


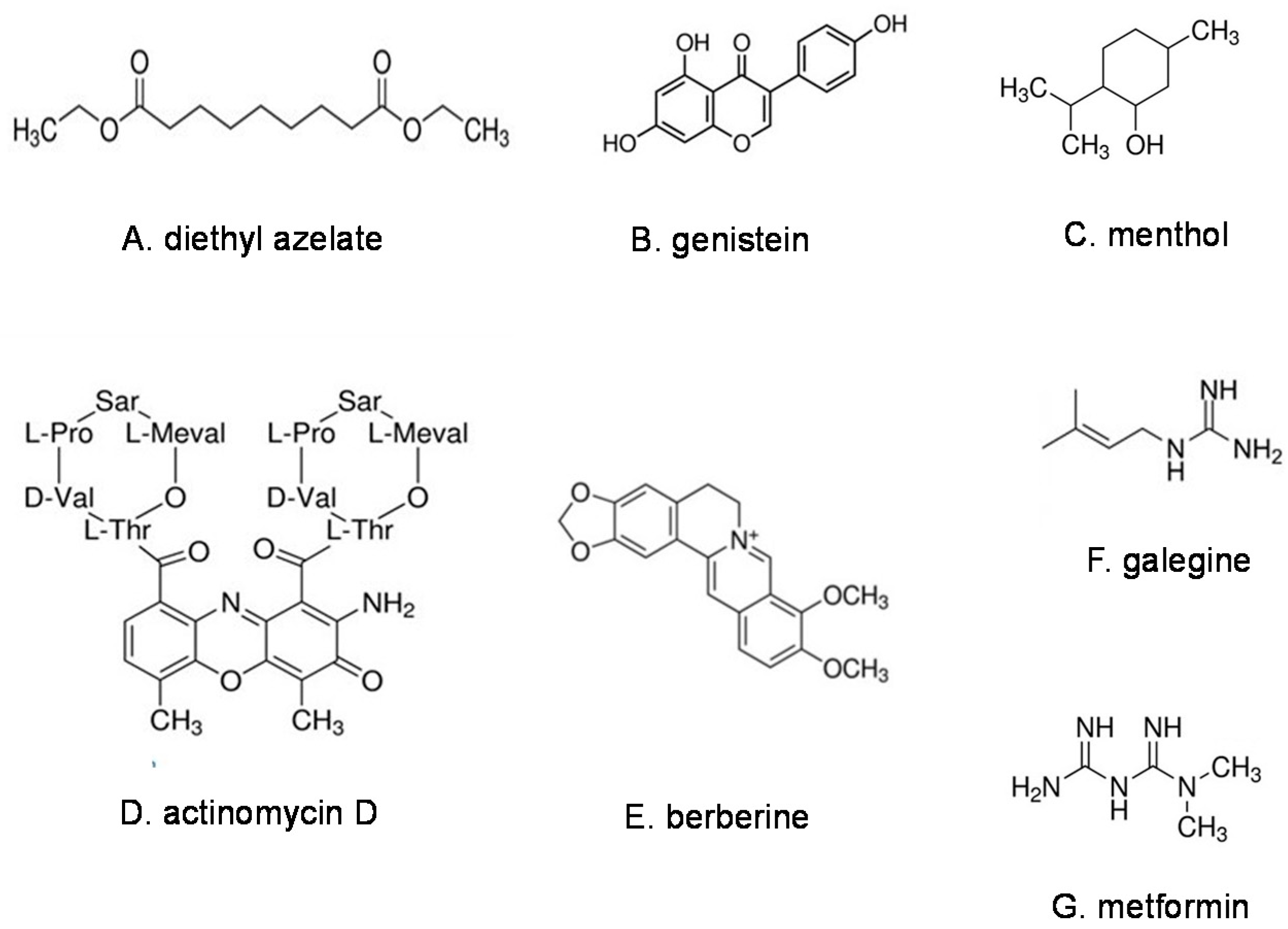
Mitigation of Insulin Resistance by Natural Products from a New Class of Molecules, Membrane-Active Immunomodulators


{kind=link}
Abstract
Share and Cite
Izbicka, E.; Streeper, R.T. Mitigation of Insulin Resistance by Natural Products from a New Class of Molecules, Membrane-Active Immunomodulators. Pharmaceuticals 2023, 16, 913. https://doi.org/10.3390/ph16070913
Izbicka E, Streeper RT. Mitigation of Insulin Resistance by Natural Products from a New Class of Molecules, Membrane-Active Immunomodulators. Pharmaceuticals. 2023; 16(7):913. https://doi.org/10.3390/ph16070913
Chicago/Turabian StyleIzbicka, Elzbieta, and Robert T. Streeper. 2023. "Mitigation of Insulin Resistance by Natural Products from a New Class of Molecules, Membrane-Active Immunomodulators" Pharmaceuticals 16, no. 7: 913. https://doi.org/10.3390/ph16070913
APA StyleIzbicka, E., & Streeper, R. T. (2023). Mitigation of Insulin Resistance by Natural Products from a New Class of Molecules, Membrane-Active Immunomodulators. Pharmaceuticals, 16(7), 913. https://doi.org/10.3390/ph16070913