
Gut-Derived Metabolite, Trimethylamine-N-oxide (TMAO) in Cardio-Metabolic Diseases: Detection, Mechanism, and Potential Therapeutics


Abstract
1. Introduction
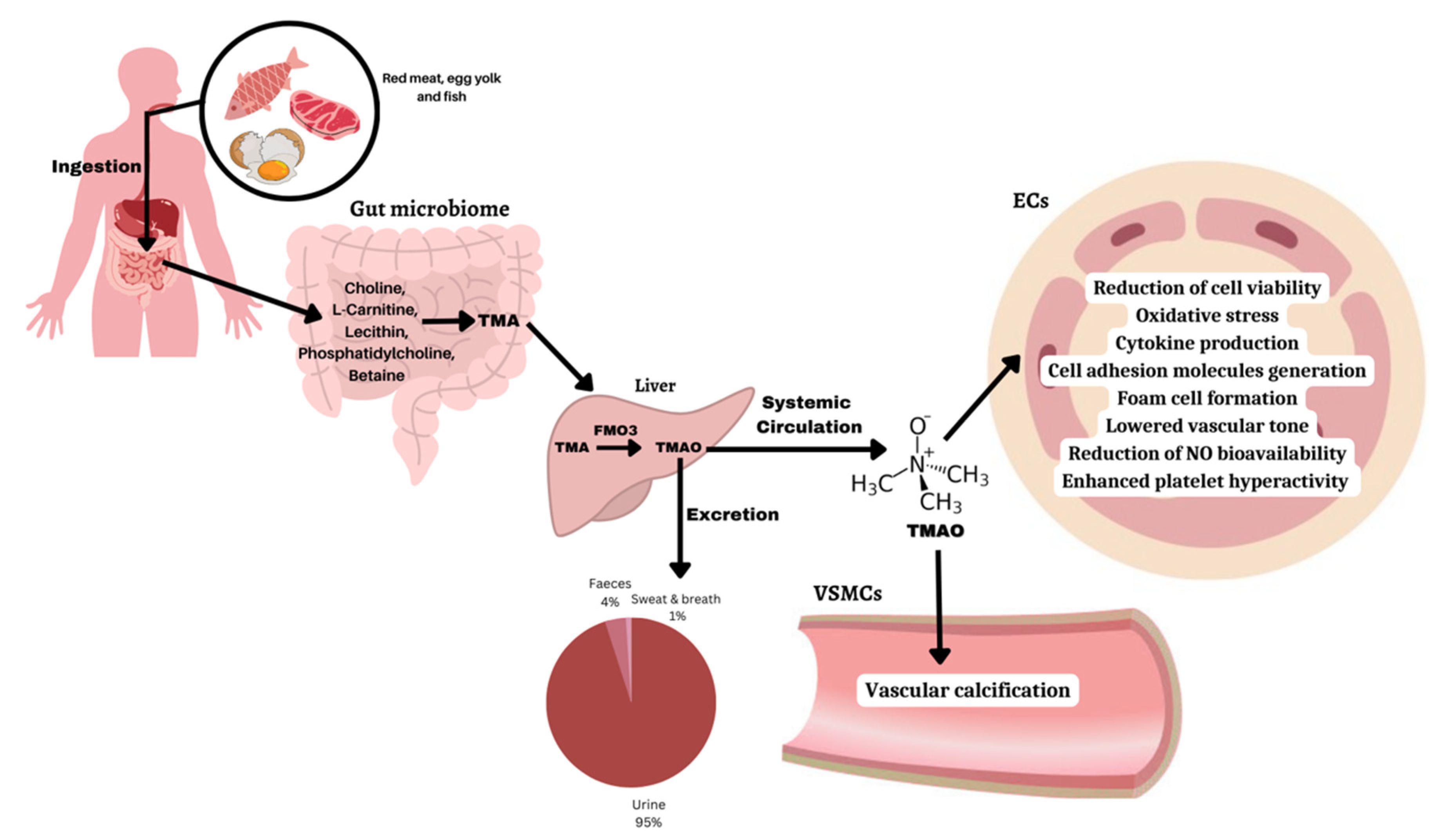
2. TMAO Metabolism, Biosynthesis, and Excretion
3. TMAO Detection and Measurement Methods
4. TMAO Level Variations and Disease Conditions
5. Molecular Mechanisms of TMAO-Induced Diseases (Table 3)
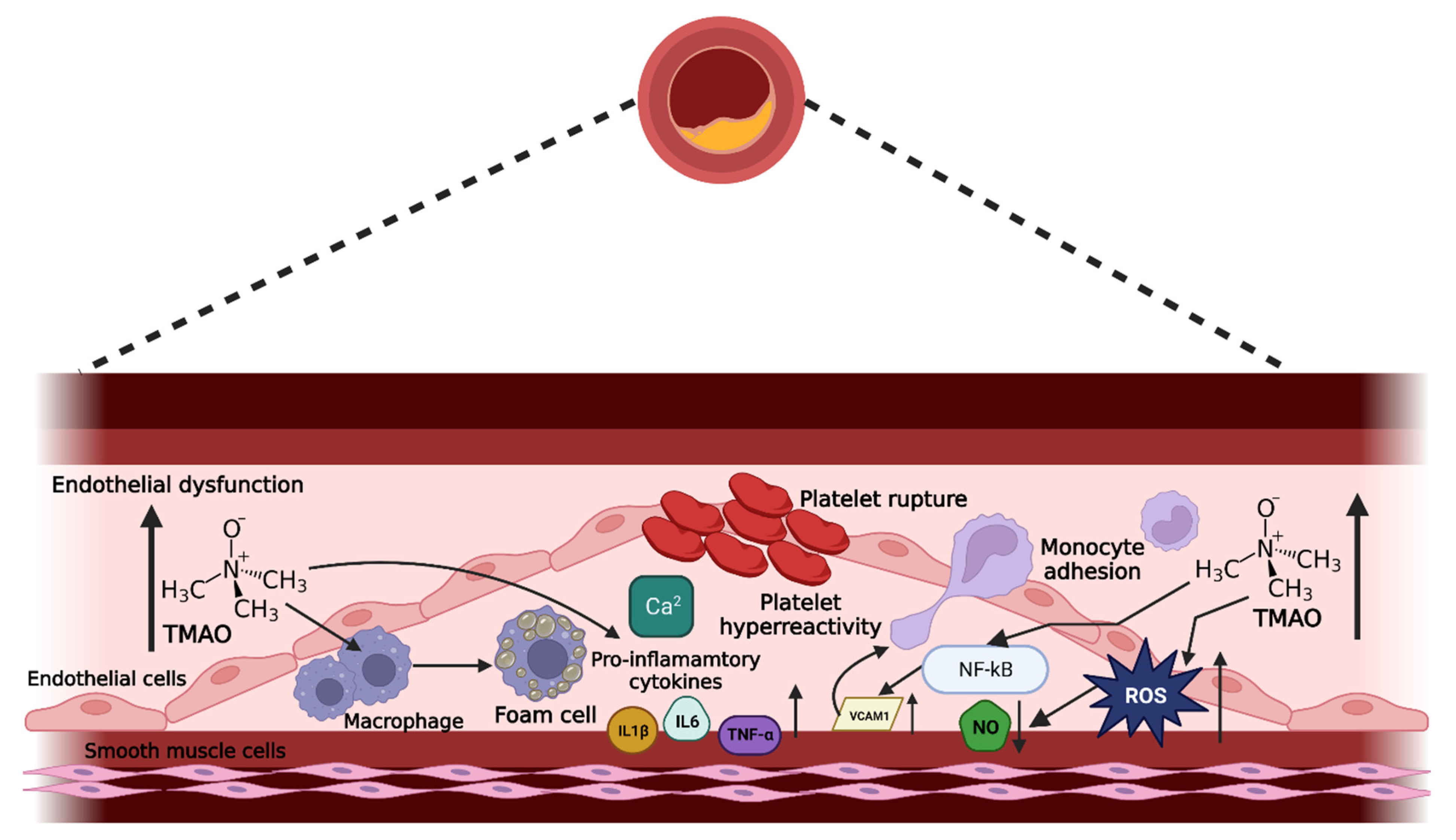
5.1. Endothelial Dysfunction Mediated by TMAO
5.1.1. Effect of TMAO on Cell Viability
5.1.2. TMAO Enhances Oxidative Stress
5.1.3. TMAO Induces Inflammation
Enhanced Cytokine Production
Activation of Adhesion Molecules
Elevated Foam Cell Formation
5.1.4. TMAO Reduces Vascular Tone
Effects of TMAO on NO Bioavailability
Association between TMAO and Hydrogen Sulfide (H2S)
Role of Prostanoids in Vasoconstriction
EDH in Endothelial Dysfunction
5.1.5. TMAO-Enhanced Platelet Hyperreactivity
5.2. TMAO Triggers Heart Failure
5.3. TMAO Promotes Metabolic Syndrome

{kind=link}
{kind=link}
| Disease | Species/Cells | Molecular Mechanisms | Potential Pathways Triggered | References |
|---|---|---|---|---|
| Atherosclerosis and CVD | THP-1y HUVECs (Human Umbilical Vein Endothelial Cells) | ↑ adhesion of monocytes, ↓ endothelial self-repair, endothelial dysfunction | Activation of PKC, NF-κB and VCAM-1 pathways | [56] |
| LDLR-/- mice fed a choline diet (aorta), HAECs (Human Aortic Endothelial Cells), VSMECs (Vascular Smooth Muscle Cells) | ↑ pro-inflammatory cytokines, ↑ leukocyte adhesion to endothelial wall | MAPK and NF-kB signaling pathway | [14] | |
| HUVECs and ApoE-/- mice (aorta) | Activation of NLRP3 inflammasome, ↑ vascular inflammation | Inhibition of SIRT3-SOD2-mitochondrial ROS signaling pathway | [61] | |
| Human and C57BL/6J ApoE-/- mice | ↓ bile acid synthetic enzyme, ↓ reverse cholesterol transport, ↓ lipid metabolism and transport | Unknown pathway, likely to be linked to Niemann Pick Cl-like1 (Npc1L1) | [11,12,114] | |
| ApoE-/- mice and ApoE-/- mice with a high-fat diet | ↑ macrophage scavenger receptors CD36 and SR-A1, ↑ lipid accumulation, ↑ foam cell formation | CD36-dependent MAPK/JNK pathway | [11,80,115] | |
| Fischer-344 rats | ↑ oxidative stress, ↑ pro-inflammatory cytokines, ↑ endothelial dysfunction and vascular inflammation | Unknown | [38] | |
| Heart failure | Male C57BL/6 mice and adult male Sprague–Dawley (SD) rats | Activation of NLRP3 inflammasome, triggers cardiac hypertrophy and fibrosis | Smad 3 signaling pathway | [116] |
| Male C57BL/6 mice and cultured cardiac fibroblasts | ↑ pro-inflammatory cytokines TNF and IL-1β, ↑ interstitial fibrosis in heart, ↑ myocardial inflammation, activation of NLRP3 inflammasome | NLRP3 inflammasome signaling pathway | [107] | |
| ICR mice and male Wistar rats | ↓ cardiac energy production, ↓ pyruvate dehydrogenase activity & fatty acid β-oxidation, ↑ glycogen synthesis, ↑ oxidative damage to proteins, mitochondrial dysfunction | Cardiac energy metabolism | [108] | |
| Kidney disease | Human and high-fat diet/low-dose streptozotocin-induced diabetes rats | ↑ pro-fibrotic factors TGF-β1, IL-1β and Smad3, ↑ phosphorylation and Smad3 activation, ↑ kidney injury molecule-1, activation of NLRP3 inflammasome, renal inflammation, renal fibrosis and renal dysfunction | NLRP3 inflammasome signaling pathway, transforming growth factor β, SMAD signaling pathway | [117,118] |
| Metabolic dysfunction | Male C57BL/6 mice with high-fat diet (HFD)-fed | ↑ insulin signaling pathway, ↑ glycogen synthesis, ↑ gluconeogenesis and glucose transport in liver, impaired glucose tolerance, ↑ insulin resistance | Hepatic insulin signaling pathway | [119] |
| Human and Male ob/ob mice and wild-type C57BL/6 | ↑ insulin resistance, ↑ FMO3, ↑ FOX01, hyperglycemia | FMO3/TMAO pathway | [111] | |
| Male wild-type C57BL/6 mice | ↑ hepatic FMO3 expression, TMAO activates PERK, ↑ FOX01 | Stress- and PERK-related pathways | [110] | |
| Human and ASO-treated mice | ↑ obesity traits and Type II diabetes, negative regulatory role for FMO3 in beiging of white adipose tissue | TMA/FMO3/TMAO pathway and transintestinal cholesterol excretion (TICE)/intestinal pathway of reverse cholesterol transport | [112] | |
| Human and cholesterol 7 alpha hydroxylase (CYP7A1) mice | ↑ Cyp7a1 in mice, ↑ hepatic lipogenesis, ↑ hepatic steatosis through bile acid metabolism | Bile acid-mediated hepatic FXR signaling pathway | [113] |
6. Potential Treatment Strategies
6.1. Targeting TMA and TMAO Formation Process through Enzymatic Inhibition
6.1.1. Targeting TMAO
6.1.2. Targeting TMA
6.2. Reduction in TMAO Generation by Modifying the Gut Microbiota
6.2.1. Prebiotics and Probiotics
6.2.2. Antibiotics
6.3. Other Therapeutic Alternatives to Lower TMAO Concentration
7. Concluding Remarks and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baselet, B.; Sonveaux, P.; Baatout, S.; Aerts, A. Pathological effects of ionizing radiation: Endothelial activation and dysfunc-tion. Cell. Mol. Life Sci. 2019, 76, 699–728. [Google Scholar] [CrossRef] [PubMed]
- Nappi, F.; Fiore, A.; Masiglat, J.; Cavuoti, T.; Romandini, M.; Nappi, P.; Singh, S.S.A.; Couetil, J.-P. Endothelium-Derived Relaxing Factors and Endothelial Function: A Systematic Review. Biomedicines 2022, 10, 2884. [Google Scholar] [CrossRef] [PubMed]
- Leo, C.H.; Jelinic, M.; Ng, H.H.; Tare, M.; Parry, L.J. Serelaxin: A Novel Therapeutic for Vascular Diseases. Trends Pharmacol. Sci. 2016, 37, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; Leo, C.H.; Girling, J.E.; Tare, M.; Beard, S.; Hannan, N.J.; Parry, L.J. Relaxin treatment reduces angiotensin II-induced vasoconstriction in pregnancy and protects against endothelial dysfunctiondagger. Biol. Reprod. 2017, 96, 895–906. [Google Scholar] [CrossRef]
- Woodman, O.L.; Leo, C.H. Flavonols in the Prevention of Diabetes-induced Vascular Dysfunction. J. Cardiovasc. Pharmacol. 2015, 65, 532–544. [Google Scholar]
- Ng, H.H.; Leo, C.H.; Prakoso, D.; Qin, C.; Ritchie, R.H.; Parry, L.J. Serelaxin treatment reverses vascular dysfunction and left ventricular hypertrophy in a mouse model of Type 1 diabetes. Sci. Rep. 2017, 7, 39604. [Google Scholar] [CrossRef]
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update from the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Subramaniam, S.; Fletcher, C. Trimethylamine N-oxide: Breathe new life. Br. J. Pharmacol. 2018, 175, 1344–1353. [Google Scholar] [CrossRef]
- Kalagi, N.A.; Thota, R.N.; Stojanovski, E.; Alburikan, K.A.; Garg, M.L. Association between Plasma Trimethylamine N-Oxide Levels and Type 2 Diabetes: A Case Control Study. Nutrients 2022, 14, 2093. [Google Scholar] [CrossRef]
- Tang, W.H.; Wang, Z.; Kennedy, D.J.; Wu, Y.; Buffa, J.A.; Agatisa-Boyle, B.; Li, X.S.; Levison, B.S.; Hazen, S.L. Gut micro-biota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mor-tality risk in chronic kidney disease. Circ. Res. 2015, 116, 448–455. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phos-phatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of l-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Senthong, V.; Li, X.S.; Hudec, T.; Coughlin, J.; Wu, Y.; Levison, B.; Wang, Z.; Hazen, S.L.; Tang, W.H. Plasma Trimethylamine N-Oxide, a Gut Microbe-Generated Phosphatidylcholine Metabolite, Is Associated with Atherosclerotic Burden. J. Am. Coll. Cardiol. 2016, 67, 2620–2628. [Google Scholar] [CrossRef]
- Seldin, M.M.; Meng, Y.; Qi, H.; Zhu, W.; Wang, Z.; Hazen, S.L.; Lusis, A.J.; Shih, D.M. Trimethylamine N-Oxide Promotes Vascular Inflammation Through Signaling of Mitogen-Activated Protein Kinase and Nuclear Factor-kappaB. J. Am. Heart Assoc. 2016, 5, e002767. [Google Scholar] [CrossRef]
- Yang, S.; Li, X.; Yang, F.; Zhao, R.; Pan, X.; Liang, J.; Tian, L.; Li, X.; Liu, L.; Xing, Y.; et al. Gut Microbiota-Dependent Marker TMAO in Promoting Cardiovascular Disease: Inflammation Mechanism, Clinical Prognostic, and Potential as a Therapeutic Target. Front. Pharmacol. 2019, 10, 1360. [Google Scholar] [CrossRef]
- Dehghan, P.; Farhangi, M.A.; Nikniaz, L.; Nikniaz, Z.; Asghari-Jafarabadi, M. Gut microbiota-derived metabolite trime-thylamine N-oxide (TMAO) potentially increases the risk of obesity in adults: An exploratory systematic review and dose-response meta- analysis. Obes. Rev. 2020, 21, e12993. [Google Scholar] [CrossRef]
- Romano, K.A.; Vivas, E.I.; Amador-Noguez, D.; Rey, F.E. Intestinal Microbiota Composition Modulates Choline Bioavailability from Diet and Accumulation of the Proatherogenic Metabolite Trimethylamine-N-Oxide. Mbio 2015, 6, e02481. [Google Scholar] [CrossRef]
- Tang, W.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef]
- Hansen, T.H.; Gobel, R.J.; Hansen, T.; Pedersen, O. The gut microbiome in cardio-metabolic health. Genome. Med. 2015, 7, 33. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, M. Trimethylamine N-Oxide Generated by the Gut Microbiota Is Associated with Vascular Inflammation: New Insights into Atherosclerosis. Mediat. Inflamm. 2020, 2020, 4634172. [Google Scholar] [CrossRef]
- Janeiro, M.H.; Ramírez, M.J.; Milagro, F.I.; Martínez, J.A.; Solas, M. Implication of Trimethylamine N-Oxide (TMAO) in Disease: Potential Biomarker or New Therapeutic Target. Nutrients 2018, 10, 1398. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, M.T.; Ramezani, A.; Manal, A.; Raj, D.S. Trimethylamine N-Oxide: The Good, the Bad and the Unknown. Toxins 2016, 8, 326. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Warrier, M. Trimethylamine N-Oxide, the Microbiome, and Heart and Kidney Disease. Annu. Rev. Nutr. 2017, 37, 157–181. [Google Scholar] [CrossRef] [PubMed]
- Zixin, Y.; Lulu, C.; Xiangchang, Z.; Qing, F.; Binjie, Z.; Chunyang, L.; Tai, R.; Dongsheng, O. TMAO as a potential biomarker and therapeutic target for chronic kidney disease: A review. Front. Pharmacol. 2022, 13, 929262. [Google Scholar] [CrossRef] [PubMed]
- Cháfer-Pericás, C.; Herráez-Hernández, R.; Campıns-Falcó, P. Liquid chromatographic determination of trimethylamine in water. J. Chromatogr. 2004, 1023, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, H.Y.; Xue, C.H.; Xin, X.Q.; Xu, J.; Chang, Y.G.; Xue, Y.; Yin, L.A. Simultaneous determination of dimethylamine, trimethylamine and trimethylamine-n-oxide in aquatic products extracts by ion chromatography with non-suppressed con-ductivity detection. J. Chromatogr. 2009, 1216, 5924–5926. [Google Scholar] [CrossRef]
- Veeravalli, S.; Karu, K.; Phillips, I.R.; Shephard, E.A. A highly sensitive liquid chromatography electrospray ionization mass spectrometry method for quantification of TMA, TMAO and creatinine in mouse urine. Methodsx 2017, 4, 310–319. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, Y.; Zhang, X.; Yang, X. A faster and simpler UPLC-MS/MS method for the simultaneous determination of tri-methylamine N-oxide, trimethylamine and dimethylamine in different types of biological samples. Food Funct. 2019, 10, 6484–6491. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Chu, Y.-H.; Wang, C.-C.; Wang, C.-H.; Tain, Y.-L.; Yang, H.-W. Rapid Detection of Gut Microbial Metabolite Trimethylamine N-Oxide for Chronic Kidney Disease Prevention. Biosensors 2021, 11, 339. [Google Scholar] [CrossRef]
- Mitrova, B.; Waffo, A.F.T.; Kaufmann, P.; Iobbi-Nivol, C.; Leimkühler, S.; Wollenberger, U. Trimethylamine N-Oxide Elec-trochemical Biosensor with a Chimeric Enzyme. ChemElectroChem 2018, 6, 1732–1737. [Google Scholar] [CrossRef]
- Lakshmi, G.B.V.S.; Yadav, A.K.; Mehlawat, N.; Jalandra, R.; Solanki, P.R.; Kumar, A. Gut microbiota derived trimethylamine N-oxide (TMAO) detection through molecularly imprinted polymer based sensor. Sci. Rep. 2021, 11, 1338. [Google Scholar] [CrossRef]
- Waffo, A.; Mitrova, B.; Tiedemann, K.; Iobbi-Nivol, C.; Leimkühler, S.; Wollenberger, U. Electrochemical Trimethylamine N-Oxide Biosensor with Enzyme-Based Oxygen-Scavenging Membrane for Long-Term Operation under Ambient Air. Biosensors 2021, 11, 98. [Google Scholar] [CrossRef]
- Yi, Y.; Liang, A.; Luo, L.; Zang, Y.; Zhao, H.; Luo, A. A novel real-time TMAO detection method based on microbial electro-chemical technology. Bioelectrochemistry 2022, 144, 108038. [Google Scholar] [CrossRef]
- Hatton, A.D.; Gibb, S.W. A Technique for the Determination of Trimethylamine-N-oxide in Natural Waters and Biological Media. Anal. Chem. 1999, 71, 4886–4891. [Google Scholar] [CrossRef]
- Mills, G.A.; Walker, V.; Mughal, H. Quantitative determination of trimethylamine in urine by solid-phase microextraction and gas chromatography–mass spectrometry. J. Chromatogr. B: Biomed. Sci. Appl. 1999, 723, 281–285. [Google Scholar] [CrossRef]
- Timm, M.; Jørgensen, B.M. Simultaneous determination of ammonia, dimethylamine, trimethylamine and trimethylamine-n-oxide in fish extracts by capillary electrophoresis with indirect UV-detection. Food Chem. 2002, 76, 509–518. [Google Scholar] [CrossRef]
- Yu, H.; Geng, W.-C.; Zheng, Z.; Gao, J.; Guo, D.-S.; Wang, Y. Facile Fluorescence Monitoring of Gut Microbial Metabolite Trimethylamine N-oxide via Molecular Recognition of Guanidinium-Modified Calixarene. Theranostics 2019, 9, 4624–4632. [Google Scholar] [CrossRef]
- Li, T.; Chen, Y.; Gua, C.; Li, X. Elevated Circulating Trimethylamine N-Oxide Levels Contribute to Endothelial Dysfunction in Aged Rats through Vascular Inflammation and Oxidative Stress. Front. Physiol. 2017, 8, 350. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, W.H.W.; Buffa, J.A.; Fu, X.; Britt, E.B.; Koeth, R.A.; Levison, B.S.; Fan, Y.; Wu, Y.; Hazen, S.L. Prognostic value of choline and betaine depends on intestinal microbiota-generated metabolite trimethylamine-N-oxide. Eur. Heart J. 2014, 35, 904–910. [Google Scholar] [CrossRef]
- Roy, S.; Yuzefpolskaya, M.; Nandakumar, R.; Colombo, P.C.; Demmer, R. Plasma Trimethylamine-N-oxide and impaired glucose regulation: Results from The Oral Infections, Glucose Intolerance and Insulin Resistance Study (ORIGINS). PLoS ONE 2020, 15, e0227482. [Google Scholar] [CrossRef]
- Dong, Z.; Liang, Z.; Guo, M.; Hu, S.; Shen, Z.; Hai, X. The Association between Plasma Levels of Trimethylamine N-Oxide and the Risk of Coronary Heart Disease in Chinese Patients with or without Type 2 Diabetes Mellitus. Dis. Markers 2018, 2018, 1578320. [Google Scholar] [CrossRef] [PubMed]
- Ufnal, M.; Zadlo, A.; Ostaszewski, R. TMAO: A small molecule of great expectations. Nutrition 2015, 31, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Kaysen, G.A.; Johansen, K.L.; Chertow, G.M.; Dalrymple, L.S.; Kornak, J.; Grimes, B.; Dwyer, T.; Chassy, A.W.; Fiehn, O. Associations of Trimethylamine N-Oxide With Nutritional and Inflammatory Biomarkers and Cardiovascular Outcomes in Patients New to Dialysis. J. Ren. Nutr. 2015, 25, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Missailidis, C.; Hällqvist, J.; Qureshi, A.R.; Barany, P.; Heimbürger, O.; Lindholm, B.; Stenvinkel, P.; Bergman, P. Serum Trimethylamine-N-Oxide Is Strongly Related to Renal Function and Predicts Outcome in Chronic Kidney Disease. PLOS ONE 2016, 11, e0141738. [Google Scholar] [CrossRef]
- Wilson, A.; Teft, W.A.; Morse, B.L.; Choi, Y.-H.; Woolsey, S.; DeGorter, M.K.; Hegele, R.A.; Tirona, R.G.; Kim, R.B. Trimethylamine-N-oxide: A Novel Biomarker for the Identification of Inflammatory Bowel Disease. Dig. Dis. Sci. 2015, 60, 3620–3630. [Google Scholar] [CrossRef]
- Fu, Q.; Zhao, M.; Wang, D.; Hu, H.; Guo, C.; Chen, W.; Li, Q.; Zheng, L.; Chen, B. Coronary Plaque Characterization Assessed by Optical Coherence Tomography and Plasma Trimethylamine-N-oxide Levels in Patients With Coronary Artery Disease. Am. J. Cardiol. 2016, 118, 1311–1315. [Google Scholar] [CrossRef]
- Li, J.; Zeng, Q.; Xiong, Z.; Xian, G.; Liu, Z.; Zhan, Q.; Lai, W.; Ao, L.; Meng, X.; Ren, H.; et al. Trimethylamine N-oxide induces osteogenic responses in human aortic valve interstitial cells in vitro and aggravates aortic valve lesions in mice. Cardiovasc. Res. 2021, 118, 2018–2030. [Google Scholar] [CrossRef]
- Wang, J.-M.; Yang, M.-X.; Wu, Q.-F.; Chen, J.; Deng, S.-F.; Chen, L.; Wei, D.-N.; Liang, F.-R. Improvement of intestinal flora: Accompany with the antihypertensive effect of electroacupuncture on stage 1 hypertension. Chin. Med. 2021, 16, 1–11. [Google Scholar] [CrossRef]
- Xiong, X.; Zhou, J.; Fu, Q.; Xu, X.; Wei, S.; Yang, S.; Chen, B. The associations between TMAO-related metabolites and blood lipids and the potential impact of rosuvastatin therapy. Lipids Heal. Dis. 2022, 21, 60. [Google Scholar] [CrossRef]
- Nieuwdorp, M.; Xie, L.; Zhao, B.-X.; Li, X.S.; Qiu, B.; Zhu, F.; Li, G.-F.; He, M.; Wang, Y.; Wang, B.; et al. Serum Trimethylamine N-Oxide Concentration Is Positively Associated With First Stroke in Hypertensive Patients. Stroke 2018, 49, 2021–2028. [Google Scholar] [CrossRef]
- Rexidamu, M.; Li, H.; Jin, H.; Huang, J. Serum levels of Trimethylamine-N-oxide in patients with ischemic stroke. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Yin, J.; Liao, S.X.; He, Y.; Wang, S.; Xia, G.H.; Liu, F.T.; Zhu, J.J.; You, C.; Chen, Q.; Zhou, L.; et al. Dysbiosis of Gut Microbiota With Reduced Trimethylamine-N-Oxide Level in Patients With Large-Artery Atherosclerotic Stroke or Transient Ischemic Attack. J. Am. Hear. Assoc. 2015, 4, e002699. [Google Scholar] [CrossRef]
- Chou, R.-H.; Chen, C.-Y.; Chen, I.-C.; Huang, H.-L.; Lu, Y.-W.; Kuo, C.-S.; Chang, C.-C.; Huang, P.-H.; Chen, J.-W.; Lin, S.-J. Trimethylamine N-Oxide, Circulating Endothelial Progenitor Cells, and Endothelial Function in Patients with Stable Angina. Sci. Rep. 2019, 9, 4249. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, F.; Zhou, Q.; Qiu, Y.; Zhang, J.; Tu, Q.; Zhou, Z.; Shao, Y.; Xu, S.; Wang, Y.; et al. Berberine Improves Vascular Dysfunction by Inhibiting Trimethylamine-N-oxide via Regulating the Gut Microbiota in Angiotensin II-Induced Hypertensive Mice. Front. Microbiol. 2022, 13, 691. [Google Scholar] [CrossRef]
- Chen, L.; Jin, Y.; Wang, N.; Yuan, M.; Lin, T.; Lu, W.; Wang, T. Trimethylamine N-oxide impairs perfusion recovery after hindlimb ischemia. Biochem. Biophys. Res. Commun. 2020, 530, 95–99. [Google Scholar] [CrossRef]
- Ma, G.; Pan, B.; Chen, Y.; Guo, C.; Zhao, M.; Zheng, L.; Chen, B. Trimethylamine N-oxide in atherogenesis: Impairing endo-thelial self-repair capacity and enhancing monocyte adhesion. Biosci. Rep. 2017, 37, BSR20160244. [Google Scholar] [CrossRef]
- Querio, G.; Antoniotti, S.; Geddo, F.; Levi, R.; Gallo, M.P. Trimethylamine N-Oxide (TMAO) Impairs Purinergic Induced In-tracellular Calcium Increase and Nitric Oxide Release in Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 3982. [Google Scholar] [CrossRef]
- Leo, C.H.; Fernando, D.T.; Tran, L.; Ng, H.H.; Marshall, S.A.; Parry, L.J. Serelaxin Treatment Reduces Oxidative Stress and Increases Aldehyde Dehydrogenase-2 to Attenuate Nitrate Tolerance. Front. Pharmacol. 2017, 8, 141. [Google Scholar] [CrossRef]
- Singh, G.B.; Zhang, Y.; Boini, K.M.; Koka, S. High Mobility Group Box 1 Mediates TMAO-Induced Endothelial Dysfunction. Int. J. Mol. Sci. 2019, 20, 3570. [Google Scholar] [CrossRef]
- Sun, X.; Jiao, X.; Ma, Y.; Liu, Y.; Zhang, L.; He, Y.; Chen, Y. Trimethylamine N-oxide induces inflammation and endothelial dysfunction in human umbilical vein endothelial cells via activating ROS-TXNIP-NLRP3 inflammasome. Biochem. Biophys. Res. Commun. 2016, 481, 63–70. [Google Scholar] [CrossRef]
- Chen, M.L.; Zhu, X.H.; Ran, L.; Lang, H.D.; Yi, L.; Mi, M.T. Trimethylamine-N-Oxide Induces Vascular Inflammation by Activating the NLRP3 Inflammasome Through the SIRT3-SOD2-mtROS Signaling Pathway. J. Am. Heart Assoc. 2017, 6, e006347. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Li, D.; Zhao, M.; Liu, C.; Liu, J.; Zeng, A.; Shi, X.; Cheng, S.; Pan, B.; Zheng, L.; et al. Gut flora-dependent metabolite Trimethylamine-N-oxide accelerates endothelial cell senescence and vascular aging through oxidative stress. Free Radic. Biol. Med. 2018, 116, 88–100, Erratum in Free Radic. Biol. Med. 2018, 129, 608–610. [Google Scholar] [CrossRef] [PubMed]
- González-Loyola, A.; Petrova, T.V. Development and aging of the lymphatic vascular system. Adv. Drug Deliv. Rev. 2021, 169, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Brunt, V.E.; LaRocca, T.J.; Bazzoni, A.E.; Sapinsley, Z.J.; Miyamoto-Ditmon, J.; Gioscia-Ryan, R.A.; Neilson, A.P.; Link, C.D.; Seals, D.R. The gut microbiome–derived metabolite trimethylamine N-oxide modulates neuroinflammation and cognitive function with aging. Geroscience 2020, 43, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and in-flammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef]
- Theofilis, P.; Sagris, M.; Oikonomou, E.; Antonopoulos, A.; Siasos, G.; Tsioufis, C.; Tousoulis, D. Inflammatory Mechanisms Contributing to Endothelial Dysfunction. Biomedicines 2021, 9, 781. [Google Scholar] [CrossRef]
- Angelovich, T.A.; Hearps, A.C.; Jaworowski, A. Inflammation-induced foam cell formation in chronic inflammatory disease. Immunol. Cell Biol. 2015, 93, 683–693. [Google Scholar] [CrossRef]
- Pober, J.S.; Sessa, W.C. Inflammation and the blood microvascular system. Cold Spring Harb. Perspect Biol. 2014, 7, a016345. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Boutagy, N.E.; Neilson, A.P.; Osterberg, K.L.; Smithson, A.T.; Englund, T.R.; Davy, B.M.; Hulver, M.W.; Davy, K.P. Pro-biotic supplementation and trimethylamine-N-oxide production following a high-fat diet. Obesity 2015, 23, 2357–2363. [Google Scholar] [CrossRef]
- Meyer, K.A. Population studies of TMAO and its precursors may help elucidate mechanisms. Am. J. Clin. Nutr. 2020, 111, 1115–1116. [Google Scholar] [CrossRef]
- Fu, B.C.; Hullar, M.A.J.; Randolph, T.W.; Franke, A.A.; Monroe, K.R.; Cheng, I.; Wilkens, L.R.; Shepherd, J.A.; Madeleine, M.M.; Le Marchand, L.; et al. Associations of plasma trimethylamine N-oxide, choline, carnitine, and betaine with inflammatory and cardiometabolic risk biomarkers and the fecal microbiome in the Multiethnic Cohort Adiposity Phe-notype Study. Am. J. Clin. Nutr. 2020, 111, 1226–1234. [Google Scholar] [CrossRef]
- Papandreou, C.; More, M.; Bellamine, A. Trimethylamine N-Oxide in Relation to Cardiometabolic Health-Cause or Effect? Nutrients 2020, 12, 1330. [Google Scholar] [CrossRef]
- Liu, H.; Jia, K.; Ren, Z.; Sun, J.; Pan, L.-L. PRMT5 critically mediates TMAO-induced inflammatory response in vascular smooth muscle cells. Cell Death Dis. 2022, 13, 299. [Google Scholar] [CrossRef]
- Durpes, M.C.; Morin, C.; Paquin-Veillet, J.; Beland, R.; Pare, M.; Guimond, M.O.; Rekhter, M.; King, G.L.; Geraldes, P. PKC-beta activation inhibits IL-18-binding protein causing endothelial dysfunction and diabetic atherosclerosis. Cardiovasc. Res. 2015, 106, 303–313. [Google Scholar] [CrossRef]
- Kong, P.; Cui, Z.-Y.; Huang, X.-F.; Zhang, D.-D.; Guo, R.-J.; Han, M. Inflammation and atherosclerosis: Signaling pathways and therapeutic intervention. Signal Transduct. Target. Ther. 2022, 7, 131. [Google Scholar] [CrossRef]
- Gui, Y.; Zheng, H.; Cao, R.Y. Foam Cells in Atherosclerosis: Novel Insights into Its Origins, Consequences, and Molecular Mechanisms. Front. Cardiovasc. Med. 2022, 9, 842. [Google Scholar] [CrossRef]
- Bentzon, J.F.; Otsuka, F.; Virmani, R.; Falk, E. Mechanisms of Plaque Formation and Rupture. Circ. Res. 2014, 114, 1852–1866. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y.; Rimm, E.B.; Hu, F.B.; Albert, C.M.; Rexrode, K.M.; Manson, J.E.; Qi, L. Dietary phosphatidylcholine and risk of all-cause and cardiovascular-specific mortality among US women and men. Am. J. Clin. Nutr. 2016, 104, 173–180. [Google Scholar] [CrossRef]
- Geng, J.; Yang, C.; Wang, B.; Zhang, X.; Hu, T.; Gu, Y.; Li, J. Trimethylamine N-oxide promotes atherosclerosis via CD36-dependent MAPK/JNK pathway. Biomed. Pharmacother. 2018, 97, 941–947. [Google Scholar] [CrossRef]
- Herrema, H.; Niess, J.H. Intestinal microbial metabolites in human metabolism and type 2 diabetes. Diabetologia 2020, 63, 2533–2547. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.M.; Zhu, W.; Schugar, R.C.; Meng, Y.; Jia, X.; Miikeda, A.; Wang, Z.; Zieger, M.; Lee, R.; Graham, M.; et al. Genetic Deficiency of Flavin-Containing Monooxygenase 3 (Fmo3) Protects Against Thrombosis but Has Only a Minor Effect on Plasma Lipid Levels—Brief Report. Arter. Thromb. Vasc. Biol. 2019, 39, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Xue, J.; Shan, J.; Hong, Y.; Zhu, W.; Nie, Z.; Zhang, Y.; Ji, N.; Luo, X.; Zhang, T.; et al. Gut-Flora-Dependent Me-tabolite Trimethylamine-N-Oxide Promotes Atherosclerosis-Associated Inflammation Responses by Indirect ROS Stimulation and Signaling Involving AMPK and SIRT1. Nutrients 2022, 14, 3338. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Kojima, M.; Takayanagi, K.; Taguchi, K.; Kobayashi, T. Role of S-Equol, Indoxyl Sulfate, and Trimethylamine N-Oxide on Vascular Function. Am. J. Hypertens. 2020, 33, 793–803. [Google Scholar] [CrossRef]
- Leo, C.H.; Jelinic, M.; Ng, H.H.; Tare, M.; Parry, L.J. Time-dependent activation of prostacyclin and nitric oxide pathways during continuous i.v. infusion of serelaxin (recombinant human H2 relaxin). Br. J. Pharmacol. 2016, 173, 1005–1017. [Google Scholar] [CrossRef]
- Leo, C.; Hart, J.; Woodman, O. Impairment of both nitric oxide-mediated and EDHF-type relaxation in small mesenteric arteries from rats with streptozotocin-induced diabetes. Br. J. Pharmacol. 2011, 162, 365–377. [Google Scholar] [CrossRef]
- Leo, C.H.; Jelinic, M.; Parkington, H.C.; Tare, M.; Parry, L.J. Acute Intravenous Injection of Serelaxin (Recombinant Human Relaxin-2) Causes Rapid and Sustained Bradykinin-Mediated Vasorelaxation. J. Am. Heart Assoc. 2014, 3, e000493. [Google Scholar] [CrossRef]
- Leo, C.H.; Jelinic, M.; Ng, H.H.; Marshall, S.A.; Novak, J.; Tare, M.; Conrad, K.P.; Parry, L.J. Vascular actions of relaxin: Nitric oxide and beyond. Br. J. Pharmacol. 2017, 174, 1002–1014. [Google Scholar] [CrossRef]
- Ng, H.H.; Jelinic, M.; Parry, L.J.; Leo, C.-H. Increased superoxide production and altered nitric oxide-mediated relaxation in the aorta of young but not old male relaxin-deficient mice. Am. J. Physiol. Circ. Physiol. 2015, 309, H285–H296. [Google Scholar] [CrossRef]
- Leo, C.-H.; Hart, J.L.; Woodman, O.L. 3′,4′-Dihydroxyflavonol Reduces Superoxide and Improves Nitric Oxide Function in Diabetic Rat Mesenteric Arteries. PLoS ONE 2011, 6, e20813. [Google Scholar] [CrossRef]
- Jelinic, M.; Kahlberg, N.; Leo, C.H.; Ng, H.H.; Rosli, S.; Deo, M.; Li, M.; Finlayson, S.; Walsh, J.; Parry, L.J.; et al. Annexin-A1 deficiency exacerabates pathological remodelling of the mesenteric vasculature in insulin-resistance but not insulin-deficiency. Br. J. Pharmacol. 2020, 177, 1677–1691. [Google Scholar] [CrossRef]
- Kahlberg, N.; Qin, C.X.; Anthonisz, J.; Jap, E.; Ng, H.H.; Jelinic, M.; Parry, L.J.; Kemp-Harper, B.K.; Ritchie, R.H.; Leo, C.H. Adverse vascular remodelling is more sensitive than endothelial dysfunction to hyperglycaemia in diabetic rat mesenteric ar-teries. Pharmacol. Res. 2016, 111, 325–335. [Google Scholar] [CrossRef]
- Jelinic, M.; Jackson, K.L.; O’Sullivan, K.; Singh, J.; Giddy, T.; Deo, M.; Parry, L.J.; Ritchie, R.H.; Woodman, O.L.; Head, G.A.; et al. Endothelium-dependent relaxation is impaired in Schlager hypertensive (BPH/2J) mice by region-specific mechanisms in conductance and resistance arteries. Life Sci. 2023, 320, 121542. [Google Scholar] [CrossRef]
- Li, T.; Gua, C.; Wu, B.; Chen, Y. Increased circulating trimethylamine N-oxide contributes to endothelial dysfunction in a rat model of chronic kidney disease. Biochem. Biophys. Res. Commun. 2018, 495, 2071–2077. [Google Scholar] [CrossRef]
- Pan, L.-L.; Qin, M.; Liu, X.-H.; Zhu, Y.-Z. The Role of Hydrogen Sulfide on Cardiovascular Homeostasis: An Overview with Update on Immunomodulation. Front. Pharmacol. 2017, 8, 686. [Google Scholar] [CrossRef]
- Bai, L.; Dai, J.; Xia, Y.; He, K.; Xue, H.; Guo, Q.; Tian, D.; Xiao, L.; Zhang, X.; Teng, X.; et al. Hydrogen Sulfide Ame-liorated High Choline-Induced Cardiac Dysfunction by Inhibiting cGAS-STING-NLRP3 Inflammasome Pathway. Oxid. Med. Cell Longev. 2022, 2022, 1392896. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, Y.; Ren, D.; Yang, X. Protective Effect of Saponins-Enriched Fraction of Gynostemma pentaphyllum against High Choline-Induced Vas-cular Endothelial Dysfunction and Hepatic Damage in Mice. Biol. Pharm. Bull. 2020, 43, 463–473. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kojima, M.; Takayanagi, K.; Taguchi, K.; Kobayashi, T. Trimethylamine-N-oxide Specifically Impairs Endothelium-Derived Hyperpolarizing Factor-Type Relaxation in Rat Femoral Artery. Biol. Pharm. Bull. 2020, 43, 569–573. [Google Scholar] [CrossRef]
- Marquardt, L.; Ruf, A.; Mansmann, U.; Winter, R.; Schuler, M.; Buggle, F.; Mayer, H.; Grau, A.J. Course of Platelet Activation Markers After Ischemic Stroke. Stroke 2002, 33, 2570–2574. [Google Scholar] [CrossRef]
- Kaur, R.; Kaur, M.; Singh, J. Endothelial dysfunction and platelet hyperactivity in type 2 diabetes mellitus: Molecular insights and therapeutic strategies. Cardiovasc. Diabetol. 2018, 17, 121. [Google Scholar] [CrossRef]
- Varga-Szabo, D.; Braun, A.; Nieswandt, B. Calcium signaling in platelets. J. Thromb. Haemost. 2009, 7, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Dean, W.L. Role of platelet plasma membrane Ca2+-ATPase in health and disease. World J. Biol. Chem. 2010, 1, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Adam, F.; Kauskot, A.; Rosa, J.; Bryckaert, M. Mitogen-activated protein kinases in hemostasis and thrombosis. J. Thromb. Haemost. 2008, 6, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Liu, X.; Huang, X.; Liu, Q.; Yang, M.; Huang, D.; Zhao, P.; Tian, J.; Wang, X.; Hou, J. Remodelling of gut microbiota by Berberine attenuates trimethylamine N-oxide-induced platelet hyperreaction and thrombus formation. Eur. J. Pharmacol. 2021, 911, 174526. [Google Scholar] [CrossRef]
- Herrera, J.; Henke, C.A.; Bitterman, P. Extracellular matrix as a driver of progressive fibrosis. J. Clin. Investig. 2018, 128, 45–53. [Google Scholar] [CrossRef]
- Li, X.; Geng, J.; Zhao, J.; Ni, Q.; Zhao, C.; Zheng, Y.; Chen, X.; Wang, L. Trimethylamine N-Oxide Exacerbates Cardiac Fibrosis via Activating the NLRP3 Inflammasome. Front. Physiol. 2019, 10, 866. [Google Scholar] [CrossRef]
- Makrecka-Kuka, M.; Volska, K.; Antone, U.; Vilskersts, R.; Grinberga, S.; Bandere, D.; Liepinsh, E.; Dambrova, M. Trime-thylamine N-oxide impairs pyruvate and fatty acid oxidation in cardiac mitochondria. Toxicol. Lett. 2017, 267, 32–38. [Google Scholar] [CrossRef]
- Guo, S. Insulin signaling, resistance, and the metabolic syndrome: Insights from mouse models into disease mechanisms. J. Endocrinol. 2014, 220, T1–T23. [Google Scholar] [CrossRef]
- Chen, P.Y.; Qin, L.; Li, G.; Wang, Z.; Dahlman, J.E.; Malagon-Lopez, J.; Gujja, S.; Cilfone, N.A.; Kauffman, K.J.; Sun, L.; et al. Endothelial TGF-beta signalling drives vascular inflammation and atherosclerosis. Nat. Metab. 2019, 1, 912–926. [Google Scholar] [CrossRef]
- Miao, J.; Ling, A.V.; Manthena, P.V.; Gearing, M.E.; Graham, M.J.; Crooke, R.M.; Croce, K.J.; Esquejo, R.M.; Clish, C.B.; Morbid Obesity Study, G.; et al. Flavin-containing monooxygenase 3 as a potential player in diabe-tes-associated atherosclerosis. Nat. Commun. 2015, 6, 6498. [Google Scholar] [CrossRef]
- Schugar, R.C.; Shih, D.M.; Warrier, M.; Helsley, R.N.; Burrows, A.; Ferguson, D.; Brown, A.L.; Gromovsky, A.D.; Heine, M.; Chatterjee, A.; et al. The TMAO-Producing Enzyme Flavin-Containing Monooxygenase 3 Regulates Obesity and the Beiging of White Adipose Tissue. Cell Rep. 2017, 19, 2451–2461. [Google Scholar] [CrossRef]
- Tan, X.; Liu, Y.; Long, J.; Chen, S.; Liao, G.; Wu, S.; Li, C.; Wang, L.; Ling, W.; Zhu, H. Trimethylamine N-Oxide Aggravates Liver Steatosis through Modulation of Bile Acid Metabolism and Inhibition of Farnesoid X Receptor Signaling in Nonalcoholic Fatty Liver Disease. Mol. Nutr. Food Res. 2019, 63, e1900257. [Google Scholar] [CrossRef]
- Koeth, R.A.; Levison, B.S.; Culley, M.K.; Buffa, J.A.; Wang, Z.; Gregory, J.C.; Org, E.; Wu, Y.; Li, L.; Smith, J.D.; et al. γ-Butyrobetaine Is a Proatherogenic Intermediate in Gut Microbial Metabolism of L-Carnitine to TMAO. Cell Metab. 2014, 20, 799–812. [Google Scholar] [CrossRef]
- Wang, Z.; Roberts, A.B.; Buffa, J.A.; Levison, B.S.; Zhu, W.; Org, E.; Gu, X.; Huang, Y.; Zamanian-Daryoush, M.; Culley, M.K.; et al. Non-lethal Inhibition of Gut Microbial Trimethylamine Production for the Treatment of Atherosclerosis. Cell 2015, 163, 1585–1595. [Google Scholar] [CrossRef]
- Li, Z.; Wu, Z.; Yan, J.; Liu, H.; Liu, Q.; Deng, Y.; Ou, C.; Chen, M. Gut microbe-derived metabolite trimethylamine N-oxide induces cardiac hypertrophy and fibrosis. Lab. Investig. 2019, 99, 346–357. [Google Scholar] [CrossRef]
- Fang, Q.; Zheng, B.; Liu, N.; Liu, J.; Liu, W.; Huang, X.; Zeng, X.; Chen, L.; Li, Z.; Ouyang, D. Trimethylamine N-Oxide Exacerbates Renal Inflammation and Fibrosis in Rats With Diabetic Kidney Disease. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef]
- El-Deeb, O.S.; Atef, M.M.; Hafez, Y.M. The interplay between microbiota-dependent metabolite trimethylamine N -oxide, Transforming growth factor β /SMAD signaling and inflammasome activation in chronic kidney disease patients: A new mechanistic perspective. J. Cell. Biochem. 2019, 120, 14476–14485. [Google Scholar] [CrossRef]
- Gao, X.; Liu, X.; Xu, J.; Xue, C.; Xue, Y.; Wang, Y. Dietary trimethylamine N-oxide exacerbates impaired glucose tolerance in mice fed a high fat diet. J. Biosci. Bioeng. 2014, 118, 476–481. [Google Scholar] [CrossRef]
- Nagatomo, Y.; Tang, W.H.W. Intersections Between Microbiome and Heart Failure: Revisiting the Gut Hypothesis. J. Card. Fail. 2015, 21, 973–980. [Google Scholar] [CrossRef]
- Conraads, V.M.; Jorens, P.G.; De Clerck, L.S.; Van Saene, H.K.; Ieven, M.M.; Bosmans, J.M.; Schuerwegh, A.; Bridts, C.H.; Wuyts, F.; Stevens, W.J.; et al. Selective intestinal decontamination in advanced chronic heart failure: A pilot trial. Eur. J. Heart Fail. 2004, 6, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation of Gut Microbiota Contributes to Alleviation of Both Genetic and Simple Obesity in Children. Ebiomedicine 2015, 2, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.P.; Wang, Y.; Sprenger, N.; Yap, I.K.; Lundstedt, T.; Lek, P.; Rezzi, S.; Ramadan, Z.; van Bladeren, P.; Fay, L.B.; et al. Probiotic modulation of symbiotic gut microbial-host metabolic inter-actions in a humanized microbiome mouse model. Mol. Syst. Biol. 2008, 4, 157. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular Risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zheng, X.; Feng, M.; Li, D.; Zhang, H. Gut Microbiota-Dependent Metabolite Trimethylamine N-Oxide Contributes to Cardiac Dysfunction in Western Diet-Induced Obese Mice. Front. Physiol. 2017, 8, 139. [Google Scholar] [CrossRef]
- Dambrova, M.; Skapare-Makarova, E.; Konrade, I.; Pugovics, O.; Grinberga, S.; Tirzite, D.; Petrovska, R.; Kalvins, I.; Liepins, E. Meldonium decreases the diet-increased plasma levels of trimethylamine N-oxide, a metabolite associated with atherosclerosis. J. Clin. Pharmacol. 2013, 53, 1095–1098. [Google Scholar] [CrossRef]
- Kuka, J.; Liepinsh, E.; Makrecka-Kuka, M.; Liepins, J.; Cirule, H.; Gustina, D.; Loza, E.; Zharkova-Malkova, O.; Grinberga, S.; Pugovics, O.; et al. Suppression of intestinal microbiota-dependent production of pro-atherogenic trimethylamine N-oxide by shifting L-carnitine microbial degradation. Life Sci. 2014, 117, 84–92. [Google Scholar] [CrossRef]
- Chen, M.-l.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.-D.; Zhang, Q.-Y.; Mi, M.-T.; Rey, F.; et al. Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabo-lism via Remodeling of the Gut Microbiota. MBio 2016, 7, e02210-15. [Google Scholar] [CrossRef]
- Wu, T.; Gao, Y.; Hao, J.; Geng, J.; Zhang, J.; Yin, J.; Liu, R.; Sui, W.; Gong, L.; Zhang, M. Capsanthin extract prevents obesity, reduces serum TMAO levels and modulates the gut microbiota composition in high-fat-diet induced obese C57BL/6J mice. Food Res. Int. 2020, 128, 108774. [Google Scholar] [CrossRef]
- Wu, T.; Gao, Y.; Hao, J.; Yin, J.; Li, W.; Geng, J.; Liu, R.; Sui, W.; Zhang, M. Lycopene, amaranth, and sorghum red pigments counteract obesity and modulate the gut microbiota in high-fat diet fed C57BL/6 mice. J. Func. Foods 2019, 60, 103437. [Google Scholar] [CrossRef]
- Wang, M.; Wang, F.; Wang, Y.; Ma, X.; Zhao, M.; Zhao, C. Metabonomics Study of the Therapeutic Mechanism of Gynostemma pentaphyllum and Atorvastatin for Hyperlipidemia in Rats. PLoS ONE 2013, 8, e78731. [Google Scholar] [CrossRef]
- Sun, B.; Wang, X.; Cao, R.; Zhang, Q.; Liu, Q.; Xu, M.; Zhang, M.; Du, X.; Dong, F.; Yan, X. NMR-based metabonomics study on the effect of Gancao in the attenuation of toxicity in rats induced by Fuzi. J. Ethnopharmacol. 2016, 193, 617–626. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Li, S.; Koh, Y.-C.; Wu, J.-C.; Yang, M.-J.; Ho, C.-T.; Pan, M.-H. Oolong Tea Extract and Citrus Peel Polymethoxyflavones Reduce Transformation of l-Carnitine to Trimethylamine-N-Oxide and Decrease Vascular Inflammation in l-Carnitine Feeding Mice. J. Agric. Food Chem. 2019, 67, 7869–7879. [Google Scholar] [CrossRef]
- Shi, Y.; Hu, J.; Geng, J.; Hu, T.; Wang, B.; Yan, W.; Jiang, Y.; Li, J.; Liu, S. Berberine treatment reduces atherosclerosis by me-diating gut microbiota in apoE-/- mice. Biomed. Pharmacother. 2018, 107, 1556–1563. [Google Scholar] [CrossRef]
- Anwar, S.; Bhandari, U.; Panda, B.P.; Dubey, K.; Khan, W.; Ahmad, S. Trigonelline inhibits intestinal microbial metabolism of choline and its associated cardiovascular risk. J. Pharm. Biomed. Anal. 2018, 159, 100–112. [Google Scholar] [CrossRef]
- Konop, M.; Radkowski, M.; Grochowska, M.; Perlejewski, K.; Samborowska, E.; Ufnal, M. Enalapril decreases rat plasma concentration of TMAO, a gut bacteria-derived cardiovascular marker. Biomarkers 2018, 23, 380–385. [Google Scholar] [CrossRef]
- Kuka, J.; Videja, M.; Makrecka-Kuka, M.; Liepins, J.; Grinberga, S.; Sevostjanovs, E.; Vilks, K.; Liepinsh, E.; Dambrova, M. Metformin decreases bacterial trimethylamine production and trimethylamine N-oxide levels in db/db mice. Sci. Rep. 2020, 10, 14555. [Google Scholar] [CrossRef]
- Shih, D.M.; Wang, Z.; Lee, R.; Meng, Y.; Che, N.; Charugundla, S.; Qi, H.; Wu, J.; Pan, C.; Brown, J.M.; et al. Flavin containing monooxygenase 3 exerts broad effects on glucose and lipid metab-olism and atherosclerosis. J. Lipid. Res. 2015, 56, 22–37. [Google Scholar] [CrossRef]
- Warrier, M.; Shih, D.M.; Burrows, A.C.; Ferguson, D.; Gromovsky, A.D.; Brown, A.L.; Marshall, S.; McDaniel, A.; Schugar, R.C.; Wang, Z.; et al. The TMAO-Generating Enzyme Flavin Monooxygenase 3 Is a Central Regulator of Cholesterol Balance. Cell Rep. 2015, 10, 326–338. [Google Scholar] [CrossRef]
- Vilskersts, R.; Zharkova-Malkova, O.; Mezhapuke, R.; Grinberga, S.; Cirule, H.; Dambrova, M. Elevated vascular gam-ma-butyrobetaine levels attenuate the development of high glucose-induced endothelial dysfunction. Clin. Exp. Pharmacol. Physiol. 2013, 40, 518–524. [Google Scholar] [CrossRef]
- Ryan, P.M.; Stanton, C.; Caplice, N.M. Bile acids at the cross-roads of gut microbiome–host cardiometabolic interactions. Diabetol. Metab. Syndr. 2017, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics con-sensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, H. The Role of Gut Microbiota in Atherosclerosis and Hypertension. Front. Pharmacol. 2018, 9, 1082. [Google Scholar] [CrossRef]
- Brugere, J.F.; Borrel, G.; Gaci, N.; Tottey, W.; O’Toole, P.W.; Malpuech-Brugere, C. Archaebiotics: Proposed therapeutic use of archaea to prevent trimethylaminuria and cardiovascular disease. Gut Microbes. 2014, 5, 5–10. [Google Scholar] [CrossRef]
- Dridi, B. Laboratory tools for detection of archaea in humans. Clin. Microbiol. Infect. 2012, 18, 825–833. [Google Scholar] [CrossRef]
- Pourrajab, B.; Fatahi, S.; Dehnad, A.; Kord Varkaneh, H.; Shidfar, F. The impact of probiotic yogurt consumption on lipid profiles in subjects with mild to moderate hypercholesterolemia: A systematic review and meta-analysis of randomized con-trolled trials. Nutr. Metab. Cardiovasc Dis. 2020, 30, 11–22. [Google Scholar] [CrossRef]
- Yousefi, B.; Eslami, M.; Ghasemian, A.; Kokhaei, P.; Salek Farrokhi, A.; Darabi, N. Probiotics importance and their immuno-modulatory properties. J. Cell. Physiol. 2019, 234, 8008–8018. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef]
- Li, X.; Hong, J.; Wang, Y.; Pei, M.; Wang, L.; Gong, Z. Trimethylamine-N-Oxide Pathway: A Potential Target for the Treatment of MAFLD. Front. Mol. Biosci. 2021, 8, 733507. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, Q.; Jiang, H. Gut microbiota in atherosclerosis: Focus on trimethylamine N-oxide. Apmis 2020, 128, 353–366. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Yamamoto, H.; Okamoto, A.; Imanishi, K.; Tokui, N.; Okamoto, T.; Suzuki, Y.; Sugiyama, N.; Imai, A.; Kudo, S.; et al. Effect of an Oral Adsorbent, AST-120, on Dialysis Initiation and Survival in Patients with Chronic Kidney Disease. Int. J. Nephrol. 2012, 2012, 376128. [Google Scholar] [CrossRef]
- Bird, J.K.; Raederstorff, D.; Weber, P.; Steinert, R.E. Cardiovascular and Antiobesity Effects of Resveratrol Mediated through the Gut Microbiota. Adv. Nutr. Int. Rev. J. 2017, 8, 839–849. [Google Scholar] [CrossRef]
- Chen, L.; He, W.; Peng, B.; Yuan, M.; Wang, N.; Wang, J.; Lu, W.; Wang, T. Sodium Tanshinone IIA sulfonate improves post-ischemic angiogenesis in hyperglycemia. Biochem. Biophys. Res. Commun. 2019, 520, 580–585. [Google Scholar] [CrossRef]
- Kant, R.; Chandra, L.; Verma, V.; Nain, P.; Bello, D.; Patel, S.; Ala, S.; Chandra, R.; Antony, M.A. Gut microbiota interactions with anti-diabetic medications and pathogenesis of type 2 diabetes mellitus. World J. Methodol. 2022, 12, 246–257. [Google Scholar] [CrossRef]
- Marshall, S.A.; Qin, C.X.; Jelinic, M.; O’Sullivan, K.; Deo, M.; Walsh, J.; Li, M.; Parry, L.J.; Ritchie, R.H.; Leo, C.H. The Novel Small-molecule Annexin-A1 Mimetic, Compound 17b, Elicits Vasoprotective Actions in Streptozotocin-induced Diabetic Mice. Int. J. Mol. Sci. 2020, 21, 1384. [Google Scholar] [CrossRef]
- Leo, C.H.; Jelinic, M.; Ng, H.H.; Parry, L.J.; Tare, M. Recent developments in relaxin mimetics as therapeutics for cardio-vascular diseases. Curr. Opin. Pharmacol. 2019, 45, 42–48. [Google Scholar] [CrossRef]
- Ng, H.H.; Leo, C.H.; Parry, L.J.; Ritchie, R.H. Relaxin as a Therapeutic Target for the Cardiovascular Complications of Diabetes. Front. Pharmacol. 2018, 9, 501. [Google Scholar] [CrossRef]
- Ong, E.S.; Low, J.; Tan, J.C.W.; Foo, S.Y.; Leo, C.H. Valorization of avocado seeds with antioxidant capacity using pressurized hot water extraction. Sci. Rep. 2022, 12, 13036. [Google Scholar] [CrossRef]
- Leo, C.H.; Foo, S.Y.; Tan, J.C.W.; Tan, U.-X.; Chua, C.K.; Ong, E.S. Green Extraction of Orange Peel Waste Reduces TNFα-Induced Vascular Inflammation and Endothelial Dysfunction. Antioxidants 2022, 11, 1768. [Google Scholar] [CrossRef]
- Ong, E.; Oh, C.; Tan, J.; Foo, S.; Leo, C. Pressurized Hot Water Extraction of Okra Seeds Reveals Antioxidant, Antidiabetic and Vasoprotective Activities. Plants 2021, 10, 1645. [Google Scholar] [CrossRef] [PubMed]


| Experimental Method | Technique | Linear Range | Limit of Detection | Sensitivity | Response Time | Advantage | Limitations | References |
|---|---|---|---|---|---|---|---|---|
| Chromatography | FIGD-IC | 40–600 nmol/dm3 | 1.35 nmol/dm3 | - | 20 min | Non-hazardous, reliable, precise (3%), sensitive | Time-consuming, expensive equipment, requires specialized technicians | [34] |
| GC-MS | SPME | 14.9–956 μmol/L | 0.01 μmol/L | 14.9 μmol/L | - | Analysis of volatile and semi-volatile compounds | Complicated, laborious, time-consuming, incomplete TMAO transformation | [35] |
| Electrophoresis | Capillary electrophoresis with indirect UV-detection | 0.025–2.5 mmol/L | 2.5 mmol/L | - | - | Analytical precision, repeatability | Time-consuming, expensive equipment, specialized technicians, restrict point-of-care testing (PCOT) | [36] |
| Liquid chromatography | SPE | 5.0–50.0 μg/mL | 0.05 μg | - | - | Selective determination in presence of other primary and secondary short chain aliphatic amines | - | [25] |
| Chromatography | Ion chromatography | 1.0–20.0 mg/mL | 0.10 mg/L | - | 16 min | Inexpensive and stable | Time-consuming, requires specialized technicians | [26] |
| Chromatography | LC-SIMs | 15–944 pg/μL | 115 pg/mL | - | 5 min | Robust, highly sensitive, reproducible, no sample pre-treatment required, only small volume of sample needed | Expensive | [27] |
| Chromatography | UPLC-M/MS | 15–1500 μg/L | 0.12 μg/L | - | 6 min | Repeatable, rapid, and economic | Not a point-of-care testing | [28] |
| Fluorescence | IDA | 0–1.22 mmol/L | 8.98 μmol/L | - | - | Low cost, easy to operate, label free, sensitive | - | [37] |
| Electrochemical | CV | 2–110 µmol/L | 2.96 nmol/L | 14.16 nA/mM | 16 s | Sensor can operate over prolonged daily measurements, quite good short-term usage stability | Complex preparation process (enzyme purification and protein reconstruction) | [30] |
| Electrochemical | DPV | 1–15 ppm | 1.5 ppm | 2.47 µA mL/ppm/cm2 | 20 min | Easy to construct and operate, highly selective | - | [31] |
| Electrochemical | Oxygen anti-interference membrane | 2 µM–15 mmol/L | 10 µmol/L | 2.75 µA/mM | 33 s | Operational stability over 3 weeks | Vulnerable to environmental interferences in clinical applications | [32] |
| Electrochemical | Microbial electrochemical technology | 0–250 µmol/L | 5.96 µmol/L | 23.92 µA/mM | 600 s | 90% accuracy in real serum, high feasibility in clinical applications | - | [33] |
| Experimental Model | Condition (If Any) | Plasma TMAO Level for Control | Plasma TMAO Level for Condition | References |
|---|---|---|---|---|
| Human and Rodents | - | 0.5–5 µmol/L | - | [11,42] |
| Human | Patients undergoing hemodialysis | 0.92–1.9 µmol/L | 28–67 µmol/L | [43] |
| Chronic Kidney Disease | - | 32.2–75.2 µmol/L | [44] | |
| Inflammatory Bowel Disease | - | 2.27 µmol/L | [45] | |
| Plaque rupture | 4.2 ± 2.4 μmol/L | 8.6 ± 4.8 µmol/L | [46] | |
| Calcified aortic valve disease | 1.4–2.8 µmol/L | 2.3–6.4 µmol/L | [47] | |
| Stage 1 hypertensive patients | - | 87.2 ng/mL | [48] | |
| Older age, BMI, lower eGFR, HDL-levels, higher choline and carnitine levels, higher TG | 2.83 ± 1.34 μmol/L | 8.43 ± 4.85 µmol/L | [49] | |
| Stroke | 1.4–3.7 µmol/L | 1.6–4.0 µmol/L | [50] | |
| First ever acute ischemic stroke and neurological deficit | 2.6–6.1 µmol/L | 0.5–18.3 µmol/L | [51] | |
| Stroke (LAA), transient ischemic attack, history of diabetes, CAD, HBP, HLP | 1.91 µmol/L | 2.70 µmol/L | [52] |
| Intervention | Therapy | Model | Intervention/Dosage | Duration | Route of Administration | Effects | Limitations | Reference |
|---|---|---|---|---|---|---|---|---|
| Targeting the gut microbiome | Prebiotics | Human | Whole grains, traditional Chinese medicinal foods, and prebiotics (WTP diet) | 30–90 days | Oral | Improves composition of gut microbiota to reduce TMAO formation | Gut microbiome is influenced by multiple components | [122] |
| Probiotics | Female germ-free mouse (C3H strain) | Basal mixed diet and probiotics supplementation in saline water | 14 days | Oral | Lowers TMAO formation in the gut | Unclear safety and effects in humans | [123] | |
| Antibiotics | Mouse | Drinking water with a cocktail of broad-spectrum antibiotics | 21 days | Oral | Suppression and inhibition of plasma TMAO levels | Inhibition of useful bacteria and induction of resistant bacteria. Not feasible in the long run | [11] | |
| Antibiotics | Human | Metronidazole (500 mg twice daily) plus ciprofloxacin (500 mg once daily) for 1 week | 7 days | Oral | Suppression and inhibition of plasma TMAO levels | Inhibition of useful bacteria and induction of resistant bacteria. Not feasible in the long run | [124] | |
| Oral non-absorbent binders | Human and rat | 10 mL solution of 800 mg of polymyxin B and 320 mg of tobramycin (selective decontamination of the digestive tract [SDD]) | 56 days | Oral | Removal of TMAO and its precursors from gut | Uncertain approach. Compound specific to TMAO has not yet been found | [121] | |
| Targeting TMA formation | Inhibition of FMO3 | Wild-type C57BL/6J, male Sprague–Dawley rat (Harlan) and human | 50 mg/kg body weight of antisense oligonucleotides (ASO) | 7 weeks or 16 weeks | Intraperitoneal injection | Inhibition of TMAO formation from TMA | Accumulation of TMA in plasma may cause other diseases. Metabolism of other compounds is also mediated by FMO3 | [111] |
| 3,3-Dimethyldimethyl-1-butanol (DMB) | Mouse | 1% DMB in drinking water | 56 days | Oral | Inhibition of TMA formation from dietary precursors choline, carnitine, corotonobetaine by inhibiting microbial TMA lyase | Complete TMAO formation cannot be avoided by DMB. Study not performed in humans. Unable to inhibit formation of TMA from γ-butyrobetaine | [125] | |
| 3,3-Dimethyldimethyl-1-butanol (DMB) | Mouse | 1%, v/v in drinking water | 16 weeks | Oral | Reorganization of gut microbial community and inhibition of TMA production | Only partial TMAO formation inhibition | [115] | |
| Meldonium | Human and rat | Single dose of 13C-GBB (100 mg/kg) or 13C-GBB in combination with meldonium (GBB + M, 100 mg/kg each) | 14 days | Oral | Lowers TMAO formation from L-carnitine and increases TMAO excretion via urine | Unable to reduce TMAO formation from choline | [126,127] | |
| Therapeutic alternatives to lower TMAO concentration | Resveratrol | Female C57BL/6J mouse and ApoE-/- mouse with a C57BL/6 genetic background | 0.4% RSV | 30 days | Oral | Alters gut microbiome composition, hence reducing bacteria that forms TMA and increasing useful bacteria | Study performed only in mice. No changes when antibiotics are used | [128] |
| Capsanthin | High-fat-diet induced obese C57BL/6J mice | Capsanthin at 200 mg kg−1 body weight | 12 weeks | Oral | Lowers body weight, effectively reduces TMAO levels, and increases microbial diversity | Study performed in mice | [129] | |
| Lycopene (Lycopersicon esculentum [M.]), amaranth (Amaranthus tricolor), and sorghum red (Sorghum bicolor (L.)) pigments | High-fat diet fed C57BL/6 mice | 200 mg/kg body of lycopene or amaranth or sorghum red administration | 12 weeks | Oral | Ameliorates lipid metabolism, and lowers TMAO levels | Study performed in mice | [130] | |
| Gynostemma pentaphyllum | Rat | 120 mg/kg/day | 28 days | Oral gavage | Lowers plasma TMAO levels and rises in lecithin levels | Study performed only in rats | [131] | |
| Gancao (root of glycyrrhiza uralensis) | Male Wistar rat | Single dose of Gancao (35.6 g kg−1 body weight) | - | Intragastric administration | Prevents increase in TMAO when administered with Fuzi (processed lateral root of Aconitum carmichaelii) | Does not lower TMAO levels when administered alone. Study performed in rats | [132] | |
| Oolong tea extract and citrus peel polymethoxyflavones | Mouse | 1 μg | Injection every 10 days for a period of 16 weeks | Intravenous injection | Lowers TMAO formation and vascular inflammation | Study performed only in mice | [133] | |
| Berberine (BBR) | ApoE-/- mouse on a C57BL/6 background | BBR treatment (50 mg/kg) twice weekly | 84 days | Intragastric administration | Lowers expression of hepatic FMO3 and serum TMAO levels | Study performed only in mice | [134] | |
| Trigonelline | C57BL/6 J mouse | Trigonelline (50 to 100 mg/kg) per day | 14 days | Oral | Inhibits conversion of TMA to TMAO by inhibiting FMO3 | Study performed only in mice | [135] | |
| Enalapril | Rat | 5.3 or 12.6 mg/kg | 14 days | Oral | Increases TMAO excretion in urine | Unclear mechanism. Does not affect TMA formation or composition of gut microbiota | [21,136] | |
| Metformin | db/db mice | 250 mg/kg/day | 8 weeks | Oral | 2-fold reduction in TMAO levels and bacteria linked to TMAO production | Study performed in only mice | [137] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shanmugham, M.; Bellanger, S.; Leo, C.H. Gut-Derived Metabolite, Trimethylamine-N-oxide (TMAO) in Cardio-Metabolic Diseases: Detection, Mechanism, and Potential Therapeutics. Pharmaceuticals 2023, 16, 504. https://doi.org/10.3390/ph16040504
Shanmugham M, Bellanger S, Leo CH. Gut-Derived Metabolite, Trimethylamine-N-oxide (TMAO) in Cardio-Metabolic Diseases: Detection, Mechanism, and Potential Therapeutics. Pharmaceuticals. 2023; 16(4):504. https://doi.org/10.3390/ph16040504
Chicago/Turabian StyleShanmugham, Meyammai, Sophie Bellanger, and Chen Huei Leo. 2023. "Gut-Derived Metabolite, Trimethylamine-N-oxide (TMAO) in Cardio-Metabolic Diseases: Detection, Mechanism, and Potential Therapeutics" Pharmaceuticals 16, no. 4: 504. https://doi.org/10.3390/ph16040504
APA StyleShanmugham, M., Bellanger, S., & Leo, C. H. (2023). Gut-Derived Metabolite, Trimethylamine-N-oxide (TMAO) in Cardio-Metabolic Diseases: Detection, Mechanism, and Potential Therapeutics. Pharmaceuticals, 16(4), 504. https://doi.org/10.3390/ph16040504