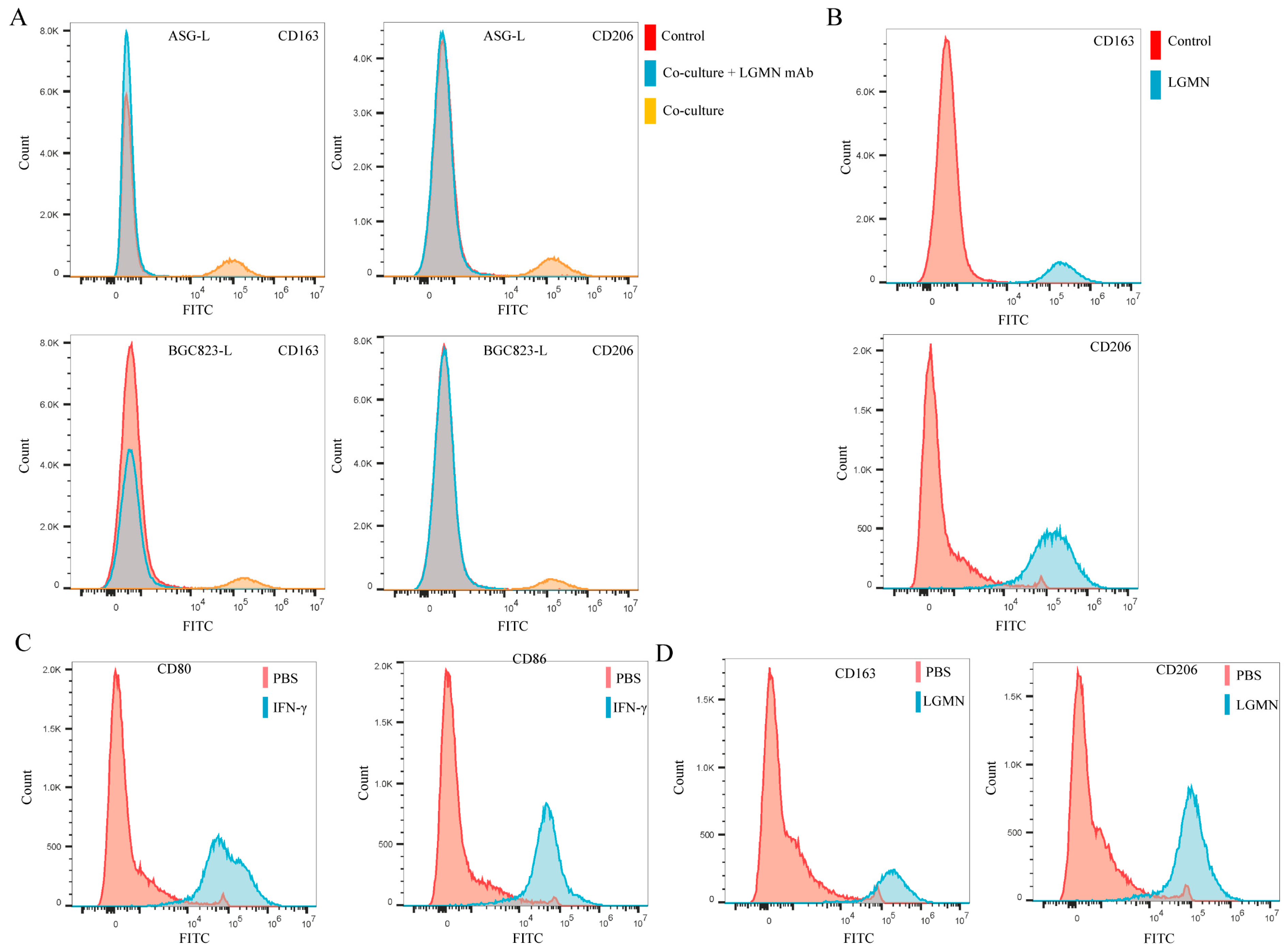
2.2. LGMN Promotes M1 Macrophages Polarization towards M2 Phenotype
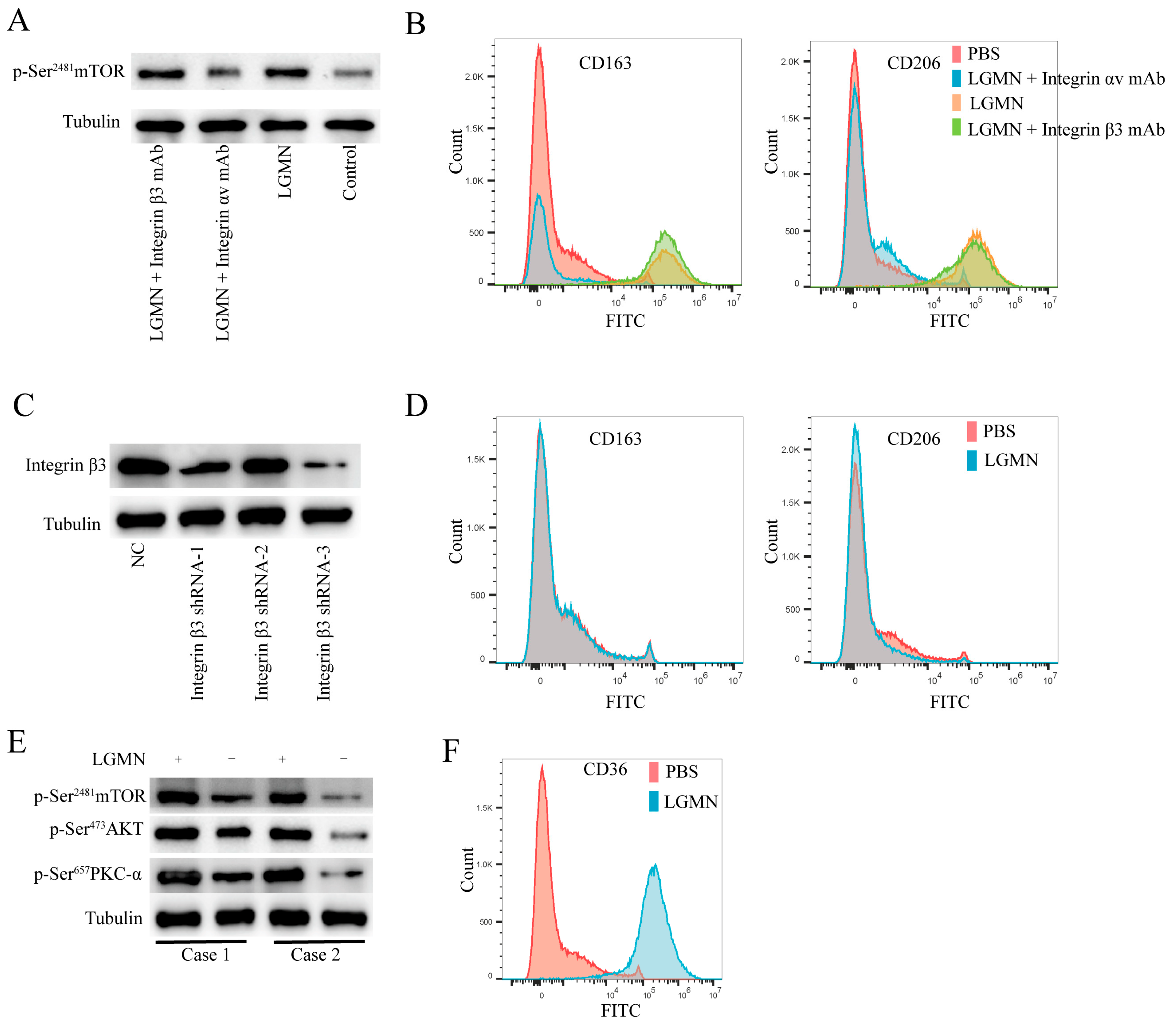
2.3. LGMN Upregulates Integrin αvβ3/AKT (Serine/Threonine Kinase)/mTORC2 Axis Signaling in Macrophages
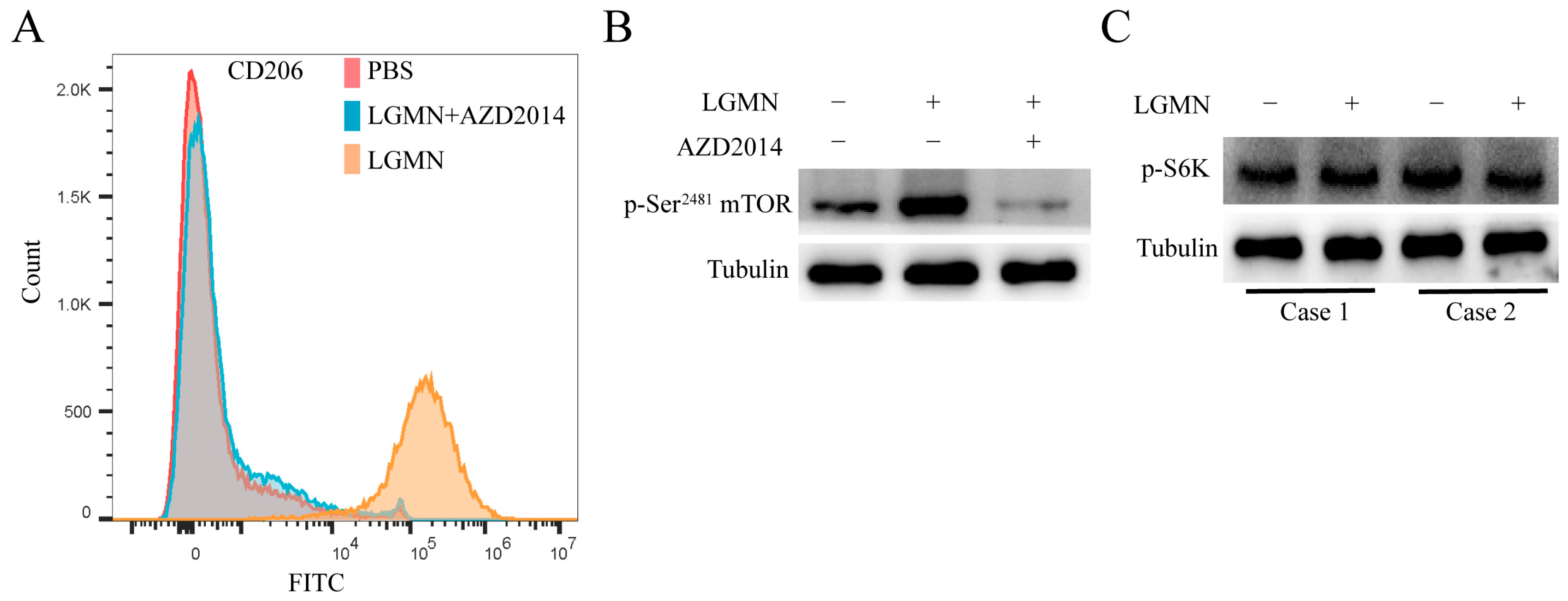
2.4. mTOR (Mammalian Target of Rapamycin) Inhibitors Block LGMN-Induced Polarization of M1 Macrophages towards M2
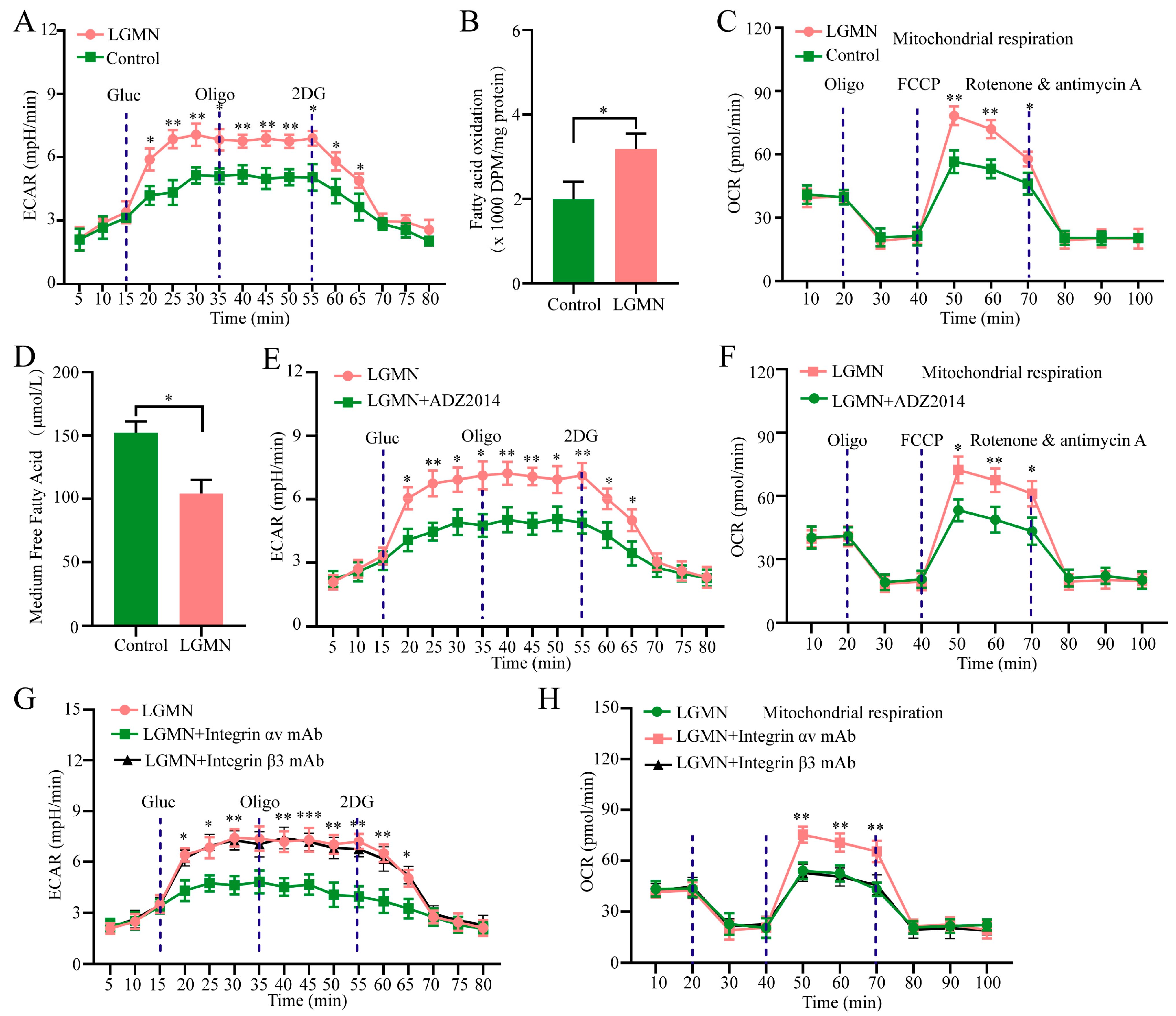
2.5. LGMN Induces Metabolic Reprogramming of M1 Macrophages via Integrin αvβ3/AKT/mTORC2 axis
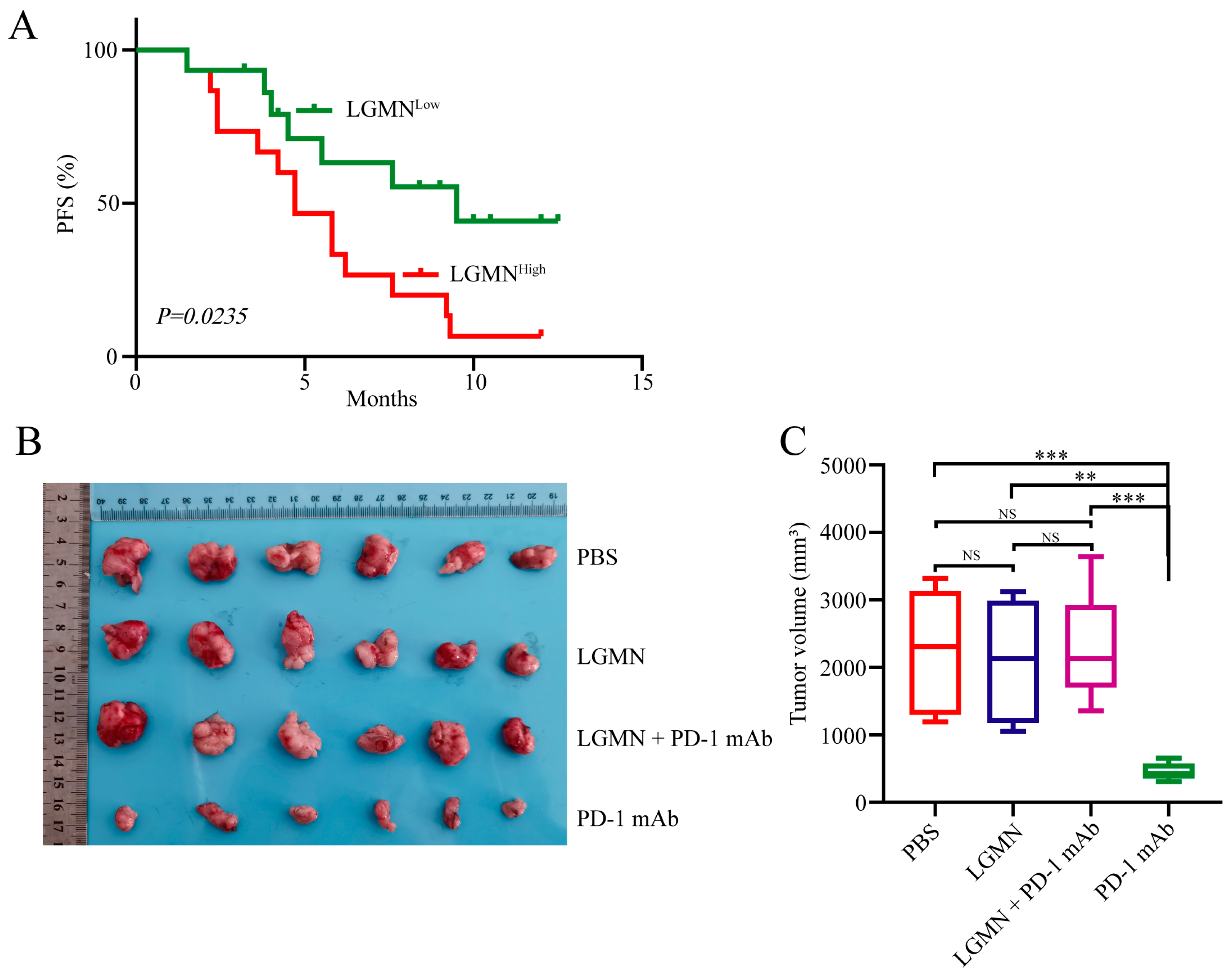
2.6. sLGMN Promotes Gastric Cancer Resistance to Anti-PD-1 Immunotherapy
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Clinical Tissues
4.2. Immunohistochemistry
4.3. Transfection Experiment
4.4. Western Blot
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Extracellular Acidification Rate (ECAR) Analysis
4.7. Flow Cytometry Analysis
4.8. Lactic Acid Measurement
4.9. Non-Targeted Metabolomics Sequencing Analysis
4.10. ROS Production Assay
4.11. Animals
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Janjigian, Y.Y.; Shitara, K.; Moehler, M.; Garrido, M.; Salman, P.; Shen, L.; Wyrwicz, L.; Yamaguchi, K.; Skoczylas, T.; Campos Bragagnoli, A.; et al. First-line nivolumab plus chemotherapy versus chemotherapy alone for advanced gastric, gastro-oesophageal junction, and oesophageal adenocarcinoma (CheckMate 649): A randomised, open-label, phase 3 trial. Lancet 2021, 398, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.K.; Boku, N.; Satoh, T.; Ryu, M.H.; Chao, Y.; Kato, K.; Chung, H.C.; Chen, J.S.; Muro, K.; Kang, W.K.; et al. Nivolumab in patients with advanced gastric or gastro-oesophageal junction cancer refractory to, or intolerant of, at least two previous chemotherapy regimens (ONO-4538-12, ATTRACTION-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 390, 2461–2471. [Google Scholar] [CrossRef]
- Takei, S.; Kawazoe, A.; Shitara, K. The New Era of Immunotherapy in Gastric Cancer. Cancers 2022, 14, 1054. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Hou, J.; Wu, X.; Lu, Y.; Jin, Z.; Yu, Z.; Yu, B.; Li, J.; Yang, Z.; Li, C.; et al. Cancer-associated fibroblasts impair the cytotoxic function of NK cells in gastric cancer by inducing ferroptosis via iron regulation. Redox Biol. 2023, 67, 102923. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Lin, Y.; Gan, L.; Wang, S.; Chen, S.; Li, C.; Hou, S.; Hu, B.; Wang, B.; Ye, Y.; et al. Potential crosstalk between SPP1 + TAMs and CD8+ exhausted T cells promotes an immunosuppressive environment in gastric metastatic cancer. J. Transl. Med. 2024, 22, 158. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, J.; Li, W.; Feng, C.; Liu, Q.; Gao, W.; He, M. Immunology and immunotherapy in gastric cancer. Clin. Exp. Med. 2023, 23, 3189–3204. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.W.; Li, Y.; Yang, Y.; Yang, H.K.; Dong, J.M.; Xiao, Z.H.; He, X.; Guo, J.H.; Wang, R.Q.; Dai, B.; et al. Tumor immunotherapy resistance: Revealing the mechanism of PD-1/PD-L1-mediated tumor immune escape. Biomed. Pharmacother. 2024, 171, 116203. [Google Scholar] [CrossRef] [PubMed]
- Haugen, M.H.; Johansen, H.T.; Pettersen, S.J.; Solberg, R.; Brix, K.; Flatmark, K.; Maelandsmo, G.M. Nuclear legumain activity in colorectal cancer. PLoS ONE 2013, 8, e52980. [Google Scholar] [CrossRef]
- Liu, C.; Wang, J.; Zheng, Y.; Zhu, Y.; Zhou, Z.; Liu, Z.; Lin, C.; Wan, Y.; Wen, Y.; Liu, C.; et al. Autocrine pro-legumain promotes breast cancer metastasis via binding to integrin αvβ3. Oncogene 2022, 41, 4091–4103. [Google Scholar] [CrossRef]
- Bai, P.; Lyu, L.; Yu, T.; Zuo, C.; Fu, J.; He, Y.; Wan, Q.; Wan, N.; Jia, D.; Lyu, A. Macrophage-Derived Legumain Promotes Pulmonary Hypertension by Activating the MMP (Matrix Metalloproteinase)-2/TGF (Transforming Growth Factor)-β1 Signaling. Arterioscler. Thromb. Vasc. Biol. 2019, 39, e130–e145. [Google Scholar] [CrossRef] [PubMed]
- Mai, C.W.; Chung, F.F.; Leong, C.O. Targeting Legumain As a Novel Therapeutic Strategy in Cancers. Curr. Drug Targets 2017, 18, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.D.; Beeraka, N.M.; Chitturi, C.M.K.; Madhunapantula, S.V. An Overview of Targeting Legumain for Inhibiting Cancers. Curr. Pharm. Des. 2021, 27, 3337–3348. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yi, M.; Wu, Y.; Dong, B.; Wu, K. Roles of tumor-associated macrophages in tumor progression: Implications on therapeutic strategies. Exp. Hematol. Oncol. 2021, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, X.; Zhang, X.; Xu, W. The promising role of tumor-associated macrophages in the treatment of cancer. Drug Resist. Updates 2024, 73, 101041. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.U.; Khan, I.M.; Khan, M.U.; Ud Din, M.A.; Khan, M.Z.; Khan, N.M.; Liu, Y. Role of LGMN in tumor development and its progression and connection with the tumor microenvironment. Front. Mol. Biosci. 2023, 10, 1121964. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Kang, L.; Wang, D.; Xun, J.; Chen, C.; Du, L.; Zhang, M.; Gong, J.; Mi, X.; Yue, S.; et al. Legumain-deficient macrophages promote senescence of tumor cells by sustaining JAK1/STAT1 activation. Cancer Lett. 2020, 472, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, B.; Lin, Y.; Zhou, Y.; Li, X. Legumain Promotes Gastric Cancer Progression Through Tumor-associated Macrophages In vitro and In vivo. Int. J. Biol. Sci. 2020, 16, 172–180. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.; Wang, H.; Cui, Y.; Wang, Z.; Cheng, X.; Li, W.; Hou, J.; Ji, Y.; Liu, T. High Level of Legumain Was Correlated with Worse Prognosis and Peritoneal Metastasis in Gastric Cancer Patients. Front. Oncol. 2020, 10, 966. [Google Scholar] [CrossRef]
- Jia, D.; Chen, S.; Bai, P.; Luo, C.; Liu, J.; Sun, A.; Ge, J. Cardiac Resident Macrophage-Derived Legumain Improves Cardiac Repair by Promoting Clearance and Degradation of Apoptotic Cardiomyocytes After Myocardial Infarction. Circulation 2022, 145, 1542–1556. [Google Scholar] [CrossRef]
- Tu, W.; Qin, M.; Li, Y.; Wu, W.; Tong, X. Metformin regulates autophagy via LGMN to inhibit choriocarcinoma. Gene 2023, 853, 147090. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, D.; Yu, Y.; Zhao, T.; Min, N.; Wu, Y.; Kang, L.; Zhao, Y.; Du, L.; Zhang, M.; et al. Legumain promotes tubular ferroptosis by facilitating chaperone-mediated autophagy of GPX4 in AKI. Cell Death Dis. 2021, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, W.; Xie, T.; Chen, Z.; Zhou, L.; Song, X.; Kan, H.; Lv, Y.; Wu, L.; Li, F.; et al. Identification of Novel Plasma Biomarkers for Abdominal Aortic Aneurysm by Protein Array Analysis. Biomolecules 2022, 12, 1853. [Google Scholar] [CrossRef] [PubMed]
- Marzagalli, M.; Ebelt, N.D.; Manuel, E.R. Unraveling the crosstalk between melanoma and immune cells in the tumor microenvironment. Semin. Cancer Biol. 2019, 59, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Simon, M.C. Cancer Cells Don’t Live Alone: Metabolic Communication within Tumor Microenvironments. Dev. Cell 2020, 54, 183–195. [Google Scholar] [CrossRef]
- Wang, N.; Wang, S.; Wang, X.; Zheng, Y.; Yang, B.; Zhang, J.; Pan, B.; Gao, J.; Wang, Z. Research trends in pharmacological modulation of tumor-associated macrophages. Clin. Transl. Med. 2021, 11, e288. [Google Scholar] [CrossRef] [PubMed]
- Goswami, K.K.; Bose, A.; Baral, R. Macrophages in tumor: An inflammatory perspective. Clin. Immunol. 2021, 232, 108875. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, B.; Chang-Strachan, D.; Chan, V.; Rosenbusch, A.; Ho, C.M.; Pryer, N.; Daniel, D.; Hwang, E.S.; Rugo, H.S.; Coussens, L.M. Macrophage IL-10 blocks CD8+ T cell-dependent responses to chemotherapy by suppressing IL-12 expression in intratumoral dendritic cells. Cancer Cell 2014, 26, 623–637. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; He, H.; Liu, H.; Li, R.; Chen, Y.; Qi, Y.; Jiang, Q.; Chen, L.; Zhang, P.; Zhang, H.; et al. Tumour-associated macrophages-derived CXCL8 determines immune evasion through autonomous PD-L1 expression in gastric cancer. Gut 2019, 68, 1764–1773. [Google Scholar] [CrossRef]
- Liu, F.; Wu, Q.; Dong, Z.; Liu, K. Integrins in cancer: Emerging mechanisms and therapeutic opportunities. Pharmacol. Ther. 2023, 247, 108458. [Google Scholar] [CrossRef]
- Pan, L.; Bai, P.; Weng, X.; Liu, J.; Chen, Y.; Chen, S.; Ma, X.; Hu, K.; Sun, A.; Ge, J. Legumain Is an Endogenous Modulator of Integrin αvβ3 Triggering Vascular Degeneration, Dissection, and Rupture. Circulation 2022, 145, 659–674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.X.; Gao, J.; Long, X.; Zhang, P.F.; Yang, X.; Zhu, S.Q.; Pei, X.; Qiu, B.Q.; Chen, S.W.; Lu, F.; et al. The circular RNA circHMGB2 drives immunosuppression and anti-PD-1 resistance in lung adenocarcinomas and squamous cell carcinomas via the miR-181a-5p/CARM1 axis. Mol. Cancer 2022, 21, 110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.F.; Gao, C.; Huang, X.Y.; Lu, J.C.; Guo, X.J.; Shi, G.M.; Cai, J.B.; Ke, A.W. Cancer cell-derived exosomal circUHRF1 induces natural killer cell exhaustion and may cause resistance to anti-PD1 therapy in hepatocellular carcinoma. Mol. Cancer 2020, 19, 110. [Google Scholar] [CrossRef]
- Jiang, Y.; Han, Q.; Zhao, H.; Zhang, J. Promotion of epithelial-mesenchymal transformation by hepatocellular carcinoma-educated macrophages through Wnt2b/β-catenin/c-Myc signaling and reprogramming glycolysis. J. Exp. Clin. Cancer Res. 2021, 40, 13. [Google Scholar] [CrossRef]
- Hao, H.; Cao, L.; Jiang, C.; Che, Y.; Zhang, S.; Takahashi, S.; Wang, G.; Gonzalez, F.J. Farnesoid X Receptor Regulation of the NLRP3 Inflammasome Underlies Cholestasis-Associated Sepsis. Cell Metab. 2017, 25, 856–867. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, X.; Zhang, S.-L.; Qiu, B.-Q.; Zhang, P.-F.; Liu, T.-S.; Wang, Y. Cancer Cell Secreted Legumain Promotes Gastric Cancer Resistance to Anti-PD-1 Immunotherapy by Enhancing Macrophage M2 Polarization. Pharmaceuticals 2024, 17, 951. https://doi.org/10.3390/ph17070951
Pei X, Zhang S-L, Qiu B-Q, Zhang P-F, Liu T-S, Wang Y. Cancer Cell Secreted Legumain Promotes Gastric Cancer Resistance to Anti-PD-1 Immunotherapy by Enhancing Macrophage M2 Polarization. Pharmaceuticals. 2024; 17(7):951. https://doi.org/10.3390/ph17070951
Chicago/Turabian StylePei, Xu, Shi-Long Zhang, Bai-Quan Qiu, Peng-Fei Zhang, Tian-Shu Liu, and Yan Wang. 2024. "Cancer Cell Secreted Legumain Promotes Gastric Cancer Resistance to Anti-PD-1 Immunotherapy by Enhancing Macrophage M2 Polarization" Pharmaceuticals 17, no. 7: 951. https://doi.org/10.3390/ph17070951
APA StylePei, X., Zhang, S.-L., Qiu, B.-Q., Zhang, P.-F., Liu, T.-S., & Wang, Y. (2024). Cancer Cell Secreted Legumain Promotes Gastric Cancer Resistance to Anti-PD-1 Immunotherapy by Enhancing Macrophage M2 Polarization. Pharmaceuticals, 17(7), 951. https://doi.org/10.3390/ph17070951