
The Therapeutic Effects of EFNB2-Fc in a Cell Model of Kawasaki Disease


Abstract
1. Introduction
2. Results
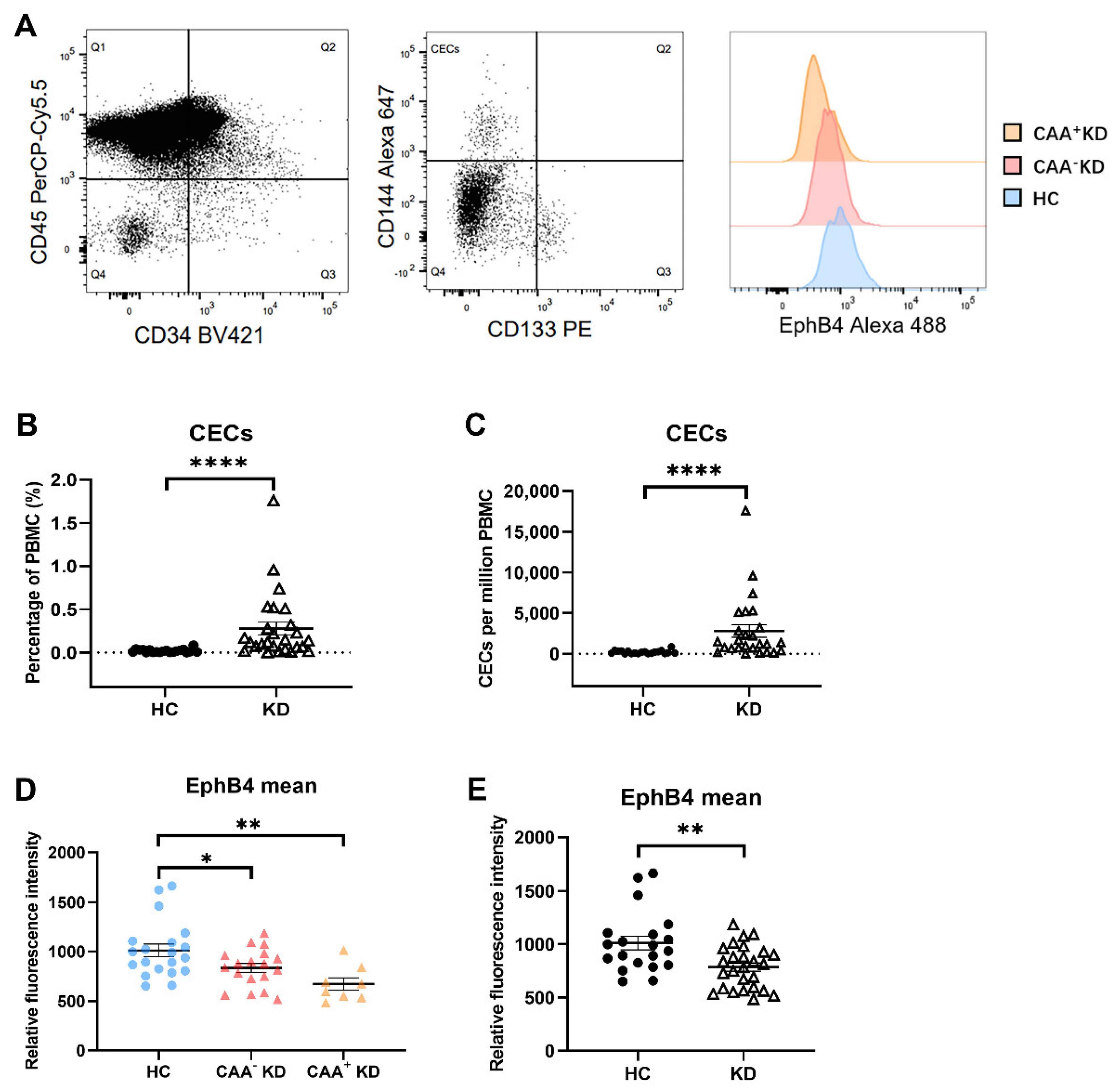
2.1. EphB4 Protein Level of CECs Were Lower in the KD Patients
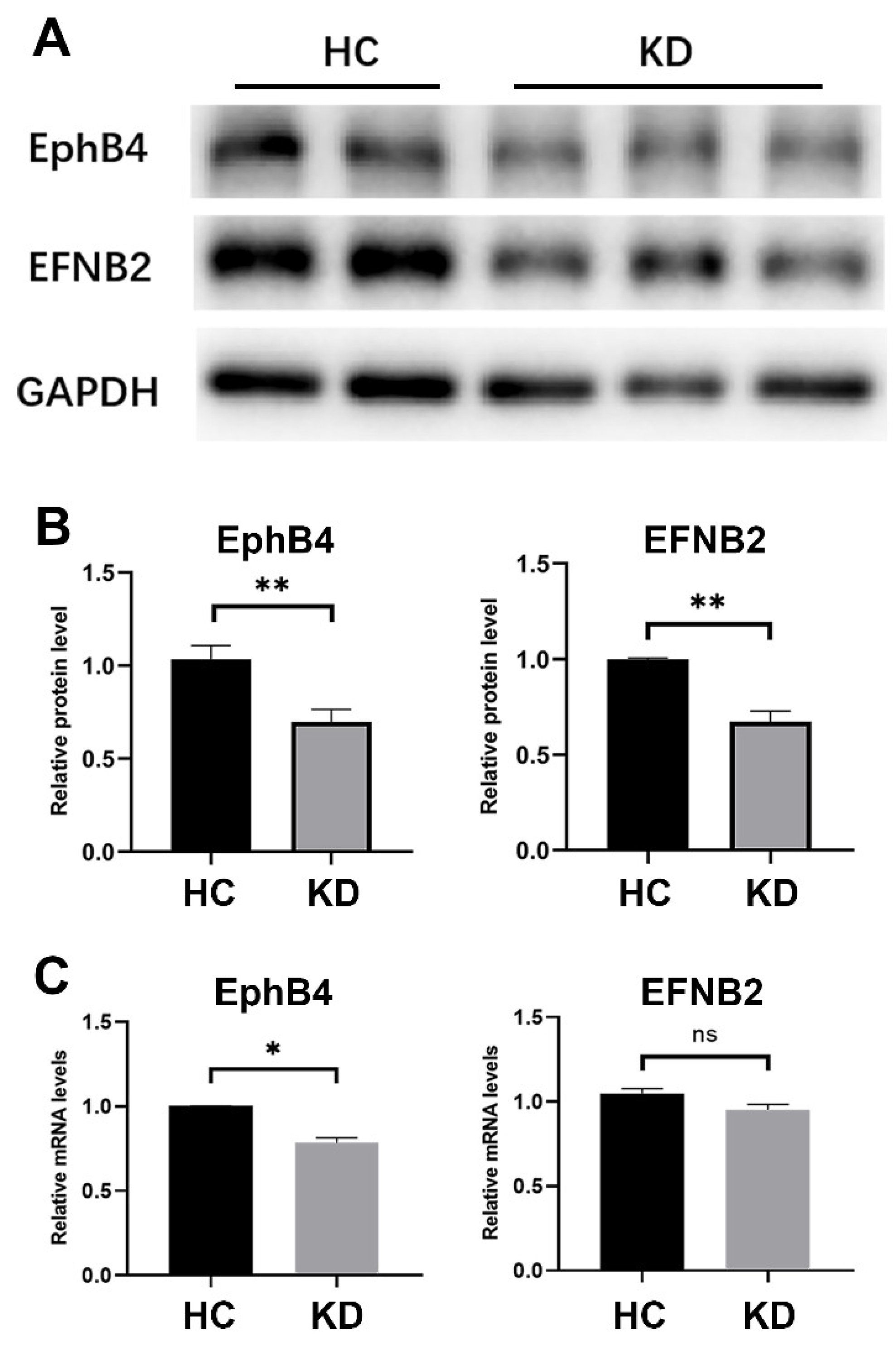
2.2. The Expression of EphB4 and EFNB2 in the Cell Model of KD Was Decreased
2.3. EphB4 Inhibition Promoted Cell Migration and Attenuated Cell Angiogenesis Ability
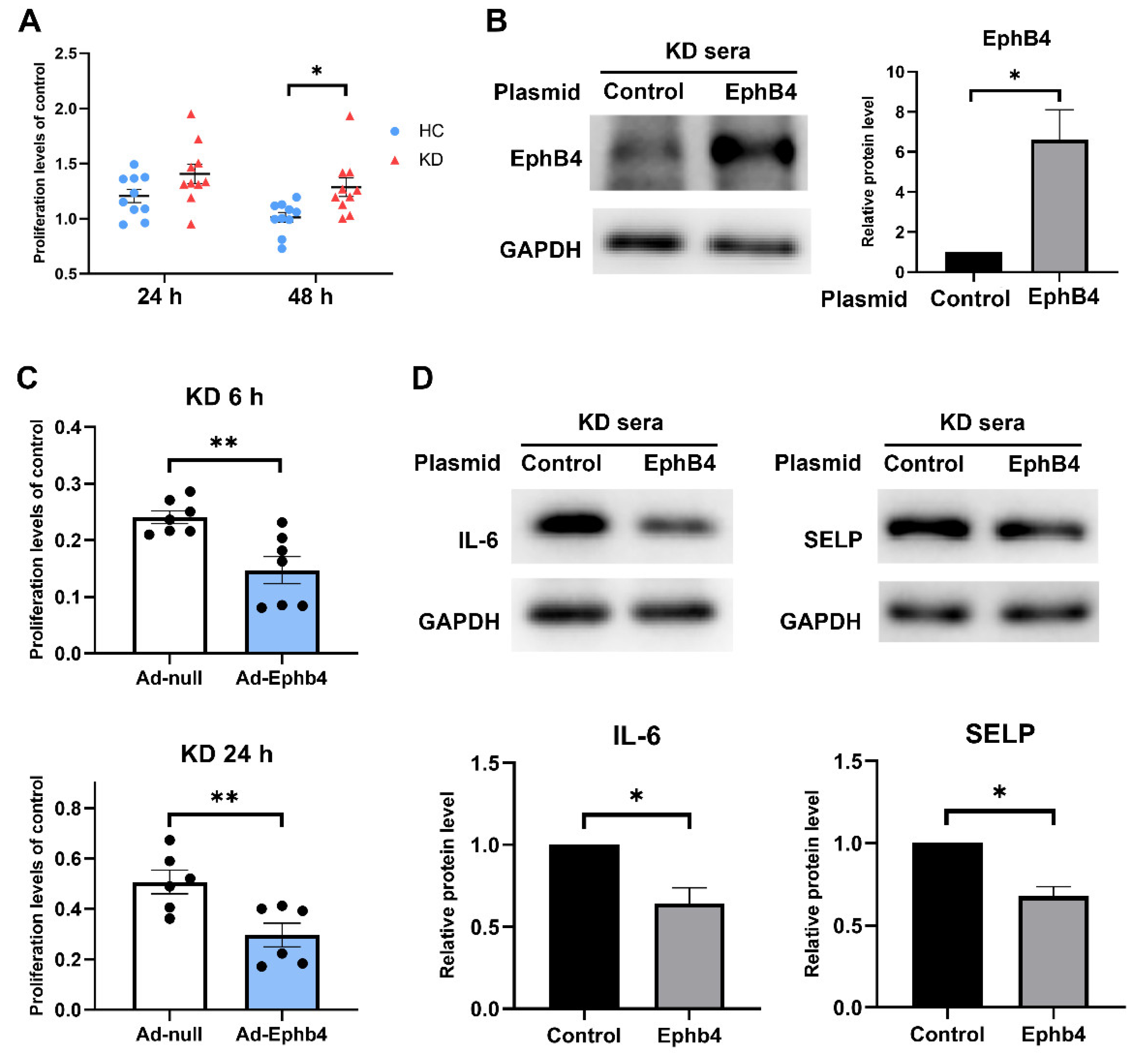
2.4. Overexpression of EphB4 Suppressed KD-Induced High Cell Viability and Inflammation
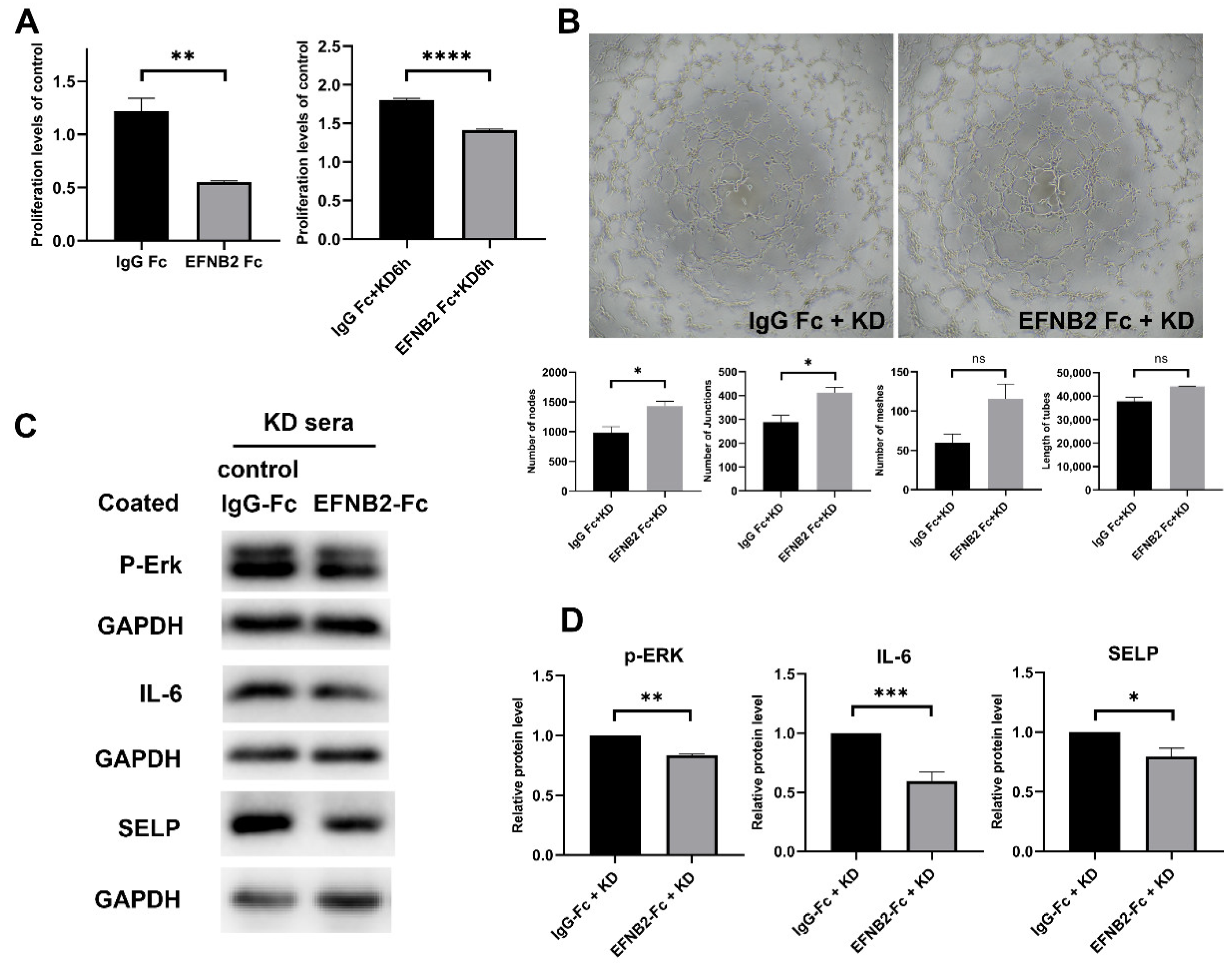
2.5. EFNB2-Fc Suppressed KD-Induced High Cell Viability and Inflammation and Elevated Cell Angiogenesis Ability
3. Discussion
4. Materials and Methods
4.1. Human Subjects
4.2. Flow Cytometry Analysis
4.3. Cell Culture and Preparation
4.4. Reverse Transcription-Real Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.5. Western Blot Assay
4.6. Recombinant Human EphrinB2 Fc Chimera
4.7. Cell Viability Assay
4.8. In Vitro Angiogenesis Assay
4.9. Wound-Healing Assay
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nakra, N.A.; Blumberg, D.A.; Herrera-Guerra, A.; Lakshminrusimha, S. Multi-System Inflammatory Syndrome in Children (MIS-C) Following SARS-CoV-2 Infection: Review of Clinical Presentation, Hypothetical Pathogenesis, and Proposed Management. Children 2020, 7, 69. [Google Scholar] [CrossRef]
- Whittaker, E.; Bamford, A.; Kenny, J.; Kaforou, M.; Jones, C.E.; Shah, P.; Ramnarayan, P.; Fraisse, A.; Miller, O.; Davies, P.; et al. Clinical Characteristics of 58 Children with a Pediatric Inflammatory Multisystem Syndrome Temporally Associated with SARS-CoV-2. JAMA 2020, 324, 259–269. [Google Scholar] [CrossRef]
- Ouldali, N.; Pouletty, M.; Mariani, P.; Beyler, C.; Blachier, A.; Bonacorsi, S.; Danis, K.; Chomton, M.; Maurice, L.; Le Bourgeois, F.; et al. Emergence of Kawasaki disease related to SARS-CoV-2 infection in an epicentre of the French COVID-19 epidemic: A time-series analysis. Lancet Child Adolesc. Health 2020, 4, 662–668. [Google Scholar] [CrossRef]
- Verdoni, L.; Mazza, A.; Gervasoni, A.; Martelli, L.; Ruggeri, M.; Ciuffreda, M.; Bonanomi, E.; D’Antiga, L. An outbreak of severe Kawasaki-like disease at the Italian epicentre of the SARS-CoV-2 epidemic: An observational cohort study. Lancet 2020, 395, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- McCrindle, B.W.; Rowley, A.H.; Newburger, J.W.; Burns, J.C.; Bolger, A.F.; Gewitz, M.; Baker, A.L.; Jackson, M.A.; Takahashi, M.; Shah, P.B.; et al. Diagnosis, Treatment, and Long-Term Management of Kawasaki Disease: A Scientific Statement for Health Professionals from the American Heart Association. Circulation 2017, 135, e927–e999. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Yamamura, K.; Sakai, Y. The up-to-date pathophysiology of Kawasaki disease. Clin. Transl. Immunol. 2021, 10, e1284. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Ninomiya, Y.; Hazeki, D.; Masuda, K.; Nomura, Y.; Kawano, Y. Disruption of Endothelial Cell Homeostasis Plays a Key Role in the Early Pathogenesis of Coronary Artery Abnormalities in Kawasaki Disease. Sci. Rep. 2017, 7, 43719. [Google Scholar] [CrossRef]
- Füller, T.; Korff, T.; Kilian, A.; Dandekar, G.; Augustin, H.G. Forward EphB4 signaling in endothelial cells controls cellular repulsion and segregation from ephrinB2 positive cells. J. Cell Sci. 2003, 116, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nakayama, M.; Pitulescu, M.E.; Schmidt, T.S.; Bochenek, M.L.; Sakakibara, A.; Adams, S.; Davy, A.; Deutsch, U.; Lüthi, U.; et al. Ephrin-B2 controls VEGF-induced angiogenesis and lymphangiogenesis. Nature 2010, 465, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Luxán, G.; Stewen, J.; Díaz, N.; Kato, K.; Maney, S.K.; Aravamudhan, A.; Berkenfeld, F.; Nagelmann, N.; Drexler, H.C.; Zeuschner, D.; et al. Endothelial EphB4 maintains vascular integrity and transport function in adult heart. eLife 2019, 8, e45863. [Google Scholar] [CrossRef]
- Capon, D.J.; Chamow, S.M.; Mordenti, J.; Marsters, S.A.; Gregory, T.; Mitsuya, H.; Byrn, R.A.; Lucas, C.; Wurm, F.M.; Groopman, J.E.; et al. Designing CD4 immunoadhesins for AIDS therapy. Nature 1989, 337, 525–531. [Google Scholar] [CrossRef]
- Strohl, W.R. Fusion Proteins for Half-Life Extension of Biologics as a Strategy to Make Biobetters. Biodrugs 2015, 29, 215–239. [Google Scholar] [CrossRef]
- Weinblatt, M.E.; Kremer, J.M.; Bankhurst, A.D.; Bulpitt, K.J.; Fleischmann, R.M.; Fox, R.I.; Jackson, C.G.; Lange, M.; Burge, D.J. A Trial of Etanercept, a Recombinant Tumor Necrosis Factor Receptor: Fc Fusion Protein, in Patients with Rheumatoid Arthritis Receiving Methotrexate. N. Engl. J. Med. 1999, 340, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Jafari, R.; Zolbanin, N.M.; Rafatpanah, H.; Majidi, J.; Kazemi, T. Fc-fusion Proteins in Therapy: An Updated View. Curr. Med. Chem. 2017, 24, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Ehlken, C.; Martin, G.; Lange, C.; Gogaki, E.G.; Fiedler, U.; Schaffner, F.; Hansen, L.L.; Augustin, H.; Agostini, H.T. Therapeutic interference with EphrinB2 signalling inhibits oxygen-induced angioproliferative retinopathy. Acta Ophthalmol. 2011, 89, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Pei, D.; Shi, L.; Cui, Y.; Hong, Z. Ephrin-B2 inhibits Abeta (25-35)-induced apoptosis by alleviating endoplasmic reticulum stress and promoting autophagy in HT22 cells. Neurosci. Lett. 2019, 704, 50–56. [Google Scholar] [CrossRef]
- Qu, F.; Song, Y.; Wu, Y.; Huang, Y.; Zhong, Q.; Zhang, Y.; Fan, Z.; Xu, C. The protective role of Ephrin-B2/EphB4 signaling in osteogenic differentiation under inflammatory environment. Exp. Cell Res. 2021, 400, 112505. [Google Scholar] [CrossRef]
- Salvucci, O.; Ohnuki, H.; Maric, D.; Hou, X.; Li, X.; Yoon, S.O.; Segarra, M.; Eberhart, C.G.; Acker-Palmer, A.; Tosato, G. EphrinB2 controls vessel pruning through STAT1-JNK3 signalling. Nat. Commun. 2015, 6, 6576. [Google Scholar] [CrossRef]
- Valverde, I.; Singh, Y.; Sanchez-De-Toledo, J.; Theocharis, P.; Chikermane, A.; Di Filippo, S.; Kuciñska, B.; Mannarino, S.; Tamariz-Martel, A.; Gutierrez-Larraya, F.; et al. Acute Cardiovascular Manifestations in 286 Children with Multisystem Inflammatory Syndrome Associated With COVID-19 Infection in Europe. Circulation 2021, 143, 21–32. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, J.; Liu, J.; Geng, Z.; Tao, Y.; Zheng, F.; Wang, Y.; Fu, S.; Wang, W.; Xie, C.; et al. The role of Ca2+/NFAT in Dysfunction and Inflammation of Human Coronary Endothelial Cells induced by Sera from patients with Kawasaki disease. Sci. Rep. 2020, 10, 4706. [Google Scholar] [CrossRef]
- Zheng, F.; Tao, Y.; Liu, J.; Geng, Z.; Wang, Y.; Wang, Y.; Fu, S.; Wang, W.; Xie, C.; Zhang, Y.; et al. KCa3.1 Inhibition of Macrophages Suppresses Inflammatory Response Leading to Endothelial Damage in a Cell Model of Kawasaki Disease. J. Inflamm. Res. 2021, 14, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Irie, N.; Takada, Y.; Shimoda, K.; Miyamoto, T.; Nishiwaki, T.; Suda, T.; Matsuo, K. Bidirectional ephrinB2-EphB4 signaling controls bone homeostasis. Cell Metab. 2006, 4, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Dietz, S.M.; Tacke, C.E.; Gort, J.; Kuipers, I.M.; De Groot, E.; Wiegman, A.; Hutten, B.A.; Kuijpers, T.W. Carotid Intima-Media Thickness in Patients with a History of Kawasaki Disease. Circ. J. 2015, 79, 2682–2687. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, K.; Takeshita, S.; Tsujimoto, H.; Kawamura, Y.; Tokutomi, T.; Sekine, I. Circulating endothelial cells in Kawasaki disease. Clin. Exp. Immunol. 2003, 131, 536–540. [Google Scholar] [CrossRef]
- Fu, S.; Gong, F.; Xie, C.; Zhu, W.; Wang, W.; Shen, H.; Tang, Y. S100A12 on Circulating Endothelial Cells Surface in Children with Kawasaki Disease. Pediatr. Res. 2010, 68, 165–168. [Google Scholar] [CrossRef]
- Chen, D.; Hughes, E.D.; Saunders, T.L.; Wu, J.; Vasquez, M.N.H.; Makinen, T.; King, P.D. Angiogenesis depends upon EPHB4-mediated export of collagen IV from vascular endothelial cells. JCI Insight 2022, 7, e156928. [Google Scholar] [CrossRef]
- Sakata, K.; Kita, M.; Imanishi, J.; Onouchi, Z.; Liu, Y.; Mitsui, Y. Effect of Kawasaki Disease on Migration of Human Umbilical Vein Endothelial Cells. Pediatr. Res. 1995, 38, 501–505. [Google Scholar] [CrossRef]
- Higashi, K.; Terai, M.; Hamada, H.; Honda, T.; Kanazawa, M.; Kohno, Y. Impairment of Angiogenic Activity in the Serum from Patients with Coronary Aneurysms Due to Kawasaki Disease. Circ. J. 2007, 71, 1052–1059. [Google Scholar] [CrossRef]
- Chiu, W.C.; Lin, J.Y.; Lee, T.S.; You, L.R.; Chiang, A.N. β2-glycoprotein I inhibits VEGF-induced endothelial cell growth and migration via suppressing phosphorylation of VEGFR2, ERK1/2, and Akt. Mol. Cell. Biochem. 2013, 372, 9–15. [Google Scholar] [CrossRef]
- Meadows, K.N.; Bryant, P.; Pumiglia, K. Vascular Endothelial Growth Factor Induction of the Angiogenic Phenotype Requires Ras Activation. J. Biol. Chem. 2001, 276, 49289–49298. [Google Scholar] [CrossRef]
- Battle, T.E.; Lynch, R.A.; Frank, D.A. Signal Transducer and Activator of Transcription 1 Activation in Endothelial Cells Is a Negative Regulator of Angiogenesis. Cancer Res. 2006, 66, 3649–3657. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.W.; Feng, K.; Liu, X.L.; Chen, H.F.; Sun, Z.Y.; Wang, C.F.; Liu, Z.Q.; Wang, H.W.; Zhang, J.W.; Yu, D.G.; et al. The Recombinant Protein EphB4-Fc Changes the Ti Particle-Mediated Imbalance of OPG/RANKL via EphrinB2/EphB4 Signaling Pathway and Inhibits the Release of Proinflammatory Factors In Vivo. Oxid. Med. Cell. Longev. 2020, 2020, 1404915. [Google Scholar] [CrossRef]
- Ye, Q.; Shao, W.-X.; Shang, S.-Q.; Zhang, T.; Hu, J.; Zhang, C.-C. A Comprehensive Assessment of the Value of Laboratory Indices in Diagnosing Kawasaki Disease. Arthritis Rheumatol. 2015, 67, 1943–1950. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, S.; Dobashi, H.; Nakatani, K.; Koike, Y.; Tsujimoto, H.; Hirayama, K.; Kawamura, Y.; Mori, K.; Sekine, I.; Yoshioka, S. Circulating soluble selectins in Kawasaki disease. Clin. Exp. Immunol. 1997, 108, 446–450. [Google Scholar] [CrossRef]
- Kwan Tat, S.; Pelletier, J.P.; Amiable, N.; Boileau, C.; Lajeunesse, D.; Duval, N.; Martel-Pelletier, J. Activation of the receptor EphB4 by its specific ligand ephrin B2 in human osteoarthritic subchondral bone osteoblasts. Arthritis Rheum. 2008, 58, 3820–3830. [Google Scholar] [CrossRef] [PubMed]
- Groppa, E.; Brkic, S.; Uccelli, A.; Wirth, G.; Korpisalo-Pirinen, P.; Filippova, M.; Dasen, B.; Sacchi, V.; Muraro, M.G.; Trani, M.; et al. EphrinB2/EphB4 signaling regulates non-sprouting angiogenesis by VEGF. EMBO Rep. 2018, 19, e45054. [Google Scholar] [CrossRef]
- Foubert, P.; Silvestre, J.-S.; Souttou, B.; Barateau, V.; Martin, C.; Ebrahimian, T.G.; Leré-Déan, C.; Contreres, J.O.; Sulpice, E.; Levy, B.I.; et al. PSGL-1–mediated activation of EphB4 increases the proangiogenic potential of endothelial progenitor cells. J. Clin. Investig. 2007, 117, 1527–1537. [Google Scholar] [CrossRef]
- Donato, C.; Kunz, L.; Castro-Giner, F.; Paasinen-Sohns, A.; Strittmatter, K.; Szczerba, B.M.; Scherrer, R.; Di Maggio, N.; Heusermann, W.; Biehlmaier, O.; et al. Hypoxia Triggers the Intravasation of Clustered Circulating Tumor Cells. Cell Rep. 2020, 32, 108105. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| EphB4 | CCACCGGGAAGGTGAATGTC | CTGGGCGCACTTTTTGTAGAA |
| EFNB2 | TATGCAGAACTGCGATTTCCAA | TGGGTATAGTACCAGTCCTTGTC |
| GAPDH | GGAGCGAGATCCCTCCAAAAT | GGCTGTTGTCATACTTCTCATGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, Y.; Wang, W.; Jin, Y.; Wang, M.; Xu, J.; Wang, Y.; Gong, F. The Therapeutic Effects of EFNB2-Fc in a Cell Model of Kawasaki Disease. Pharmaceuticals 2023, 16, 500. https://doi.org/10.3390/ph16040500
Tao Y, Wang W, Jin Y, Wang M, Xu J, Wang Y, Gong F. The Therapeutic Effects of EFNB2-Fc in a Cell Model of Kawasaki Disease. Pharmaceuticals. 2023; 16(4):500. https://doi.org/10.3390/ph16040500
Chicago/Turabian StyleTao, Yijing, Wei Wang, Yihua Jin, Min Wang, Jiawen Xu, Yujia Wang, and Fangqi Gong. 2023. "The Therapeutic Effects of EFNB2-Fc in a Cell Model of Kawasaki Disease" Pharmaceuticals 16, no. 4: 500. https://doi.org/10.3390/ph16040500
APA StyleTao, Y., Wang, W., Jin, Y., Wang, M., Xu, J., Wang, Y., & Gong, F. (2023). The Therapeutic Effects of EFNB2-Fc in a Cell Model of Kawasaki Disease. Pharmaceuticals, 16(4), 500. https://doi.org/10.3390/ph16040500