
GDF3 Protects Mice against Sepsis-Induced Acute Lung Injury by Suppression of Macrophage Pyroptosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
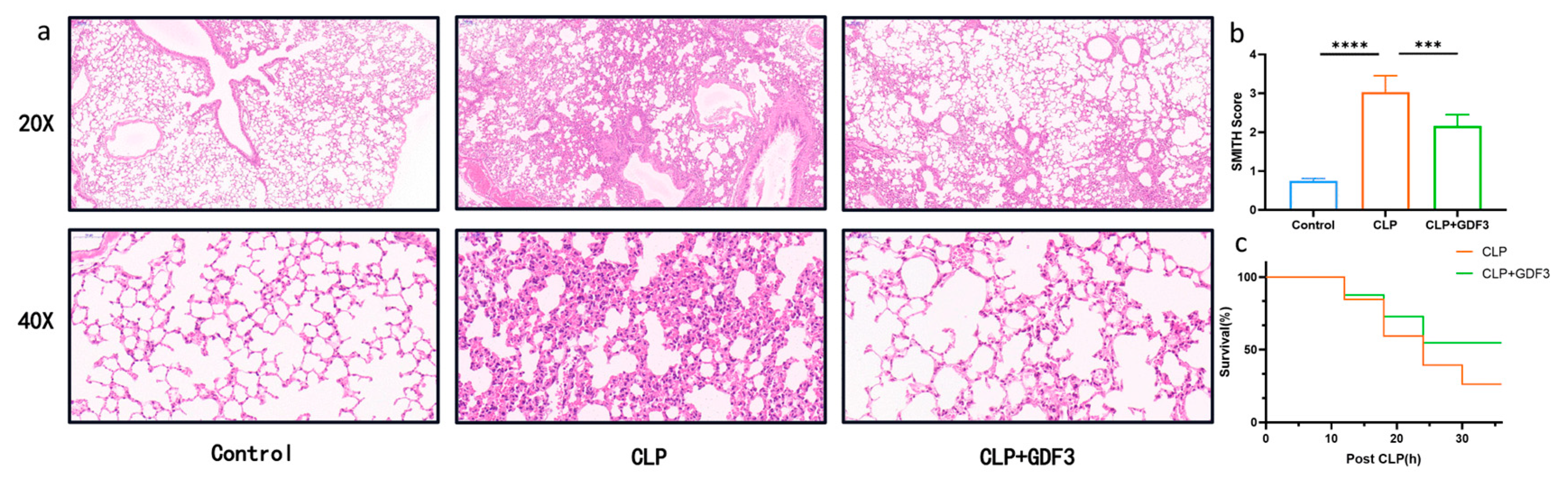
2.1. GDF3 Protects Mice against CLP-Induced ALI
2.2. GDF3 Alleviated the Increased Alveolar–Capillary Permeability in Lung Tissue
2.3. GDF3 Inhibits the Levels of Cytokines in CLP-Induced ALI
2.4. GDF3 Reduces Pyroptosis in CLP-Induced ALI
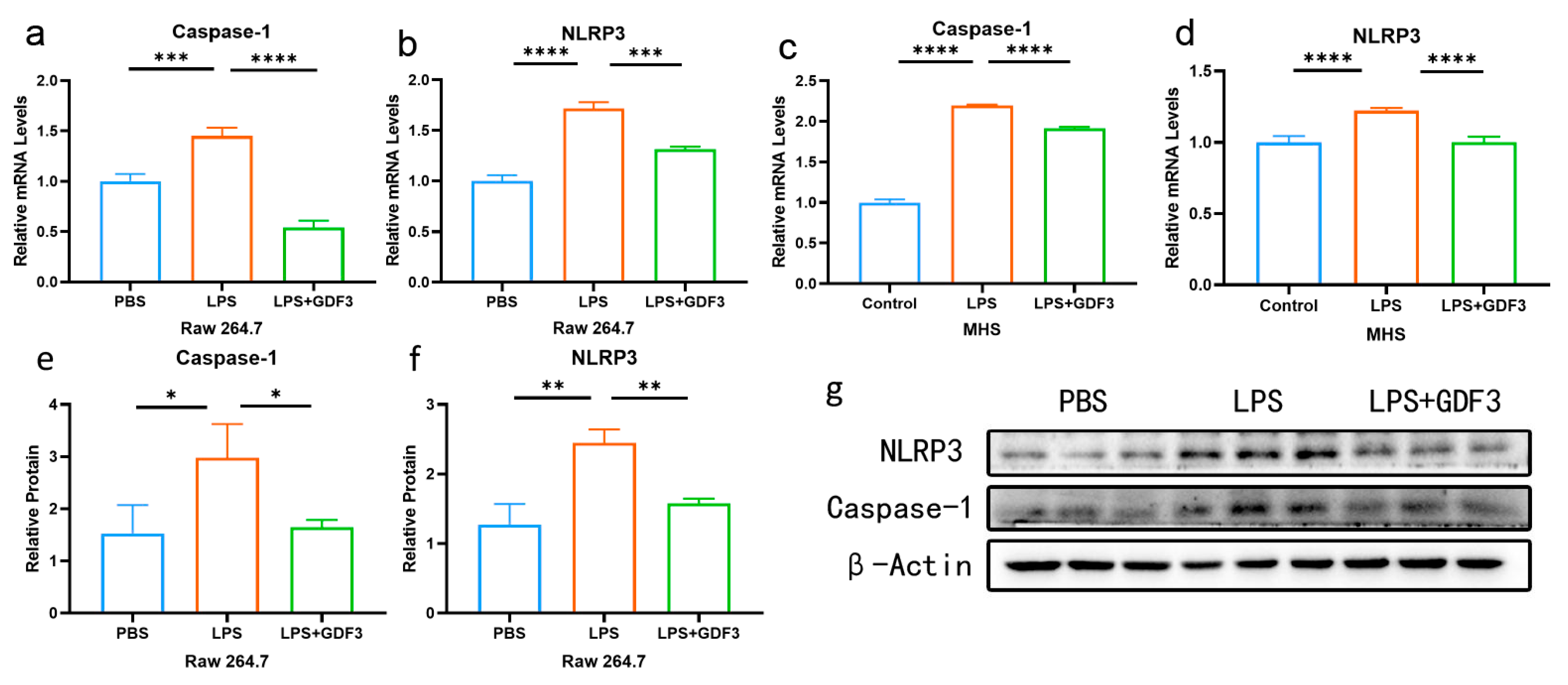
2.5. GDF3 Inhibited LPS-Induced Pyroptosis in Macrophages
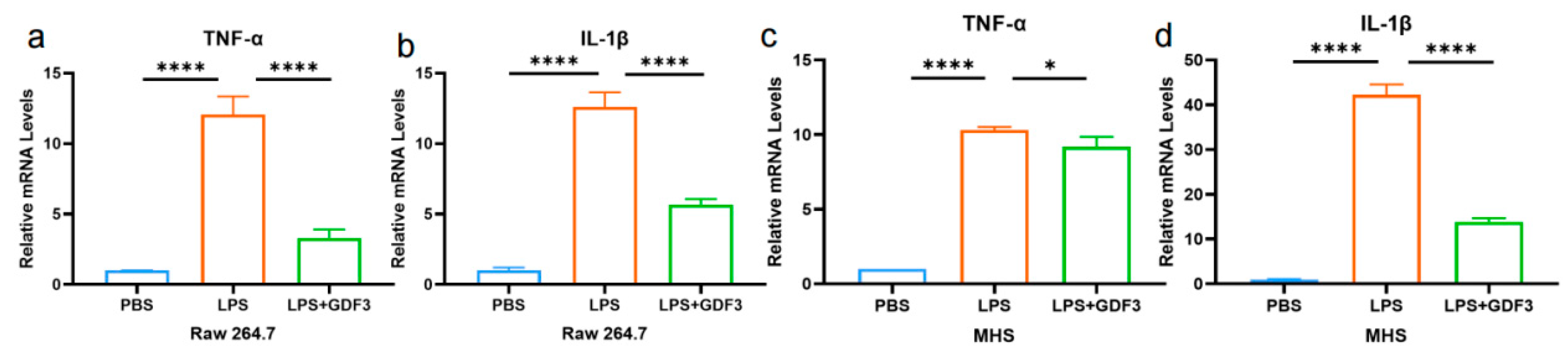
2.6. GDF3 Reduced the Levels of Cytokines in LPS-Induced Macrophages
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cell Culture
4.3. Mouse Model Preparation and Treatment
4.4. Cell Count and Classification of Bronchoalveolar Lavage Fluid (BALF)
4.5. Lung Histology and Lung Injury Score
4.6. Tissue Wet-to-Dry Weight Ratio
4.7. Wright’s Giemsa Staining
4.8. Cytokines of the Serum and the BALF
4.9. Immunohistochemistry
4.10. Immunofluorescence Staining
4.11. Isolation of RNA and Quantitative Real-Time PCR (qRT-PCR)
4.12. Western Blot
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Torres, L.K.; Pickkers, P.; van der Poll, T. Sepsis-Induced Immunosuppression. Annu. Rev. Physiol. 2022, 84, 157–181. [Google Scholar] [CrossRef]
- He, R.; Liu, B.; Xiong, R.; Geng, B.; Meng, H.; Lin, W.; Hao, B.; Zhang, L.; Wang, W.; Jiang, W.; et al. Itaconate inhibits ferroptosis of macrophage via Nrf2 pathways against sepsis-induced acute lung injury. Cell Death Discov. 2022, 8, 43. [Google Scholar] [CrossRef]
- Ning, L.; Shishi, Z.; Bo, W.; Huiqing, L. Targeting immunometabolism against acute lung injury. Clin. Immunol. 2023, 249, 109289. [Google Scholar] [CrossRef]
- Vichare, R.; Janjic, J.M. Macrophage-Targeted Nanomedicines for ARDS/ALI: Promise and Potential. Inflammation 2022, 45, 2124–2141. [Google Scholar] [CrossRef]
- Gao, Y.-L.; Zhai, J.-H.; Chai, Y.-F. Recent Advances in the Molecular Mechanisms Underlying Pyroptosis in Sepsis. Mediat. Inflamm. 2018, 2018, 1–7. [Google Scholar] [CrossRef]
- Sun, J.; Ge, X.; Wang, Y.; Niu, L.; Tang, L.; Pan, S. USF2 knockdown downregulates THBS1 to inhibit the TGF-β signaling pathway and reduce pyroptosis in sepsis-induced acute kidney injury. Pharmacol. Res. 2022, 176, 105962. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, W.; Gong, F.; Chen, Y.; Chen, E. The Role and Mechanism of Pyroptosis and Potential Therapeutic Targets in Sepsis: A Review. Front. Immunol. 2021, 12, 711939. [Google Scholar] [CrossRef]
- Rimmelé, T.; Payen, D.; Cantaluppi, V.; Marshall, J.; Gomez, H.; Gomez, A.; Murray, P.; Kellum, J.A.O.B.O.T.A.X.W. Immune Cell Phenotype and Function in Sepsis. Shock 2016, 45, 282–291. [Google Scholar] [CrossRef]
- Liu, B.; Wang, Z.; He, R.; Xiong, R.; Li, G.; Zhang, L.; Fu, T.; Li, C.; Li, N.; Geng, Q. Buformin alleviates sepsis-induced acute lung injury via inhibiting NLRP3-mediated pyroptosis through an AMPK-dependent pathway. Clin. Sci. 2022, 136, 273–289. [Google Scholar] [CrossRef]
- Zamani, N.; Brown, C.W. Emerging roles for the transforming growth factor-β superfamily in regulating adiposity and energy expenditure. Endocr. Rev. 2011, 32, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Camell, C.D.; Sander, J.; Spadaro, O.; Lee, A.; Nguyen, K.Y.; Wing, A.; Goldberg, E.L.; Youm, Y.-H.; Brown, C.W.; Elsworth, J.; et al. Inflammasome-driven catecholamine catabolism in macrophages blunts lipolysis during ageing. Nature 2017, 550, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Patsalos, A.; Simandi, Z.; Hays, T.T.; Peloquin, M.; Hajian, M.; Restrepo, I.; Coen, P.M.; Russell, A.J.; Nagy, L. In vivo GDF3 administration abrogates aging related muscle regeneration delay following acute sterile injury. Aging Cell 2018, 17, e12815. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Mu, X.; Zhao, H.; Li, Y.; Wang, L.; Wolfe, V.; Cui, S.-N.; Wang, X.; Peng, T.; Zingarelli, B.; et al. Administration of GDF3 Into Septic Mice Improves Survival via Enhancing LXRα-Mediated Macrophage Phagocytosis. Front. Immunol. 2021, 12, 647070. [Google Scholar] [CrossRef]
- Levitt, J.E.; Matthay, M.A. Clinical review: Early treatment of acute lung injury-paradigm shift toward prevention and treatment prior to respiratory failure. Crit. Care 2012, 16, 223. [Google Scholar] [CrossRef]
- Herold, S.; Gabrielli, N.M.; Vadász, I. Novel concepts of acute lung injury and alveolar-capillary barrier dysfunction. Am. J. Physiol. Cell. Mol. Physiol. 2013, 305, L665–L681. [Google Scholar] [CrossRef]
- Sun, B.; Lei, M.; Zhang, J.; Kang, H.; Liu, H.; Zhou, F. Acute lung injury caused by sepsis: How does it happen? Front. Med. 2023, 10, 1289194. [Google Scholar] [CrossRef]
- Zhuang, C.; Kang, M.; Lee, M. Delivery systems of therapeutic nucleic acids for the treatment of acute lung injury/acute respiratory distress syndrome. J. Control. Release 2023, 360, 1–14. [Google Scholar] [CrossRef]
- An, Y.; Zhang, H.; Wang, R.; Shen, Y.-F.; Zhao, J.-L.; Zou, Y.-S.; Wang, G.-Z. Biomarkers, signaling pathways, and programmed cell death in acute lung injury and its treatment with Traditional Chinese Medicine: A narrative review. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 10157–10170. [Google Scholar] [CrossRef]
- Xu, H.; Sheng, S.; Luo, W.; Xu, X.; Zhang, Z. Acute respiratory distress syndrome heterogeneity and the septic ARDS subgroup. Front. Immunol. 2023, 14, 1277161. [Google Scholar] [CrossRef]
- Levine, A.J.; Brivanlou, A.H. GDF3 at the crossroads of TGF-beta signaling. Cell Cycle 2006, 5, 1069–1073. [Google Scholar] [CrossRef]
- Finicelli, M.; Benedetti, G.; Squillaro, T.; Pistilli, B.; Marcellusi, A.; Mariani, P.; Santinelli, A.; Latini, L.; Galderisi, U.; Giordano, A. Expression of stemness genes in primary breast cancer tissues: The role of SOX2 as a prognostic marker for detection of early recurrence. Oncotarget 2014, 5, 9678–9688. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Kang, G.; Wang, J.; Li, T.; Chen, J.; Wang, J.; Li, W.; Wang, B. A novel variation of GDF3 in Chinese Han children with a broad phenotypic spectrum of non-syndromic CHDs. Cardiol. Young 2014, 25, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Varga, T.; Mounier, R.; Patsalos, A.; Gogolák, P.; Peloquin, M.; Horvath, A.; Pap, A.; Daniel, B.; Nagy, G.; Pintye, E.; et al. Macrophage PPARγ, a Lipid Activated Transcription Factor Controls the Growth Factor GDF3 and Skeletal Muscle Regeneration. Immunity 2016, 45, 1038–1051. [Google Scholar] [CrossRef] [PubMed]
- Bu, Y.; Okunishi, K.; Yogosawa, S.; Mizuno, K.; Irudayam, M.J.; Brown, C.W.; Izumi, T. Insulin Regulates Lipolysis and Fat Mass by Upregulating Growth/Differentiation Factor 3 in Adipose Tissue Macrophages. Diabetes 2018, 67, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Dong, X.; Li, G.; Chen, Y.; Chen, J.; He, X.; Sun, H.; Kim, D.-H.; Kemper, J.K.; Chen, L.-F. Brd4 modulates diet-induced obesity via PPARγ-dependent Gdf3 expression in adipose tissue macrophages. J. Clin. Investig. 2021, 6, 143379. [Google Scholar] [CrossRef]
- Wang, L.; Li, Y.; Wang, X.; Wang, P.; Essandoh, K.; Cui, S.; Huang, W.; Mu, X.; Liu, Z.; Wang, Y.; et al. GDF3 Protects Mice against Sepsis-Induced Cardiac Dysfunction and Mortality by Suppression of Macrophage Pro-Inflammatory Phenotype. Cells 2020, 9, 120. [Google Scholar] [CrossRef]
- Wang, A.; Wang, Y.; Du, C.; Yang, H.; Wang, Z.; Jin, C.; Hamblin, M.R. Pyroptosis and the tumor immune microenvironment: A new battlefield in ovarian cancer treatment. Biochim. et Biophys. Acta (BBA)-Rev. Cancer 2023, 1879, 189058. [Google Scholar] [CrossRef]
- Liu, Y.; Lei, H.; Zhang, W.; Xing, Q.; Liu, R.; Wu, S.; Liu, Z.; Yan, Q.; Li, W.; Liu, X.; et al. Pyroptosis in renal inflammation and fibrosis: Current knowledge and clinical significance. Cell Death Dis. 2023, 14, 472. [Google Scholar] [CrossRef]
- You, H.-M.; Wang, L.; Meng, H.-W.; Huang, C.; Fang, G.-Y.; Li, J. Pyroptosis: Shedding light on the mechanisms and links with cancers. Front. Immunol. 2023, 14, 1290885. [Google Scholar] [CrossRef]
- Jiao, C.; Zhang, H.; Li, H.; Fu, X.; Lin, Y.; Cao, C.; Liu, S.; Liu, Y.; Li, P. Caspase-3/GSDME mediated pyroptosis: A potential pathway for sepsis. Int. Immunopharmacol. 2023, 124 Pt B, 111022. [Google Scholar] [CrossRef]
- Chen, P.; Liu, J.; Sun, K.; Wang, L.; Li, X.; Li, X.; Jia, W.; Yuan, X. Methylation of TTC4 interaction with HSP70 inhibits pyroptosis in macrophages of sepsis-induced lung injury by NLRP3 inflammation. Am. J. Cancer Res. 2023, 13, 5122–5137. [Google Scholar] [PubMed]
- Tao, Y.; Xu, X.; Yang, B.; Zhao, H.; Li, Y. Mitigation of Sepsis-Induced Acute Lung Injury by BMSC-Derived Exosomal miR-125b-5p Through STAT3-Mediated Suppression of Macrophage Pyroptosis. Int. J. Nanomed. 2023, ume 18, 7095–7113. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.; Radbruch, A.; Abrignani, S.; Addo, R.; Akdis, M.; Andrä, I.; Andreata, F.; Annunziato, F.; Arranz, E.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (third edition). Eur. J. Immunol. 2021, 51, 2708–3145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lu, H.; Yu, L.; Yuan, J.; Qin, S.; Li, C.; Ge, R.-S.; Chen, H.; Ye, L. Effects of gestational exposure to perfluorooctane sulfonate on the lung development of offspring rats. Environ. Pollut. 2021, 272, 115535. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, J.; Wang, L.; Zou, L.; Wang, H.; Zhang, Y.; Liu, S.; Pan, M.; Zhu, X.; Zhan, L. GDF3 Protects Mice against Sepsis-Induced Acute Lung Injury by Suppression of Macrophage Pyroptosis. Pharmaceuticals 2024, 17, 268. https://doi.org/10.3390/ph17030268
Lei J, Wang L, Zou L, Wang H, Zhang Y, Liu S, Pan M, Zhu X, Zhan L. GDF3 Protects Mice against Sepsis-Induced Acute Lung Injury by Suppression of Macrophage Pyroptosis. Pharmaceuticals. 2024; 17(3):268. https://doi.org/10.3390/ph17030268
Chicago/Turabian StyleLei, Jiaxi, Lu Wang, Lijuan Zou, Huijuan Wang, Yunlong Zhang, Shiping Liu, Mingliang Pan, Xue Zhu, and Liying Zhan. 2024. "GDF3 Protects Mice against Sepsis-Induced Acute Lung Injury by Suppression of Macrophage Pyroptosis" Pharmaceuticals 17, no. 3: 268. https://doi.org/10.3390/ph17030268
APA StyleLei, J., Wang, L., Zou, L., Wang, H., Zhang, Y., Liu, S., Pan, M., Zhu, X., & Zhan, L. (2024). GDF3 Protects Mice against Sepsis-Induced Acute Lung Injury by Suppression of Macrophage Pyroptosis. Pharmaceuticals, 17(3), 268. https://doi.org/10.3390/ph17030268