
Neuroprotective Effect of Nosustrophine in a 3xTg Mouse Model of Alzheimer’s Disease





Abstract
:1. Introduction
2. Results
2.1. Biochemical Effect of Nosustrophine in Serum from 3xTg-AD Mice
2.2. In Vivo Experimentation
2.3. The Neurotrophic Action of Nosustrophine Induces Neuronal Development
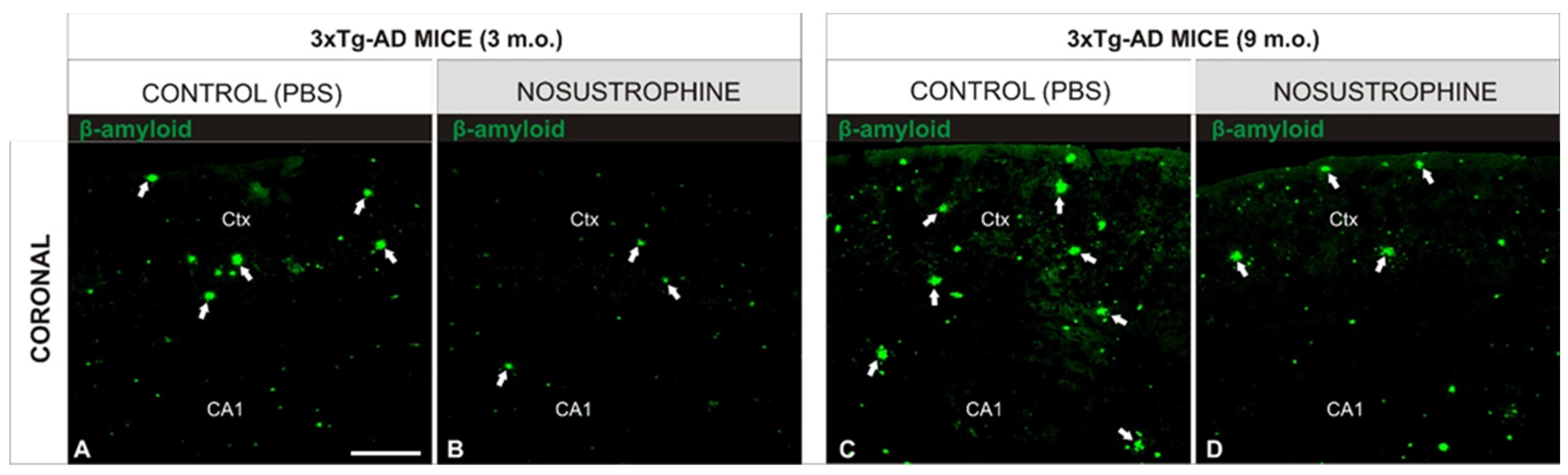
2.4. Nosustrophine Disrupts the Progressive Neuropathology in 3xTg-AD Mice
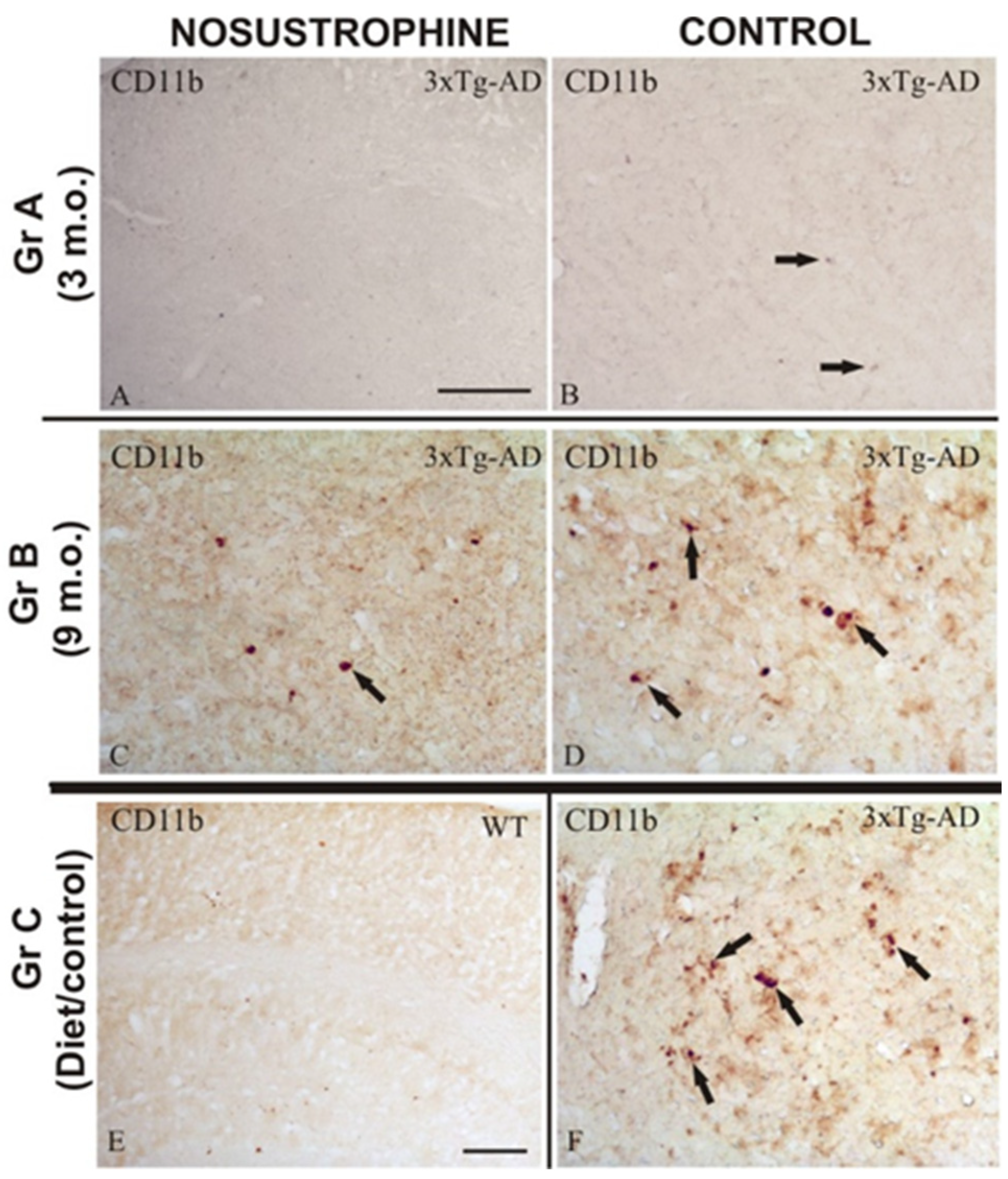
2.5. Nosustrophine Reduces Inflammatory Activity in Cortical and Hippocampal Regions
2.6. Neuroprotective Effect of Nosustrophine on 3xTg-AD Mice Neuropathology
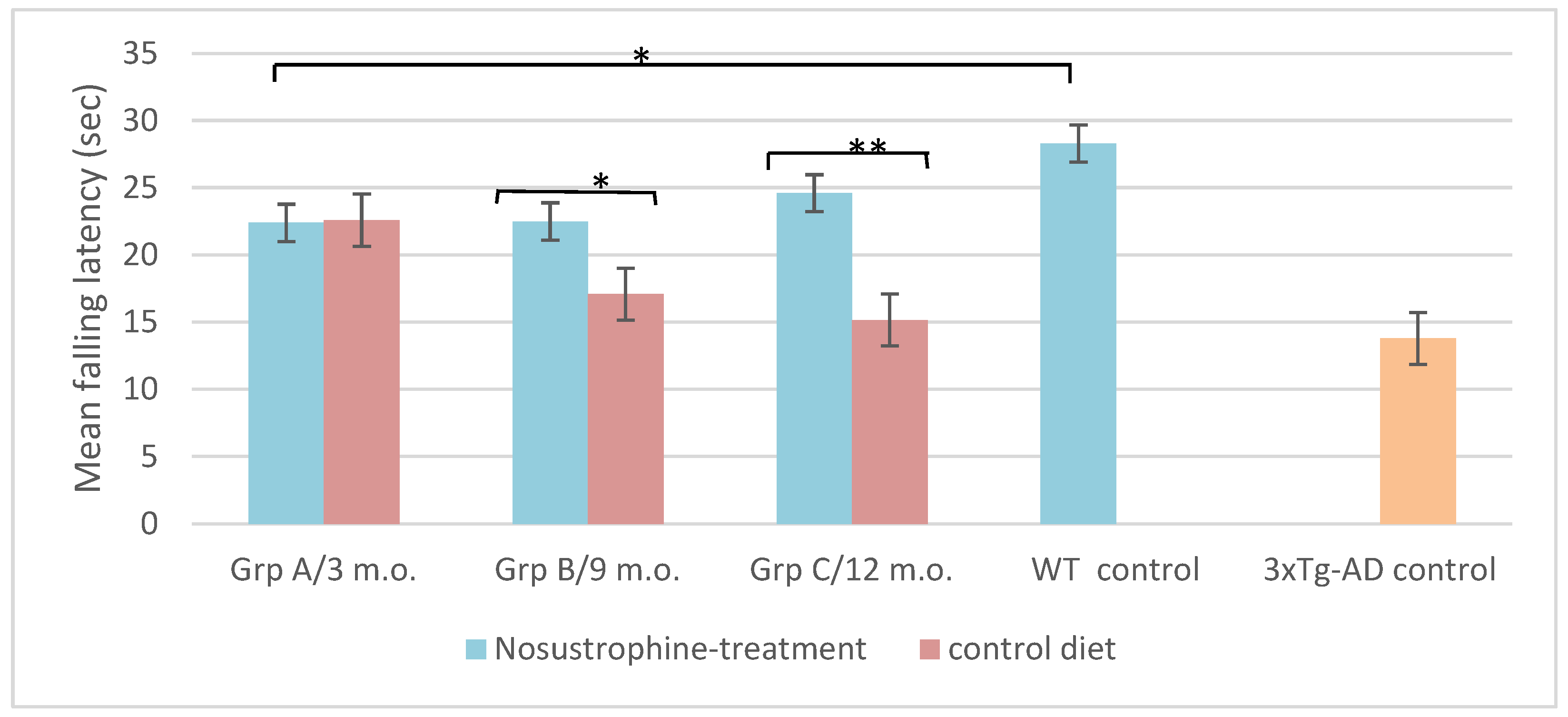
2.7. Nosustrophine Modulate Motor Coordination Performances in 3xTg-AD Mice
3. Discussion
4. Materials and Methods
4.1. Mouse Models
4.2. Biochemical Characterization of Nosustrophine
4.3. Experimental Design
4.4. Immunohistochemistry and Motor Function Evaluation
4.5. Imaging
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Cheng, F.; Takeda, S.; Lai, K.; Lin, L.; Sakata, R. Effects of porcine brain hydrolysate on impairment of cognitive learning ability in amyloid β(1-40)-infused rats. Anim. Sci. J. 2019, 90, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Tuszynski, M.; Yang, J.; Barba, D.; Hoi-Sang, U.; Bakay, R.; Pay, M.; Masliah, E.; Conner, J.; Kobalka, P.; Roy, S.; et al. Nerve Growth Factor Gene Therapy: Activation of Neuronal Responses in Alzheimer Disease. JAMA Neurol. 2015, 72, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Froestl, W.; Muhs, A.; Pfeifer, A. Cognitive enhancers (Nootropics). Part 1: Drugs interacting with receptors. J. Alzheimer’s Dis. 2014, 41, 961–1019. [Google Scholar] [CrossRef] [PubMed]
- Binder, D.; Scharfman, H. Brain-derived neurotrophic factor. Growth Factors 2004, 22, 123–131. [Google Scholar] [CrossRef]
- Zhang, L.; Fang, Y.; Lian, Y.; Chen, Y.; Wu, T.; Zheng, Y.; Zong, H.; Sun, L.; Zhang, R.; Wang, Z.; et al. Brain-Derived Neurotrophic Factor Ameliorates Learning Deficits in a Rat Model of Alzheimer’s Disease Induced by Aβ1-42. PLoS ONE 2015, 10, e0122415. [Google Scholar] [CrossRef]
- Zhang, T.; Kim, M.J.; Kim, M.J.; Wu, X.; Yang, H.J.; Yuan, H.; Huang, S.; Yoon, S.M.; Kim, K.N.; Park, S. Long-Term Effect of Porcine Brain Enzyme Hydrolysate Intake on Scopolamine-Induced Memory Impairment in Rats. Int. J. Mol. Sci. 2022, 23, 3361. [Google Scholar] [CrossRef]
- Thukham-Mee, W.; Wattanathorn, J.; Muchimapura, S.; Bunchonglikitkul, C. Neuroprotective Effect against Alzheimer’s Disease of Porcine Brain Extract. Am. J. Appl. Sci. 2012, 9, 700–708. [Google Scholar]
- Chao, M.; Rajagopal, R.; Lee, F. Neurotrophin signalling in health and disease. Clin. Sci. 2006, 110, 167–173. [Google Scholar] [CrossRef]
- Kimura, A.; Namekata, K.; Guo, X.; Harada, C.; Harada, T. Neuroprotection, Growth Factors and BDNF-TrkB Signalling in Retinal Degeneration. Int. J. Mol. Sci. 2016, 17, 1584. [Google Scholar] [CrossRef]
- Venkatesan, R.; Ji, E.; Kim, S. Phytochemicals that regulate neurodegenerative disease by targeting neurotrophins: A comprehensive review. BioMed. Res. Int. 2015, 2015, 814068. [Google Scholar] [CrossRef]
- Mitre, M.; Mariga, A.; Chao, M. Neurotrophin signalling: Novel insights into mechanisms and pathophysiology. Clin. Sci. 2017, 131, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Sisodiya, S. Precision medicine and therapies of the future. Epilepsia 2021, 62, S90–S105. [Google Scholar] [CrossRef] [PubMed]
- Carrera, I.; Fernandez-Novoa, L.; Aliev, G.; Vigo, C.; Cacabelos, R. Validating Immunotherapy in Alzheimer’s Disease: The EB101 Vaccine. Curr. Pharm. Des. 2016, 22, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Sadler, M.; Altman, R. Promises and challenges in pharmacoepigenetics. Camb. Prism. Precis. Med. 2023, 1, e18. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Iglesias, O.; Naidoo, V.; Carrera, I.; Corzo, L.; Cacabelos, R. Nosustrophine: An Epinutraceutical Bioproduct with Effects on DNA Methylation, Histone Acetylation and Sirtuin Expression in Alzheimer’s disease. Pharmaceutics 2022, 14, 2447. [Google Scholar] [CrossRef]
- Carrera, I.; Novoa, L.; Teijido, O.; Sampedro, C.; Seoane, S.; Lakshmana, M. Comparative Characterization Profile of Transgenic Mouse Models of Alzheimer’s Disease. J. Genom. Med. Pharmacogenom. 2017, 2, 331–337. [Google Scholar]
- Martínez-Iglesias, O.; Naidoo, V.; Carrera, I.; Cacabelos, R. Epigenetic Studies in the Male APP/BIN1/COPS5 Triple-Transgenic Mouse Model of Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 3, 2446. [Google Scholar] [CrossRef]
- Moneim, A. Oxidant/Antioxidant imbalance and the risk of Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 335–349. [Google Scholar] [CrossRef]
- Sopova, K.; Gatsiou, K.; Stellos, K.; Laske, C. Dysregulation of neurotrophic and haematopoietic growth factors in Alzheimer’s disease: From pathophysiology to novel treatment strategies. Curr. Alzheimer Res. 2014, 11, 27–39. [Google Scholar] [CrossRef]
- Martin, E.; Delarasse, C. Complex role of chemokine mediators in animal models of Alzheimer’s Disease. Biomed. J. 2018, 41, 34–40. [Google Scholar] [CrossRef]
- Eugenín, J.; Vecchiola, A.; Murgas, P.; Arroyo, P.; Cornejo, F.; von Bernhardi, R. Expression Pattern of Scavenger Receptors and Amyloid-β Phagocytosis of Astrocytes and Microglia in Culture are Modified by Acidosis: Implications for Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 53, 857–873. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cui, G.; Zhu, M.; Kang, X.; Guo, H. Neuroinflammation in Alzheimer’s disease: Chemokines produced by astrocytes and chemokine receptors. Int. J. Clin. Exp. Pathol. 2014, 7, 8342–8355. [Google Scholar] [PubMed]
- Boon, B.D.C.; Hoozemans, J.J.M.; Lopuhaä, B.; Eigenhuis, K.N.; Scheltens, P.; Kamphorst, W.; Rozemuller, A.J.M.; Bouwman, F.H. Neuroinflammation is increased in the parietal cortex of atypical Alzheimer’s disease. J. Neuroinflammation 2018, 15, 170. [Google Scholar] [CrossRef] [PubMed]
- Ballou, S.; Kushner, I. Laboratory evaluation of inflammation. In Kelley’s Textbook of Rheumatology, 7th ed.; Harris, E.D., Jr., Budd, R.C., Firestein, G.S., Genovese, M.C., Sergent, J.S., Ruddy, S., Sledge, C.B., Eds.; Elsevier: Philadelphia, PA, USA, 2005; pp. 720–727. [Google Scholar]
- Toriumi, K.; Miyashita, M.; Suzuki, K.; Yamasaki, N.; Yasumura, M.; Horiuchi, Y.; Yoshikawa, A.; Asakura, M.; Usui, N.; Itokawa, M.; et al. Vitamin B6 deficiency hyperactivates the noradrenergic system, leading to social deficits and cognitive impairment. Transl. Psychiatry 2021, 11, 262. [Google Scholar] [CrossRef]
- Tardy, A.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients 2020, 12, 228. [Google Scholar] [CrossRef] [PubMed]
- Douaud, G.; Refsum, H.; de Jager, C.A.; Jacoby, R.; Nichols, T.E.; Smith, S.M.; Smith, A.D. Preventing Alzheimer’s disease-related gray matter atrophy by B-vitamin treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 9523–9528. [Google Scholar] [CrossRef]
- Vural, H.; Demirin, H.; Kara, Y.; Eren, I.; Delibas, N. Alterations of plasma magnesium, copper, zinc, iron and selenium concentrations and some related erythrocyte antioxidant enzyme activities in patients with Alzheimer’s disease. J. Trace Elem. Med. Biol. 2010, 24, 169–173. [Google Scholar] [CrossRef]
- Hristova, M. Metabolic syndrome-from the neurotrophic hypothesis to a theory. Med. Hypotheses 2013, 81, 627–634. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, Y.; Guo, T.; Tang, C.; Liu, L.; Gao, D. High Concentration of Glial Cell Line-Derived Neurotrophic Factor Protects Primary Astrocytes from Apoptosis. Dev. Neurosci. 2018, 40, 134–144. [Google Scholar] [CrossRef]
- Khan, N.; Smith, M.T. Neurotrophins and Neuropathic Pain: Role in Pathobiology. Molecules 2015, 20, 10657–10688. [Google Scholar] [CrossRef]
- Bonomini, F.; Favero, G.; Castrezzati, S.; Borsani, E. Role of Neurotrophins in Orofacial Pain Modulation: A Review of the Latest Discoveries. Int. J. Mol. Sci. 2023, 24, 12438. [Google Scholar] [CrossRef] [PubMed]
- Jittiwat, J.; Wattanathorn, J.; Tongun, T.; Muchimapura, S.; Bunchonglikitkul, C. Porcine brain extract attenuates memory impairments induced by focal cerebral ischemia. Am. J. Appl. Sci. 2009, 6, 1662–1668. [Google Scholar] [CrossRef]
- Tsinman, O.; Tsinman, K.; Sun, N.; Avdeef, A. Physicochemical selectivity of the BBB microenvironment governing passive diffusion-matching with a porcine brain lipid extract artificial membrane permeability model. Pharm. Res. 2011, 28, 337–363. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, V.; Korolev, O.; Mares, V.; Pastalkova, E.; Bures, J. Hippocampal damage induced by carbon monoxide poisoning and spreading depression is alleviated by chronic treatment with brain derived polypeptides. Brain Res. 1999, 816, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Buresh, Y.; Koroleva, V.I.; Korolev, O.; Maresh, V. Changes in the constant potential in brain structures in rats during focal ischemia and systemic hypoxia. Neurosci. Behav. Physiol. 1999, 29, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Razavi, S.; Nazem, G.; Mardani, M.; Esfandiari, E.; Esfahani, S.; Salehi, H. Neurotrophic factors and their effects in the treatment of multiple sclerosis. Adv. Biomed. Res. 2015, 4, 53. [Google Scholar] [CrossRef]
- Mirowska-Guzel, D. The role of neurotrophic factors in the pathology and treatment of multiple sclerosis. Immunopharmacol. Immunotoxicol. 2009, 31, 32–38. [Google Scholar] [CrossRef]
- Bathina, S.; Das, U. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. AMS 2015, 11, 1164–1178. [Google Scholar] [CrossRef]
- Xu, J.; Sun, Y.; Wu, T.; Wang, B.; Liu, Y.; Zhang, J.; Lee, W.; Kang, Q.; Chai, Y.; Li, G. Porcine brain extract promotes osteogenic differentiation of bone marrow derived mesenchymal stem cells and bone consolidation in a rat distraction osteogenesis model. PLoS ONE 2017, 12, e0187362. [Google Scholar] [CrossRef]
- Reinhoud, N.; Brouwer, H.; van Heerwaarden, L.; Korte-Bouws, G. Analysis of Glutamate, GABA, Noradrenaline, Dopamine, Serotonin and Metabolites using microbore UHPLC with Electrochemical Detection. ACS Chem. Neurosci. 2013, 4, 888–894. [Google Scholar] [CrossRef]
- Nguyen, A.; Aerts, T.; Van Dam, D.; De Deyn, P. Biogenic amines and their metabolites in mouse brain tissue: Development, optimization and validation of an analytical HPLC method. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 3003–3014. [Google Scholar] [CrossRef] [PubMed]
- Yamatodani, A.; Fukuda, H.; Wada, H.; Iwaeda, T.; Watanabe, T. High-performance liquid chromatographic determination of plasma and brain histamine without previous purification of biological samples: Cation-exchange chromatography coupled with post-column derivatization fluorometry. J. Chromatogr. 1985, 344, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Burbano, C.; Cuadrado, C.; Muzquiz, M.; Cubero, J. Variation of favism-inducing factors (vicine, convicine and L-DOPA) during pod development in Vicia faba L. Plant Foods Hum. Nutr. 1995, 47, 265–275. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Diet | Nosustrophine | |||
| (g) | (% kcal) | (g) | (% kcal) | |
| Protein | 18.6 | 20 | 48 | 33 |
| Carbohydrates | 68.2 | 70 | 0 | 0 |
| Fat | 4.4 | 10 | 44 | 67 |
| Saturated | (2) | - | (18.9) | - |
| Monosaturated | (1.3) | - | (14.5) | - |
| Polyunsaturated | (1.1) | - | (10.6) | - |
| Total | 91.2 | 100 | 92 | 100 |
| Antibody | Antigen | Type | Source | Dilution | Ref. |
|---|---|---|---|---|---|
| β-Amyloid (IHC) | Aβ1-42 (mouse) | Mouse monoclonal | Millipore | 1:600 | 05-831-I |
| β-Amyloid (IF) | Aβ1-42 (rabbit) | Rabbit polyclonal | Invitrogen | 1:500 | 51-2700 |
| GFAP (IHC) | Glial fibrillary acidic protein (mouse) | Mouse monoclonal | Sigma | 1:100 | G-3893 |
| CD11b (IHC) | CD11/B-cells (rat) | Rat polyclonal | Millipore | 1:500 | MABF514 |
| TH (IF) | Tyrosine Hydroxylase (mouse) | Mouse monoclonal | Millipore | 1:500 | IHCR1005-6 |
| NeuN (IF) | Neuronal Nuclei (mouse) | Mouse monoclonal | Millipore | 1:500 | MAB377 |
| Cox-2 (IF) | Cyclooxygenase 2 (rabbit) | Mouse monoclonal | Vector | 1:400 | PA5-96081 |
| IL-17 (IF) | Interleukin 17 (rat) | Rat polyclonal | Invitrogen | 1:500 | 12-7177-81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrera, I.; Corzo, L.; Martínez-Iglesias, O.; Naidoo, V.; Cacabelos, R. Neuroprotective Effect of Nosustrophine in a 3xTg Mouse Model of Alzheimer’s Disease. Pharmaceuticals 2023, 16, 1306. https://doi.org/10.3390/ph16091306
Carrera I, Corzo L, Martínez-Iglesias O, Naidoo V, Cacabelos R. Neuroprotective Effect of Nosustrophine in a 3xTg Mouse Model of Alzheimer’s Disease. Pharmaceuticals. 2023; 16(9):1306. https://doi.org/10.3390/ph16091306
Chicago/Turabian StyleCarrera, Iván, Lola Corzo, Olaia Martínez-Iglesias, Vinogran Naidoo, and Ramón Cacabelos. 2023. "Neuroprotective Effect of Nosustrophine in a 3xTg Mouse Model of Alzheimer’s Disease" Pharmaceuticals 16, no. 9: 1306. https://doi.org/10.3390/ph16091306
APA StyleCarrera, I., Corzo, L., Martínez-Iglesias, O., Naidoo, V., & Cacabelos, R. (2023). Neuroprotective Effect of Nosustrophine in a 3xTg Mouse Model of Alzheimer’s Disease. Pharmaceuticals, 16(9), 1306. https://doi.org/10.3390/ph16091306