

Production and Immunogenicity Assessment of LTp50: An Escherichia coli-Made Chimeric Antigen Targeting S1- and S2-Epitopes from the SARS-CoV-2/BA.5 Spike Protein


 , , , ,
, , , ,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Structural Description of LTp50
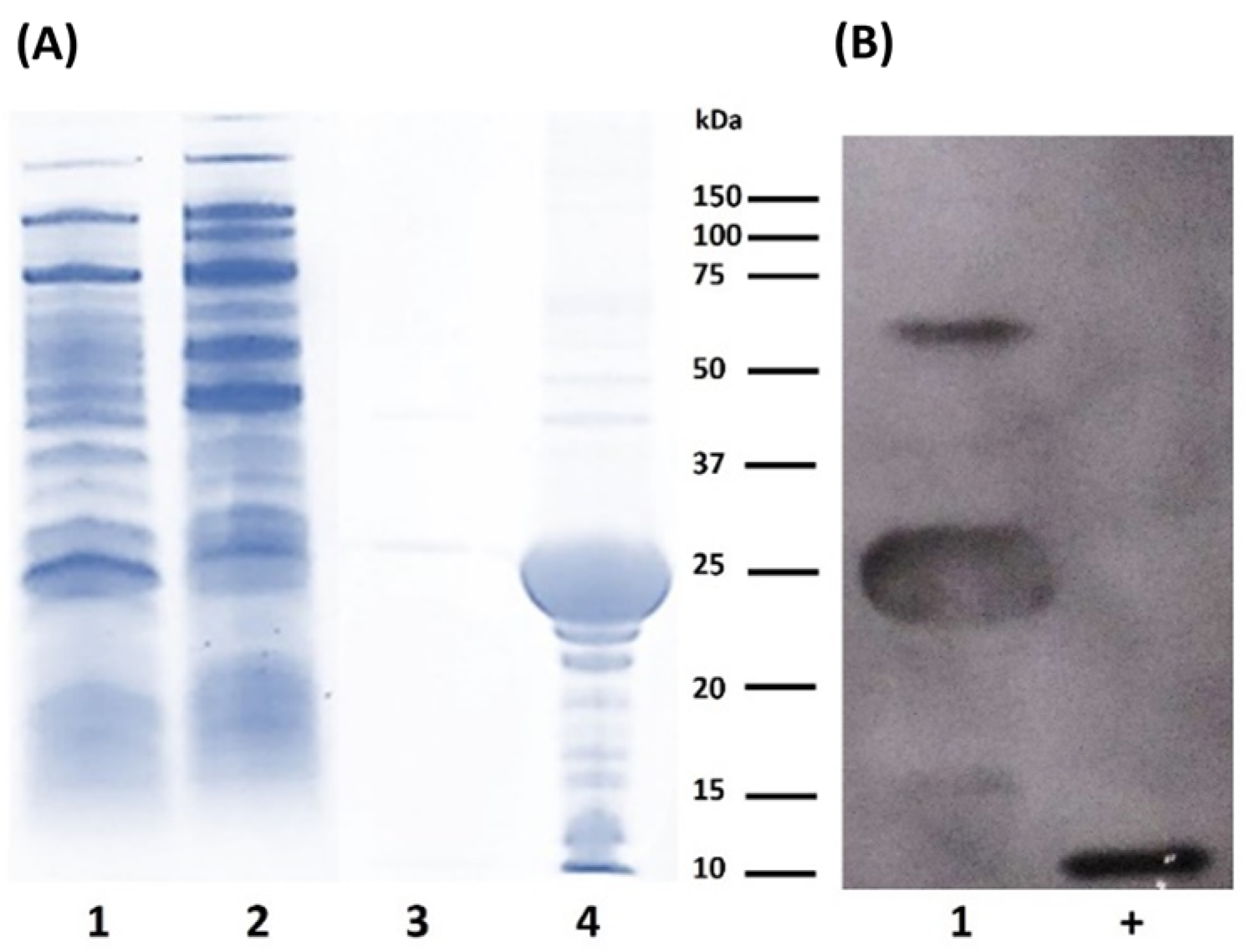
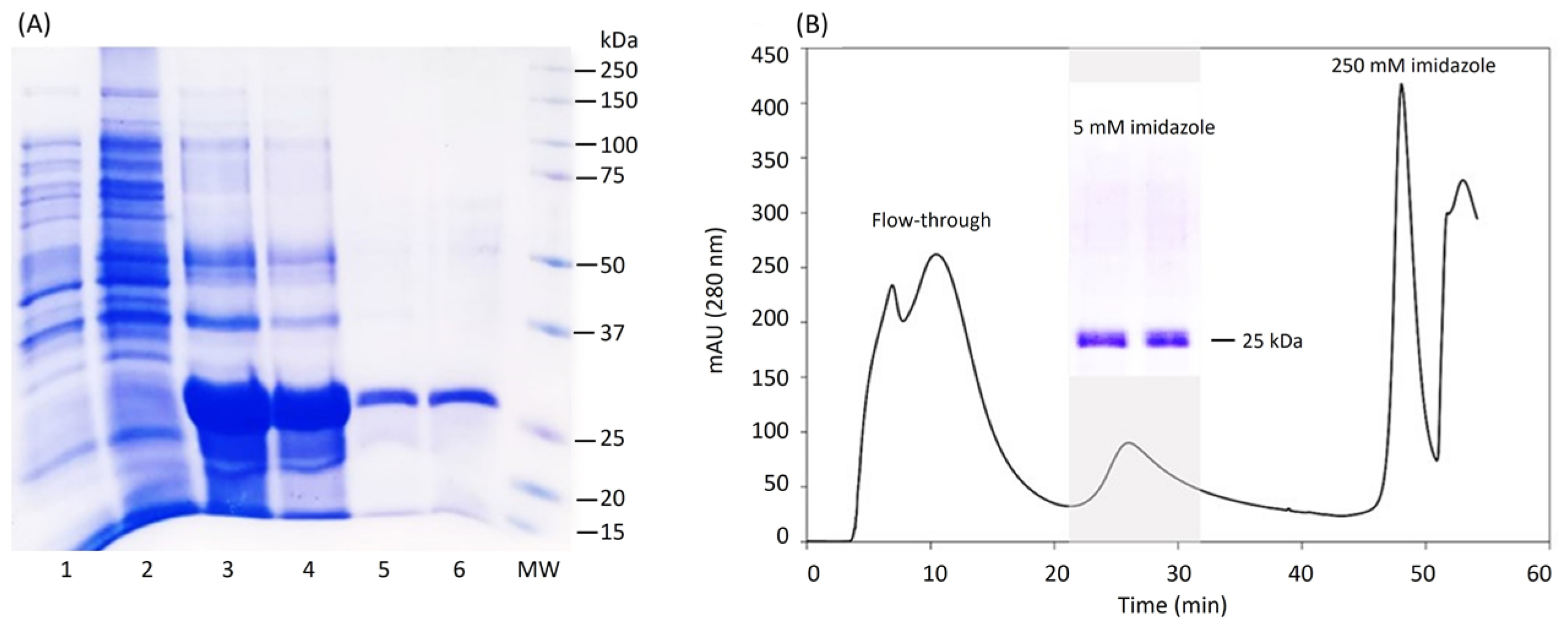
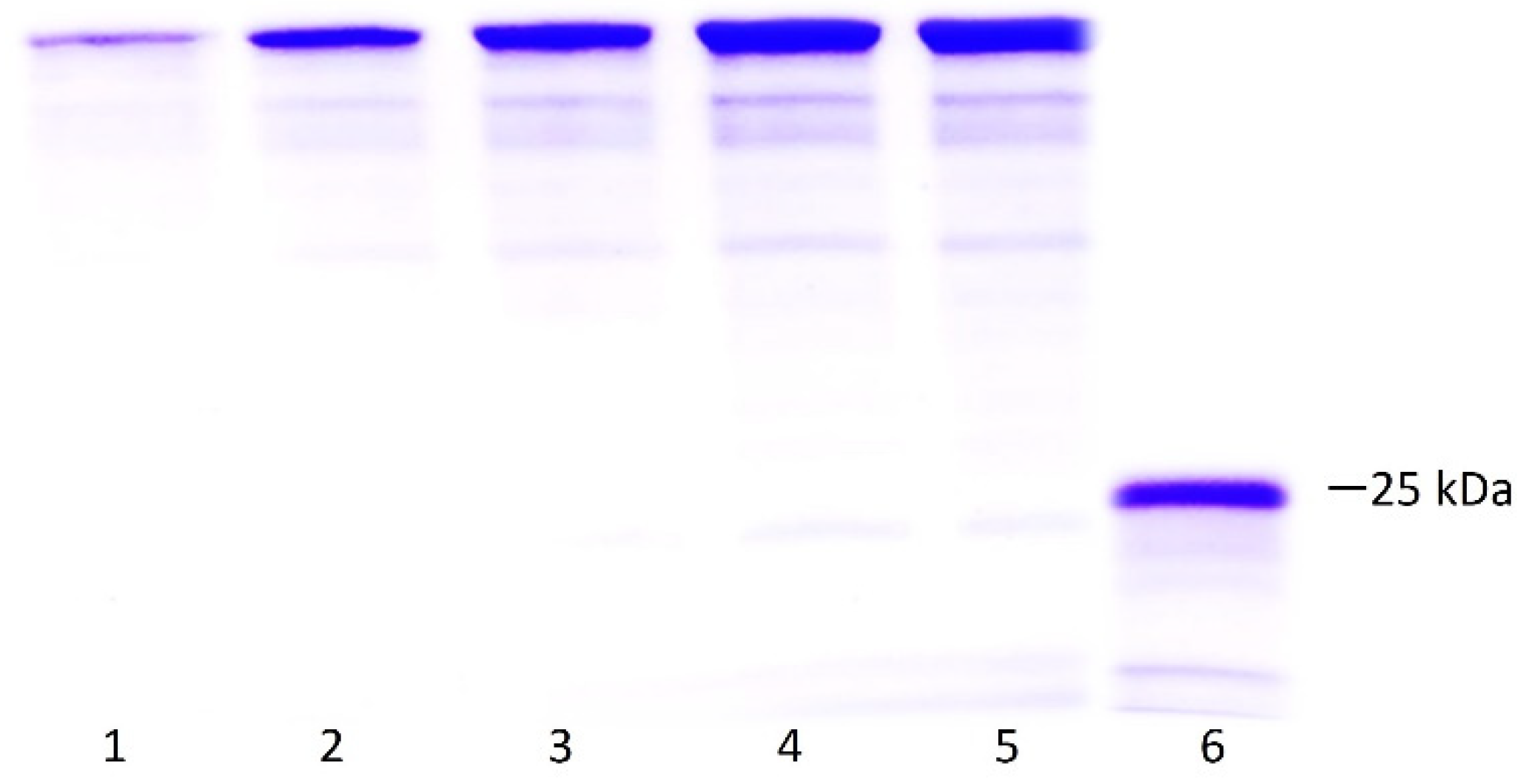
2.2. Production and Purification of LTp50
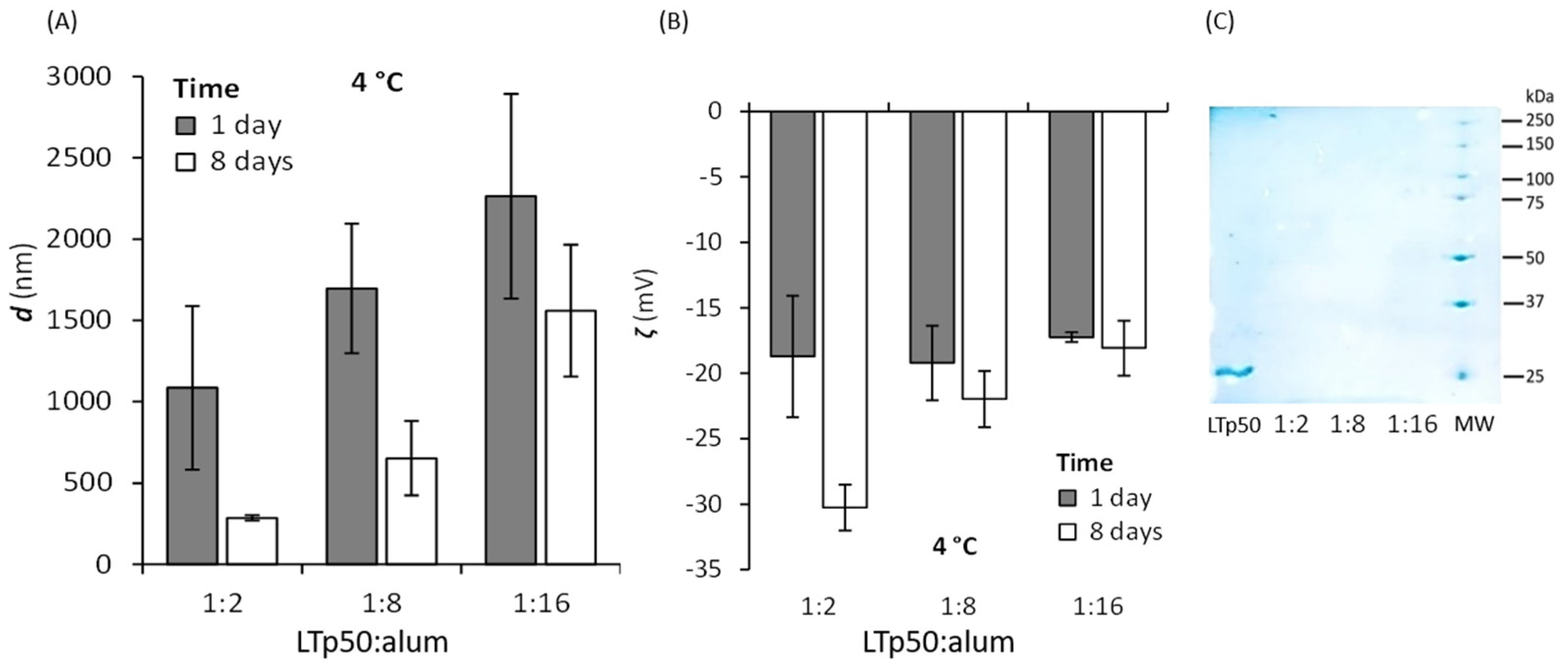
2.3. LTp50 Formulation with Alum
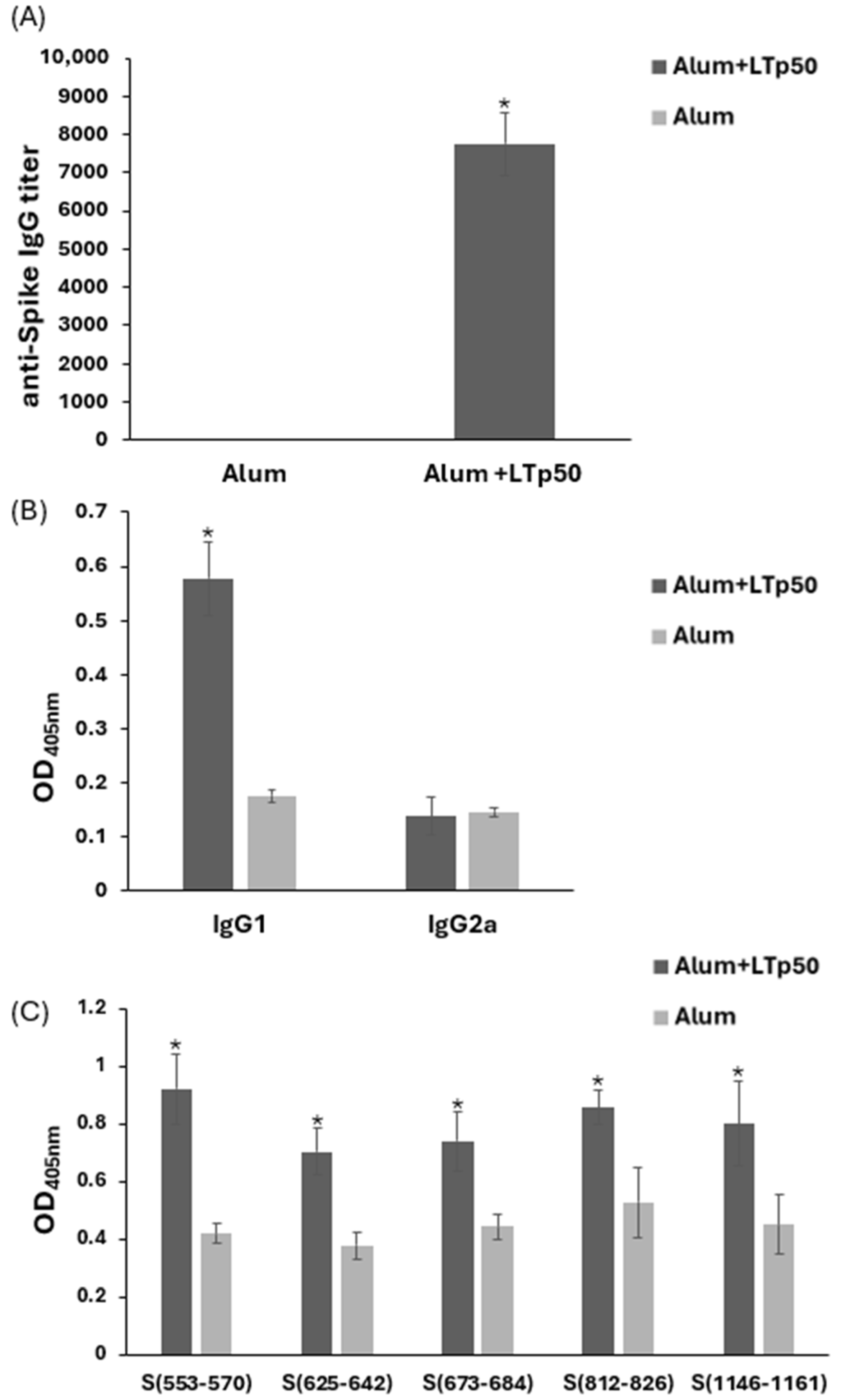
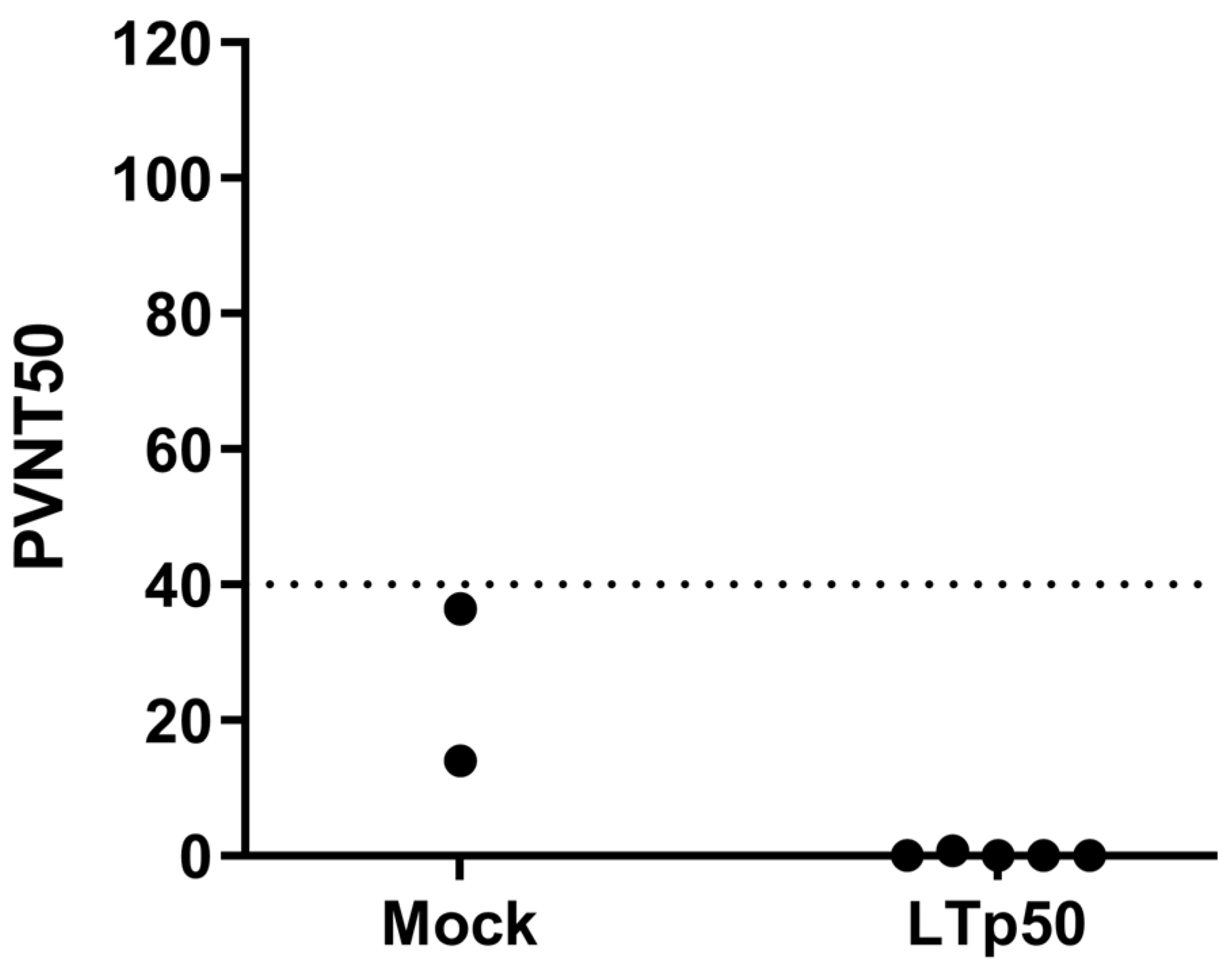
2.4. Assessment of LTp50 Immunogenicity
3. Discussion
4. Materials and Methods
4.1. Gene Design and Expression Vector Construction
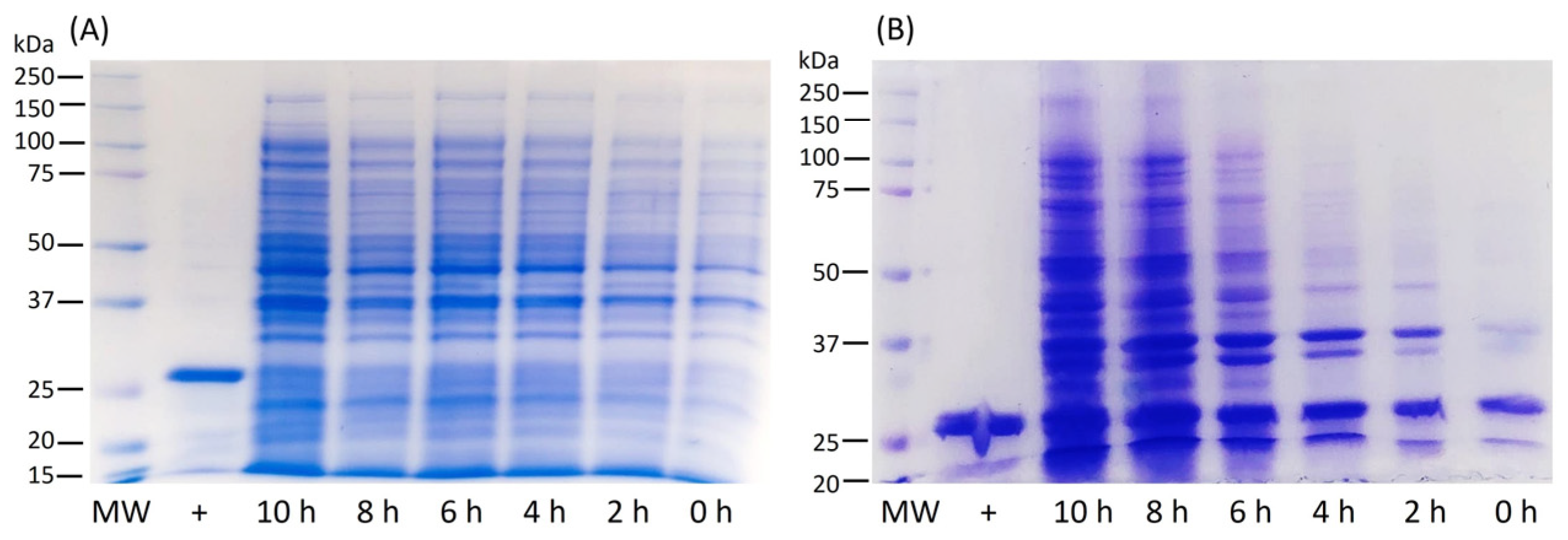
4.2. Protein Expression in Flasks
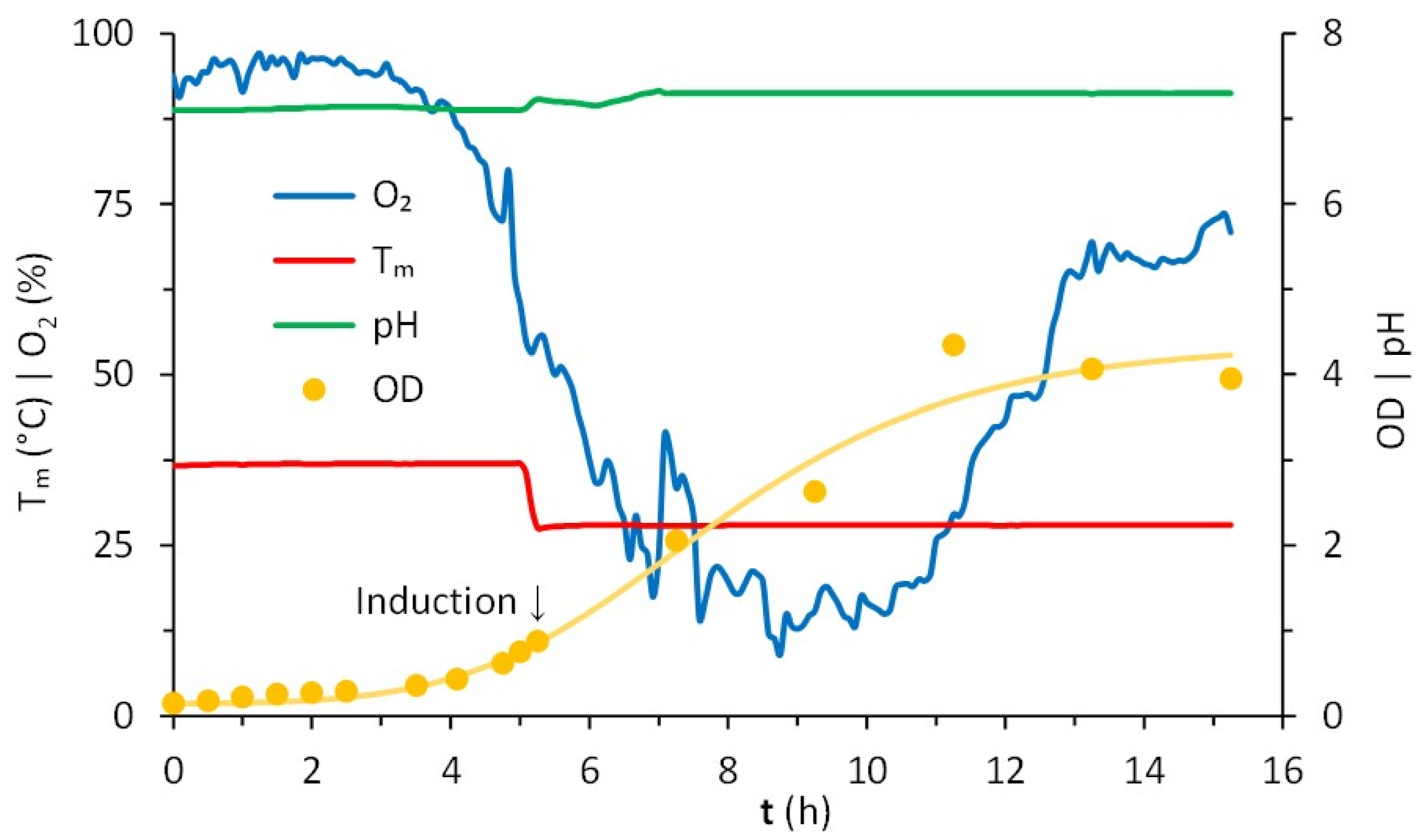
4.3. Protein Expression in Bioreactor
4.4. Protein Extraction and Purification
4.5. Protein Analysis Protocols
4.6. Vaccine Formulation
4.7. Immunogenicity Assessment
4.8. Pseudotyped Virus-Based Neutralization Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Coronavirus Disease (COVID-19) Pandemic. Available online: https://www.who.int/europe/emergencies/situations/covid-19 (accessed on 10 June 2023).
- WHO. WHO Chief Declares End to COVID-19 as a Global Health Emergency. Available online: https://news.un.org/en/story/2023/05/1136367 (accessed on 10 June 2023).
- WHO. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 10 June 2023).
- Chen, C.; Haupert, S.R.; Zimmermann, L.; Shi, X.; Fritsche, L.G.; Mukherjee, B. Global Prevalence of Post-Coronavirus Disease 2019 (COVID-19) Condition or Long COVID: A Meta-Analysis and Systematic Review. J. Infect. Dis. 2022, 226, 1593–1607. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.; Andreadakis, Z.; Jumar, A.; Gomez-Roman, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug Dicov. 2020, 19, 305–306. [Google Scholar] [CrossRef]
- Mathieu, E.; Ritchie, H.; Rodés-Guirao, L.; Appel, C.; Giattino, C.; Hasell, J.; Macdonald, B.; Dattani, S.; Beltekian, D.; Ortiz-Ospina, E. Max Roser Coronavirus Pandemic (COVID-19). 2020. Available online: https://ourworldindata.org/coronavirus (accessed on 10 June 2023).
- Gasmi, A.; Srinath, S.; Dadar, M.; Pivina, L.; Menzel, A.; Gasmi-Benahmed, A.; Chirumbolo, S.; Bjørklund, G. A global survey in the developmental landscape of possible vaccination strategies for COVID-19. Clin. Immunol. 2022, 237, 108958. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.K.; Rahman, M.M.; Siddiqi, U.R.; Howlader, A.; Tushar, M.A.; Qazi, A. Global landscape of COVID-19 vaccination progress: Insight from an exploratory data analysis. Hum. Vaccin. Immunother. 2022, 18, 2025009. [Google Scholar] [CrossRef] [PubMed]
- Calvo Fernández, E.; Zhu, L.Y. Racing to immunity: Journey to a COVID-19 vaccine and lessons for the future. Br. J. Clin. Pharmacol. 2021, 87, 3408–3424. [Google Scholar] [CrossRef]
- Huang, H.Y.; Wang, S.H.; Tang, Y.; Sheng, W.; Zuo, C.J.; Wu, D.W.; Fang, H.; Du, Q.; Li, N. Landscape and progress of global COVID-19 vaccine development. Hum. Vaccines Immunother. 2021, 17, 3276–3280. [Google Scholar] [CrossRef] [PubMed]
- Garçon, N.; Vaughn, D.W.; Didierlaurent, A.M. Development and evaluation of AS03, an Adjuvant System containing α-tocopherol and squalene in an oil-in-water emulsion. Expert. Rev. Vaccines 2012, 11, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.G.; Su, D.; Song, T.Z.; Zeng, Y.; Huang, W.; Wu, J.; Xu, R.; Luo, P.; Yang, X.; Zhang, X.; et al. S-Trimer, a COVID-19 subunit vaccine candidate, induces protective immunity in nonhuman primates. Nat. Commun. 2021, 12, 1346. [Google Scholar] [CrossRef]
- Tran, T.N.M.; May, B.P.; Ung, T.T.; Nguyen, M.K.; Nguyen, T.T.T.; Dinh, V.L.; Doan, C.C.; Tran, T.V.; Khong, H.; Nguyen, T.T.T.; et al. Preclinical Immune Response and Safety Evaluation of the Protein Subunit Vaccine Nanocovax for COVID-19. Front. Immunol. 2021, 12, 766112. [Google Scholar] [CrossRef]
- Dai, L.; Zheng, T.; Xu, K.; Han, Y.; Xu, L.; Huang, E.; An, Y.; Cheng, Y.; Li, S.; Liu, M.; et al. A Universal Design of Betacoronavirus Vaccines against COVID-19, MERS, and SARS. Cell 2020, 182, 722–733.e11. [Google Scholar] [CrossRef]
- Valdes-Balbin, Y.; Santana-Mederos, D.; Quintero, L.; Fernández, S.; Rodriguez, L.; Sanchez Ramirez, B.; Perez-Nicado, R.; Acosta, C.; Méndez, Y.; Ricardo, M.G.; et al. SARS-CoV-2 RBD-Tetanus Toxoid Conjugate Vaccine Induces a Strong Neutralizing Immunity in Preclinical Studies. ACS Chem. Biol. 2021, 16, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Wei, J.; Kundu, R.T.; Adhikari, R.; Liu, Z.; Lee, J.; Versteeg, L.; Poveda, C.; Keegan, B.; Villar, M.J.; et al. Genetic modification to design a stable yeast-expressed recombinant SARS-CoV-2 receptor binding domain as a COVID-19 vaccine candidate. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129893. [Google Scholar] [CrossRef] [PubMed]
- Ryzhikov, A.B.; Ryzhikov, E.А.; Bogryantseva, M.P.; Danilenko, E.D.; Imatdinov, I.; Nechaeva, E.A.; Pyankov, O.V.; Pyankova, O.G.; Susloparov, I.M.; Taranov, O.S.; et al. Immunogenicity and protectivity of the peptide vaccine against SARS-CoV-2. Ann. Russ. Acad. Med. Sci. 2021, 76, 5–19. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.M.; Liu, M.C.; Chen, Y.H.; Lee, W.S.; Hwang, S.J.; Cheng, S.H.; Ko, W.C.; Hwang, K.P.; Wang, N.C.; Lee, Y.L.; et al. Safety and immunogenicity of CpG 1018 and aluminium hydroxide-adjuvanted SARS-CoV-2 S-2P protein vaccine MVC-COV1901: Interim results of a large-scale, double-blind, randomised, placebo-controlled phase 2 trial in Taiwan. Lancet Respir. Med. 2021, 9, 1396–1406. [Google Scholar] [CrossRef] [PubMed]
- Richmond, P.; Hatchuel, L.; Dong, M.; Ma, B.; Hu, B.; Smolenov, I.; Li, P.; Liang, P.; Han, H.H.; Liang, J.; et al. Safety and immunogenicity of S-Trimer (SCB-2019), a protein subunit vaccine candidate for COVID-19 in healthy adults: A phase 1, randomised, double-blind, placebo-controlled trial. Lancet 2021, 397, 682–694. [Google Scholar] [CrossRef] [PubMed]
- BioSpace. Clover Biopharmaceuticals Completes Enrollment of Adult and Elderly Population in SPECTRA Global Phase 2/3 Clinical Trial for its COVID-19 Vaccine Candidate. 2021. Available online: https://www.biospace.com/article/releases/clover-biopharmaceuticals-completes-enrollment-of-adult-and-elderly-population-in-spectra-global-phase-2-3-clinical-trial-for-its-covid-19-vaccine-candidate/ (accessed on 10 June 2023).
- Clover Biopharmaceuticals. Clover’s COVID-19 Vaccine Candidate Demonstrates 79% Efficacy Against Delta in Global Phase 2/3 SPECTRA Trial Dominated by Variants of Concern and Interest. 2021. Available online: https://www.prnewswire.com/news-releases/clovers-covid-19-vaccine-candidate-demonstrates-79-efficacy-against-delta-in-global-phase-23-spectra-trial-dominated-by-variants-of-concern-and-interest-301382628.html (accessed on 10 June 2023).
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li Ch Fang, X.; Wang Ch Zhao, X.; Huang, E.; Wu Ch Zhong, Z.; Wang, F.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Bernal, F.; Ricardo-Cobas, M.C.; Martín-Bauta, Y.; Navarro-Rodríguez, Z.; Piñera-Martínez, M.; Quintana-Guerra, J.; Urrutia-Pérez, K.; Urrutia-Pérez, K.; Chávez-Chong, C.O.; Azor-Hernández, J.L.; et al. Safety, tolerability, and immunogenicity of a SARS-CoV-2 recombinant spike protein vaccine: A randomised, double-blind, placebo-controlled, phase 1-2 clinical trial (ABDALA Study). eClinicalMedicine 2022, 46, 101383. [Google Scholar] [CrossRef]
- Lemos-Perez, G.; Chavez-Valdes, S.; Gonzalez-Formental, H.; Freyre-Corrales, G.; Vazquez-Arteaga, A.; Alvarez-Acevedo, B.; Avila-Díaz, L.; Martínez-Rosales, R.U.; Chacon-Quintero, Y.; Coizeau-Rodriguez, E.; et al. Elevated Antibody Titers in Abdala Vaccinees Evaluated by Elecsys® Anti-SARS-Cov-2 S Highly Correlate with UMELISA SARS-Cov-2 ANTI RBD, ACE-2 Binding Inhibition and Viral Neutralization Assays. J. Biotechnol. Biomed. 2022, 5, 151–157. [Google Scholar] [CrossRef]
- Pollet, J.; Chen, W.H.; Versteeg, L.; Keegan, B.; Zhan, B.; Wei, J.; Liu, Z.; Lee, J.; Kundu, R.; Adhikari, R.; et al. SARS-CoV-2 RBD219-N1C1: A yeast-expressed SARS-CoV-2 recombinant receptor-binding domain candidate vaccine stimulates virus neutralizing antibodies and T-cell immunity in mice. Hum. Vaccines Immunother. 2021, 17, 2356–2366. [Google Scholar] [CrossRef]
- Ryzhikov, A.B.; Ryzhikov, Е.А.; Bogryantseva, M.P.; Usova, S.V.; Danilenko, E.D.; Nechaeva, E.A.; Pyankov, O.V.; Pyankova, O.G.; Gudymo, A.S.; Bodnev, S.A.; et al. A single blind, placebo-controlled randomized study of the safety, reactogenicity and immunogenicity of the “EpiVacCorona” Vaccine for the prevention of COVID-19, in volunteers aged 18–60 years (phase I–II). J. Infect. Immun. 2021, 11, 283–296. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Madan, R.; Singh, S. Immunology to Immunotherapeutics of SARS-CoV-2: Identification of Immunogenic Epitopes for Vaccine Development. Curr. Microbiol. 2022, 79, 306. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.X.; Lim, J.; Jazayeri, S.D.; Poppema, S.; Poh, C.L. Development of multi-epitope peptide-based vaccines against SARS-CoV-2. Biomed. J. 2021, 44, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Weltzin, R.; Guy, B.; Thomas, W.D., Jr.; Giannasca, P.J.; Monath, T.P. Parenteral adjuvant activities of Escherichia coli heat-labile toxin and its B subunit for immunization of mice against gastric Helicobacter pylori infection. Infect. Immun. 2000, 68, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Schödel, F.; Will, H.; Johansson, S.; Sanchez, J.; Holmgren, J. Synthesis in Vibrio cholerae and secretion of hepatitis B virus antigens fused to Escherichia coli heat-labile enterotoxin subunit B. Gene 1991, 99, 255–259. [Google Scholar] [CrossRef] [PubMed]
- IEBD. Immune Epitope Database & Tools (IEBD). Available online: https://www.iedb.org/ (accessed on 15 July 2023).
- Seow, J.; Khan, H.; Rosa, A.; Calvaresi, V.; Graham, C.; Pickering, S.; Pye, V.E.; Cronin, N.B.; Huettner, I.; Malim, M.H.; et al. A neutralizing epitope on the SD1 domain of SARS-CoV-2 spike targeted following infection and vaccination. Cell Rep. 2022, 40, 111276. [Google Scholar] [CrossRef] [PubMed]
- Robson, B. COVID-19 Coronavirus spike protein analysis for synthetic vaccines, a peptidomimetic antagonist, and therapeutic drugs, and analysis of a proposed achilles’ heel conserved region to minimize probability of escape mutations and drug resistance. Comput. Biol. Med. 2020, 121, 103749. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, F.; Crivelli, V.; Abernathy, M.E.; Guerra, C.; Palus, M.; Muri, J.; Marcotte, H.; Piralla, A.; Pedotti, M.; De Gasparo, R.; et al. Human neutralizing antibodies to cold linear epitopes and to subdomain 1 of SARS-CoV-2. bioRxiv 2022. [Google Scholar] [CrossRef]
- Li, Y.; Ma, M.L.; Lei, Q.; Wang, F.; Hong, W.; Lai, D.Y.; Hou, H.; Xu, Z.W.; Zhang, B.; Chen, H.; et al. Linear epitope landscape of the SARS-CoV-2 Spike protein constructed from 1051 COVID-19 patients. Cell Rep. 2021, 34, 108915. [Google Scholar] [CrossRef]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.; Chee, R.S.; Fong, S.W.; Yeo, N.K.; Lee, W.H.; Torres-Ruesta, A.; et al. Two linear epitopes on the SARS-CoV-2 spike protein that elicit neutralising antibodies in COVID-19 patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.A.; Singh, M. Acceptable levels of endotoxin in vaccine formulations during preclinical research. J. Pharm. Sci. 2011, 100, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tian, X.; Jia, X.; Wan, J.; Lu, L.; Jiang, S.; Lan, F.; Lu, Y.; Wu, Y.; Ying, T. The impact of receptor-binding domain natural mutations on antibody recognition of SARS-CoV-2. Signal Transduct. Target. Ther. 2021, 6, 132. [Google Scholar] [CrossRef] [PubMed]
- Kremer, E.J. Pros and Cons of Adenovirus-Based SARS-CoV-2 Vaccines. Mol. Ther. 2020, 28, 2303–2304. [Google Scholar] [CrossRef] [PubMed]
- Vitiello, A.; Ferrara, F. Brief review of the mRNA vaccines COVID-19. Inflammopharmacology 2021, 29, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, A.; Leach, S.; Tobias, J.; Carlin, N.; Gustafsson, B.; Jertborn, M.; Bourgeois, L.; Walker, R.; Holmgren, J.; Svennerholm, A.M. Clinical trial to evaluate safety and immunogenicity of an oral inactivated enterotoxigenic Escherichia coli prototype vaccine containing CFA/I overexpressing bacteria and recombinantly produced LTB/CTB hybrid protein. Vaccine 2013, 31, 1163–1170. [Google Scholar] [CrossRef]
- Balasubramaniyam, A.; Ryan, E.; Brown, D.; Hamza, T.; Harrison, W.; Gan, M.; Sankhala, R.S.; Chen, W.-H.; Martinez, E.J.; Jensen, J.L.; et al. Unglycosylated Soluble SARS-CoV-2 Receptor Binding Domain (RBD) Produced in E. coli Combined with the Army Liposomal Formulation Containing QS21 (ALFQ) Elicits Neutralizing Antibodies against Mismatched Variants. Vaccines 2023, 11, 42. [Google Scholar] [CrossRef]
- Khow, O.; Suntrarachun, S. Strategies for production of active eukaryotic proteins in bacterial expression system. Asian Pac. J. Trop. Biomed. 2012, 2, 159–162. [Google Scholar] [CrossRef]
- Pidiyar, V.; Kumraj, G.; Ahmed, K.; Ahmed, S.; Shah, S.; Majumder, P.; Verma, B.; Pathak, S.; Mukherjee, S. COVID-19 management landscape: A need for an affordable platform to manufacture safe and efficacious biotherapeutics and prophylactics for the developing countries. Vaccine 2022, 40, 5302–5312. [Google Scholar] [CrossRef]
- Knapp, M.A.; Johnson, T.A.; Ritter, M.K.; Rainer, R.O.; Fiester, S.E.; Grier, J.T.; Connell, T.D.; Arce, S. Immunomodulatory regulation by heat-labile enterotoxins and potential therapeutic applications. Expert. Rev. Vaccines 2021, 20, 975–987. [Google Scholar] [CrossRef]
- Ma, Y. Recent advances in nontoxic Escherichia coli heat-labile toxin and its derivative adjuvants. Expert. Rev. Vaccines 2016, 15, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- HogenEsch, H. Mechanisms of stimulation of the immune response by aluminum adjuvants. Vaccine 2002, 20, S34–S39. [Google Scholar] [CrossRef] [PubMed]
- Tarigan, S.; Dharmayanti, N.L.P.I.; Sugiartanti, D.; Putri, R.; Andriani Nuradji, H.; Robinson, M.; Wiendayanthi, N.; Djufri, F. Characterization of two linear epitopes SARS CoV-2 spike protein formulated in tandem repeat. PLoS ONE 2023, 18, e0280627. [Google Scholar] [CrossRef] [PubMed]
- Abebe, E.C.; Dejenie, T.A. Protective roles and protective mechanisms of neutralizing antibodies against SARS-CoV-2 infection and their potential clinical implications. Front. Immunol. 2023, 14, 1055457. [Google Scholar] [CrossRef] [PubMed]
- Ubol, S.; Halstead, S.B. How innate immune mechanisms contribute to antibody-enhanced viral infections. Clin. Vaccine Immunol. 2010, 17, 1829–1835. [Google Scholar] [CrossRef] [PubMed]
- Gartlan, C.; Tipton, T.; Salguero, F.J.; Sattentau, Q.; Gorringe, A.; Carroll, M.W. Vaccine-Associated Enhanced Disease and Pathogenic Human Coronaviruses. Front. Immunol. 2022, 13, 882972. [Google Scholar] [CrossRef]
- King, R.G.; Silva-Sanchez, A.; Peel, J.N.; Botta, D.; Dickson, A.M.; Pinto, A.K.; Meza-Perez, S.; Allie, S.R.; Schultz, M.D.; Liu, M.; et al. Single-Dose Intranasal Administration of AdCOVID Elicits Systemic and Mucosal Immunity against SARS-CoV-2 and Fully Protects Mice from Lethal Challenge. Vaccines 2021, 9, 881. [Google Scholar] [CrossRef]
- Bewley, K.R.; Gooch, K.; Thomas, K.M.; Longet, S.; Wiblin, N.; Hunter, L.; Chan, K.; Brown, P.; Russell, R.A.; Ho, C.; et al. Immunological and pathological outcomes of SARS-CoV-2 challenge following formalin-inactivated vaccine in ferrets and rhesus macaques. Sci. Adv. 2021, 7, eabg7996. [Google Scholar] [CrossRef]
- Snider, D.P.; Marshall, J.S.; Perdue, M.H.; Liang, H. Production of IgE antibody and allergic sensitization of intestinal and peripheral tissues after oral immunization with protein Ag and cholera toxin. J. Immunol. 1994, 153, 647–657. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to Image J: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Solis-Andrade, K.I.; Gonzalez-Ortega, O.; Govea-Alonso, D.O.; Comas-Garcia, M.; Rosales-Mendoza, S. Production and Purification of LTB-RBD: A Potential Antigen for Mucosal Vaccine Development against SARS-CoV-2. Vaccines 2022, 10, 1759. [Google Scholar] [CrossRef]
- Farfán-Castro, S.; García-Soto, M.J.; Comas-García, M.; Arévalo-Villalobos, J.I.; Palestino, G.; González-Ortega, O.; Rosales-Mendoza, S. Synthesis and immunogenicity assessment of a gold nanoparticle conjugate for the delivery of a peptide from SARS-CoV-2. Nanomed. Nanotechnol. Biol. Med. 2021, 34, 102372. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scales | LTp50 in Crude Extract (mg/L Culture) | Pure LT-p50 (mg/L Culture) | Yield * | Purity |
|---|---|---|---|---|
| Flask (1 L) | 33.7 mg/L | 16.7 mg/L | 25.6% | >90% |
| Bioreactor (1.5 L) | 52.6 mg/L | 22.5 mg/L | 42.8% | >90% |
| Stage | Buffer | Rounds | Incubation time |
|---|---|---|---|
| Washing | PBS 1×, 1% (v/v) Triton X-100 | 2 | 20 min |
| Washing | PBS 1× | 2 | 20 min |
| Washing | 20 mM phosphate buffer, 500 mM NaCl, 4 M urea, pH 7.4 | 1 | 20 min |
| Solubilization | 20 mM phosphate buffer, 500 mM NaCl, 8 M urea, pH 7.4 | 1 | 20 min |
| Solubilization | 20 mM phosphate buffer, 500 mM NaCl, 8 M urea, pH 7.4 | 1 | overnight |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong-Arce, A.; Gonzalez-Ortega, O.; Romero-Maldonado, A.; Miranda-López, A.; García-Soto, M.; Farfán-Castro, S.; Betancourt-Mendiola, L.; Teeravechyan, S.; Srisutthisamphan, K.; Comas-García, M.; et al. Production and Immunogenicity Assessment of LTp50: An Escherichia coli-Made Chimeric Antigen Targeting S1- and S2-Epitopes from the SARS-CoV-2/BA.5 Spike Protein. Pharmaceuticals 2024, 17, 302. https://doi.org/10.3390/ph17030302
Wong-Arce A, Gonzalez-Ortega O, Romero-Maldonado A, Miranda-López A, García-Soto M, Farfán-Castro S, Betancourt-Mendiola L, Teeravechyan S, Srisutthisamphan K, Comas-García M, et al. Production and Immunogenicity Assessment of LTp50: An Escherichia coli-Made Chimeric Antigen Targeting S1- and S2-Epitopes from the SARS-CoV-2/BA.5 Spike Protein. Pharmaceuticals. 2024; 17(3):302. https://doi.org/10.3390/ph17030302
Chicago/Turabian StyleWong-Arce, Alejandra, Omar Gonzalez-Ortega, Andrea Romero-Maldonado, Arleth Miranda-López, Mariano García-Soto, Susan Farfán-Castro, Lourdes Betancourt-Mendiola, Samaporn Teeravechyan, Kanjana Srisutthisamphan, Mauricio Comas-García, and et al. 2024. "Production and Immunogenicity Assessment of LTp50: An Escherichia coli-Made Chimeric Antigen Targeting S1- and S2-Epitopes from the SARS-CoV-2/BA.5 Spike Protein" Pharmaceuticals 17, no. 3: 302. https://doi.org/10.3390/ph17030302
APA StyleWong-Arce, A., Gonzalez-Ortega, O., Romero-Maldonado, A., Miranda-López, A., García-Soto, M., Farfán-Castro, S., Betancourt-Mendiola, L., Teeravechyan, S., Srisutthisamphan, K., Comas-García, M., Solís Andrade, K. I., & Rosales-Mendoza, S. (2024). Production and Immunogenicity Assessment of LTp50: An Escherichia coli-Made Chimeric Antigen Targeting S1- and S2-Epitopes from the SARS-CoV-2/BA.5 Spike Protein. Pharmaceuticals, 17(3), 302. https://doi.org/10.3390/ph17030302