
The Effects of Seleno-Methionine in Cadmium-Challenged Human Primary Chondrocytes


 ,
,
Abstract
1. Introduction
2. Results
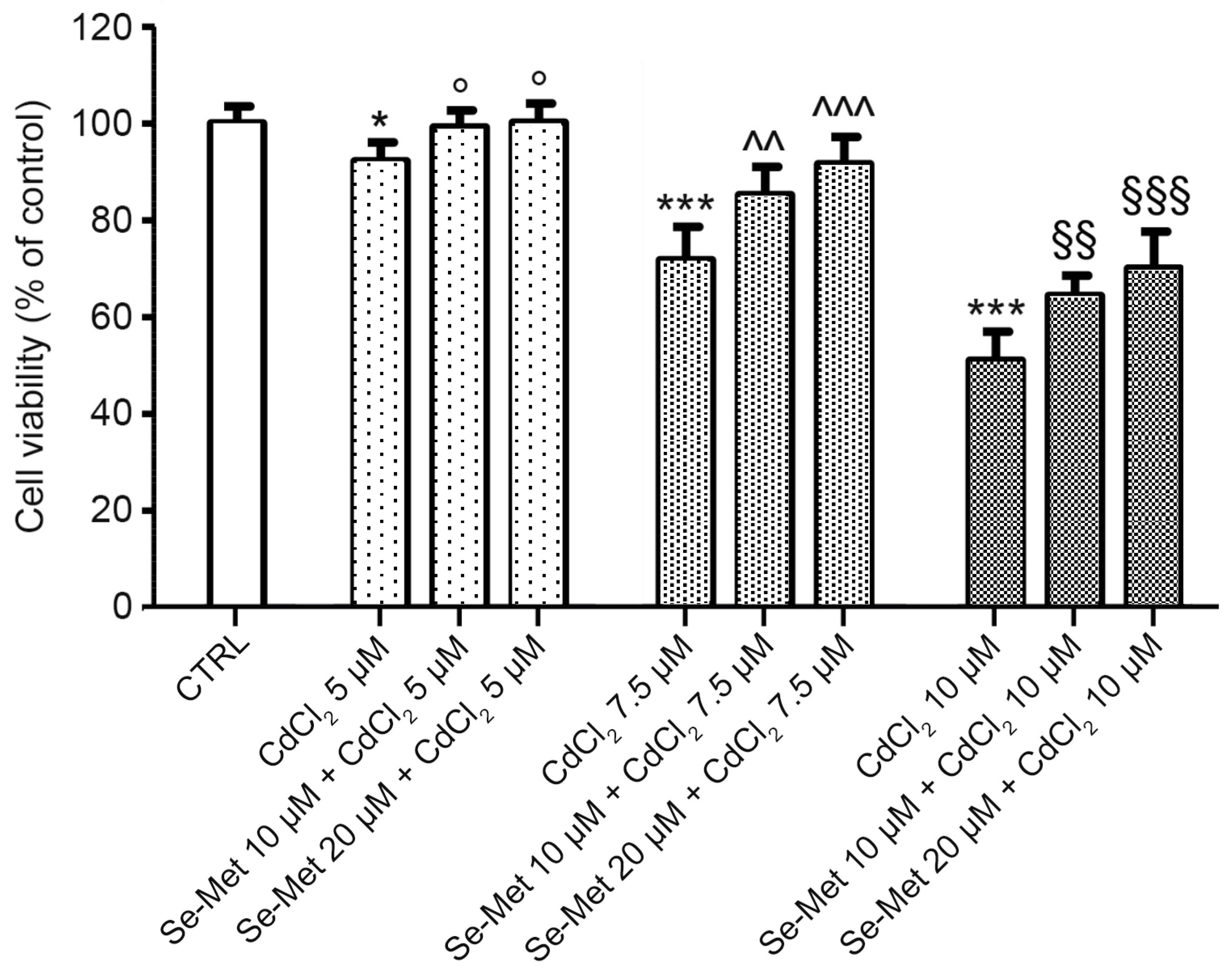
2.1. Effects of Se-Met on CdCl2-Induced Cell Injury
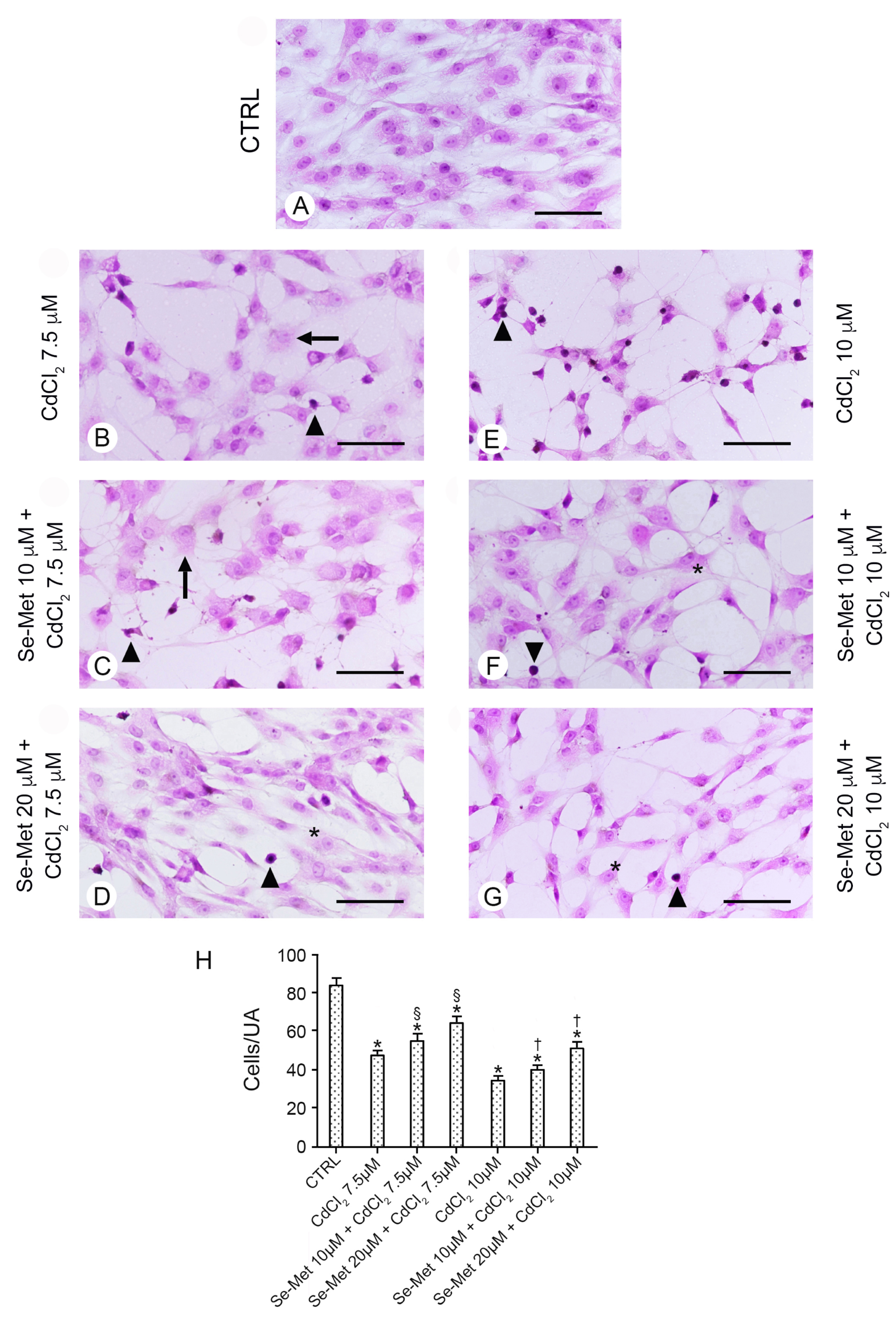
2.2. Se-Met Preserves Chondrocytes from the Damage Induced by CdCl2
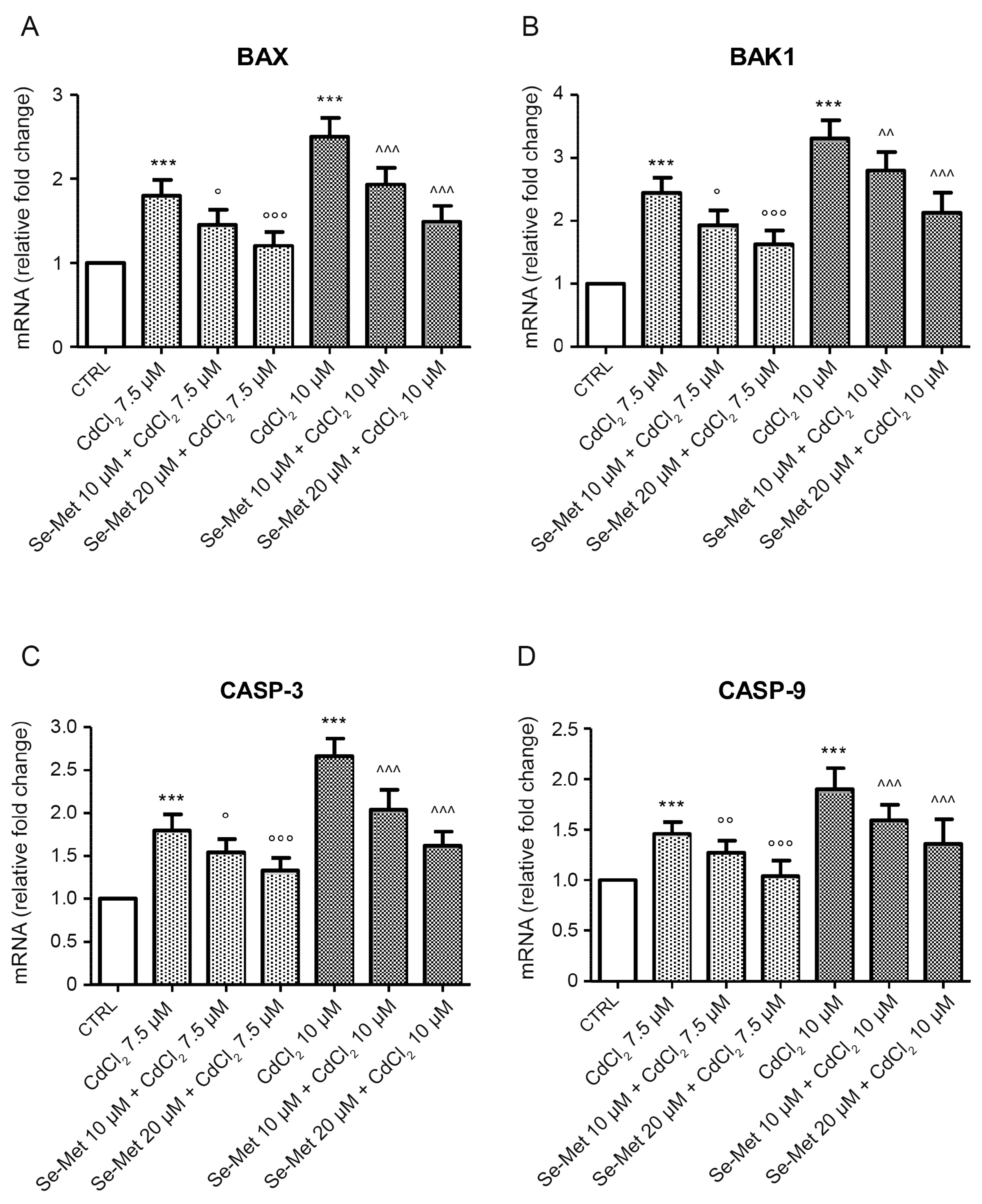
2.3. Se-Met Effects on Transcription of Apoptotic Genes
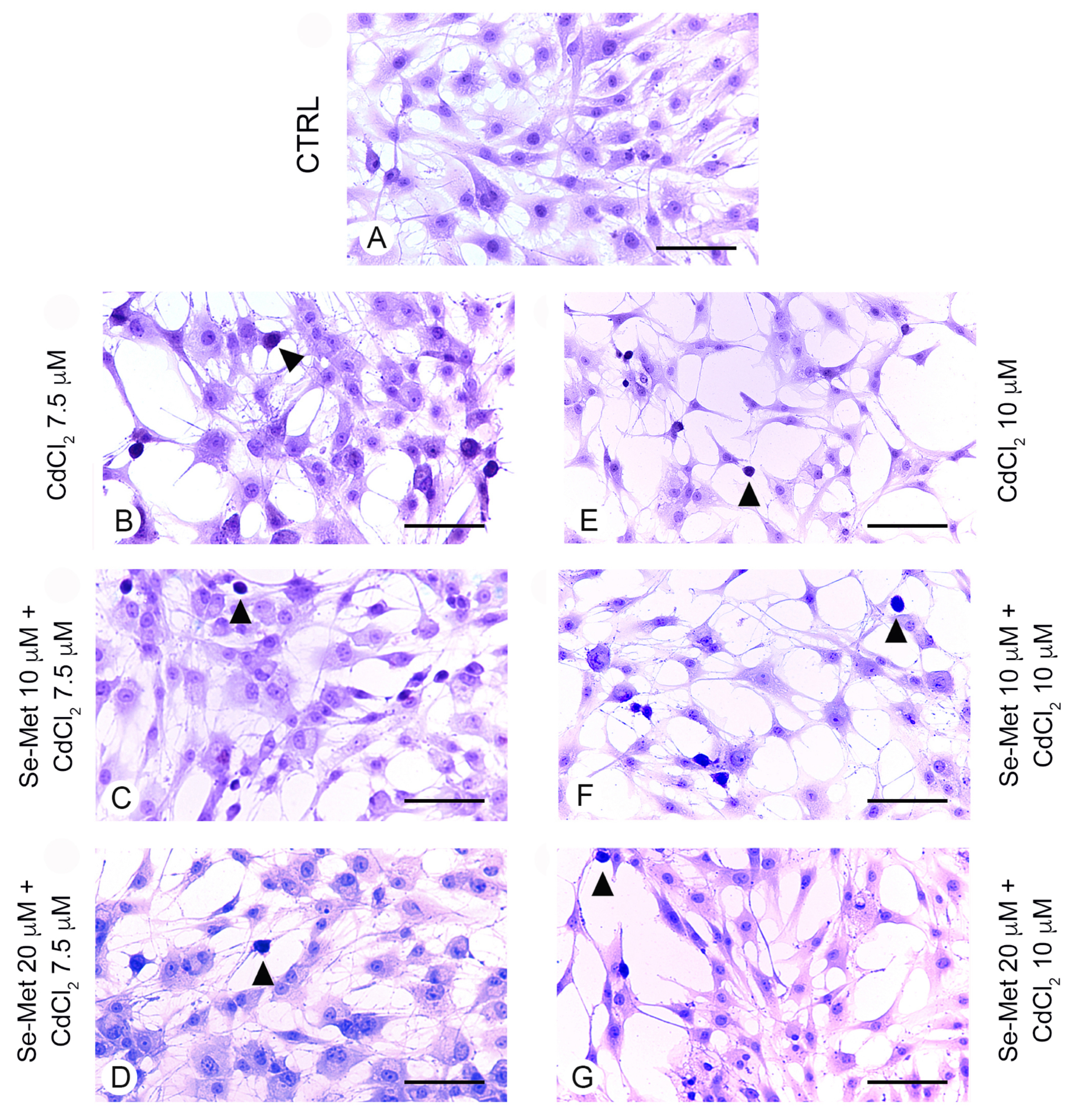
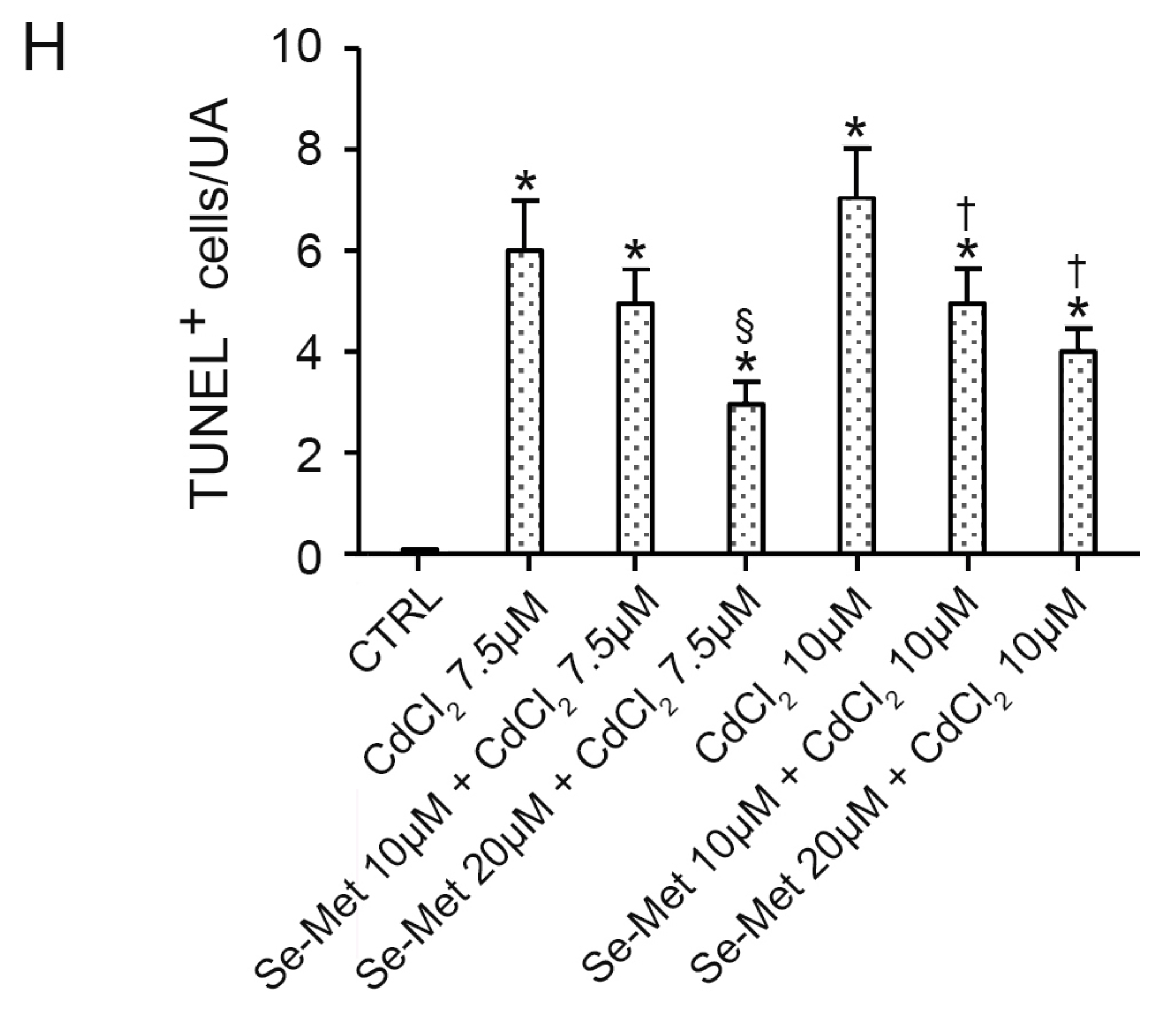
2.4. Se-Met Effects on Apoptosis with TUNEL Assay
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Treatment
4.3. MTT Assay
4.4. Histological Evaluation
4.5. Evidence of Apoptosis with TUNEL Assay
4.6. Morphometric Evaluation
4.7. RNA Extraction and Quantitative Reverse Transcription-PCR (qRT-PCR)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pourret, O.; Hursthouse, A. It’s Time to Replace the Term “Heavy Metals” with “Potentially Toxic Elements” When Reporting Environmental Research. Int. J. Environ. Res. Public Health 2019, 16, 4446. [Google Scholar] [CrossRef] [PubMed]
- Rigby, H.; Smith, S.R. The significance of cadmium entering the human food chain via livestock ingestion from the agricultural use of biosolids, with special reference to the UK. Environ. Int. 2020, 143, 105844. [Google Scholar] [CrossRef] [PubMed]
- Peana, M.; Pelucelli, A.; Medici, S.; Cappai, R.; Nurchi, V.M.; Zoroddu, M.A. Metal Toxicity and Speciation: A Review. Curr. Med. Chem. 2021, 28, 7190–7208. [Google Scholar] [CrossRef]
- Marini, H.R.; Bellone, F.; Catalano, A.; Squadrito, G.; Micali, A.; Puzzolo, D.; Freni, J.; Pallio, G.; Minutoli, L. Nutraceuticals as Alternative Approach against Cadmium-Induced Kidney Damage: A Narrative Review. Metabolites 2023, 13, 722. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, M.; Jiang, L.; Song, L. New insight into molecular interaction of heavy metal pollutant—Cadmium (II) with human serum albumin. Environ. Sci. Pollut. Res. Int. 2016, 23, 4959. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sharma, A. Cadmium toxicity: Effects on human reproduction and fertility. Rev. Environ. Health 2019, 34, 327–338. [Google Scholar] [CrossRef]
- Satarug, S.; Gobe, G.C.; Ujjin, P.; Vesey, D.A. A Comparison of the Nephrotoxicity of Low Doses of Cadmium and Lead. Toxics 2020, 8, 18. [Google Scholar] [CrossRef]
- Lee, W.K.; Thévenod, F. Cell organelles as targets of mammalian cadmium toxicity. Arch. Toxicol. 2020, 94, 1017–1049. [Google Scholar] [CrossRef]
- Charkiewicz, A.E.; Omeljaniuk, W.J.; Nowak, K.; Garley, M.; Nikliński, J. Cadmium Toxicity and Health Effects-A Brief Summary. Molecules 2023, 28, 6620. [Google Scholar] [CrossRef]
- Cui, Z.G.; Ahmed, K.; Zaidi, S.F.; Muhammad, J.S. Ins and outs of cadmium-induced carcinogenesis: Mechanism and prevention. Cancer Treat. Res. Commun. 2021, 27, 100372. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.H.; Lee, S.M.; Kim, N.H.; Moon, Y.G.; Tak, T.K.; Hyun, M.; Heo, J.D. Cadmium induces cytotoxicity in normal mouse renal MM55.K cells. Int. J. Environ. Health Res. 2022, 32, 131–140. [Google Scholar] [CrossRef]
- Zhuang, J.; Nie, G.; Yang, F.; Dai, X.; Cao, H.; Xing, C.; Hu, G.; Zhang, C. Cadmium induces cytotoxicity through oxidative stress-mediated apoptosis pathway in duck renal tubular epithelial cells. Toxicol. In Vitro 2019, 61, 104625. [Google Scholar] [CrossRef]
- Li, Z.; Chi, H.; Zhu, W.; Yang, G.; Song, J.; Mo, L.; Zhang, Y.; Deng, Y.; Xu, F.; Yang, J.; et al. Cadmium induces renal inflammation by activating the NLRP3 inflammasome through ROS/MAPK/NF-κB pathway in vitro and in vivo. Arch. Toxicol. 2021, 95, 3497–3513. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.J.; Cao, Q.J.; Xu, M.Y.; Yang, L.; Wu, Y.J. Individual and combined hepatocytotoxicity of DDT and cadmium in vitro. Toxicol. Ind. Health 2021, 37, 270–279. [Google Scholar] [CrossRef]
- Zou, H.; Wang, T.; Yuan, J.; Sun, J.; Yuan, Y.; Gu, J.; Liu, X.; Bian, J.; Liu, Z. Cadmium-induced cytotoxicity in mouse liver cells is associated with the disruption of autophagic flux via inhibiting the fusion of autophagosomes and lysosomes. Toxicol. Lett. 2020, 321, 32–43. [Google Scholar] [CrossRef]
- Del Pino, J.; Zeballos, G.; Anadón, M.J.; Moyano, P.; Díaz, M.J.; García, J.M.; Frejo, M.T. Cadmium-induced cell death of basal forebrain cholinergic neurons mediated by muscarinic M1 receptor blockade, increase in GSK-3β enzyme, β-amyloid and tau protein levels. Arch. Toxicol. 2016, 90, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhang, Y.; Zhao, S.; Chen, J.; Yang, J.; Wang, T.; Zou, H.; Wang, Y.; Gu, J.; Liu, X.; et al. Cadmium-induced apoptosis in neuronal cells is mediated by Fas/FasL-mediated mitochondrial apoptotic signaling pathway. Sci. Rep. 2018, 8, 8837. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Xu, M.; Zhang, W.; Song, R.; Zou, H.; Gu, J.; Liu, X.; Bian, J.; Liu, Z.; Yuan, Y. Cadmium induces mitochondrial dysfunction via SIRT1 suppression-mediated oxidative stress in neuronal cells. Environ. Toxicol. 2023, 38, 743–753. [Google Scholar] [CrossRef]
- Varmazyari, A.; Taghizadehghalehjoughi, A.; Sevim, C.; Baris, O.; Eser, G.; Yildirim, S.; Hacimuftuoglu, A.; Buha, A.; Wallace, D.R.; Tsatsakis, A.; et al. Cadmium sulfide-induced toxicity in the cortex and cerebellum: In vitro and in vivo studies. Toxicol. Rep. 2020, 7, 637–648. [Google Scholar] [CrossRef]
- Rinaldi, M.; Micali, A.; Marini, H.R.; Adamo, E.B.; Puzzolo, D.; Pisani, A.; Trichilo, V.; Altavilla, D.; Squadrito, F.; Minutoli, L. Cadmium, Organ Toxicity and Therapeutic Approaches: A Review on Brain, Kidney and Testis Damage. Curr. Med. Chem. 2017, 24, 3879–3893. [Google Scholar] [CrossRef]
- Wang, M.; Liu, J.; Zhu, G.; Chen, X. Low levels of cadmium exposure affect bone by inhibiting Lgr4 expression in osteoblasts and osteoclasts. J. Trace Elem. Med. Biol. 2022, 73, 127025. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Liu, W.; Chen, Y.; Guo, Y.; Gao, M.; Chen, W.; Liu, Y.; Liu, S. Diagnostic significance of metallothionein members in recognizing cadmium exposure in various organs under low-dose exposure. Chemosphere 2019, 229, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Noorafshan, A.; Niazi, B.; Mohamadpour, M.; Hoseini, L.; Hoseini, N.; Owji, A.A.; Rafati, A.; Sadeghi, Y.; Karbalay-Doust, S. First and second order stereology of hyaline cartilage: Application on mice femoral cartilage. Ann. Anat. 2016, 208, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Afridi, H.I.; Kazi, T.G.; Kazi, N.; Talpur, F.N.; Shah, F.; Naeemullah-Arain, S.S.; Brahman, K.D. Evaluation of status of arsenic, cadmium, lead and zinc levels in biological samples of normal and arthritis patients of age groups (46–60) and (61–75) years. Clin. Lab. 2013, 59, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, A.; Lemire, D.; Driessnack, M.; Naderi, M.; Niyogi, S. Interactive effects of chronic dietary selenomethionine and cadmium exposure in rainbow trout (Oncorhynchus mykiss): A preliminary study. Chemosphere 2018, 197, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Benvenga, S.; Marini, H.R.; Micali, A.; Freni, J.; Pallio, G.; Irrera, N.; Squadrito, F.; Altavilla, D.; Antonelli, A.; Ferrari, S.M.; et al. Protective Effects of Myo-Inositol and Selenium on Cadmium-Induced Thyroid Toxicity in Mice. Nutrients 2020, 12, 1222. [Google Scholar] [CrossRef] [PubMed]
- Benvenga, S.; Micali, A.; Ieni, A.; Antonelli, A.; Fallahi, P.; Pallio, G.; Irrera, N.; Squadrito, F.; Picciolo, G.; Puzzolo, D.; et al. The Association of Myo-Inositol and Selenium Contrasts Cadmium-Induced Thyroid C Cell Hyperplasia and Hypertrophy in Mice. Front. Endocrinol. 2021, 12, 608697. [Google Scholar] [CrossRef]
- Satarug, S.; Gobe, G.C.; Vesey, D.A. Multiple Targets of Toxicity in Environmental Exposure to Low-Dose Cadmium. Toxics 2022, 10, 472. [Google Scholar] [CrossRef]
- Jamwal, A.; Naderi, M.; Niyogi, S. An in vitro examination of selenium-cadmium antagonism using primary cultures of rainbow trout (Oncorhynchus mykiss) hepatocytes. Metallomic 2016, 8, 218–227. [Google Scholar] [CrossRef]
- Yan, J.; Fei, Y.; Han, Y.; Lu, S. Selenoprotein O deficiencies suppress chondrogenic differentiation of ATDC5 cells. Cell. Biol. Int. 2016, 40, 1033–1040. [Google Scholar] [CrossRef]
- Branca, J.J.V.; Morucci, G.; Maresca, M.; Tenci, B.; Cascella, R.; Paternostro, F.; Ghelardini, C.; Gulisano, M.; Di Cesare Mannelli, L.; Pacini, A. Selenium and zinc: Two key players against cadmium-induced neuronal toxicity. Toxicol. In Vitro 2018, 48, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements. Am. J. Clin. Nutr. 2010, 92, 1002. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cheng, T.; Yu, X. The Impact of Trace Elements on Osteoarthritis. Front. Med. 2021, 8, 771297. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Torres, J.; Zamudio-Cuevas, Y.; Martínez-Nava, G.A.; Aztatzi-Aguilar, O.G.; Sierra-Vargas, M.P.; Lozada-Pérez, C.A.; Suárez-Ahedo, C.; Landa-Solís, C.; Olivos-Meza, A.; Del Razo, L.M.; et al. Impact of Cadmium Mediated by Tobacco Use in Musculoskeletal Diseases. Biol. Trace Elem. Res. 2022, 200, 2008–2015. [Google Scholar] [CrossRef] [PubMed]
- Babazadeh, D.; Ahmadi Simab, P. Methionine in Poultry Nutrition: A Review. J. World’s Poult. Sci. 2022, 1, 1–11. [Google Scholar] [CrossRef]
- Chwalba, A.; Orłowska, J.; Słota, M.; Jeziorska, M.; Filipecka, K.; Bellanti, F.; Dobrakowski, M.; Kasperczyk, A.; Zalejska-Fiolka, J.; Kasperczyk, S. Effect of Cadmium on Oxidative Stress Indices and Vitamin D Concentrations in Children. J. Clin. Med. 2023, 12, 1572. [Google Scholar] [CrossRef] [PubMed]
- Branca, J.J.V.; Carrino, D.; Paternostro, F.; Morucci, G.; Fiorillo, C.; Nicoletti, C.; Gulisano, M.; Ghelardini, C.; Di Cesare Mannelli, L.; Becatti, M.; et al. The Protection of Zinc against Acute Cadmium Exposure: A Morphological and Molecular Study on a BBB In Vitro Model. Cells. 2022, 11, 1646. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Torres, J.; Plata-Rodríguez, R.; Zamudio-Cuevas, Y.; Martínez-Nava, G.A.; Landa-Solís, C.; Mendoza-Soto, L.; Olivos-Meza, A.; Suárez-Ahedo, C.; Barbier, O.C.; Narváez-Morales, J.; et al. Effect of cadmium on the viability on monolayer cultures of synoviocytes, chondrocytes, and Hoffa: A preliminary study. Toxicol. Ind. Health 2020, 36, 940–945. [Google Scholar] [CrossRef]
- Zamudio-Cuevas, Y.E.; Martínez-Nava, G.; Reyes-Hinojosa, D.; Mendoza-Soto, L.; Fernández-Torres, J.; López-Reyes, A.; Olivos-Meza, A.; Armienta-Hernández, M.A.; Ruíz-Huerta, E.A.; de Jesús González-Guadarrama, M.; et al. Impact of cadmium toxicity on cartilage loss in a 3D in vitro model. Environ. Toxicol. Pharmacol. 2020, 74, 103307. [Google Scholar]
- Gu, J.; Li, S.; Wang, G.; Zhang, X.; Yuan, Y.; Liu, X.; Bian, J.; Tong, X.; Liu, Z. Cadmium Toxicity on Chondrocytes and the Palliative Effects of 1α, 25-Dihydroxy Vitamin D3 in White Leghorns Chicken’s Embryo. Front. Vet. Sci. 2021, 8, 637369. [Google Scholar] [CrossRef]
- Martínez-Nava, G.A.; Mendoza-Soto, L.; Fernández-Torres, J.; Zamudio-Cuevas, Y.; Reyes-Hinojosa, D.; Plata-Rodríguez, R.; Olivos-Meza, A.; Ruíz-Huerta, E.A.; Armienta-Hernández, M.A.; Hernández-Álvarez, E.; et al. Effect of cadmium on the concentration of essential metals in a human chondrocyte micromass culture. J. Trace Elem. Med. Biol. 2020, 62, 126614. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ran, D.; Shi, X.; Zhao, H.; Liu, Z. Cadmium toxicity: A role in bone cell function and teeth development. Sci. Total Environ. 2021, 769, 144646. [Google Scholar] [CrossRef]
- Wang, Q.; Ying, L.; Wei, B.; Ji, Y.; Xu, Y. Effects of quercetin on apoptosis and extracellular matrix degradation of chondrocytes induced by oxidative stress-mediated pyroptosis. J. Biochem. Mol. Toxicol. 2022, 36, e22951. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Kondo, T.; Zhao, Q.L.; Li, F.J.; Tanabe, K.; Arai, Y.; Zhou, Z.C.; Kasuya, M. Apoptosis induced by cadmium in human lymphoma U937 cells through Ca2+-calpain and caspase-mitochondria- dependent pathways. J. Biol. Chem. 2000, 275, 39702–39709. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Cheon, H.S.; Kim, S.Y.; Juhnn, Y.S.; Kim, Y.Y. Cadmium induces neuronal cell death through reactive oxygen species activated by GADD153. BMC Cell. Biol. 2013, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Cheon, H.; Kim, S.M.; Kim, Y.Y. GSK-3β-mediated regulation of cadmium-induced cell death and survival. Cell. Mol. Biol. Lett. 2018, 23, 9. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Micali, A.; Rinaldi, M.; Irrera, N.; Marini, H.; Puzzolo, D.; Pisani, A.; Lorenzini, C.; Valenti, A.; Laurà, R.; et al. Polydeoxyribonucleotide, an Adenosine-A2A Receptor Agonist, Preserves Blood Testis Barrier from Cadmium-Induced Injury. Front. Pharmacol. 2017, 7, 537, Erratum in Front. Pharmacol. 2022, 13, 1073543. [Google Scholar] [CrossRef] [PubMed]
- Pallio, G.; Micali, A.; Benvenga, S.; Antonelli, A.; Marini, H.R.; Puzzolo, D.; Macaione, V.; Trichilo, V.; Santoro, G.; Irrera, N.; et al. Myo-inositol in the protection from cadmium-induced toxicity in mice kidney: An emerging nutraceutical challenge. Food Chem. Toxicol. 2019, 132, 110675. [Google Scholar] [CrossRef] [PubMed]
- Pérez Díaz, M.F.F.; Plateo Pignatari, M.G.; Filippa, V.P.; Mohamed, F.H.; Marchevsky, E.J.; Gimenez, M.S.; Ramirez, D.C. A soybean-based diet modulates cadmium-induced vascular apoptosis. J. Trace Elem. Med. Biol. 2019, 52, 239–246. [Google Scholar] [CrossRef]
- Ferlazzo, N.; Micali, A.; Marini, H.R.; Freni, J.; Santoro, G.; Puzzolo, D.; Squadrito, F.; Pallio, G.; Navarra, M.; Cirmi, S.; et al. A Flavonoid-Rich Extract from Bergamot Juice, Alone or in Association with Curcumin and Resveratrol, Shows Protective Effects in a Murine Model of Cadmium-Induced Testicular Injury. Pharmaceuticals 2021, 14, 386. [Google Scholar] [CrossRef]
- Cirmi, S.; Maugeri, A.; Micali, A.; Marini, H.R.; Puzzolo, D.; Santoro, G.; Freni, J.; Squadrito, F.; Irrera, N.; Pallio, G.; et al. Cadmium-Induced Kidney Injury in Mice Is Counteracted by a Flavonoid-Rich Extract of Bergamot Juice, Alone or in Association with Curcumin and Resveratrol, via the Enhancement of Different Defense Mechanisms. Biomedicines 2021, 9, 1797. [Google Scholar] [CrossRef]
- Fang, J.; Xie, S.; Chen, Z.; Wang, F.; Chen, K.; Zuo, Z.; Cui, H.; Guo, H.; Ouyang, P.; Chen, Z.; et al. Protective Effect of Vitamin E on Cadmium-Induced Renal Oxidative Damage and Apoptosis in Rats. Biol. Trace Elem. Res. 2021, 199, 4675–4687. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ding, L.; Wang, K.; Huang, R.; Yu, W.; Yan, B.; Wang, H.; Zhang, C.; Yang, Z.; Liu, Z. Role of endoplasmic reticulum stress in cadmium-induced hepatocyte apoptosis and the protective effect of quercetin. Ecotoxicol. Environ. Saf. 2022, 241, 113772. [Google Scholar] [CrossRef] [PubMed]
- Al Kahtani, M.A. Effect of both selenium and biosynthesized nanoselenium particles on cadmium-induced neurotoxicity in albino rats. Hum. Exp. Toxicol. 2020, 39, 159–172. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, S.; Gavrilović, A.; Li, D.; Tang, R. Selenium alleviates cadmium-induced oxidative stress, endoplasmic reticulum stress, and apoptosis in L8824 cells. Ecotoxicol. Environ. Saf. 2023, 262, 115337. [Google Scholar] [CrossRef]
- Zwolak, I. The Role of Selenium in Arsenic and Cadmium Toxicity: An Updated Review of Scientific Literature. Biol. Trace Elem. Res. 2020, 193, 44–63. [Google Scholar] [CrossRef] [PubMed]
- Wan, N.; Xu, Z.; Liu, T.; Min, Y.; Li, S. Ameliorative Effects of Selenium on Cadmium-Induced Injury in the Chicken Ovary: Mechanisms of Oxidative Stress and Endoplasmic Reticulum Stress in Cadmium-Induced Apoptosis. Biol. Trace Elem. Res. 2018, 184, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Wang, S.; Zhang, C.; Hu, X.; Zhou, L.; Li, Y.; Xu, L. Selenium ameliorates cadmium-induced mouse leydig TM3 cell apoptosis via inhibiting the ROS/JNK/c-jun signaling pathway. Ecotoxicol. Environ. Saf. 2020, 192, 110266. [Google Scholar] [CrossRef]
- Sun, M.; Hussain, S.; Hu, Y.; Yan, J.; Min, Z.; Lan, X.; Guo, Y.; Zhao, Y.; Huang, H.; Feng, M.; et al. Maintenance of SOX9 stability and ECM homeostasis by selenium-sensitive PRMT5 in cartilage. Osteoarthr. Cartil. 2019, 7, 932–944. [Google Scholar] [CrossRef]
- Kurokawa, S.; Berry, M.J. Selenium. Role of the essential metalloid in health. Met. Ions Life Sci. 2013, 13, 499–534. [Google Scholar]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Asadi, M.; Taghizadeh, S.; Kaviani, E.; Vakili, O.; Taheri-Anganeh, M.; Tahamtan, M.; Savardashtaki, A. Caspase-3: Structure, function, and biotechnological aspects. Biotechnol. Appl. Biochem. 2022, 69, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Vailati-Riboni, M.; Xu, T.; Qadir, B.; Bucktrout, R.; Parys, C.; Loor, J.J. In vitro methionine supplementation during lipopolysaccharide stimulation modulates immunometabolic gene network expression in isolated polymorphonuclear cells from lactating Holstein cows. J. Dairy Sci. 2019, 102, 8343–8351. [Google Scholar] [CrossRef] [PubMed]
- Minutoli, L.; Squadrito, F.; Altavilla, D.; Marini, H. Chapter 21: Therapy with Selenium Cocktails and Co-Use of Lycopene and Selenium. In Selenium: Chemistry, Analysis, Function and Effects; Preedy, V.R., Ed.; Series: Food and Nutritional Components in Focus; Royal Society of Chemistry: London, UK, 2015; Volume 9, pp. 363–376. [Google Scholar] [CrossRef]
- Marini, H.R.; Micali, A.; Squadrito, G.; Puzzolo, D.; Freni, J.; Antonuccio, P.; Minutoli, L. Nutraceuticals: A New Challenge against Cadmium-Induced Testicular Injury. Nutrients 2022, 14, 663. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Rawal, R. Influence of heavy metal exposure on gut microbiota: Recent advances. J. Biochem. Mol. Toxicol. 2023, 37, e23485. [Google Scholar] [CrossRef] [PubMed]
- Porru, S.; Esplugues, A.; Llop, S.; Delgado-Saborit, J.M. The effects of heavy metal exposure on brain and gut microbiota: A systematic review of animal studies. Environ. Pollut. 2024, 348, 123732. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, R.M.; D’Ascola, A.; Vicchio, T.M.; Campo, S.; Gianì, F.; Giovinazzo, S.; Frasca, F.; Cannavò, S.; Campennì, A.; Trimarchi, F. Selenium exerts protective effects against oxidative stress and cell damage in human thyrocytes and fibroblasts. Endocrine 2020, 68, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Marini, H.R. Mediterranean Diet and Soy Isoflavones for Integrated Management of the Menopausal Metabolic Syndrome. Nutrients 2022, 14, 1550. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, F.; Mostafalou, S.; Bahadar, H.; Abdollahi, M. Review of endocrine disorders associated with environmental toxicants and possible involved mechanisms. Life Sci. 2016, 145, 265–273. [Google Scholar] [CrossRef]
- Holz, J.D.; Sheu, T.J.; Drissi, H.; Matsuzawa, M.; Zuscik, M.J.; Puzas, J.E. Environmental agents affect skeletal growth and development. Birth Defects Res. C Embryo Today 2007, 81, 41–50. [Google Scholar] [CrossRef]
- Huang, W.; Wang, X.; Zheng, S.; Wu, R.; Liu, C.; Wu, K. Effect of bisphenol A on craniofacial cartilage development in zebrafish (Danio rerio) embryos: A morphological study. Ecotoxicol. Environ. Saf. 2021, 212, 111991. [Google Scholar] [CrossRef] [PubMed]
- Deprouw, C.; Courties, A.; Fini, J.B.; Clerget-Froidevaux, M.S.; Demeneix, B.; Berenbaum, F.; Sellam, J.; Louati, K. Pollutants: A candidate as a new risk factor for osteoarthritis-results from a systematic literature review. RMD Open 2022, 8, e001983. [Google Scholar] [CrossRef] [PubMed]
- Scuruchi, M.; D’Ascola, A.; Avenoso, A.; Zappone, A.; Mandraffino, G.; Campo, S.; Campo, G.M. miR9 inhibits 6-mer HA-induced cytokine production and apoptosis in human chondrocytes by reducing NF-kB activation. Arch. Biochem. Biophys. 2022, 718, 109139. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Fu, M.; Bi, R.; Zheng, X.; Fu, B.; Tian, S.; Liu, C.; Li, Q.; Liu, J. Cadmium induced BEAS-2B cells apoptosis and mitochondria damage via MAPK signaling pathway. Chemosphere 2021, 263, 128346. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| β-actin | Fw: 5′-AGAGCTACGAGCTGCCTGAC-3′ |
| Rw: 5′-AGCACTGTGTTGGCGTACAG-3′ | |
| BAX | Fw: 5′-GACGAACTGGACAGTAACATGG-3′ |
| Rw: 5′-TCAGAAAACATGTCAGCTGCC-3′ | |
| BAK1 | Fw: 5′-TGGGACACTGTGAACCAGGA-3′ |
| Rw: 5′-GAGGAAGCCAAACACCAGTAGG-3′ | |
| CASP-3 | Fw: 5′-TGAGGCATGGTGAAGAAGGA-3′ |
| Rw: 5′-TCCAGTTCTGTACCACGGCA-3′ | |
| CASP-9 | Fw: 5′-CTGGACGCCATATCTAGTTTGC-3′ |
| Rw: 5′-AACGTACCAGGAGCCACTCTT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urzì Brancati, V.; Aliquò, F.; Freni, J.; Pantano, A.; Galipò, E.; Puzzolo, D.; Minutoli, L.; Marini, H.R.; Campo, G.M.; D’Ascola, A. The Effects of Seleno-Methionine in Cadmium-Challenged Human Primary Chondrocytes. Pharmaceuticals 2024, 17, 936. https://doi.org/10.3390/ph17070936
Urzì Brancati V, Aliquò F, Freni J, Pantano A, Galipò E, Puzzolo D, Minutoli L, Marini HR, Campo GM, D’Ascola A. The Effects of Seleno-Methionine in Cadmium-Challenged Human Primary Chondrocytes. Pharmaceuticals. 2024; 17(7):936. https://doi.org/10.3390/ph17070936
Chicago/Turabian StyleUrzì Brancati, Valentina, Federica Aliquò, José Freni, Alice Pantano, Erika Galipò, Domenico Puzzolo, Letteria Minutoli, Herbert Ryan Marini, Giuseppe Maurizio Campo, and Angela D’Ascola. 2024. "The Effects of Seleno-Methionine in Cadmium-Challenged Human Primary Chondrocytes" Pharmaceuticals 17, no. 7: 936. https://doi.org/10.3390/ph17070936
APA StyleUrzì Brancati, V., Aliquò, F., Freni, J., Pantano, A., Galipò, E., Puzzolo, D., Minutoli, L., Marini, H. R., Campo, G. M., & D’Ascola, A. (2024). The Effects of Seleno-Methionine in Cadmium-Challenged Human Primary Chondrocytes. Pharmaceuticals, 17(7), 936. https://doi.org/10.3390/ph17070936