
Sinomenium acutum Modulates Platelet Aggregation and Thrombus Formation by Regulating the Glycoprotein VI-Mediated Signalosome in Mice


Abstract
:1. Introduction
2. Results
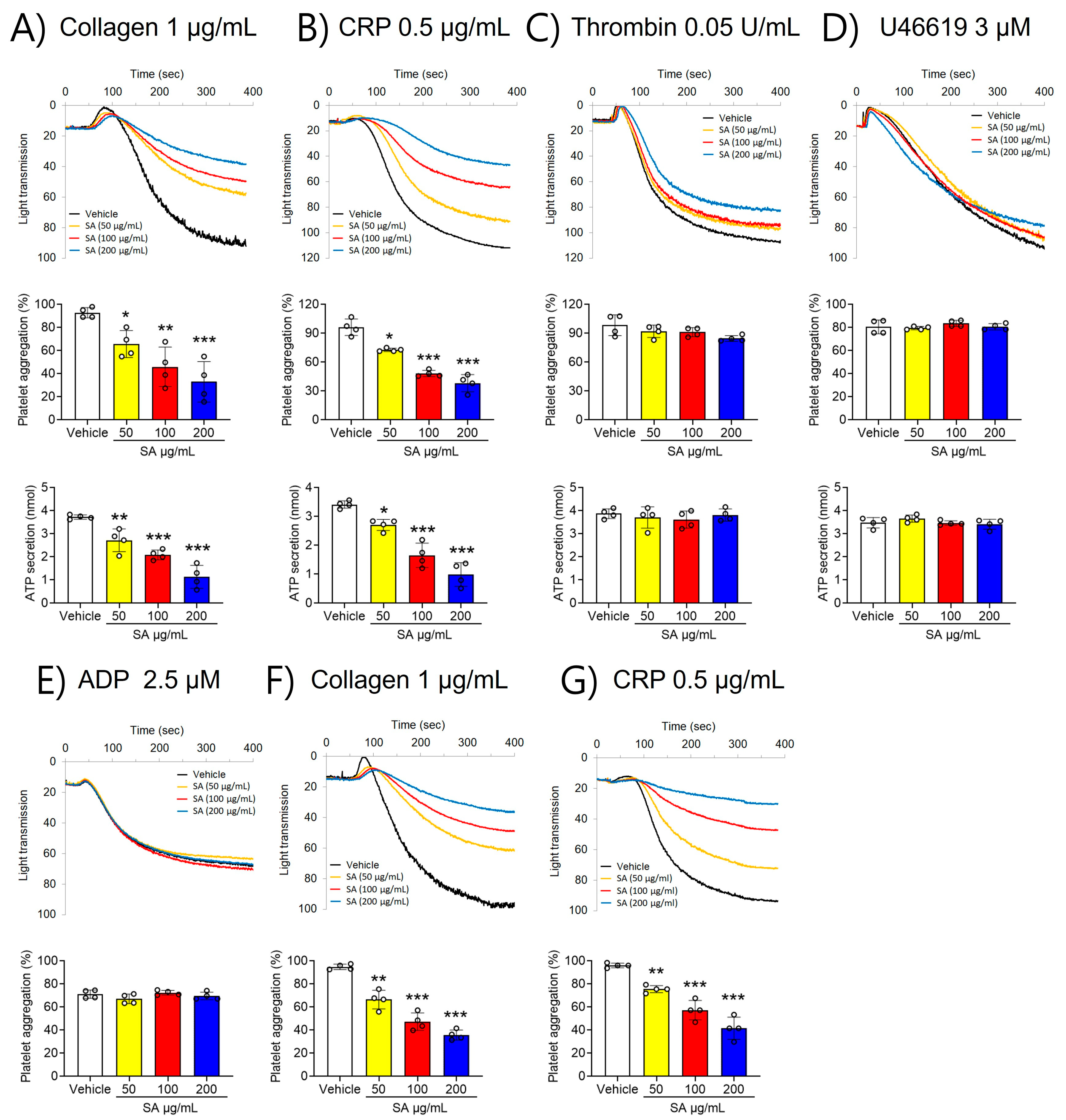
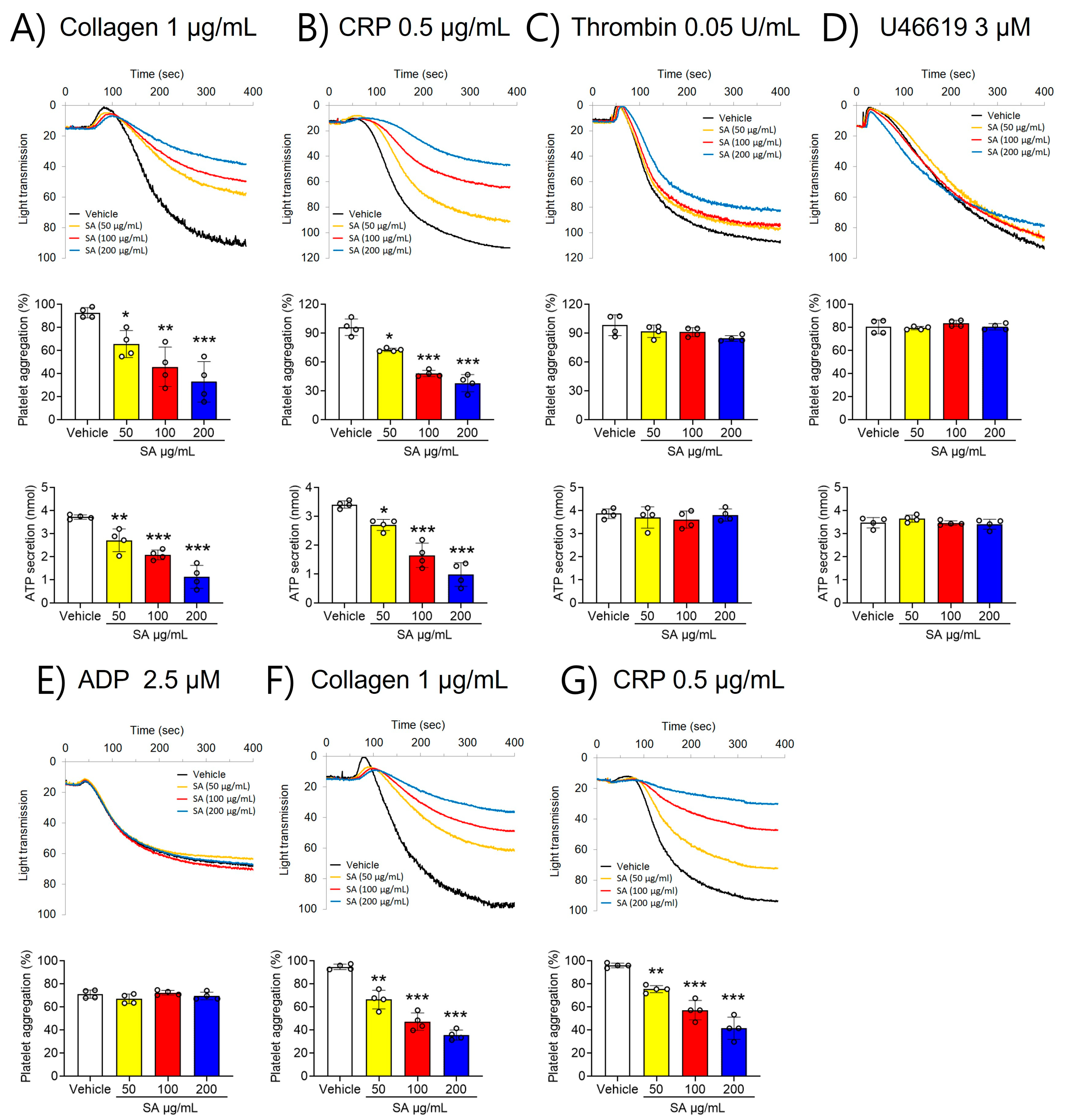
2.1. SA Suppresses GPVI-Activated Platelet Aggregation and ATP Secretion
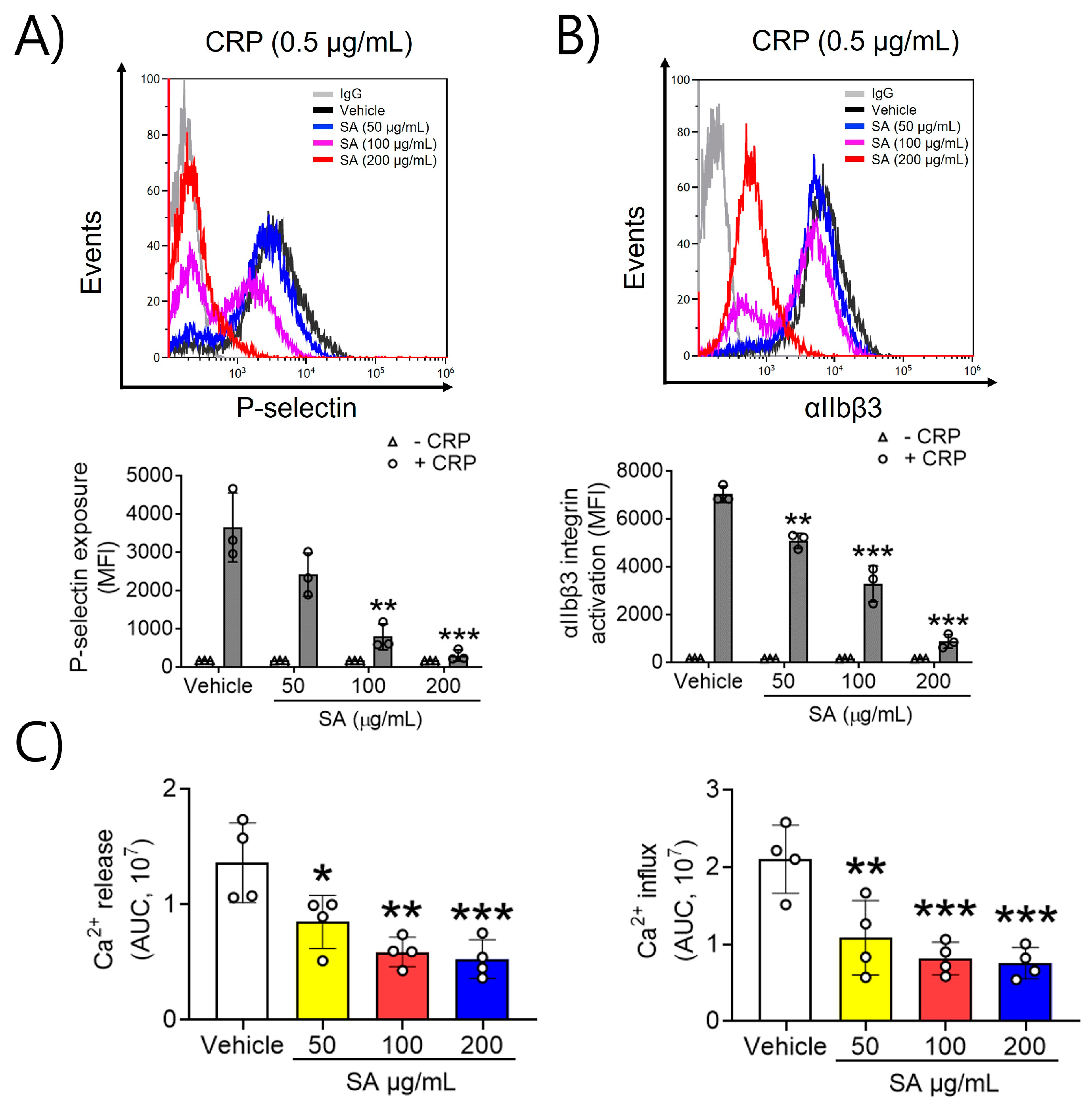
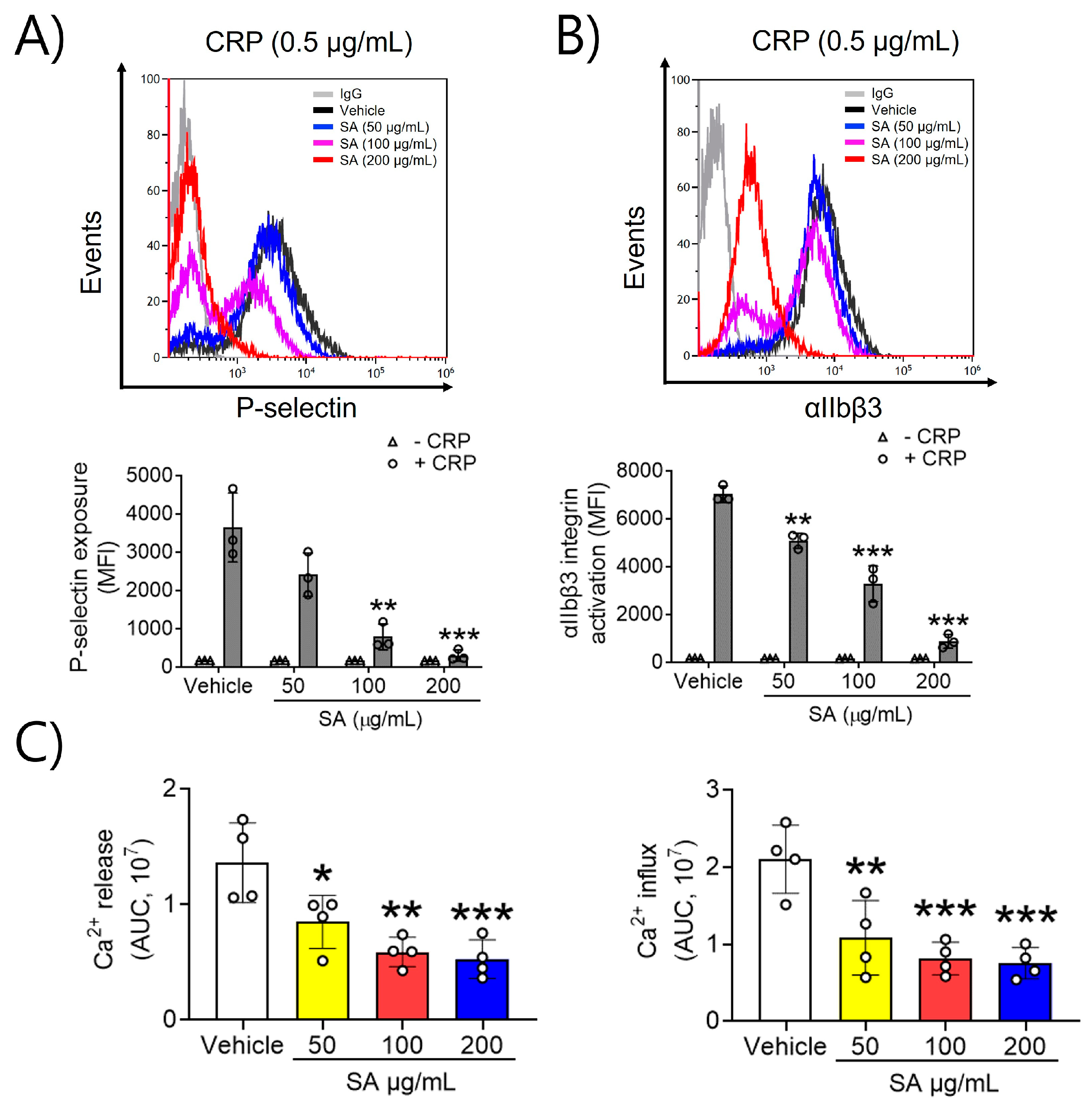
2.2. SA Is Actively Involved in P-Selectin Exposure, αIIbβ3 Integrin Activation, and Ca2+ Mobilization after CRP Induction
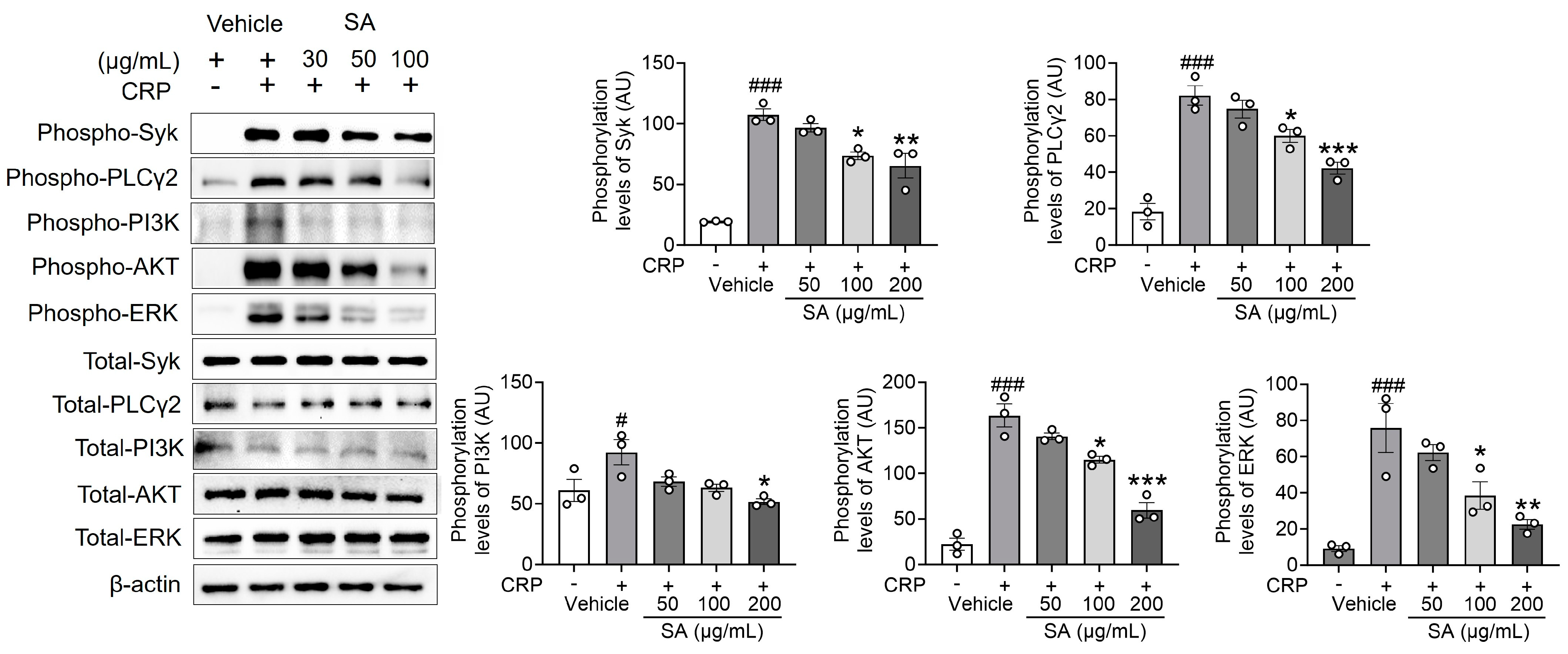
2.3. SA Is Essential in Regulating Syk, PLCγ2, PI3K, AKT, and ERK Phosphorylation Following CRP Activation
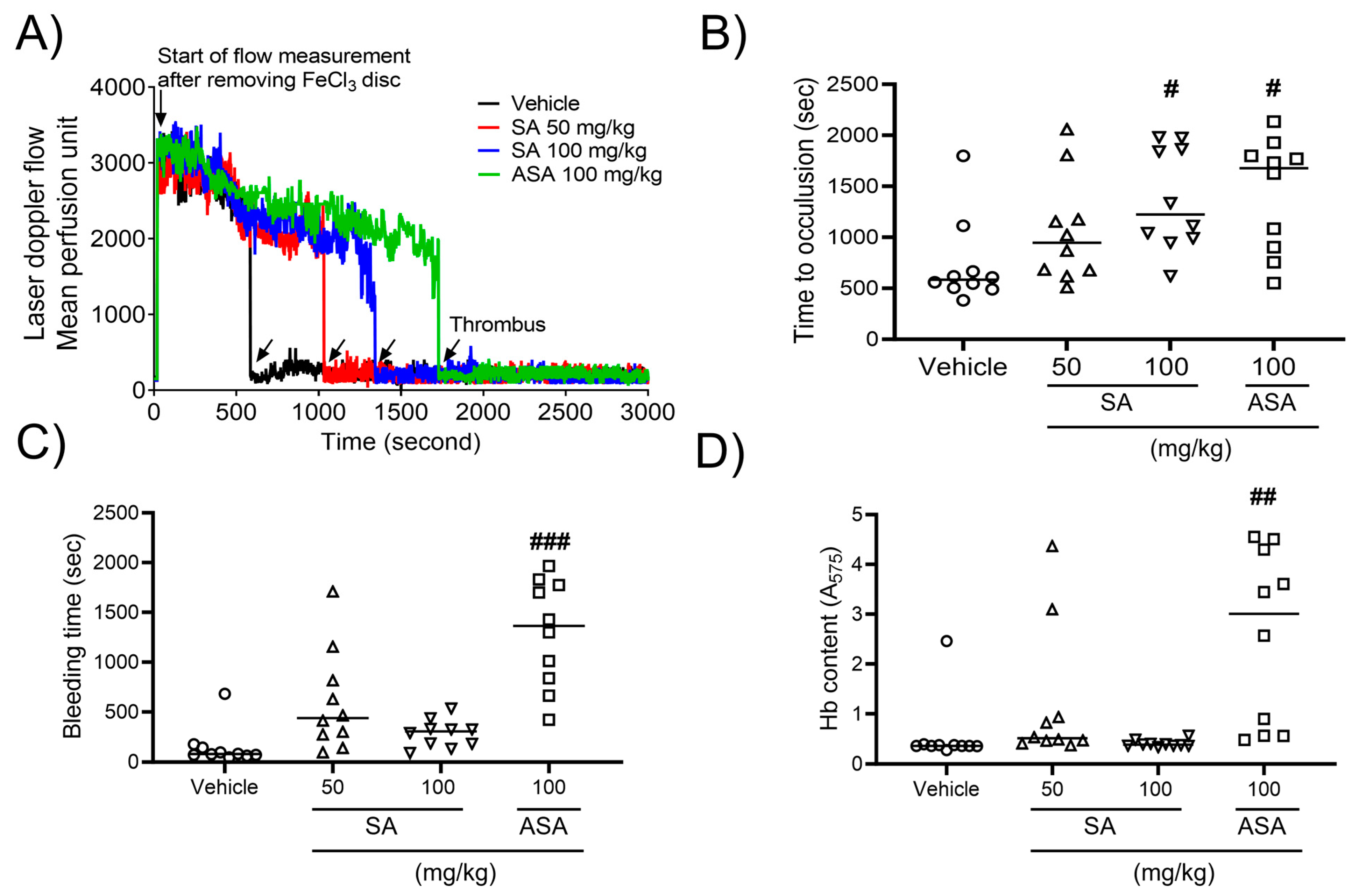
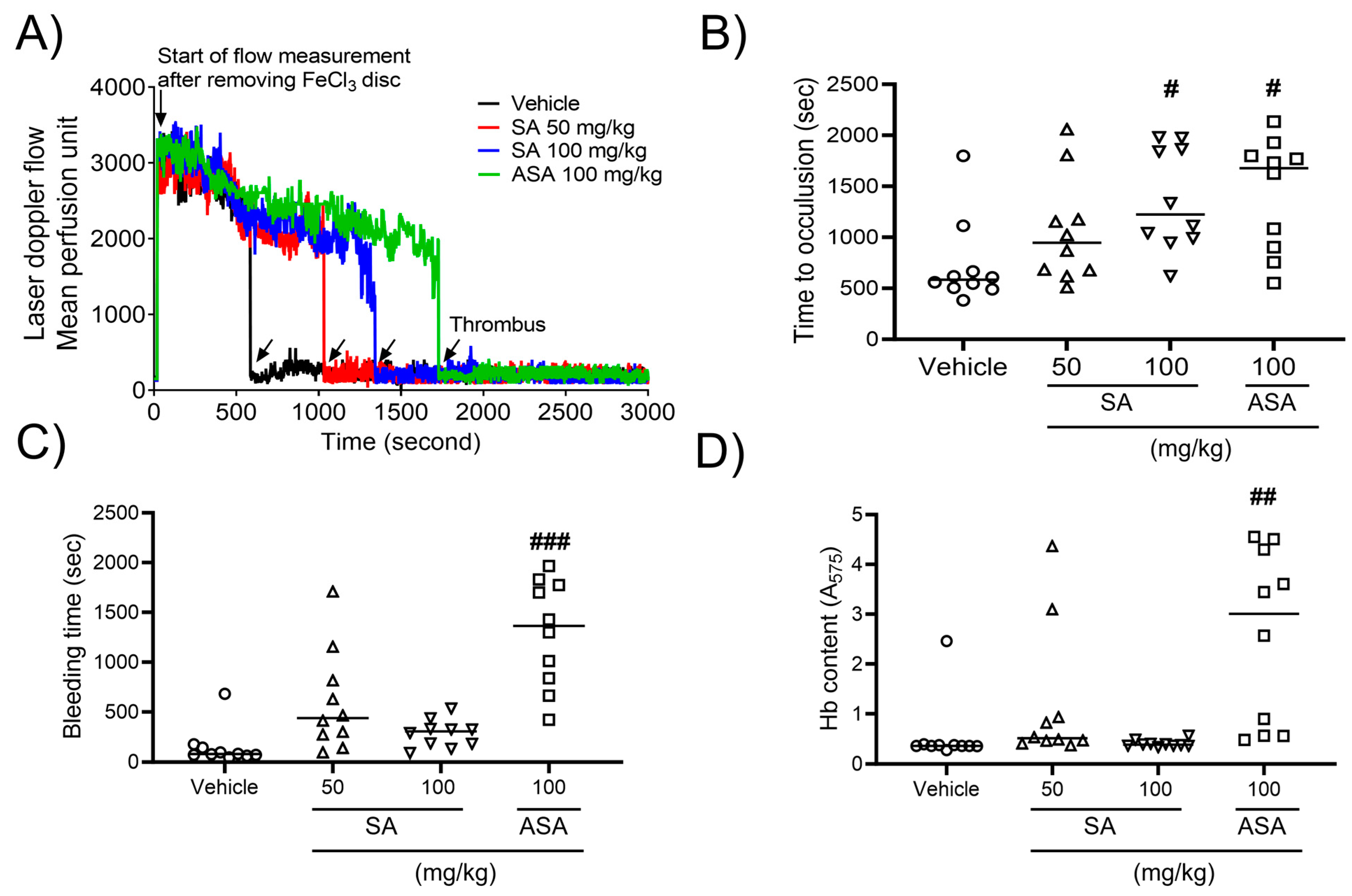
2.4. SA prevents In Vivo Thrombosis, While Having No Impact on Hemostasis
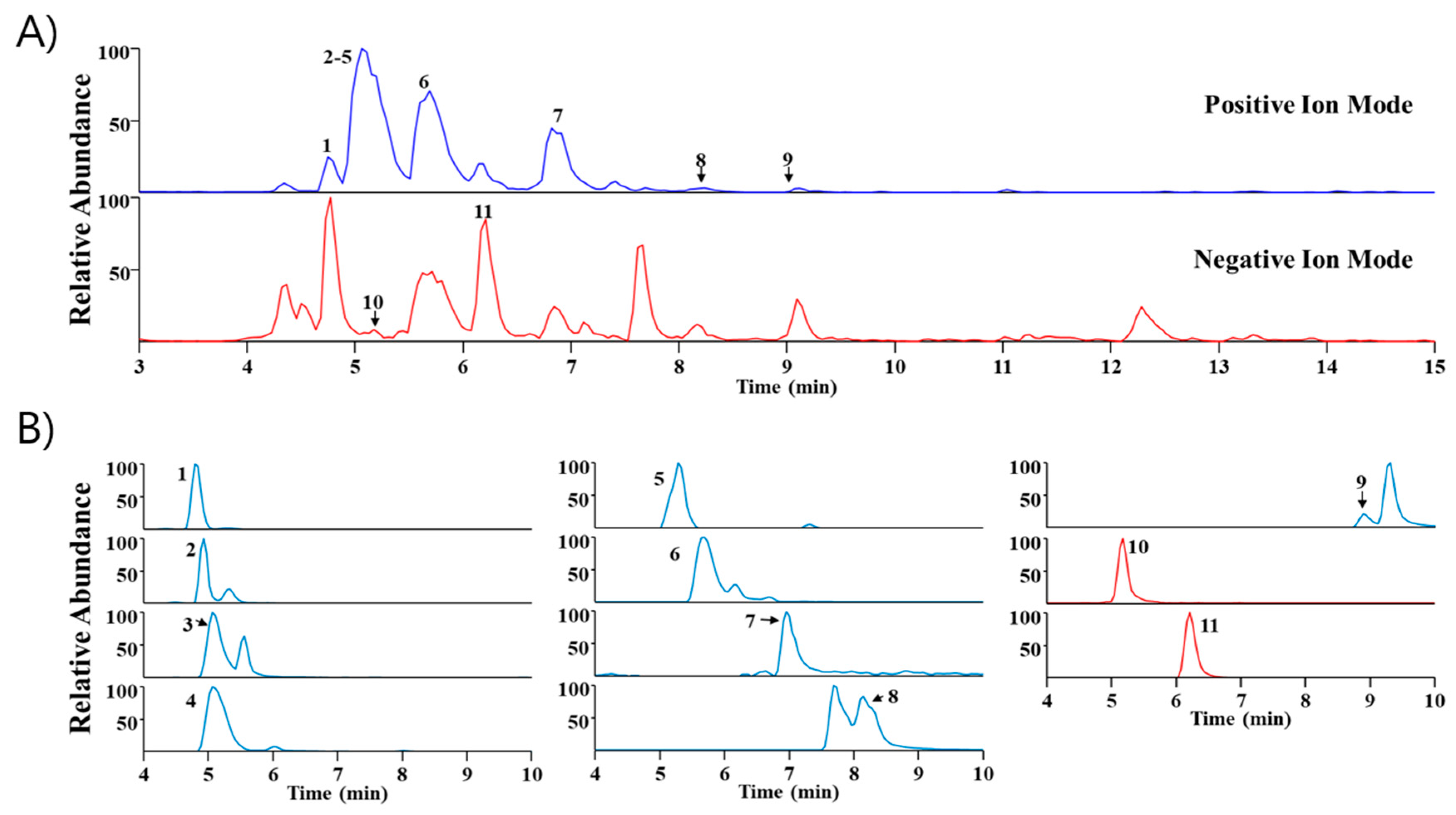
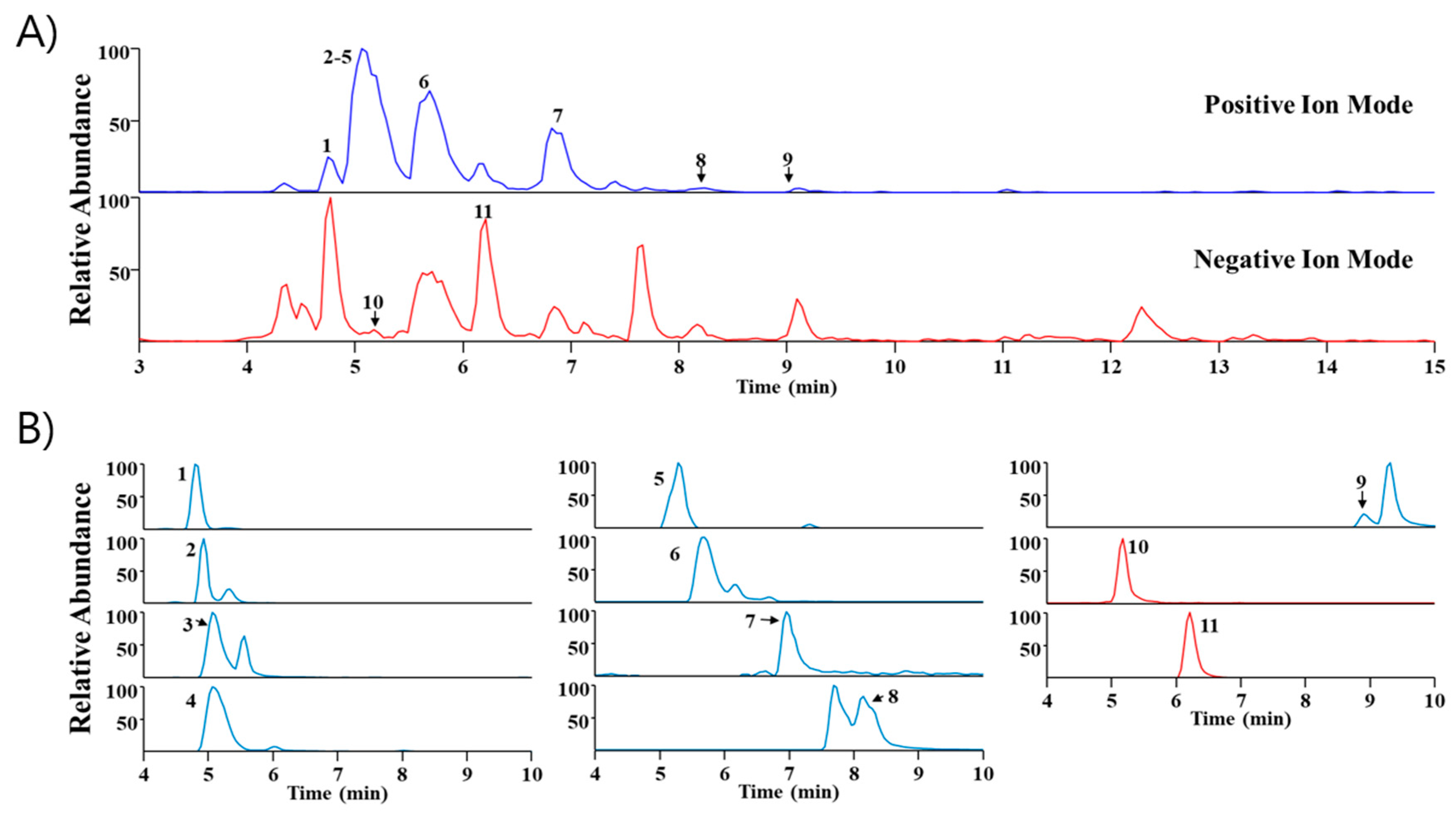
2.5. Phytochemical Profiling of SA
3. Discussion
4. Materials and Methods
4.1. Reagent
4.2. Herbal Medicine Preparation
4.3. Animals
4.4. Platelet Preparation
4.5. Platelet Aggregation and ATP Secretion
4.6. Flow Cytometric Analysis
4.7. Ca2+ Mobilization
4.8. Immunoblotting
4.9. FeCl3-Induced In Vivo Thrombosis and Tail Bleeding Time
4.10. Phytochemical Profiling of Sinomenium acutum with Ultra-High-Performance Liquid Chromatography–Tandem Mass Spectrometry (UHPLC–MS/MS)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, Y.; Zhang, D.; Tan, P.; Xian, B.; Jiang, H.; Wu, Q.; Huang, X.; Zhang, P.; Xiao, X.; Pei, J. Mechanism of platelet activation and potential therapeutic effects of natural drugs. Phytomedicine 2023, 108, 154463. [Google Scholar] [CrossRef]
- Furie, B.; Furie, B.C. Mechanisms of thrombus formation. N. Engl. J. Med. 2008, 359, 938–949. [Google Scholar] [CrossRef]
- Nieswandt, B.; Watson, S.P. Platelet-collagen interaction: Is gpvi the central receptor? Blood 2003, 102, 449–461. [Google Scholar] [CrossRef]
- Lowell, C.A. Src-family and syk kinases in activating and inhibitory pathways in innate immune cells: Signaling cross talk. Cold Spring Harb. Perspect. Biol. 2011, 3, a002352. [Google Scholar] [CrossRef]
- Watson, S.P.; Asazuma, N.; Atkinson, B.; Berlanga, O.; Best, D.; Bobe, R.; Jarvis, G.; Marshall, S.; Snell, D.; Stafford, M.; et al. The role of itam- and itim-coupled receptors in platelet activation by collagen. Thromb. Haemost. 2001, 86, 276–288. [Google Scholar]
- Dütting, S.; Bender, M.; Nieswandt, B. Platelet gpvi: A target for antithrombotic therapy?! Trends Pharmacol. Sci. 2012, 33, 583–590. [Google Scholar] [CrossRef]
- Munnix, I.C.; Strehl, A.; Kuijpers, M.J.; Auger, J.M.; van der Meijden, P.E.; van Zandvoort, M.A.; oude Egbrink, M.G.; Nieswandt, B.; Heemskerk, J.W. The glycoprotein vi-phospholipase cgamma2 signaling pathway controls thrombus formation induced by collagen and tissue factor in vitro and in vivo. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2673–2678. [Google Scholar] [CrossRef]
- Varga-Szabo, D.; Braun, A.; Nieswandt, B. Calcium signaling in platelets. J. Thromb. Haemost. 2009, 7, 1057–1066. [Google Scholar] [CrossRef]
- Nurden, A.T. Clinical significance of altered collagen-receptor functioning in platelets with emphasis on glycoprotein vi. Blood Rev. 2019, 38, 100592. [Google Scholar] [CrossRef]
- Ding, C.; Li, Y.; Sun, Y.; Wu, Y.; Wang, F.; Liu, C.; Zhang, H.; Jiang, Y.; Zhang, D.; Song, X. Sinomenium acutum: A comprehensive review of its botany, phytochemistry, pharmacology and clinical application. Am. J. Chin. Med. 2022, 50, 1219–1253. [Google Scholar] [CrossRef]
- Zhao, X.X.; Peng, C.; Zhang, H.; Qin, L.P. Sinomenium acutum: A review of chemistry, pharmacology, pharmacokinetics, and clinical use. Pharm. Biol. 2012, 50, 1053–1061. 47. [Google Scholar] [CrossRef]
- Stefanini, L.; Roden, R.C.; Bergmeier, W. Caldag-gefi is at the nexus of calcium-dependent platelet activation. Blood 2009, 114, 2506–2514. [Google Scholar] [CrossRef]
- Berlanga, O.; Bobe, R.; Becker, M.; Murphy, G.; Leduc, M.; Bon, C.; Barry, F.A.; Gibbins, J.M.; Garcia, P.; Frampton, J.; et al. Expression of the collagen receptor glycoprotein vi during megakaryocyte differentiation. Blood 2000, 96, 2740–2745. [Google Scholar] [CrossRef]
- Zou, Y.T.; Long, F.; Wu, C.Y.; Zhou, J.; Zhang, W.; Xu, J.D.; Zhang, Y.Q.; Li, S.L. A dereplication strategy for identifying triterpene acid analogues in poria cocos by comparing predicted and acquired uplc-esi-qtof-ms/ms data. Phytochem. Anal. 2019, 30, 292–310. [Google Scholar] [CrossRef]
- Jang, S.; Lee, A.; Hwang, Y.-H. Qualitative profiling and quantitative analysis of major constituents in jinmu-tang by uhplc-q-orbitrap-ms and uplc-tq-ms/ms. Molecules 2022, 27, 7887. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, X.; Wang, J.; Tang, M.; Yu, B.; Wang, Z.; Cao, L.; Chen, X.; Qian, M.; Wang, S.; et al. Identification and characterization of major alkaloid from sinomenium acutum stem and their metabolites after oral administration in rat plasma, urine, bile and feces based on uplc-q-tof/ms. J. Pharm. Biomed. Anal. 2022, 220, 115005. [Google Scholar] [CrossRef]
- Huang, Y.-F.; He, F.; Wang, C.-J.; Xie, Y.; Zhang, Y.-Y.; Sang, Z.; Qiu, P.; Luo, P.; Xiao, S.-Y.; Li, J.; et al. Discovery of chemical markers for improving the quality and safety control of Sinomenium acutum stem by the simultaneous determination of multiple alkaloids using uhplc-qqq-ms/ms. Sci. Rep. 2020, 10, 14182. [Google Scholar] [CrossRef]
- McFadyen, J.D.; Schaff, M.; Peter, K. Current and future antiplatelet therapies: Emphasis on preserving haemostasis. Nat. Rev. Cardiol. 2018, 15, 181–191. [Google Scholar] [CrossRef]
- Lebozec, K.; Jandrot-Perrus, M.; Avenard, G.; Favre-Bulle, O.; Billiald, P. Design, development and characterization of act017, a humanized fab that blocks platelet’s glycoprotein vi function without causing bleeding risks. mAbs 2017, 9, 945–958. [Google Scholar] [CrossRef]
- Ungerer, M.; Rosport, K.; Bultmann, A.; Piechatzek, R.; Uhland, K.; Schlieper, P.; Gawaz, M.; Munch, G. Novel antiplatelet drug revacept (dimeric glycoprotein vi-fc) specifically and efficiently inhibited collagen-induced platelet aggregation without affecting general hemostasis in humans. Circulation 2011, 123, 1891–1899. [Google Scholar] [CrossRef]
- Law, D.A.; Nannizzi-Alaimo, L.; Ministri, K.; Hughes, P.E.; Forsyth, J.; Turner, M.; Shattil, S.J.; Ginsberg, M.H.; Tybulewicz, V.L.; Phillips, D.R. Genetic and pharmacological analyses of syk function in alphaiibbeta3 signaling in platelets. Blood 1999, 93, 2645–2652. [Google Scholar] [CrossRef]
- van Eeuwijk, J.M.; Stegner, D.; Lamb, D.J.; Kraft, P.; Beck, S.; Thielmann, I.; Kiefer, F.; Walzog, B.; Stoll, G.; Nieswandt, B. The novel oral syk inhibitor, bl1002494, protects mice from arterial thrombosis and thromboinflammatory brain infarction. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1247–1253. [Google Scholar] [CrossRef]
- Braselmann, S.; Taylor, V.; Zhao, H.; Wang, S.; Sylvain, C.; Baluom, M.; Qu, K.; Herlaar, E.; Lau, A.; Young, C.; et al. R406, an orally available spleen tyrosine kinase inhibitor blocks fc receptor signaling and reduces immune complex-mediated inflammation. J. Pharmacol. Exp. Ther. 2006, 319, 998–1008. [Google Scholar] [CrossRef]
- Senis, Y.A.; Atkinson, B.T.; Pearce, A.C.; Wonerow, P.; Auger, J.M.; Okkenhaug, K.; Pearce, W.; Vigorito, E.; Vanhaesebroeck, B.; Turner, M.; et al. Role of the p110delta pi 3-kinase in integrin and itam receptor signalling in platelets. Platelets 2005, 16, 191–202. [Google Scholar] [CrossRef]
- Lian, L.; Wang, Y.; Draznin, J.; Eslin, D.; Bennett, J.S.; Poncz, M.; Wu, D.; Abrams, C.S. The relative role of plcbeta and pi3kgamma in platelet activation. Blood 2005, 106, 110–117. [Google Scholar] [CrossRef]
- Adam, F.; Kauskot, A.; Rosa, J.P.; Bryckaert, M. Mitogen-activated protein kinases in hemostasis and thrombosis. J. Thromb. Haemost. 2008, 6, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Gilio, K.; Munnix, I.C.; Mangin, P.; Cosemans, J.M.; Feijge, M.A.; van der Meijden, P.E.; Olieslagers, S.; Chrzanowska-Wodnicka, M.B.; Lillian, R.; Schoenwaelder, S.; et al. Non-redundant roles of phosphoinositide 3-kinase isoforms alpha and beta in glycoprotein vi-induced platelet signaling and thrombus formation. J. Biol. Chem. 2009, 284, 33750–33762. [Google Scholar] [CrossRef] [PubMed]
- Tseng, M.T.; Dozier, A.; Haribabu, B.; Graham, U.M. Transendothelial migration of ferric ion in fecl3 injured murine common carotid artery. Thromb. Res. 2006, 118, 275–280. [Google Scholar] [CrossRef]
- Eckly, A.; Hechler, B.; Freund, M.; Zerr, M.; Cazenave, J.P.; Lanza, F.; Mangin, P.H.; Gachet, C. Mechanisms underlying fecl3-induced arterial thrombosis. J. Thromb. Haemost. 2011, 9, 779–789. [Google Scholar] [CrossRef]
- Kurz, K.D.; Main, B.W.; Sandusky, G.E. Rat model of arterial thrombosis induced by ferric chloride. Thromb. Res. 1990, 60, 269–280. [Google Scholar] [CrossRef]
- Chukwumah, Y.; Walker, L.; Vogler, B.; Verghese, M. Changes in the phytochemical composition and profile of raw, boiled, and roasted peanuts. J. Agric. Food Chem. 2007, 55, 9266–9273. [Google Scholar] [CrossRef]
- Kim, T.W.; Han, J.M.; Han, Y.K.; Chung, H. Anti-inflammatory effects of Sinomenium acutum extract on endotoxin-induced uveitis in lewis rats. Int. J. Med. Sci. 2018, 15, 758–764. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, T.I.; Kim, K. An optimized herbal medicine containing Scutellaria baicalensis Georgi, Alisma orientale Juzepzuk, and Atractylodes japonica Koidzumi has potent antiplatelet and antithrombotic activities. J. Tradit. Complement. Med. 2023, 13, 285–296. [Google Scholar] [CrossRef]
- Kim, T.I.; Kim, Y.J.; Kim, K. Extract of seaweed inhibits integrin αiibβ3-induced outside-in signaling and arterial thrombosis. Front. Pharmacol. 2021, 12, 685948. [Google Scholar] [CrossRef]
- Amirou, A.; Bnouham, M.; Legssyer, A.; Ziyyat, A.; Aziz, M.; Berrabah, M.; Mekhfi, H. Effects of root bark extract on platelet aggregation, bleeding time, and plasmatic coagulation: And experiments. Evid.-Based Complement. Altern. Med. 2018, 2018, 7313517. [Google Scholar] [CrossRef]
- Endale, M.; Lee, W.M.; Kamruzzaman, S.M.; Kim, S.D.; Park, J.Y.; Park, M.H.; Park, T.Y.; Park, H.J.; Cho, J.Y.; Rhee, M.H. Ginsenoside-rp1 inhibits platelet activation and thrombus formation via impaired glycoprotein vi signalling pathway, tyrosine phosphorylation and mapk activation. Br. J. Pharmacol. 2012, 167, 109–127. [Google Scholar] [CrossRef]
- Kamruzzaman, S.M.; Endale, M.; Oh, W.J.; Park, S.C.; Kim, K.S.; Hong, J.H.; Kwak, Y.S.; Yun, B.S.; Rhee, M.H. Inhibitory effects of aqueous extract on agonist-induced platelet activation and thrombus formation involves mitogen-activated protein kinases. J. Ethnopharmacol. 2010, 130, 614–620. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Jang, S.A.; Kim, T.; Ha, H. Anti-osteoporotic and anti-adipogenic effects of rhus chinensis nutgalls in ovariectomized mice fed with a high-fat diet. Planta Med. 2019, 85, 1128–1135. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Rt (min) | Calculated (m/z) | Measured (m/z) | Error (ppm) | Adduct | Formula | MS/MS Fragments (m/z) | Identification |
|---|---|---|---|---|---|---|---|---|
| 1 | 4.79 | 272.1281 | 272.1283 | 0.6464 | M+H | C16H17NO3 | 255, 107, 161 | Higenamine * |
| 2 | 4.93 | 384.1208 | 384.1212 | 0.8195 | M+H | C18H22ClNO6 | 305, 341 | Acutumidine [16] |
| 3 | 5.06 | 398.1365 | 398.1369 | 1.0345 | M+H | C19H24ClNO6 | 305, 341 | Acutumine [17] |
| 4 | 5.06 | 330.17 | 330.1703 | 0.9331 | M+H | C19H23NO4 | - | Sinomenine [17] |
| 5 | 5.29 | 323.1754 | 323.1761 | 2.0296 | M+H | C20H22N2O2 | 278 | Gelsemine * |
| 6 | 5.69 | 342.17 | 342.1703 | 0.9004 | M+ | C20H24NO4 | 265, 297 | Magnoflorine * |
| 7 | 6.96 | 193.0495 | 193.0499 | 1.9244 | M+H | C10H8O4 | 105, 133, 161 | Scopoletin * |
| 8 | 8.15 | 338.1387 | 338.1389 | 0.5175 | M+ | C20H20NO4 | - | Columbamine or Jatrorrhizine * |
| 9 | 8.89 | 352.1543 | 352.1546 | 0.6859 | M+ | C21H22NO4 | 336 | Palmatine * |
| 10 | 5.17 | 417.1402 | 417.1412 | 2.4101 | M+HCO2 | C17H24O9 | 149, 271 | Syringin * |
| 11 | 6.21 | 787.2666 | 787.2681 | 1.8373 | M+HCO2 | C34H46O18 | 417 | Eleutheroside E * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-J.; Kim, T.I.; Lee, A.; Kim, K.; Hwang, Y.-H. Sinomenium acutum Modulates Platelet Aggregation and Thrombus Formation by Regulating the Glycoprotein VI-Mediated Signalosome in Mice. Pharmaceuticals 2024, 17, 6. https://doi.org/10.3390/ph17010006
Kim Y-J, Kim TI, Lee A, Kim K, Hwang Y-H. Sinomenium acutum Modulates Platelet Aggregation and Thrombus Formation by Regulating the Glycoprotein VI-Mediated Signalosome in Mice. Pharmaceuticals. 2024; 17(1):6. https://doi.org/10.3390/ph17010006
Chicago/Turabian StyleKim, Yeon-Ji, Tae In Kim, Ami Lee, Kyungho Kim, and Youn-Hwan Hwang. 2024. "Sinomenium acutum Modulates Platelet Aggregation and Thrombus Formation by Regulating the Glycoprotein VI-Mediated Signalosome in Mice" Pharmaceuticals 17, no. 1: 6. https://doi.org/10.3390/ph17010006
APA StyleKim, Y.-J., Kim, T. I., Lee, A., Kim, K., & Hwang, Y.-H. (2024). Sinomenium acutum Modulates Platelet Aggregation and Thrombus Formation by Regulating the Glycoprotein VI-Mediated Signalosome in Mice. Pharmaceuticals, 17(1), 6. https://doi.org/10.3390/ph17010006