
EVOO Polyphenols Exert Anti-Inflammatory Effects on the Microglia Cell through TREM2 Signaling Pathway

 ,
,  ,
, 
 , , ,
, , , 
Abstract

1. Introduction
2. Results
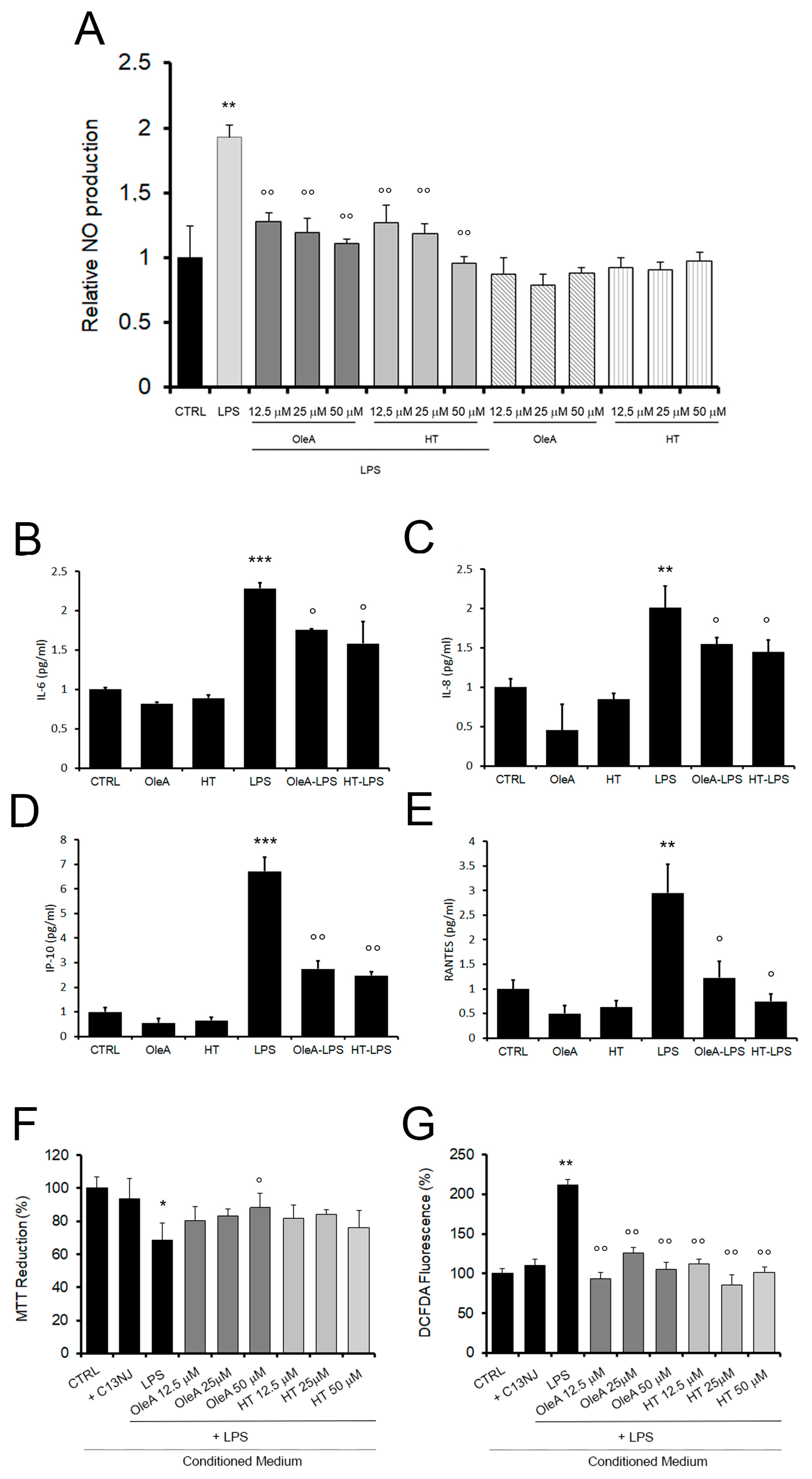
2.1. OleA and HT Reduce Oxidative Stress and Mitochondrial Damage in BV2 Cells Exposed to LPS
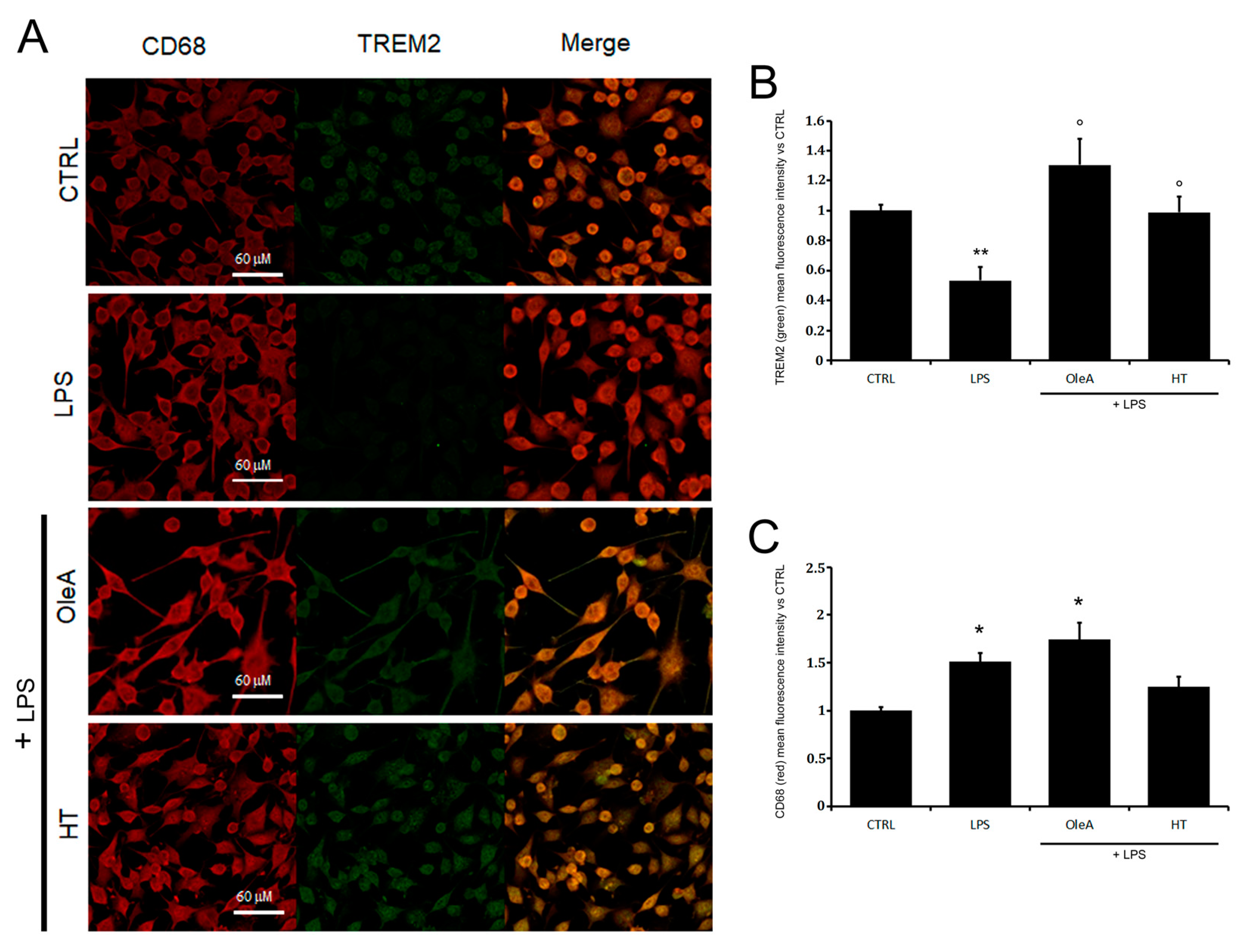
2.2. LPS Induces a Decrease in TREM2 Levels Which Is Prevented by Cell Exposure to Either Polyphenol
2.3. OleA and HT Ameliorate the Imbalance of TREM2 Signaling
2.4. OleA and HT Suppress the LPS-Induced Phosphorylation and Nuclear Translocation of NF-κB
2.5. Olive Polyphenols Decrease the LPS-Induced Production of Cytokine, Chemokine and Nitric Oxide
2.6. M2 Phenotype Characterization
3. Discussion
4. Materials and Methods
4.1. Cell Line and Culture Conditions
4.2. Preparation of Oleuropein Aglycone (OleA) and Hydroxytyrosol (HT) Samples
4.3. MTT Reduction Assay
4.4. Intracellular ROS Determination
4.5. Western Blot Analysis
4.6. Immunofluorescence
4.7. NO Assay
4.8. Determination of Cytokine Release
4.9. Assay of Cytotoxicity to Neuronal Cells of the Factors Released by Microglia
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Sarlus, H.; Heneka, M.T. Microglia in Alzheimer’s disease. J. Clin. Investig. 2017, 127, 3240–3249. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Jedrychowski, M.P.; Moore, C.S.; Cialic, R.; Lanser, A.J.; Gabriely, G.; Koeglsperger, T.; Dake, B.; Wu, P.M.; Doykan, C.E.; et al. Identification of a unique TGF-β-dependent molecular and functional signature in microglia. Nat. Neurosci. 2014, 17, 131–143. [Google Scholar] [CrossRef]
- Gautier, E.L.; Shay, T.; Miller, J.; Greter, M.; Jakubzick, C.; Ivanov, S.; Helft, J.; Chow, A.; Elpek, K.G.; Gordonov, S.; et al. Immunological Genome Consortium. Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat. Immunol. 2012, 13, 1118–1128. [Google Scholar] [CrossRef]
- Hickman, S.; Izzy, S.; Sen, P.; Morsett, L.; El Khoury, J. Microglia in neurodegeneration. Nat. Neurosci. 2018, 21, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Lambertsen, K.L.; Clausen, B.H.; Babcock, A.A.; Gregersen, R.; Fenger, C.; Nielsen, H.H.; Haugaard, L.S.; Wirenfeldt, M.; Nielsen, M.; Dagnaes-Hansen, F.; et al. Microglia protect neurons against ischemia by synthesis of tumor necrosis factor. J. Neurosci. 2009, 29, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, R.; Lu, H.; Butovsky, O.; Ohno, N.; Rietsch, A.M.; Cialic, R.; Wu, P.M.; Doykan, C.E.; Lin, J.; Cotleur, A.C.; et al. Differential roles of microglia and monocytes in the inflamed central nervous system. J. Exp. Med. 2014, 211, 1533–1549. [Google Scholar] [CrossRef]
- Krasemann, S.; Madore, C.; Cialic, R.; Baufeld, C.; Calcagno, N.; El Fatimy, R.; Beckers, L.; O’Loughlin, E.; Xu, Y.; Fanek, Z.; et al. The TREM2-APOE Pathway Drives the Transcriptional Phenotype of Dysfunctional Microglia in Neurodegenerative Diseases. Immunity 2017, 47, 566–581. [Google Scholar] [CrossRef]
- Efthymiou, A.G.; Goate, A.M. Late onset Alzheimer’s disease genetics implicates microglial pathways in disease risk. Mol. Neurodegener. 2017, 12, 43. [Google Scholar] [CrossRef]
- Leyns, C.E.G.; Ulrich, J.D.; Finn, M.B.; Stewart, F.R.; Koscal, L.J.; Remolina Serrano, J.; Robinson, G.O.; Anderson, E.; Colonna, M.; Holtzman, D.M. TREM2 deficiency attenuates neuroinflammation and protects against neurodegeneration in a mouse model of tauopathy. Proc. Natl. Acad. Sci. USA 2017, 114, 11524–11529. [Google Scholar] [CrossRef]
- Paloneva, J.; Kestilä, M.; Wu, J.; Salminen, A.; Böhling, T.; Ruotsalainen, V.; Hakola, P.; Bakker, A.B.; Phillips, J.H.; Pekkarinen, P.; et al. Loss-of-function mutations in TYROBP (DAP12) result in a presenile dementia with bone cysts. Nat. Genet. 2000, 25, 357–361. [Google Scholar] [CrossRef]
- Ulland, T.K.; Song, W.M.; Huang, S.C.; Ulrich, J.D.; Sergushichev, A.; Beatty, W.L.; Loboda, A.A.; Zhou, Y.; Cairns, N.J.; Kambal, A.; et al. TREM2 maintains microglial metabolic fitness in Alzheimer’s disease. Cell 2017, 170, 649–663.E13. [Google Scholar] [CrossRef]
- Colonna, M.; Wang, Y. TREM2 variants: New keys to decipher Alzheimer disease pathogenesis. Nat. Rev. Neurosci. 2016, 17, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Cheng, B.; Li, Y.; Li, X.; Chen, X.; Zhang, Y.W. TREM2 in Alzheimer’s Disease: Microglial Survival and Energy Metabolism. Front. Aging Neurosci. 2018, 10, 395. [Google Scholar] [CrossRef]
- Guerreiro, R.; Wojtas, A.; Bras, J.; Carrasquillo, M.; Rogaeva, E.; Majounie, E.; Cruchaga, C.; Sassi, C.; Kauwe, J.S.; Younkin, S.; et al. Alzheimer Genetic Analysis Group. TREM2 variants in Alzheimer’s disease. N. Engl. J. Med. 2013, 368, 117–127. [Google Scholar] [CrossRef]
- Jonsson, T.; Stefansson, H.; Steinberg, S.; Jonsdottir, I.; Jonsson, P.V.; Snaedal, J.; Bjornsson, S.; Huttenlocher, J.; Levey, A.I.; Lah, J.J.; et al. Variant of TREM2 associated with the risk of Alzheimer’s disease. N. Engl. J. Med. 2013, 368, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.D.; Daggett, A.; Gu, X.; Jiang, L.L.; Langfelder, P.; Li, X.; Wang, N.; Zhao, Y.; Park, C.S.; Cooper, Y.; et al. Elevated TREM2 Gene Dosage Reprograms Microglia Responsivity and Ameliorates Pathological Phenotypes in Alzheimer’s Disease Models. Neuron 2018, 7, 1032–1048.e5. [Google Scholar] [CrossRef] [PubMed]
- Bemiller, S.M.; McCray, T.J.; Allan, K.; Formica, S.V.; Xu, G.; Wilson, G.; Kokiko-Cochran, O.N.; Crish, S.D.; Lasagna-Reeves, C.A.; Ransohoff, R.M.; et al. TREM2 deficiency exacerbates tau pathology through dysregulated kinase signaling in a mouse model of tauopathy. Mol. Neurodegener. 2017, 12, 74. [Google Scholar] [CrossRef]
- Xu, X.; Du, L.; Jiang, J.; Yang, M.; Wang, Z.; Wang, Y.; Tang, T.; Fu, X.; Hao, J. Microglial TREM2 Mitigates Inflammatory Responses and Neuronal Apoptosis in Angiotensin II-Induced Hypertension in Middle-Aged Mice. Front. Aging Neurosci. 2021, 13, 716917. [Google Scholar] [CrossRef]
- Zhong, L.; Chen, X.F.; Zhang, Z.L.; Wang, Z.; Shi, X.Z.; Xu, K.; Zhang, Y.W.; Xu, H.; Bu, G. DAP12 stabilizes the C-terminal fragment of the triggering receptor expressed on myeloid cells-2 (TREM2) and protects against lps-induced pro-inflammatory response. J. Biol. Chem. 2015, 290, 15866–15877. [Google Scholar] [CrossRef]
- Zhong, L.; Chen, X.F.; Wang, T.; Wang, Z.; Liao, C.; Wang, Z.; Huang, R.; Wang, D.; Li, X.; Wu, L.; et al. Soluble TREM2 induces inflammatory responses and enhances microglial survival. J. Exp. Med. 2017, 214, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Jia, L.; Liu, C.C.; Rong, Z.; Zhong, L.; Yang, L.; Chen, X.F.; Fryer, J.D.; Wang, X.; Zhang, Y.W.; et al. TREM2 promotes microglial survival by activating Wnt/beta-catenin pathway. J. Neurosci. 2017, 37, 1772–1784. [Google Scholar] [CrossRef] [PubMed]
- Jay, T.R.; von Saucken, V.E.; Landreth, G.E. TREM2 in neurodegenerative diseases. Mol. Neurodegener. 2017, 12, 56. [Google Scholar] [CrossRef]
- Wunderlich, P.; Glebov, K.; Kemmerling, N.; Tien, N.T.; Neumann, H.; Walter, J. Sequential proteolytic processing of the triggering receptor expressed on myeloid cells-2 (TREM2) protein by ectodomain shedding and γ-secretase-dependent intramembranous cleavage. J. Biol. Chem. 2013, 288, 33027–33036. [Google Scholar] [CrossRef]
- Lessard, C.B.; Malnik, S.L.; Zhou, Y.; Ladd, T.B.; Cruz, P.E.; Ran, Y.; Mahan, T.E.; Chakrabaty, P.; Holtzman, D.M.; Ulrich, J.D.; et al. High-affinity interactions and signal transduction between Aβ oligomers and TREM2. EMBO Mol. Med. 2018, 10, e9027. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, X.; Li, X.; Jiang, L.L.; Gui, X.; Liu, Y.; Sun, Y.; Zhu, B.; Piña-Crespo, J.C.; Zhang, M.; et al. TREM2 Is a Receptor for β-Amyloid that Mediates Microglial Function. Neuron 2018, 97, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Udeochu, J.; Sayed, F.A.; Gan, L. TREM2 and amyloid β: A love-hate relationship. Neuron 2018, 97, 991–993. [Google Scholar] [CrossRef]
- Parhizkar, S.; Arzberger, T.; Brendel, M.; Kleinberger, G.; Deussing, M.; Focke, C.; Nuscher, B.; Xiong, M.; Ghasemigharagoz, A.; Katzmarski, N.; et al. Loss of TREM2 function increases amyloid seeding but reduces plaque-associated ApoE. Nat. Neurosci. 2019, 22, 191–204. [Google Scholar] [CrossRef]
- Hou, J.; Chen, Y.; Grajales-Reyes, G.; Colonna, M. TREM2 dependent and independent functions of microglia in Alzheimer’s disease. Mol. Neurodegener. 2022, 17, 84. [Google Scholar] [CrossRef]
- Leri, M.; Natalello, A.; Bruzzone, E.; Stefani, M.; Bucciantini, M. Oleuropein aglycone and hydroxytyrosol interfere differently with toxic Aβ 1-42 aggregation. Food Chem. Toxicol. 2019, 129, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef]
- Rigacci, S.; Miceli, C.; Nediani, C.; Berti, A.; Cascella, R.; Pantano, D.; Nardiello, P.; Luccarini, I.; Casamenti, F.; Stefani, M. Oleuropein aglycone induces autophagy via the AMPK/mTOR signalling pathway: A mechanistic insight. Oncotarget 2015, 6, 35344–35357. [Google Scholar] [CrossRef] [PubMed]
- Grossi, C.; Rigacci, S.; Ambrosini, S.; Ed Dami, T.; Luccarini, I.; Traini, C.; Failli, P.; Berti, A.; Casamenti, F.; Stefani, M. The polyphenol oleuropein aglycone protects TgCRND8 mice against Aß plaque pathology. PLoS ONE 2013, 8, e71702. [Google Scholar] [CrossRef] [PubMed]
- Luccarini, I.; Grossi, C.; Rigacci, S.; Coppi, E.; Pugliese, A.M.; Pantano, D.; la Marca, G.; Ed Dami, T.; Berti, A.; Stefani, M.; et al. Oleuropein aglycone protects against pyroglutamylated-3 amyloid-ß toxicity: Biochemical, epigenetic and functional correlates. Neurobiol. Aging 2015, 36, 648–663. [Google Scholar] [CrossRef] [PubMed]
- Fuccelli, R.; Fabiani, R.; Rosignoli, P. Hydroxytyrosol Exerts Anti-Inflammatory and Anti-Oxidant Activities in a Mouse Model of Systemic Inflammation. Molecules 2018, 23, 3212. [Google Scholar] [CrossRef] [PubMed]
- Wendimu, M.Y.; Hooks, S.B. Microglia Phenotypes in Aging and Neurodegenerative Diseases. Cells 2022, 11, 2091. [Google Scholar] [CrossRef]
- Serra, A.; Rubió, L.; Borràs, X.; Macià, A.; Romero, M.P.; Motilva, M.J. Distribution of olive oil phenolic compounds in rat tissues after administration of a phenolic extract from olive cake. Mol. Nutr. Food Res. 2012, 56, 486–496. [Google Scholar] [CrossRef]
- Angeloni, C.; Malaguti, M.; Barbalace, M.C.; Hrelia, S. Bioactivity of Olive Oil Phenols in Neuroprotection. Int. J. Mol. Sci. 2017, 18, 2230. [Google Scholar] [CrossRef] [PubMed]
- Grubić Kezele, T.; Ćurko-Cofek, B. Neuroprotective Panel of Olive Polyphenols: Mechanisms of Action, Anti-Demyelination, and Anti-Stroke Properties. Nutrients 2022, 14, 4533. [Google Scholar] [CrossRef]
- Guijarro-Muñoz, I.; Compte, M.; Álvarez-Cienfuegos, A.; Álvarez-Vallina, L.; Sanz, L. Lipopolysaccharide activates Toll-like receptor 4 (TLR4)-mediated NF-κB signaling pathway and proinflammatory response in human pericytes. J. Biol. Chem. 2014, 289, 2457–2468. [Google Scholar] [CrossRef] [PubMed]
- Azizi, G.; Navabi, S.S.; Al-Shukaili, A.; Seyedzadeh, M.H.; Yazdani, R.; Mirshafiey, A. The Role of Inflammatory Mediators in the Pathogenesis of Alzheimer’s Disease. Sultan Qaboos Univ. Med. J. 2015, 15, e305–e316. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.S.; Yang, X.; Chen, Z.G.; Lu, M.O.; Morris, C.; Winblad, B.; Zhu, J. Decreased fractalkine and increased IP-10 expression in aged brain of APP(swe) transgenic mice. Neurochem. Res. 2008, 33, 1085–1089. [Google Scholar] [CrossRef]
- Tripathy, D.; Thirumangalakudi, L.; Grammas, P. RANTES upregulation in the Alzheimer’s disease brain: A possible neuroprotective role. Neurobiol. Aging 2010, 31, 8–16. [Google Scholar] [CrossRef]
- Pey, P.; Pearce, R.K.; Kalaitzakis, M.E.; Griffin, W.S.; Gentleman, S.M. Phenotypic profile of alternative activation marker CD163 is different in Alzheimer’s and Parkinson’s disease. Acta Neuropathol. Commun. 2014, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Hou, X.H.; Yang, S.B. Lidocaine potentiates SOCS3 to attenuate inflammation in microglia and sup-press neuropathic pain. Cell. Mol. Neurobiol. 2019, 39, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Ikram, M.; Ullah, R.; Rehman, S.U.; Kim, M.O. Hesperetin, a citrus flavonoid, attenuates LPS-induced neuroinflammation, apoptosis and memory impairments by modulating TLR4/NF-κB signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef]
- Azam, S.; Haque, M.E.; Balakrishnan, R.; Kim, I.S.; Choi, D.K. The Ageing Brain: Molecular and Cellular Basis of Neurodegeneration. Front. Cell. Dev. Biol. 2021, 9, 683459. [Google Scholar] [CrossRef]
- Calvo-Rodriguez, M.; García-Rodríguez, C.; Villalobos, C.; Núñez, L. Role of Toll Like Receptor 4 in Alzheimer’s Disease. Front. Immunol. 2020, 11, 1588. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, W.; Zhang, M.; Tian, X.; Li, Y.; Lü, Y. Imbalance of Microglial TLR4/TREM2 in LPS-Treated APP/PS1 trans-genic mice: A potential link between Alzheimer’s disease and systemic inflammation. Neurochem. Res. 2019, 44, 1138–1151. [Google Scholar] [CrossRef]
- Ruganzu, J.B.; Peng, X.; He, Y.; Wu, X.; Zheng, Q.; Ding, B.; Lin, C.; Guo, H.; Yang, Z.; Zhang, X.; et al. Downregulation of TREM2 expression exacerbates neu-roinflammatory responses through TLR4-mediated MAPK signaling pathway in a trans-genic mouse model of Alzheimer’s disease. Mol. Immunol. 2022, 142, 22–36. [Google Scholar] [CrossRef]
- Fang, H.L.; Lai, J.T.; Lin, W.C. Inhibitory effect of olive oil on fibrosis induced by carbon tetrachloride in rat liver. Clin. Nutr. 2008, 27, 900–907. [Google Scholar] [CrossRef]
- Pojero, F.; Aiello, A.; Gervasi, F.; Caruso, C.; Ligotti, M.E.; Calabrò, A.; Procopio, A.; Candore, G.; Accardi, G.; Allegra, M. Effects of Oleuropein and Hydroxytyrosol on Inflammatory Mediators: Consequences on Inflammaging. Int. J. Mol. Sci. 2022, 24, 380. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, X.; Ding, B.; Zi, M.; Ma, Y. Oleuropein ameliorated lung ischemia-reperfusion injury by inhibiting TLR4 signaling cascade in alveolar macrophages. Transpl. Immunol. 2022, 74, 101664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, J.; Jiang, X.; Yang, L.; Zhang, Q.; Wang, B.; Cui, L.; Wang, X. Hydroxytyrosol inhibits LPS-induced neuroinflammatory responses via suppression of TLR-4-mediated NF-κB P65 activation and ERK aignaling pathway. Neuroscience 2020, 426, 189–200. [Google Scholar] [CrossRef]
- Blasi, E.; Barluzi, R.; Bocchini, V.; Mazolla, R.; Bistoni, F. Immortalization of murine microglia cells by a v-raf/v-myc carrying retrovirus. J. Neuroimmunol. 1990, 27, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Peudenier, S.; Héry, C.; Montagnier, L.; Tardieu, M. Human microglial cells: Characterization in cerebral tissue and in primary cultures and study of their susceptibility to HIV-I infection. Ann. Neurol. 1991, 29, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Branca, J.J.; Morucci, G.; Malentacchi, F.; Gelmini, S.; Ruggiero, M.; Pacini, S. Effects of oxaliplatin and oleic acid Gc-protein-derived macrophage-activating factor on murine and human microglia. J. Neurosci. Res. 2015, 93, 1364–1377. [Google Scholar] [CrossRef]
- Rigacci, S.; Guidotti, V.; Bucciantini, M.; Parri, M.; Nediani, C.; Cerbai, E.; Stefani, M.; Berti, A. Oleuropein aglycon prevents cytotoxic amyloid aggregation of human amylin. J. Nutr. Biochem. 2010, 21, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Zafra-Gómez, A.; Luzón-Toro, B.; Capel-Cuevas, S.; Morales, J.C. Stability of Hydroxytyrosol in Aqueous Solutions at Different Concentration, Temperature and with Different Ionic Content: A Study Using UPLC-MS. Food Nutr. Sci. 2011, 2, 1114–1120. [Google Scholar] [CrossRef]
- Vasarri, M.; Leri, M.; Barletta, E.; Ramazzotti, M.; Marzocchini, R.; Degl’Innocenti, D. Anti-inflammatory properties of the marine plant Posidonia oceanica (L.) Delile. J. Ethnopharmacol. 2020, 247, 112252. [Google Scholar] [CrossRef] [PubMed]
- Cabaro, S.; D’Esposito, V.; Di Matola, T.; Sale, S.; Cennamo, M.; Terracciano, D.; Parisi, V.; Oriente, F.; Portella, G.; Beguinot, F.; et al. Cytokine signature and COVID-19 prediction models in the two waves of pandemics. Sci. Rep. 2021, 11, 20793. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pg/mL | vs. CTRL | p Value vs. CTRL | p Value vs. LPS | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LPS | OleA | HT | OleA-LPS | HT-LPS | LPS | OleA | HT | OleA-LPS | HT-LPS | |
| PDGF-bb | 1.0903 ± 0.09 | 0.6379 ± 0.168 | 0.7284 ± 0.09 | 1.2715 ± 0.09 | 1 ± 0.12 | 0.21 | 0.052 | 0.047 | 0.21 | |
| IL-1b | 2.1052 ± 0.10 | 1.3421 ± 0.1438 | 1.2105 ± 0.1053 | 1.5526 ± 0.4038 | 1.5 ± 0.3163 | 0.0042 | 0.040 | 0.036 | 0.22 | 0.36 |
| IL-1ra | 1.1088 ± 0.0982 | 0.4309 ± 0.5 | 0.5899± 0.2696 | 1 ± 0.1 | 1.0560 ± 0.053 | 0.21 | 0.061 | 0.21 | 0.35 | |
| IL-2 | OOR< | OOR< | OOR< | OOR< | OOR< | |||||
| IL-4 | 1.5913 ± 0.1613 | 0.5484 ± 0.2391 | 0.9139 ± 0.1811 | 1.3118 ±0.1613 | 1.5806 ± 0.1839 | 0.035 | 0.069 | 0.33 | 0.47 | |
| IL-5 | 1.2829 ± 0.0368 | 0.7541 ± 0.0262 | 0.8543 ± 0.054 | 1.1453 ± 0.0393 | 1.1514 ± 0.1415 | 0.004 | 0.018 | 0.046 | 0.027 | 0.24 |
| IL-6 | 2.2838 ± 0.0731 | 0.8197 ± 0.0167 | 0.8890 ± 0.0409 | 1.7589 ± 0.0134 | 1.5844 ± 0.2757 | 0.0008 | 0.11 | 0.047 | 0.042 | 0.013 |
| IL-7 | OOR< | OOR< | OOR< | OOR< | OOR< | |||||
| IL-8 | 2.1090 ± 0.2778 | 0.4524 ± 0.3283 | 0.8465 ± 0.077 | 1.5480 ± 0.0831 | 1.4489 ± 0.3538 | 0.0033 | 0.0539 | 0.092 | 0.024 | 0.026 |
| IL-9 | 1.3405 ± 0.0719 | 0.6024 ± 0.1038 | 0.8301 ± 0.0639 | 1.0610 ± 0.061 | 1.0457 ± 0.1105 | 0.033 | 0.18 | 0.057 | 0.023 | 0.15 |
| IL-10 | OOR< | OOR< | OOR< | OOR< | OOR< | |||||
| IL-12(p70) | OOR< | OOR< | OOR< | OOR< | OOR< | |||||
| IL-13 | OOR< | OOR< | OOR< | OOR< | OOR< | |||||
| IL-15 | 0.8931 ± 0.13 | 0.3265 ± 0.5023 | 0.7995 ± 0.1969 | 0.6237 ± 0.2631 | 1.0892 ± 0.2832 | 0.23 | 0.16 | 0.14 | 0.30 | |
| IL-17 | OOR< | OOR< | OOR< | OOR< | OOR< | |||||
| Eotaxin | 1.2051 ± 0.1702 | 0.5448 ± 0.3412 | 0.9102 ± 0.0986 | 0.8653 ± 0.0519 | 1.1666 ± 0.1429 | 0.211 | 0.067 | 0.21 | 0.12 | 0.44 |
| FGF basic | 1.0424 ± 0.3307 | 0.2324 ± 0.1673 | 1.3788 ± 0.1958 | 0.8969 ± 0.1562 | 1.9322 ± 0.2904 | 0.45 | 0.19 | 0.11 | 0.34 | 0.12 |
| G-CSF | 1.4032 ± 0.2475 | 0.6091 ± 0.2208 | 0.6871 ± 0.0709 | 1.2981 ± 0.0703 | 0.6234 ± 0.4963 | 0.17 | 0.057 | 0.023 | 0.46 | 0.08 |
| GM-CSF | 3.6612 ± 0.3376 | 1.0414 ± 0.1134 | 0.6312 ± 0.1391 | 3.5369 ± 0.1074 | 1.0522 ± 0.47 | 0.07 | 0.37 | 0.045 | 0.26 | 0.14 |
| IFN-γ | 1.2878 ± 0.0959 | 0.5227 ± 0.4842 | 0.6060 ± 0.0758 | 1.0227 ± 0.3418 | 1.1439 ± 0.0964 | 0.057 | 0.10 | 0.017 | 0.39 | 0.11 |
| IP-10 | 6.7186 ± 0.6911 | 0.5578 ± 0.3781 | 0.6473 ± 0.1995 | 2.7464± 0.3379 | 2.4714 ± 0.4727 | 0.00016 | 0.12 | 0.10 | 0.0023 | 0.0022 |
| MCP-1 (MCAF) | 1.1680 ± 0.4202 | 0.1596 ± 0.4202 | 0.5798 ± 0.5942 | 0.5966 ± 0.844 | 1.2941 ± 0.4313 | 0.36 | 0.091 | 0.21 | 0.16 | 0.21 |
| MIP-1a | 1.4 ± 0.233 | 0.85 ± 0.522 | 0.625 ± 0.39 | 1 ±0.212 | 0.4 ± 0.15 | 0.15 | 0.14 | 0.15 | 0.028 | |
| MIP-1b | 1.6257 ± 0.130 | 0.7449 ± 0.122 | 0.8499 ± 0.048 | 1.3832 ± 0.093 | 1.3519 ± 0.144 | 0.048 | 0.054 | 0.029 | 0.21 | 0.22 |
| RANTES | 2.9518 ± 0.58 | 0.4961 ± 0.17 | 0.6270 ± 0.138 | 1.2278 ± 0.3339 | 0.7438 ± 0.150 | 0.0018 | 0.03 | 0.051 | 0.021 | 0.015 |
| TNF-α | 1.4048 ± 0.190 | 0.5788 ± 0.307 | 1.1433 ± 0.098 | 1.3056 ± 0.083 | 1.2037 ± 0.130 | 0.12 | 0.078 | 0.14 | 0.36 | 0.26 |
| VEGF | 1.4359 ± 0.091 | OOR< | 1.6765 ± 0.092 | 1.2007 ± 0.377 | 1.6316 ± 0.441 | 0.014 | 0.072 | 0.32 | 0.40 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leri, M.; Vasarri, M.; Carnemolla, F.; Oriente, F.; Cabaro, S.; Stio, M.; Degl’Innocenti, D.; Stefani, M.; Bucciantini, M. EVOO Polyphenols Exert Anti-Inflammatory Effects on the Microglia Cell through TREM2 Signaling Pathway. Pharmaceuticals 2023, 16, 933. https://doi.org/10.3390/ph16070933
Leri M, Vasarri M, Carnemolla F, Oriente F, Cabaro S, Stio M, Degl’Innocenti D, Stefani M, Bucciantini M. EVOO Polyphenols Exert Anti-Inflammatory Effects on the Microglia Cell through TREM2 Signaling Pathway. Pharmaceuticals. 2023; 16(7):933. https://doi.org/10.3390/ph16070933
Chicago/Turabian StyleLeri, Manuela, Marzia Vasarri, Federica Carnemolla, Francesco Oriente, Serena Cabaro, Maria Stio, Donatella Degl’Innocenti, Massimo Stefani, and Monica Bucciantini. 2023. "EVOO Polyphenols Exert Anti-Inflammatory Effects on the Microglia Cell through TREM2 Signaling Pathway" Pharmaceuticals 16, no. 7: 933. https://doi.org/10.3390/ph16070933
APA StyleLeri, M., Vasarri, M., Carnemolla, F., Oriente, F., Cabaro, S., Stio, M., Degl’Innocenti, D., Stefani, M., & Bucciantini, M. (2023). EVOO Polyphenols Exert Anti-Inflammatory Effects on the Microglia Cell through TREM2 Signaling Pathway. Pharmaceuticals, 16(7), 933. https://doi.org/10.3390/ph16070933