
Supplementation of SkQ1 Increases Mouse In Vitro Oocyte Maturation and Subsequent Embryonic Development by Reducing Oxidative Stress

Abstract
1. Introduction
2. Results
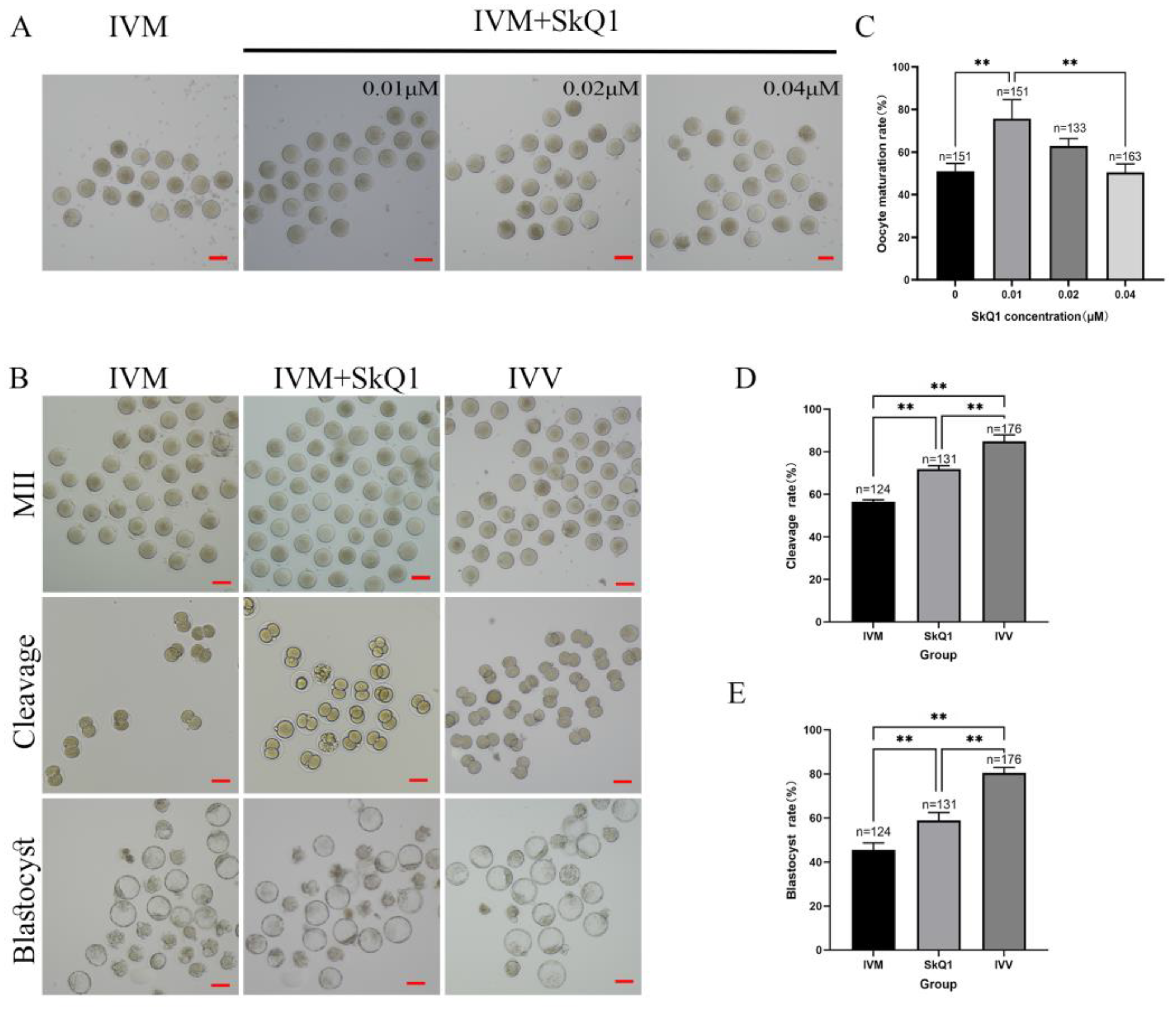
2.1. SkQ1 Improved the Maturation Rates and Developmental Potential of In Vitro-Matured Mouse Oocytes
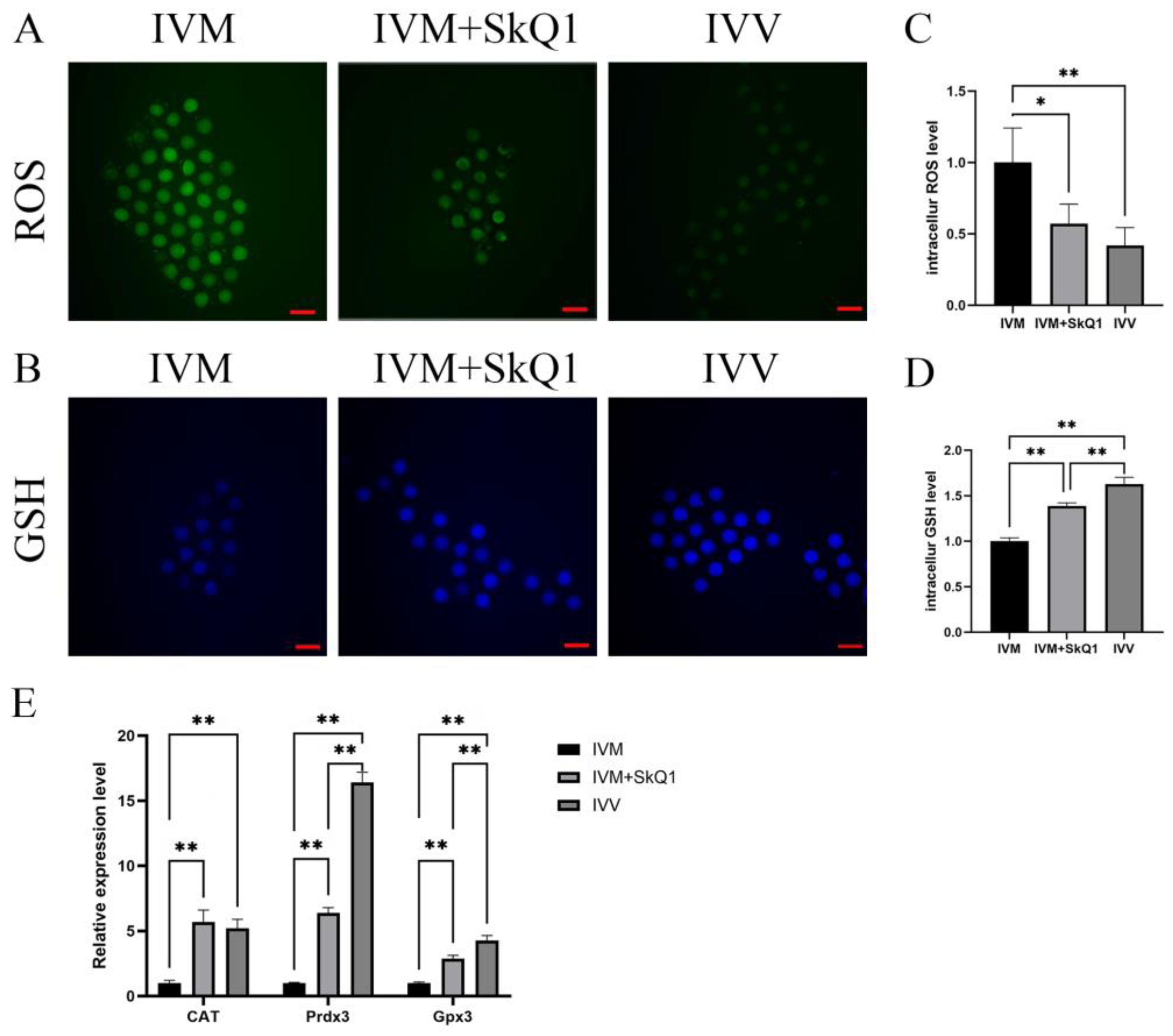
2.2. SkQ1 Reduced Oxidative Stress Levels and Enhanced Antioxidant Ability in In Vitro-Matured Mouse Oocytes
2.3. SkQ1 Increased Mitochondrial Membrane Potential in In Vitro-Matured Mouse Oocytes
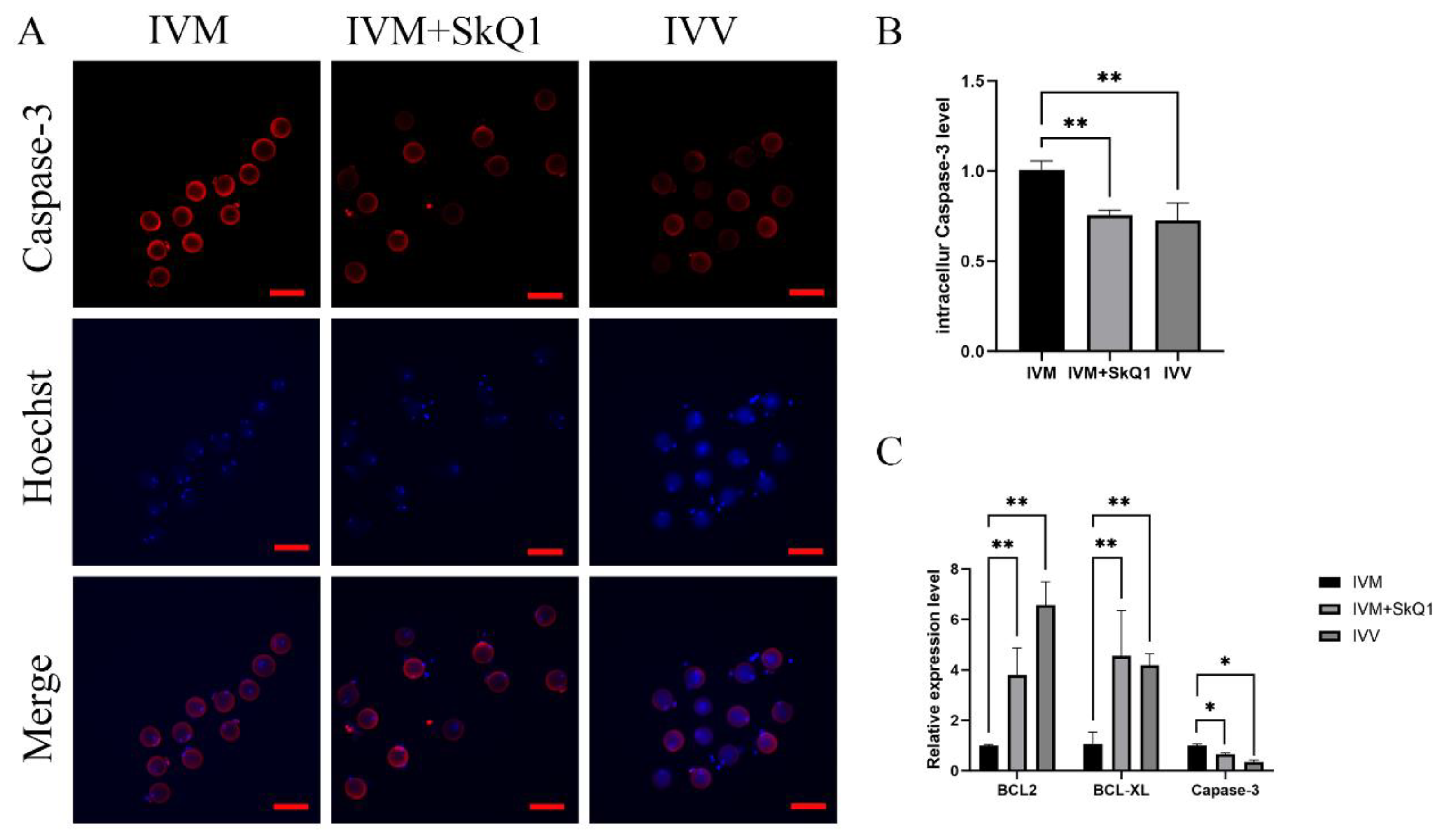
2.4. SkQ1 Inhibited Apoptosis in In Vitro-Matured Mouse Oocytes
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Germinal Vesicle Oocyte Collection
4.3. Oocyte In Vitro Maturation and Drug Treatment
4.4. Collection of In Vivo-Matured Oocytes
4.5. Experimental Design
4.6. In Vitro Fertilization
4.7. Quantitative PCR
4.8. ROS Level Measurement
4.9. GSH Level Measurement
4.10. Mitochondrial Membrane Potential Measurement
4.11. Immunofluorescence
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IVM | In vitro oocyte maturation |
| ETC | Electron transport chain |
| DNA | Deoxyribonucleic acid |
| RNA | Ribonucleic acid |
| mtDNA | Mitochondrial DNA |
| ICR | Institute of Cancer Research |
| PMSG | Pregnant mare serum gonadotropin |
| hCG | Human chorionic gonadotropin |
| PB | Polar body |
| ZP | Zona pellucida |
| IVF | In vitro fertilization |
| qPCR | Quantitative real-time PCR |
| MMP | Mitochondrial membrane potential |
| GSH | Glutathione |
| DPBS | Dulbecco’s Phosphate-Buffered Saline |
| COCs | Cumulus–oocyte complexes |
| GV | Germinal vesicle |
| MII | Metaphase II |
| DMSO | Dimethyl sulfoxide |
| HTF | Human tubal fluid |
| h | Hour |
| m | Minute |
| ROS | Reactive oxygen species |
| TMRE | Tetramethylrhodamine |
| % | Percentage |
References
- Ata, B.; Shalom-Paz, E.; Chian, R.C.; Tan, S.L. In vitro maturation of oocytes as a strategy for fertility preservation. Clin. Obstet. Gynecol. 2010, 53, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Marthandan, S.; Murphy, M.P.; Billett, E.; Barnett, Y. An investigation of the effects of MitoQ on human peripheral mononuclear cells. Free Radic. Res. 2011, 45, 351–358. [Google Scholar] [CrossRef] [PubMed]
- De Vos, M.; Grynberg, M.; Ho, T.M.; Yuan, Y.; Albertini, D.F.; Gilchrist, R.B. Perspectives on the development and future of oocyte IVM in clinical practice. J. Assist. Reprod. Genet. 2021, 38, 1265–1280. [Google Scholar] [CrossRef] [PubMed]
- Guerin, P.; El, M.S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative stress in an assisted reproductive techniques setting. Fertil. Steril. 2006, 86, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, M.; Machado, S.A.; Stojkovic, P.; Zakhartchenko, V.; Hutzler, P.; Goncalves, P.B.; Wolf, E. Mitochondrial distribution and adenosine triphosphate content of bovine oocytes before and after in vitro maturation: Correlation with morphological criteria and developmental capacity after in vitro fertilization and culture. Biol. Reprod. 2001, 64, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Amoushahi, M.; Salehnia, M.; Ghorbanmehr, N. The mitochondrial DNA copy number, cytochrome c oxidase activity and reactive oxygen species level in metaphase II oocytes obtained from in vitro culture of cryopreserved ovarian tissue in comparison with in vivo-obtained oocyte. J. Obstet. Gynaecol. Res. 2018, 44, 1937–1946. [Google Scholar] [CrossRef] [PubMed]
- Al-Zubaidi, U.; Adhikari, D.; Cinar, O.; Zhang, Q.; Yuen, W.S.; Murphy, M.P.; Rombauts, L.; Robker, R.L.; Carroll, J. Mitochondria-targeted therapeutics, MitoQ and BGP-15, reverse aging-associated meiotic spindle defects in mouse and human oocytes. Hum. Reprod. 2021, 36, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Zhuan, Q.; Luo, Y.; Liu, H.; Meng, L.; Du, X.; Wu, G.; Hou, Y.; Li, J.; Fu, X. Mito-Q promotes porcine oocytes maturation by maintaining mitochondrial thermogenesis via UCP2 downregulation. Theriogenology 2022, 187, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, S.M.; Amidi, F.; Shamsara, M.; Nazarian, H.; Eini, F.; Hosseinzadeh, S.F.; Majidi, Z.M.; Ghaffari, N.M.; Daliri, J.M. Exposing Mouse Oocytes to MitoQ During In Vitro Maturation Improves Maturation and Developmental Competence. Iran. J. Biotechnol. 2020, 18, e2454. [Google Scholar]
- Nikmard, F.; Hosseini, E.; Bakhtiyari, M.; Ashrafi, M.; Amidi, F.; Aflatoonian, R. The boosting effects of melatonin on the expression of related genes to oocyte maturation and antioxidant pathways: A polycystic ovary syndrome- mouse model. J. Ovarian Res. 2022, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Kala, M.; Shaikh, M.V.; Nivsarkar, M. Equilibrium between anti-oxidants and reactive oxygen species: A requisite for oocyte development and maturation. Reprod. Med. Biol. 2017, 16, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.Y.; Gao, Y.; Jiao, G.Z.; Sun, M.J.; Wu, X.F.; Wang, T.Y.; Li, H.; Tan, J.H. Antioxidant supplementation overcomes the deleterious effects of maternal restraint stress-induced oxidative stress on mouse oocytes. Reproduction 2013, 146, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhao, H.; Wang, Z.; Zhang, C.; Bian, Y.; Liu, X.; Zhang, C.; Zhang, X.; Zhao, Y. Quercetin promotes in vitro maturation of oocytes from humans and aged mice. Cell Death Dis. 2020, 11, 965. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Troger, A.; Spahiu, V.; Perekhvatova, N.; Skulachev, M.; Petrov, A.; Chernyak, B.; Asbell, P. The Role of SKQ1 (Visomitin) in Inflammation and Wound Healing of the Ocular Surface. Ophthalmol. Ther. 2019, 8, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Vays, V.B.; Eldarov, C.M.; Vangely, I.M.; Kolosova, N.G.; Bakeeva, L.E.; Skulachev, V.P. Antioxidant SkQ1 delays sarcopenia-associated damage of mitochondrial ultrastructure. Aging 2014, 6, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Skulachev, V.P. Cationic antioxidants as a powerful tool against mitochondrial oxidative stress. Biochem. Biophys. Res. Commun. 2013, 441, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Labarta, E.; de Los, S.M.; Escriba, M.J.; Pellicer, A.; Herraiz, S. Mitochondria as a tool for oocyte rejuvenation. Fertil. Steril. 2019, 111, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Chiaratti, M.R.; Garcia, B.M.; Carvalho, K.F.; Machado, T.S.; Ribeiro, F.; Macabelli, C.H. The role of mitochondria in the female germline: Implications to fertility and inheritance of mitochondrial diseases. Cell Biol. Int. 2018, 42, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, F.C.; Leal, C.; Zvi, R.; Hansen, P.J. Effects of melatonin on production of reactive oxygen species and developmental competence of bovine oocytes exposed to heat shock and oxidative stress during in vitro maturation. Zygote 2019, 27, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Menezo, Y.J.; Silvestris, E.; Dale, B.; Elder, K. Oxidative stress and alterations in DNA methylation: Two sides of the same coin in reproduction. Reprod. Biomed. Online 2016, 33, 668–683. [Google Scholar] [CrossRef] [PubMed]
- Brad, A.M.; Bormann, C.L.; Swain, J.E.; Durkin, R.E.; Johnson, A.E.; Clifford, A.L.; Krisher, R.L. Glutathione and adenosine triphosphate content of in vivo and in vitro matured porcine oocytes. Mol. Reprod. Dev. 2003, 64, 492–498. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kang, J.T.; Park, S.J.; Kim, S.J.; Moon, J.H.; Saadeldin, I.M.; Jang, G.; Lee, B.C. Effect of 7,8-dihydroxyflavone as an antioxidant on in vitro maturation of oocytes and development of parthenogenetic embryos in pigs. J. Reprod. Dev. 2013, 59, 450–456. [Google Scholar] [CrossRef]
- Simkin, J.L.; Work, T.S. Biochemical approaches to the problem of protein synthesis. Nature 1957, 179, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Zare, Z.; Masteri Farahani, R.; Salehi, M.; Piryaei, A.; Ghaffari Novin, M.; Fadaei Fathabadi, F.; Mohammadi, M.; Dehghani-Mohammadabadi, M. Effect of L-carnitine supplementation on maturation and early embryo development of immature mouse oocytes selected by brilliant cresyle blue staining. J. Assist. Reprod. Genet. 2015, 32, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Galkin, I.I.; Pletjushkina, O.Y.; Zinovkin, R.A.; Zakharova, V.V.; Birjukov, I.S.; Chernyak, B.V.; Popova, E.N. Mitochondria-targeted antioxidants prevent TNFalpha-induced endothelial cell damage. Biochemistry 2014, 79, 124–130. [Google Scholar] [PubMed]
- Ubaldi, F.; Rienzi, L. Morphological selection of gametes. Placenta 2008, 29 (Suppl. B), 115–120. [Google Scholar] [CrossRef]
- Xing, X.; Liang, Y.; Li, Y.; Zhao, Y.; Zhang, Y.; Li, Z.; Li, Z.; Wu, Z. Fisetin Delays Postovulatory Oocyte Aging by Regulating Oxidative Stress and Mitochondrial Function through Sirt1 Pathway. Molecules 2023, 28, 5533. [Google Scholar] [CrossRef] [PubMed]
- Zare, Z.; Abouhamzeh, B.; Masteri, F.R.; Salehi, M.; Mohammadi, M. Supplementation of L-carnitine during in vitro maturation of mouse oocytes affects expression of genes involved in oocyte and embryo competence: An experimental study. Int. J. Reprod. Biomed. 2017, 15, 779–786. [Google Scholar] [CrossRef]
- Zhuan, Q.; Ma, H.; Chen, J.; Luo, Y.; Luo, Y.; Gao, L.; Hou, Y.; Zhu, S.; Fu, X. Cytoplasm lipids can be modulated through hormone-sensitive lipase and are related to mitochondrial function in porcine IVM oocytes. Reprod. Fertil. Dev. 2020, 32, 667–675. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′ → 3′) | Product Length (bp) | GenBank Acc No. | Annealing Temp (°C) |
|---|---|---|---|---|
| Gapdh | F:CAGTCCATGCCATCACTGCCA R:ATGCCAGTGAGCTTCCCGTTC | 163 | NC_000072 | 59.76 |
| CAT | F:GACAAAATGCTTCAGGGCCGC R:ACCCTGGTTGTCATGCATGCA | 156 | NC_009804 | 58.00 |
| Prdx3 | F:GTGCTGTTGGAAAGTGCTGGC R:TGGGCAGACTTCTCCATGGGT | 171 | NM_007452 | 59.76 |
| Gpx3 | F:AAACAGGAGCCAGGCGAGAAC R:AGTGGGAGGGCAGGAGTTCTT | 162 | NM_001329860 | 59.76 |
| Bcl2 | F:GCATCTGCACACCTGGATCCA R:ACTTGTGGCCCAGGTATGCAC | 170 | NM_009741 | 59.50 |
| Bcl-XL | F:TTTTTCTCCTTTGGCGGGGCA R:TCCACAAAAGTGTCCCAGCCG | 152 | NM_001289716 | 59.50 |
| Caspase-3 | F:TTCATCATTCAGGCCTGCCGG R:TGAACCACGACCCGTCCTTTG | 175 | NM_001284409 | 59.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhang, Y.; Cao, J.; Xing, X.; Liang, Y.; Zhang, Y.; Tang, X.; Lin, S.; Wu, Z.; Li, Z.; et al. Supplementation of SkQ1 Increases Mouse In Vitro Oocyte Maturation and Subsequent Embryonic Development by Reducing Oxidative Stress. Pharmaceuticals 2024, 17, 455. https://doi.org/10.3390/ph17040455
Li Z, Zhang Y, Cao J, Xing X, Liang Y, Zhang Y, Tang X, Lin S, Wu Z, Li Z, et al. Supplementation of SkQ1 Increases Mouse In Vitro Oocyte Maturation and Subsequent Embryonic Development by Reducing Oxidative Stress. Pharmaceuticals. 2024; 17(4):455. https://doi.org/10.3390/ph17040455
Chicago/Turabian StyleLi, Zheng, Yiqian Zhang, Jinping Cao, Xupeng Xing, Yalin Liang, Yuxing Zhang, Xiaopeng Tang, Shengyi Lin, Zhenfang Wu, Zicong Li, and et al. 2024. "Supplementation of SkQ1 Increases Mouse In Vitro Oocyte Maturation and Subsequent Embryonic Development by Reducing Oxidative Stress" Pharmaceuticals 17, no. 4: 455. https://doi.org/10.3390/ph17040455
APA StyleLi, Z., Zhang, Y., Cao, J., Xing, X., Liang, Y., Zhang, Y., Tang, X., Lin, S., Wu, Z., Li, Z., & Huang, S. (2024). Supplementation of SkQ1 Increases Mouse In Vitro Oocyte Maturation and Subsequent Embryonic Development by Reducing Oxidative Stress. Pharmaceuticals, 17(4), 455. https://doi.org/10.3390/ph17040455