
Discovery of Novel Allosteric SHP2 Inhibitor Using Pharmacophore-Based Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and Principal Component Analysis



Abstract
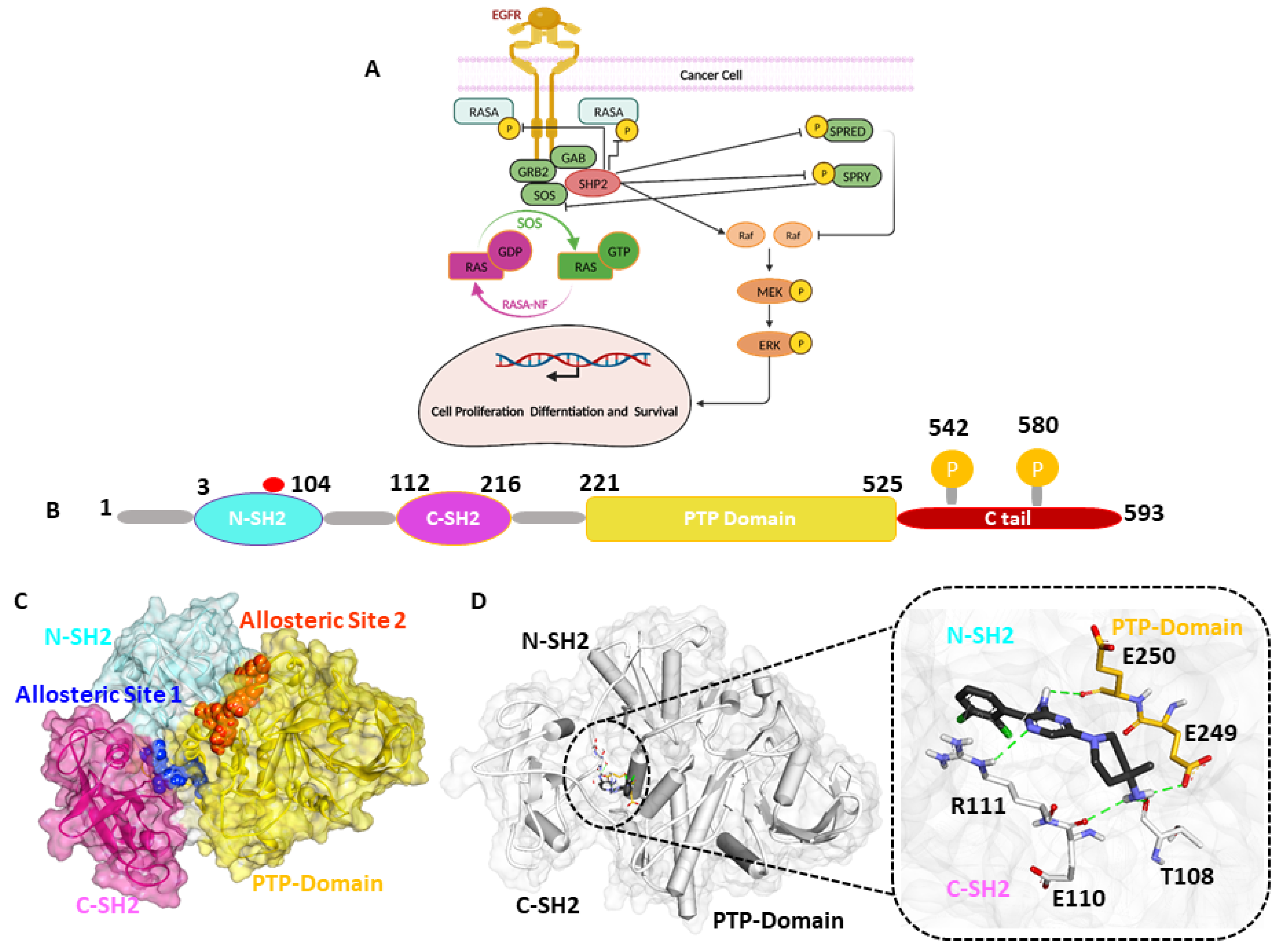
1. Introduction
2. Results
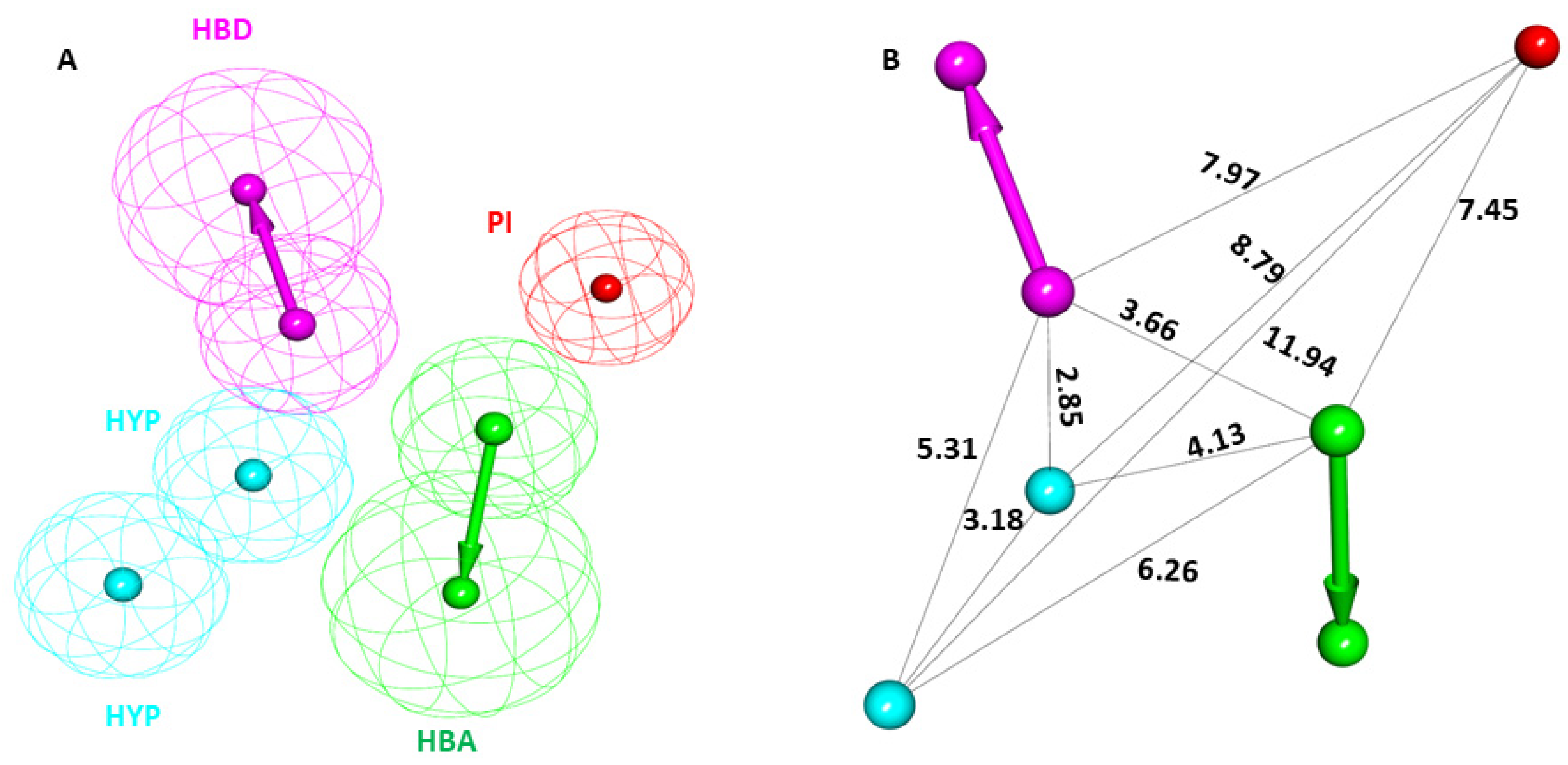
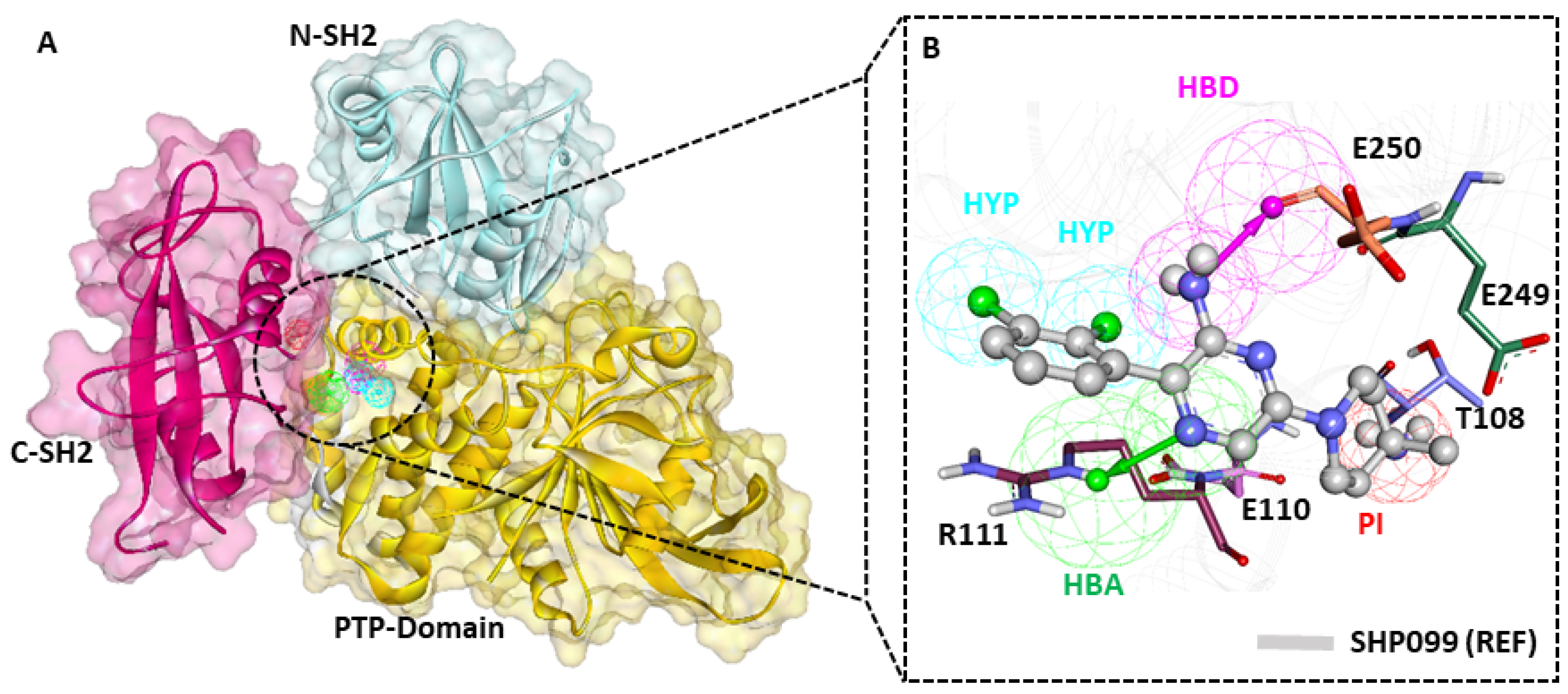
2.1. Generation of Receptor-Based Pharmacophore Model
2.2. Pharmacophore Model Validation
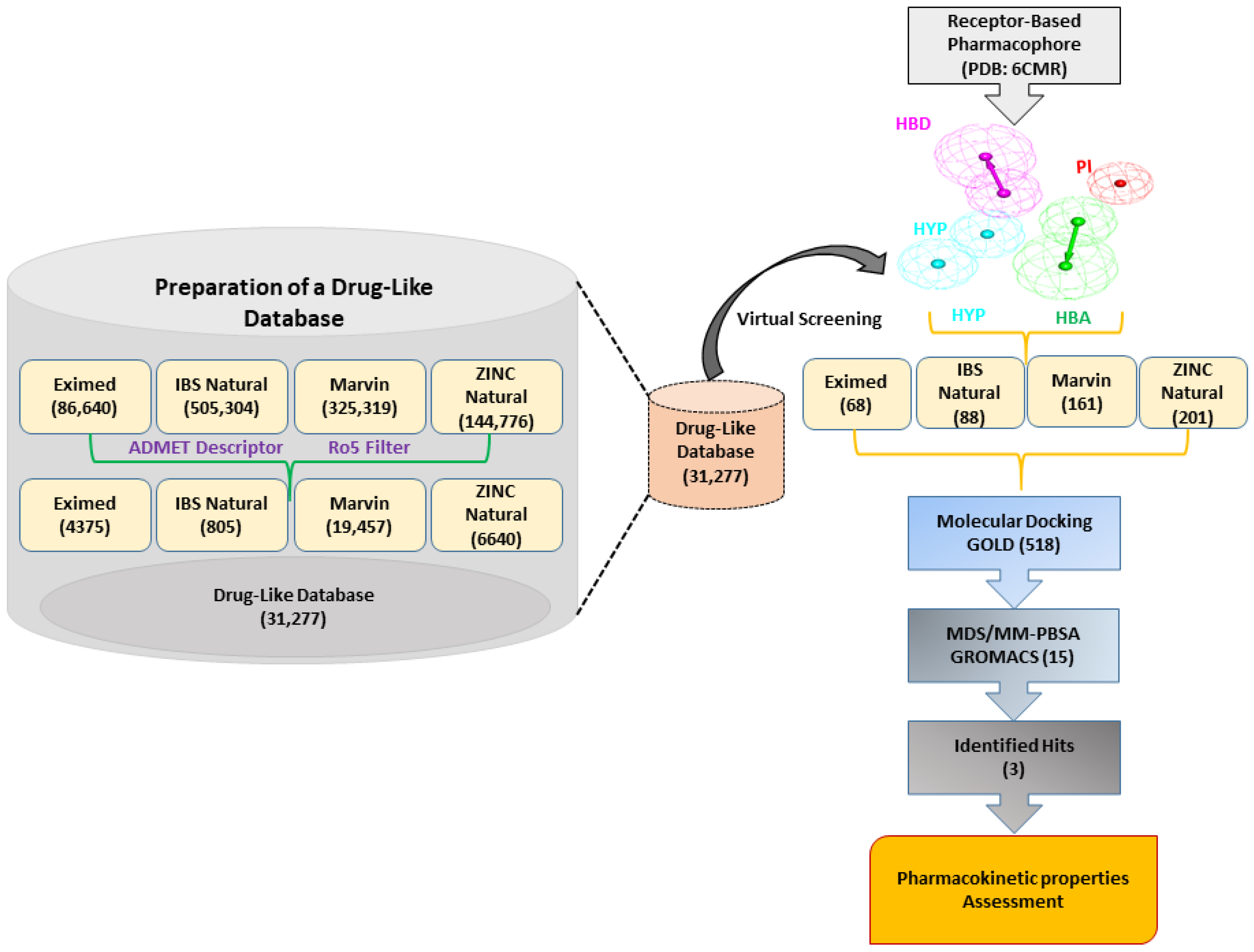
2.3. Virtual Screening
2.4. Molecular Docking
2.5. Molecular Dynamics Simulation
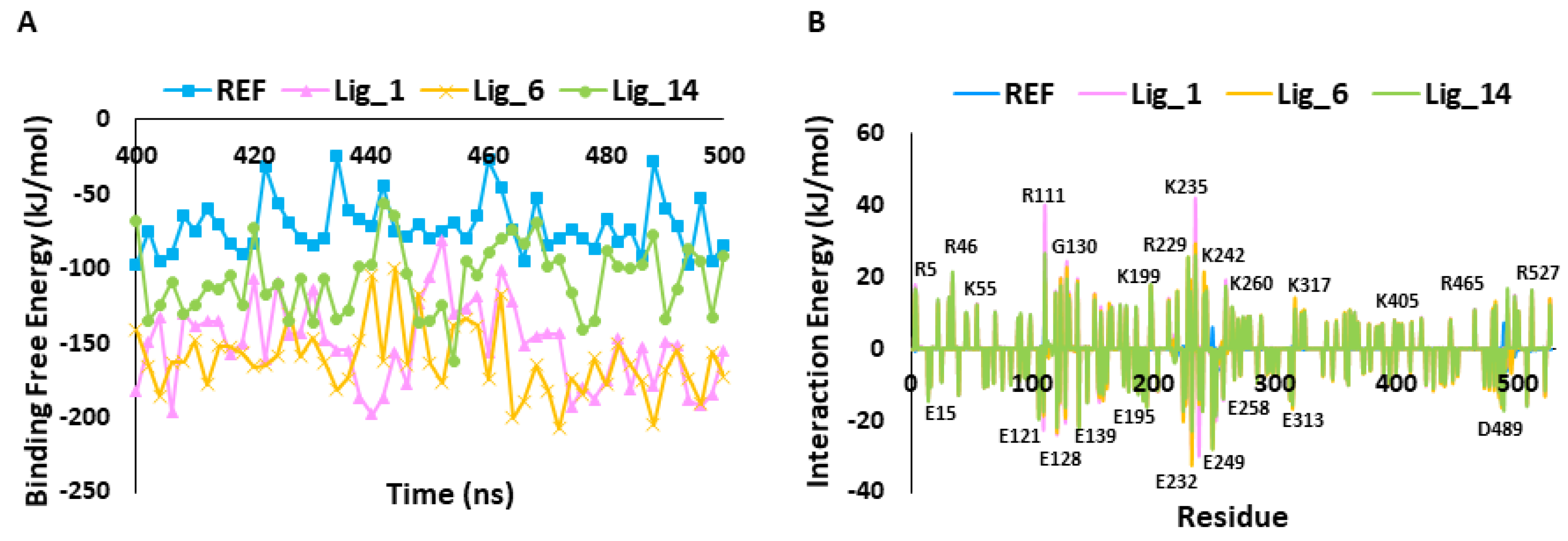
2.5.1. Stability Assessment of Simulated Complexes
2.5.2. MM-PBSA Calculation
2.5.3. Principal Component Analysis
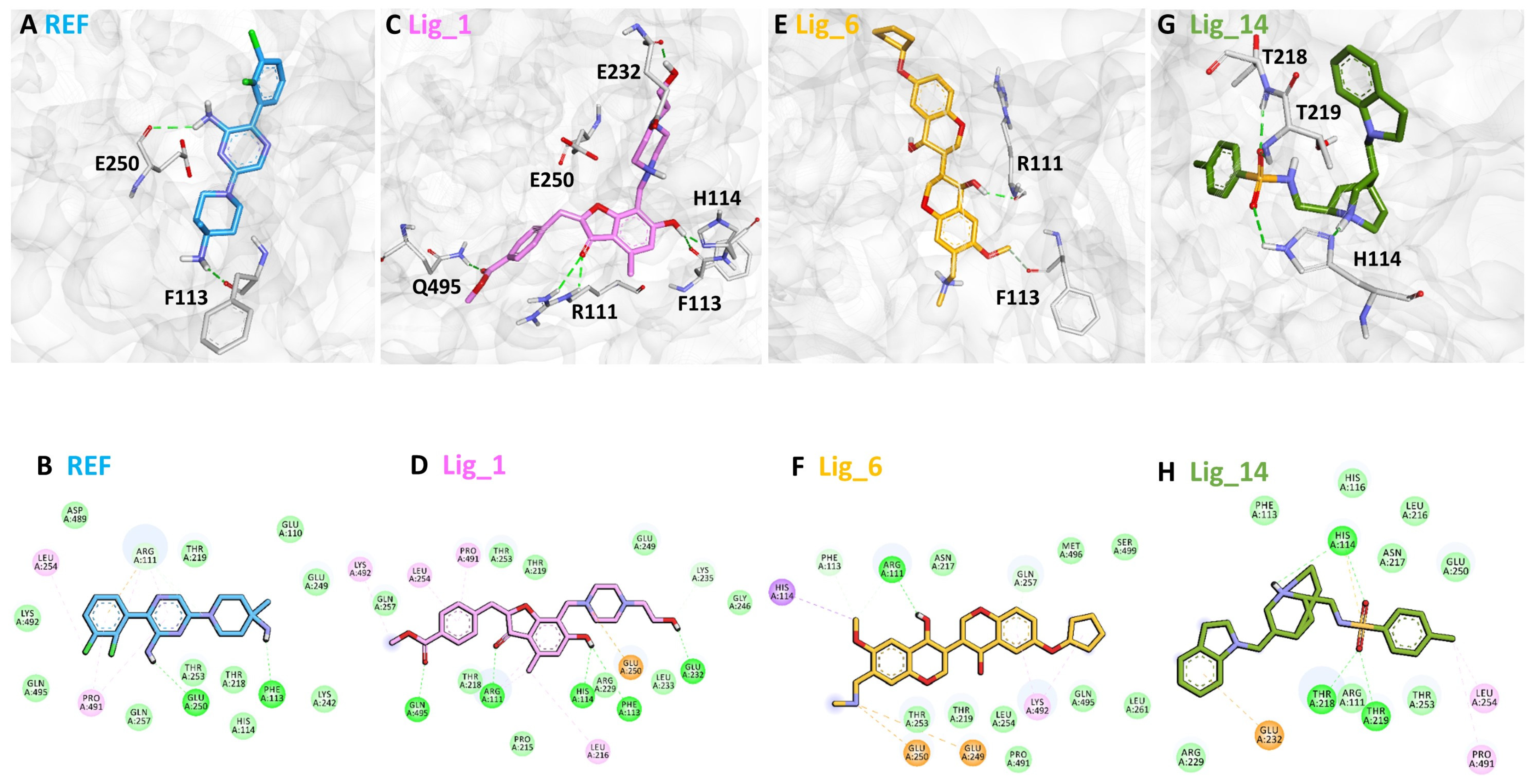
2.6. Binding Mode Analysis
2.7. Pharmacokinetic Property Assessment
3. Discussion
4. Materials and Methods
4.1. Generation of Receptor-Based Pharmacophore Model
4.2. Pharmacophore Model Validation
4.3. Virtual Screening
4.4. Molecular Docking
4.5. Molecular Dynamics Simulation
4.5.1. Stability of Simulation Systems
4.5.2. MM-PBSA Binding Free Energy Calculation
4.5.3. Principal Component and Free Energy Landscape Analysis
4.6. Pharmacokinetic Properties Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garcia Fortanet, J.; Chen, C.H.-T.; Chen, Y.-N.P.; Chen, Z.; Deng, Z.; Firestone, B.; Fekkes, P.; Fodor, M.; Fortin, P.D.; Fridrich, C.; et al. Allosteric Inhibition of SHP2: Identification of a Potent, Selective, and Orally Efficacious Phosphatase Inhibitor. J. Med. Chem. 2016, 59, 7773–7782. [Google Scholar] [CrossRef] [PubMed]
- Bobone, S.; Pannone, L.; Biondi, B.; Solman, M.; Flex, E.; Canale, V.C.; Calligari, P.; De Faveri, C.; Gandini, T.; Quercioli, A.; et al. Targeting Oncogenic Src Homology 2 Domain-Containing Phosphatase 2 (SHP2) by Inhibiting Its Protein–Protein Interactions. J. Med. Chem. 2021, 64, 15973–15990. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-N.P.; LaMarche, M.J.; Chan, H.M.; Fekkes, P.; Garcia-Fortanet, J.; Acker, M.G.; Antonakos, B.; Chen, C.H.-T.; Chen, Z.; Cooke, V.G.; et al. Allosteric Inhibition of SHP2 Phosphatase Inhibits Cancers Driven by Receptor Tyrosine Kinases. Nature 2016, 535, 148–152. [Google Scholar] [CrossRef]
- Song, Y.; Wang, S.; Zhao, M.; Yang, X.; Yu, B. Strategies Targeting Protein Tyrosine Phosphatase SHP2 for Cancer Therapy. J. Med. Chem. 2022, 65, 3066–3079. [Google Scholar] [CrossRef]
- Liu, M.; Gao, S.; Elhassan, R.M.; Hou, X.; Fang, H. Strategies to Overcome Drug Resistance Using SHP2 Inhibitors. Acta Pharm. Sin. B 2021, 11, 3908–3924. [Google Scholar] [CrossRef] [PubMed]
- Östman, A.; Hellberg, C.; Böhmer, F.D. Protein-Tyrosine Phosphatases and Cancer. Nat. Rev. Cancer 2006, 6, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Raveendra-Panickar, D.; Finlay, D.; Layng, F.I.; Lambert, L.J.; Celeridad, M.; Zhao, M.; Barbosa, K.; De Backer, L.J.S.; Kwong, E.; Gosalia, P.; et al. Discovery of Novel Furanylbenzamide Inhibitors That Target Oncogenic Tyrosine Phosphatase SHP2 in Leukemia Cells. J. Biol. Chem. 2022, 298, 101477. [Google Scholar] [CrossRef]
- Araki, T.; Mohi, M.G.; Ismat, F.A.; Bronson, R.T.; Williams, I.R.; Kutok, J.L.; Yang, W.; Pao, L.I.; Gilliland, D.G.; Epstein, J.A.; et al. Mouse Model of Noonan Syndrome Reveals Cell Type- and Gene Dosage-Dependent Effects of Ptpn11 Mutation. Nat. Med. 2004, 10, 849–857. [Google Scholar] [CrossRef]
- Tartaglia, M.; Gelb, B.D. Germ-Line and Somatic PTPN11 Mutations in Human Disease. Eur. J. Med. Genet. 2005, 48, 81–96. [Google Scholar] [CrossRef]
- Cai, T.; Nishida, K.; Hirano, T.; Khavari, P.A. Gab1 and SHP-2 Promote Ras/MAPK Regulation of Epidermal Growth and Differentiation. J. Cell Biol. 2002, 159, 103–112. [Google Scholar] [CrossRef]
- Yuan, Y.; Fan, Y.; Gao, Z.; Sun, X.; Zhang, H.; Wang, Z.; Cui, Y.; Song, W.; Wang, Z.; Zhang, F.; et al. SHP2 Promotes Proliferation of Breast Cancer Cells through Regulating Cyclin D1 Stability via the PI3K/AKT/GSK3β Signaling Pathway. Cancer Biol. Med. 2020, 17, 707–725. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Morales, L.D.; Jang, I.-S.; Cho, Y.-Y.; Kim, D.J. Protein Tyrosine Phosphatases as Potential Regulators of STAT3 Signaling. Int. J. Mol. Sci. 2018, 19, 2708. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Qu, C.-K. Protein Tyrosine Phosphatases in the JAK/STAT Pathway. Front. Biosci. 2008, 13, 4925–4932. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Bu, H.; Zhou, J.; Yang, C.-Y.; Zhang, H. Recent Advances of SHP2 Inhibitors in Cancer Therapy: Current Development and Clinical Application. J. Med. Chem. 2020, 63, 11368–11396. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yu, B.; Xu, G.; Xu, W.-R.; Loh, M.L.; Tang, L.-D.; Qu, C.-K. Identification of Cryptotanshinone as an Inhibitor of Oncogenic Protein Tyrosine Phosphatase SHP2 (PTPN11). J. Med. Chem. 2013, 56, 7212–7221. [Google Scholar] [CrossRef] [PubMed]
- Lade, D.M.; Agazie, Y.M. Targeting SHP2 with an Active Site Inhibitor Blocks Signaling and Breast Cancer Cell Phenotypes. ACS Bio Med. Chem. Au 2023, 3, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, H.; Zhao, G.; Wang, R. Allosteric Inhibitors of SHP2: An Updated Patent Review (2015–2020). Curr. Med. Chem. 2021, 28, 3825–3842. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Wang, M.; Ge, Y.; Chen, X.-P.; Xu, Z.; Sun, Y.; Xiong, X.-F. Tyrosine Phosphatase SHP2 Inhibitors in Tumor-Targeted Therapies. Acta Pharm. Sin. B 2021, 11, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Pádua, R.A.P.; Sun, Y.; Marko, I.; Pitsawong, W.; Stiller, J.B.; Otten, R.; Kern, D. Mechanism of Activating Mutations and Allosteric Drug Inhibition of the Phosphatase SHP2. Nat. Commun. 2018, 9, 4507. [Google Scholar] [CrossRef]
- Guner, F.O. History and Evolution of the Pharmacophore Concept in Computer-Aided Drug Design. Curr. Top. Med. Chem. 2002, 2, 1321–1332. [Google Scholar] [CrossRef]
- Guner, O.; Waldman, M.; Hoffman, R.; Kim, J.-H. Strategies for Database Mining and Pharmacophore Development. In Pharmacophore Perception, Development, and Use in Drug Design; International University Line: Portland, OR, USA, 2000; pp. 193–212. ISBN 0-9936817-6-1. [Google Scholar]
- Singh, P.; Kumar, V.; Lee, G.; Jung, T.S.; Ha, M.W.; Hong, J.C.; Lee, K.W. Pharmacophore-Oriented Identification of Potential Leads as CCR5 Inhibitors to Block HIV Cellular Entry. Int. J. Mol. Sci. 2022, 23, 16122. [Google Scholar] [CrossRef] [PubMed]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, Flexible, and Free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Amadei, A.; Linssen, A.B.; Berendsen, H.J. Essential Dynamics of Proteins. Proteins 1993, 17, 412–425. [Google Scholar] [CrossRef] [PubMed]
- David, C.C.; Jacobs, D.J. Principal Component Analysis: A Method for Determining the Essential Dynamics of Proteins. Methods Mol. Biol. 2014, 1084, 193–226. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yang, W.Y.; Zhou, L.; Li, L.P.; Wu, J.W.; Li, W.Y.; Du, S.; Ma, Y.; Wang, R.L. Exploring the Cause of the Dual Allosteric Targeted Inhibition Attaching to Allosteric Sites Enhancing SHP2 Inhibition. Mol. Divers. 2022, 26, 1567–1580. [Google Scholar] [CrossRef]
- Pires, D.E.V.; Blundell, T.L.; Ascher, D.B. PkCSM: Predicting Small-Molecule Pharmacokinetic and Toxicity Properties Using Graph-Based Signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef] [PubMed]
- Ruess, D.A.; Heynen, G.J.; Ciecielski, K.J.; Ai, J.; Berninger, A.; Kabacaoglu, D.; Görgülü, K.; Dantes, Z.; Wörmann, S.M.; Diakopoulos, K.N.; et al. Mutant KRAS-Driven Cancers Depend on PTPN11/SHP2 Phosphatase. Nat. Med. 2018, 24, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Petrocchi, A.; Grillo, A.; Ferrante, L.; Randazzo, P.; Prandi, A.; De Matteo, M.; Iaccarino, C.; Bisbocci, M.; Cellucci, A.; Alli, C.; et al. Discovery of a Novel Series of Potent SHP2 Allosteric Inhibitors. ACS Med. Chem. Lett. 2023, 14, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular Docking: A Powerful Approach for Structure-Based Drug Discovery. Curr. Comput. Aided. Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and Validation of a Genetic Algorithm for Flexible Docking11Edited by F. E. Cohen. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef]
- Kontoyianni, M. Docking and Virtual Screening in Drug Discovery. Methods Mol. Biol. 2017, 1647, 255–266. [Google Scholar] [CrossRef]
- Hess, B.; Kutzner, C.; Van Der Spoel, D.; Lindahl, E. GRGMACS 4: Algorithms for Highly Efficient, Load-Balanced, and Scalable Molecular Simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Kumar, R.; Lynn, A. g_mmpbsa—A GROMACS Tool for High-Throughput MM-PBSA Calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef]
- Khan, M.T.; Ali, S.; Zeb, M.T.; Kaushik, A.C.; Malik, S.I.; Wei, D.-Q. Gibbs Free Energy Calculation of Mutation in PncA and RpsA Associated with Pyrazinamide Resistance. Front. Mol. Biosci. 2020, 7, 52. [Google Scholar] [CrossRef]
- Studio, D. Dassault Systemes BIOVIA, Discovery Studio Modelling Environment, Release 4.5. Accelrys Softw. Inc. 2015, 17, 98–104. [Google Scholar] [CrossRef]
- van de Waterbeemd, H.; Gifford, E. ADMET in Silico Modelling: Towards Prediction Paradise? Nat. Rev. Drug Discov. 2003, 2, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26, 1PII of Original Article: S0169-409X(96)00423-1. The Article Was Originally Published in Advanced Drug Delivery Reviews 23 (1997) 3. [Google Scholar] [CrossRef]
- Kumar, V.; Parate, S.; Thakur, G.; Lee, G.; Ro, H.-S.; Kim, Y.; Kim, H.J.; Kim, M.O.; Lee, K.W. Identification of CDK7 Inhibitors from Natural Sources Using Pharmacoinformatics and Molecular Dynamics Simulations. Biomedicines 2021, 9, 1197. [Google Scholar] [CrossRef]
- Yadav, M.B.; Singh, P.; Jeong, Y.T. Regioselective Synthesis and Molecular Docking Studies of Functionalized Imidazo [1,2-a]Pyridine Derivatives through MCRs. Mol. Divers. 2023, 28, 171–182. [Google Scholar] [CrossRef]
- Sapay, N.; Tieleman, D.P. Combination of the CHARMM27 Force Field with United-Atom Lipid Force Fields. J. Comput. Chem. 2011, 32, 1400–1410. [Google Scholar] [CrossRef]
- Zoete, V.; Cuendet, M.A.; Grosdidier, A.; Michielin, O. SwissParam: A Fast Force Field Generation Tool for Small Organic Molecules. J. Comput. Chem. 2011, 32, 2359–2368. [Google Scholar] [CrossRef] [PubMed]
- Maisuradze, G.; Liwo, A.; Scheraga, H. Principal Component Analysis for Protein Folding Dynamics. J. Mol. Biol. 2008, 385, 312–329. [Google Scholar] [CrossRef] [PubMed]
- Kumbhar, N.; Nimal, S.; Barale, S.; Kamble, S.; Bavi, R.; Sonawane, K.; Gacche, R. Identification of Novel Leads as Potent Inhibitors of HDAC3 Using Ligand-Based Pharmacophore Modeling and MD Simulation. Sci. Rep. 2022, 12, 1712. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kumar, R.; Parate, S.; Danishuddin; Lee, G.; Kwon, M.; Jeong, S.-H.; Ro, H.-S.; Lee, K.W.; Kim, S.-W. Identification of Activated Cdc42-Associated Kinase Inhibitors as Potential Anticancer Agents Using Pharmacoinformatic Approaches. Biomolecules 2023, 13, 217. [Google Scholar] [CrossRef]
- Hoang, T.X.; Trovato, A.; Seno, F.; Banavar, J.R.; Maritan, A. Geometry and Symmetry Presculpt the Free-Energy Landscape of Proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 7960–7964. [Google Scholar] [CrossRef]
- Gautam, R.K.; Chattopadhyaya, M.C. Kinetics and Equilibrium Isotherm Modeling: Graphene-Based Nanomaterials for the Removal of Heavy Metals from Water. In Nanomaterials for Wastewater Remediation; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Parameters | Calculated Values |
|---|---|---|
| 1 | Total no. of molecules in the database (D) | 270 |
| 2 | Total number of active molecules in the database (A) | 20 |
| 3 | Total number of active molecules in the retrieved hits (Ht) | 24 |
| 4 | Number of retrieved hits by pharmacophore (Ha) | 19 |
| 5 | % Yield of actives [(Ha/Ht) × 100] | 79.16% |
| 6 | % Ratio of actives [(Ha/A × 100)] | 95% |
| 7 | False negative [A-Ha] | 1 |
| 8 | False positive [Ht-Ha] | 5 |
| 9 | Goodness of fit | 0.81 |
| 10 | Enrichment factor (EF) | 10.68 |
| Systems | Docking Score | RMSD (nm) | RMSF (nm) | Potential Energy (kJ/mol) | Number of Hydrogen Bonds | Binding Free Energy (ΔGbinding kJ/mol) | |
|---|---|---|---|---|---|---|---|
| Goldscore | Chemscore | Backbone Atoms | Backbone Atoms | ||||
| REF | 55.28 | −28.35 | 0.31 | 0.12 | −1,019,870 | 1.53 | −71.48 |
| Lig_1 | 79.37 | −37.47 | 0.28 | 0.12 | −1,019,239 | 3.57 | −151.28 |
| Lig_6 | 73.16 | −20.03 | 0.27 | 0.11 | −1,020,092 | 1.08 | −161.49 |
| Lig_14 | 68.37 | −26.43 | 0.24 | 0.12 | −1,019,950 | 2.49 | −107.13 |
| Name | Hydrogen Bond Interactions | van der Waals Interactions | π-π/π–Alkyl Interactions | |||
|---|---|---|---|---|---|---|
| Amino Acid | Amino Acid Atom | Ligand Atom | Distance (<3.5 Å) | |||
| Lig_1 | Arg111 | HE | O1 | 2.13 | Glu110, His114, Thr219, Lys242, Glu249, Asp489, Lys492, Gln495 | Leu216, Glu250, Leu254, Lys492, Pro491 |
| Phe113 | O | H43 | 2.76 | |||
| His114 | ND1 | H43 | 1.93 | |||
| Glu232 | O | H62 | 1.80 | |||
| Gln495 | HE21 | O28 | 2.18 | |||
| Lig_6 | Arg111 | O | H41 | 2.05 | Phe113, Asn217, Thr219, Thr253, Leu254, Gln257, Leu261, Pro491, Gln495, Met499 | His114, Glu249, Glu250, Lys492 |
| Lig_14 | His114 | HE2 | O9 | 2.89 | Arg111, Phe113, His116, Leu216, Asn217, Arg229, Glu250, Thr253 | Glu232, Leu254, Pro491 |
| Thr219 | NH | O10 | 2.37 | |||
| REF | Phe113 | O | H42 | 2.06 | Glu110, His114, Thr219, Lys292, Glu249, Asp489, Lys492, Gln495 | Leu254, Pro491 |
| Glu250 | O | H37 | 2.68 | |||
| Characters | Lig_1 | Lig_6 | Lig_14 |
|---|---|---|---|
| IUPAC name | methyl 4-[[6-hydroxy-7-[[4-(2-hydroxyethyl)piperazin-1-ium-1-yl]methyl]-4-methyl-3-oxo-benzofuran-2-yl]methyl]benzoate | [(3R,4R)-3-[(3S,4S)-6-(cyclopentoxy)-4-hydroxy-chroman-3-yl]-4-hydroxy-6-methoxy-chroman-7-yl]methyl-methyl-ammonium | N-[[(1S,2R,4S,5S)-5-(indolin-1-ylmethyl)quinuclidin-1-ium-2-yl]methyl]-4-methyl-benzenesulfonamide |
| SMILE ID | COC(=O)c1ccc(CC2Oc3c(C[NH+]4CCN(CCO)CC4)c(O)cc(C)c3C2=O)cc1 | C[NH2]Cc1cc2OC[C@@H]([C@H]3COc4ccc(OC5CCCC5)cc4[C@H]3O)[C@@H](O)c2cc1OC | Cc1ccc(cc1)S(=O)(=O)NC[C@H]2C[C@@H]3CC[N@H]2C[C@@H]3CN4CCc5ccccc45 |
| 2D Structure | 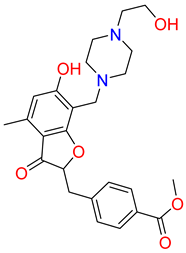 | 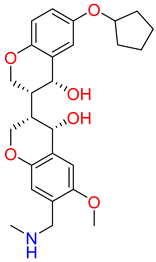 | 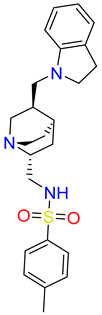 |
| ADME Properties | Lig_1 | Lig_6 | Lig_14 | REF (SHP099) | Cutoff | |
|---|---|---|---|---|---|---|
| Absorption | WS (log mol/L) | −3.3 | −4.28 | −3.17 | −3.84 | <−10 insoluble to <0 highly soluble |
| Caco-2 Permeability (log cm/s) | 0.18 | 1.042 | 1.66 | 0.999 | >0.90 | |
| IA human (% abs) | 63.17 | 90.35 | 96.62 | 92.23 | >30 | |
| SP (log KP) | −2.74 | −2.75 | −2.81 | −3.01 | >−2.5 | |
| P-glycoprotein Substrate | Yes | Yes | Yes | No | No | |
| P-glycoprotein I inhibitor | No | Yes | Yes | No | No | |
| P-glycoprotein II inhibitor | No | No | Yes | No | No | |
| Distribution | VDss (human) | 0.856 | 0.857 | 1.152 | 0.865 | <0.71 low to >2.81 high |
| Fraction unbound (human) | 0.262 | 0.138 | 0.146 | 0.308 | Numeric (Fu) | |
| BBBP (logBB) | −1.153 | −0.751 | −0.262 | −0.451 | >0.3 high to <−1 poor | |
| CNS permeability | −3.685 | −3.224 | −2.606 | −2.88 | >−0.2 high to <−3 poor | |
| Metabolism | CYP2D6 substrate | No | No | No | No | No |
| CYP2D6 inhibitor | No | No | No | No | No | |
| CYP3A4 substrate | Yes | Yes | Yes | Yes | No | |
| CYP3A4 inhibitor | No | Yes | Yes | No | No | |
| CYP1A2 inhibitor | No | No | No | Yes | No | |
| CYP2C19 inhibitor | No | No | No | No | No | |
| CYP2C9 inhibitor | No | No | No | No | No | |
| Excretion | TC (mL/min/kg) | 1.312 | 1.145 | 1.005 | 0.66 | Numeric (mL/min/kg) |
| Renal OCT2 substrate | No | No | Yes | No | No | |
| Toxicity Parameters | Lig_1 | Lig_6 | Lig_14 | REF (SHP099) | Cutoff | |
|---|---|---|---|---|---|---|
| Toxicity | AMES toxicity | No | No | No | No | Categorical (Yes/No) |
| Max. tolerated dose (human) | −0.15 | −0.12 | −0.42 | −0.18 | >0.477 mg/kg/day | |
| hERG I inhibitor | No | No | No | No | Categorical (Yes/No) | |
| hERG II inhibitor | Yes | Yes | Yes | Yes | Categorical (Yes/No) | |
| Oral Rat Acute Toxicity (LD50) | 2.944 | 2.72 | 2.61 | 2.932 | Numeric (mol/kg) | |
| Oral Rat Chronic Toxicity (LOAEL) | 2.852 | 1.76 | 1.86 | 0.508 | Numeric (mg/kg_bw/day) | |
| Hepatotoxicity | Yes | No | Yes | Yes | Categorical (Yes/No) | |
| SS | No | No | No | No | Categorical (Yes/No) | |
| T. Pyriformis toxicity | 0.29 | 0.294 | 0.33 | 0.513 | >0.5 μg/L Toxic | |
| Minnow toxicity | 1.488 | 1.498 | 1.107 | 2.056 | <−0.3 Toxic | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.; Kumar, V.; Lee, K.W.; Hong, J.C. Discovery of Novel Allosteric SHP2 Inhibitor Using Pharmacophore-Based Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and Principal Component Analysis. Pharmaceuticals 2024, 17, 935. https://doi.org/10.3390/ph17070935
Singh P, Kumar V, Lee KW, Hong JC. Discovery of Novel Allosteric SHP2 Inhibitor Using Pharmacophore-Based Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and Principal Component Analysis. Pharmaceuticals. 2024; 17(7):935. https://doi.org/10.3390/ph17070935
Chicago/Turabian StyleSingh, Pooja, Vikas Kumar, Keun Woo Lee, and Jong Chan Hong. 2024. "Discovery of Novel Allosteric SHP2 Inhibitor Using Pharmacophore-Based Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and Principal Component Analysis" Pharmaceuticals 17, no. 7: 935. https://doi.org/10.3390/ph17070935
APA StyleSingh, P., Kumar, V., Lee, K. W., & Hong, J. C. (2024). Discovery of Novel Allosteric SHP2 Inhibitor Using Pharmacophore-Based Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and Principal Component Analysis. Pharmaceuticals, 17(7), 935. https://doi.org/10.3390/ph17070935