

Hesperidin Mitigates Cyclophosphamide-Induced Testicular Dysfunction via Altering the Hypothalamic Pituitary Gonadal Axis and Testicular Steroidogenesis, Inflammation, and Apoptosis in Male Rats


Abstract
1. Introduction
2. Results
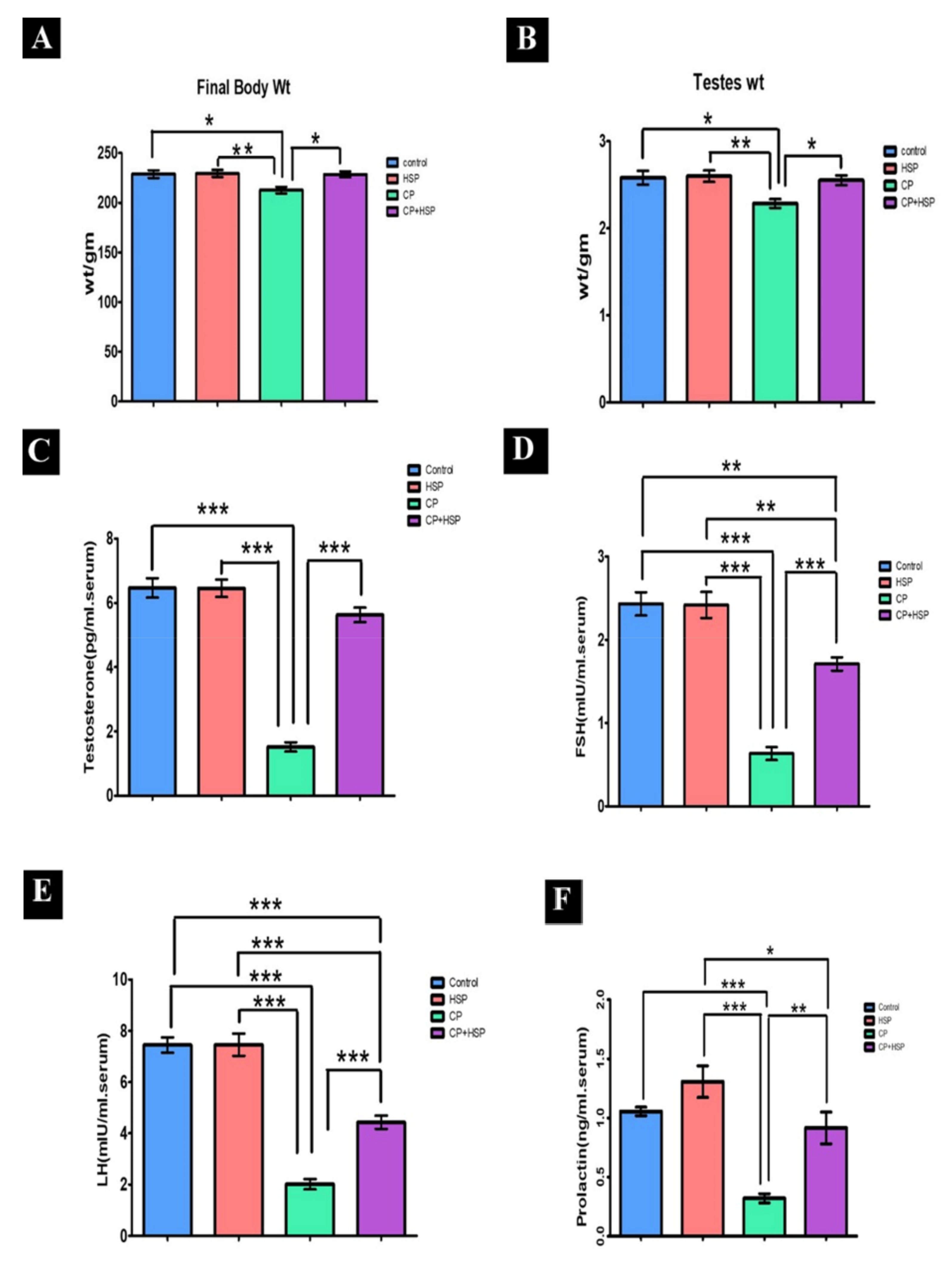
2.1. Effect on the Final Body, Testicular Weights and Serum Hormone Levels
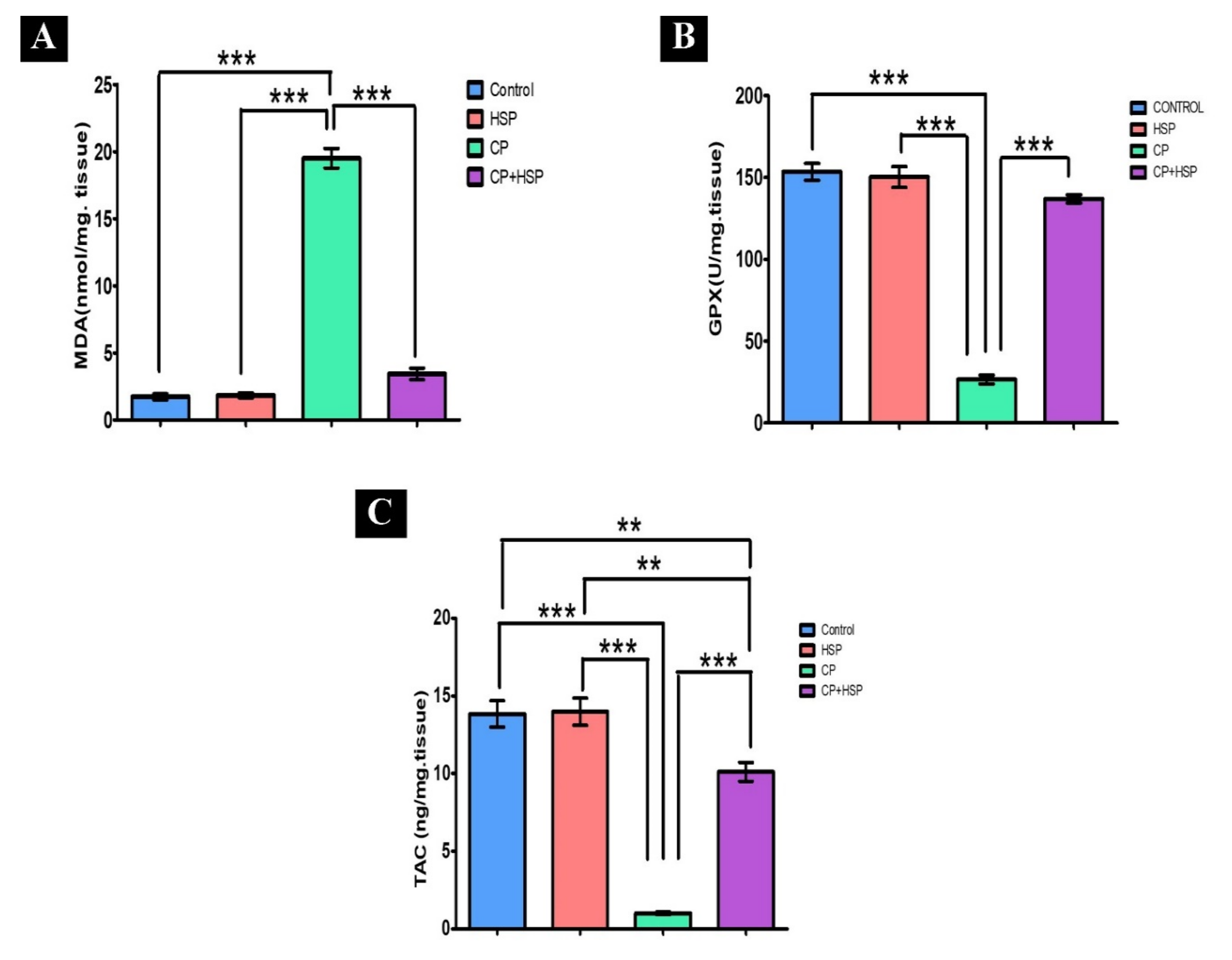
2.2. Effect on Testicular Lipid Peroxidation and Oxidative Stress Markers
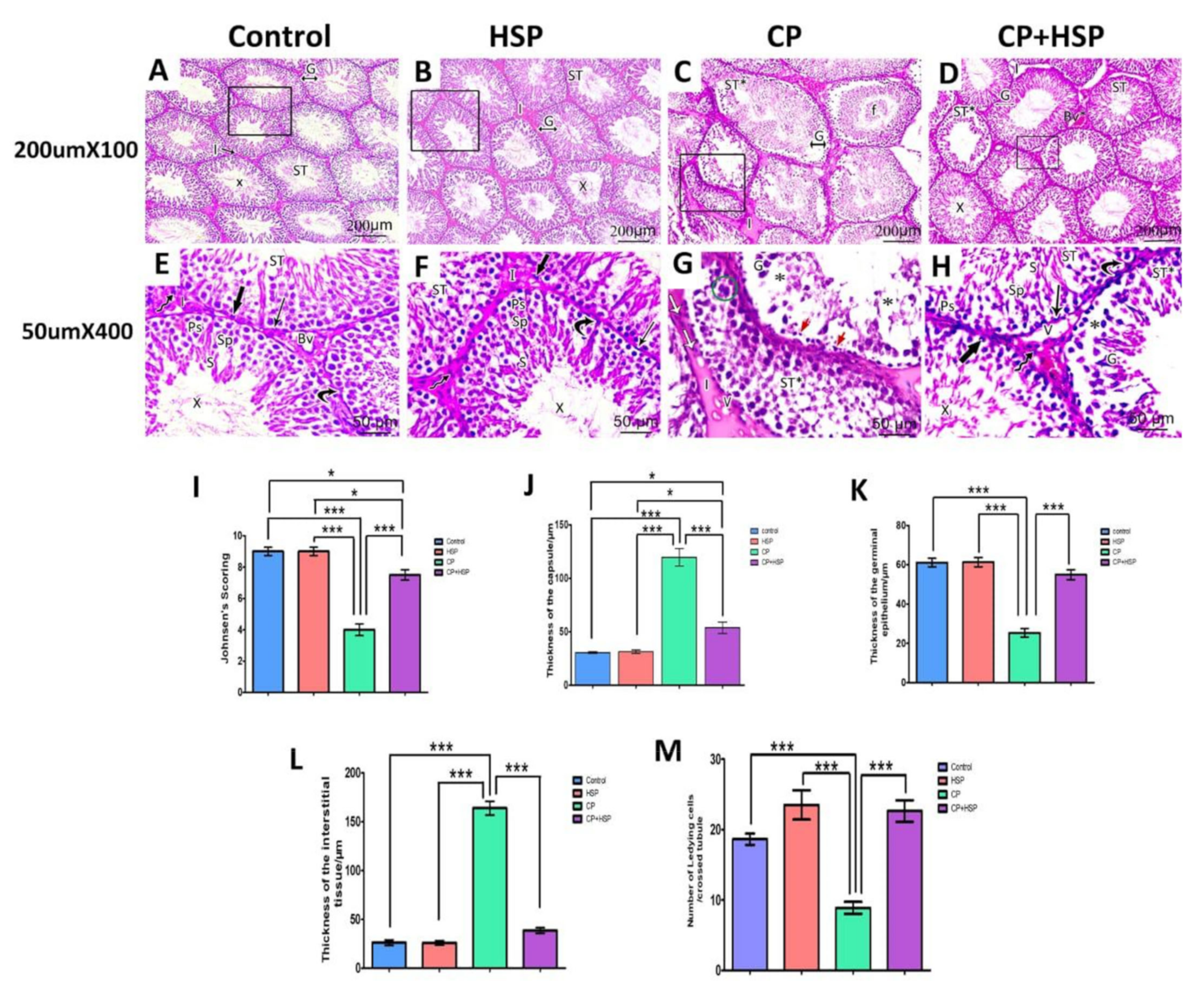
2.3. Effect on Testicular Histopathology and Morphology
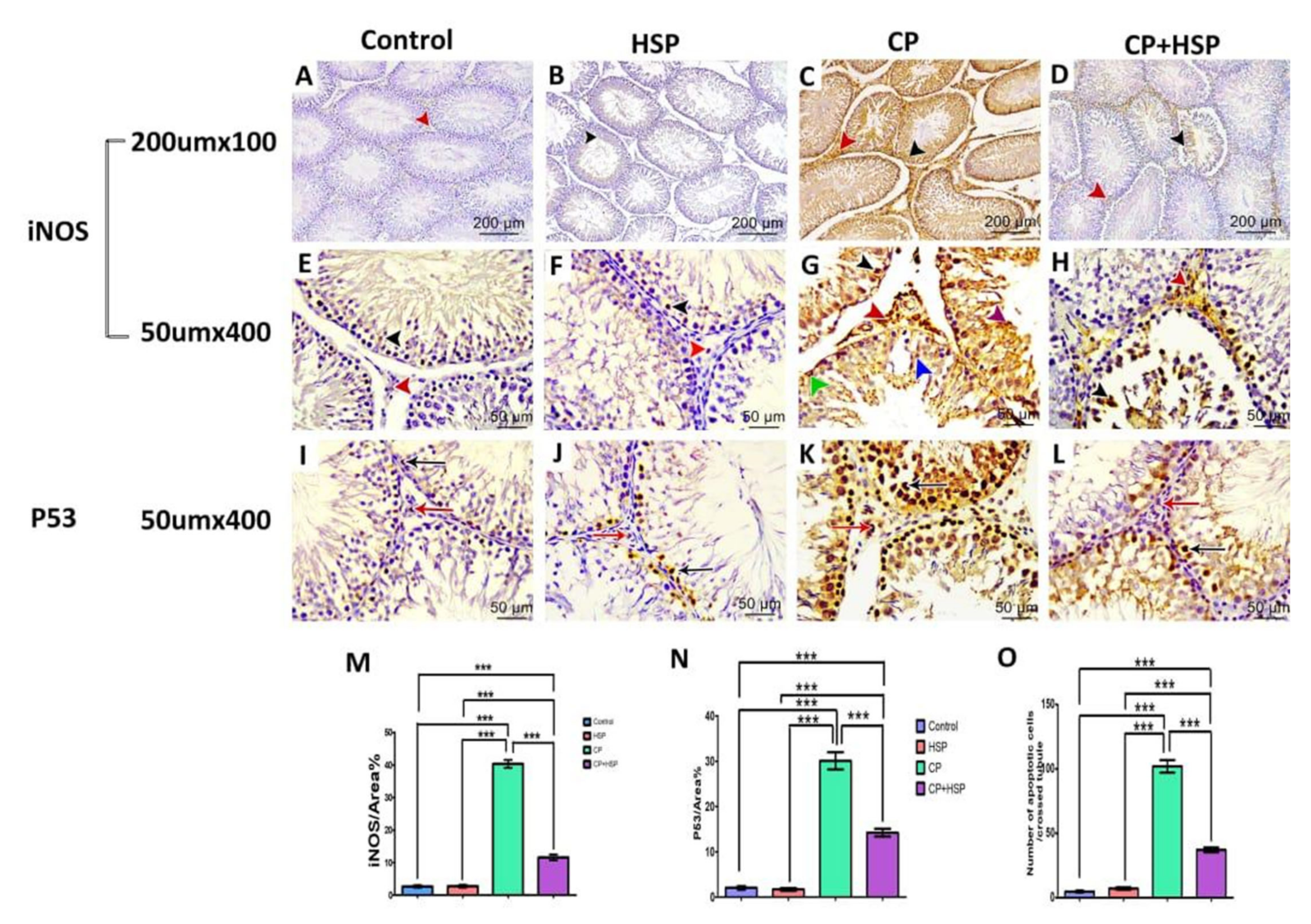
2.4. Effect on Testicular Immunohistochemistry of iNOS and P53
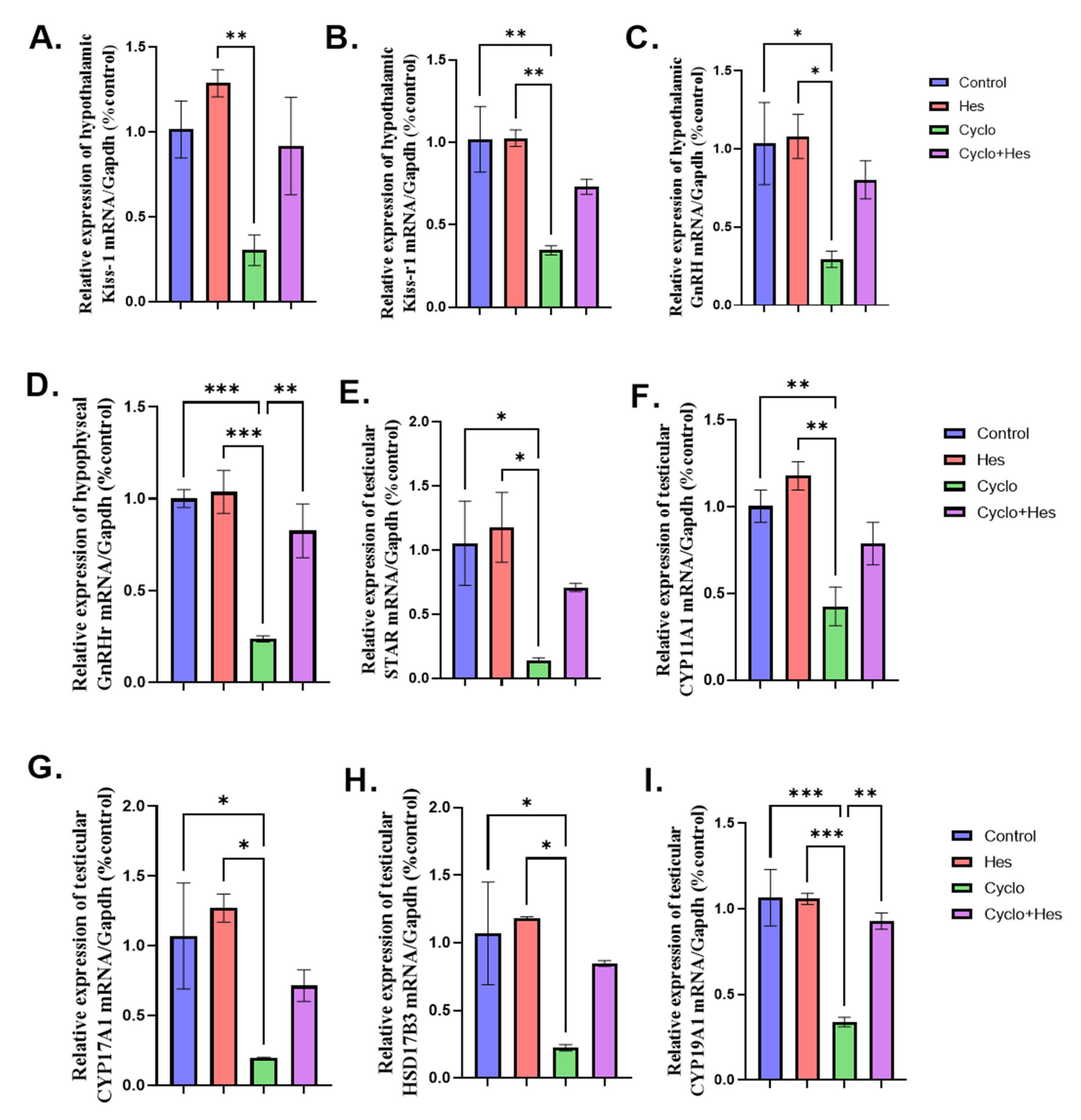
2.5. Effect on mRNA Expression of Hypothalamic KISS-1, KISS-1r, GnRH, Hypophyseal GnRHr and Testicular Steroidogenic Enzymes
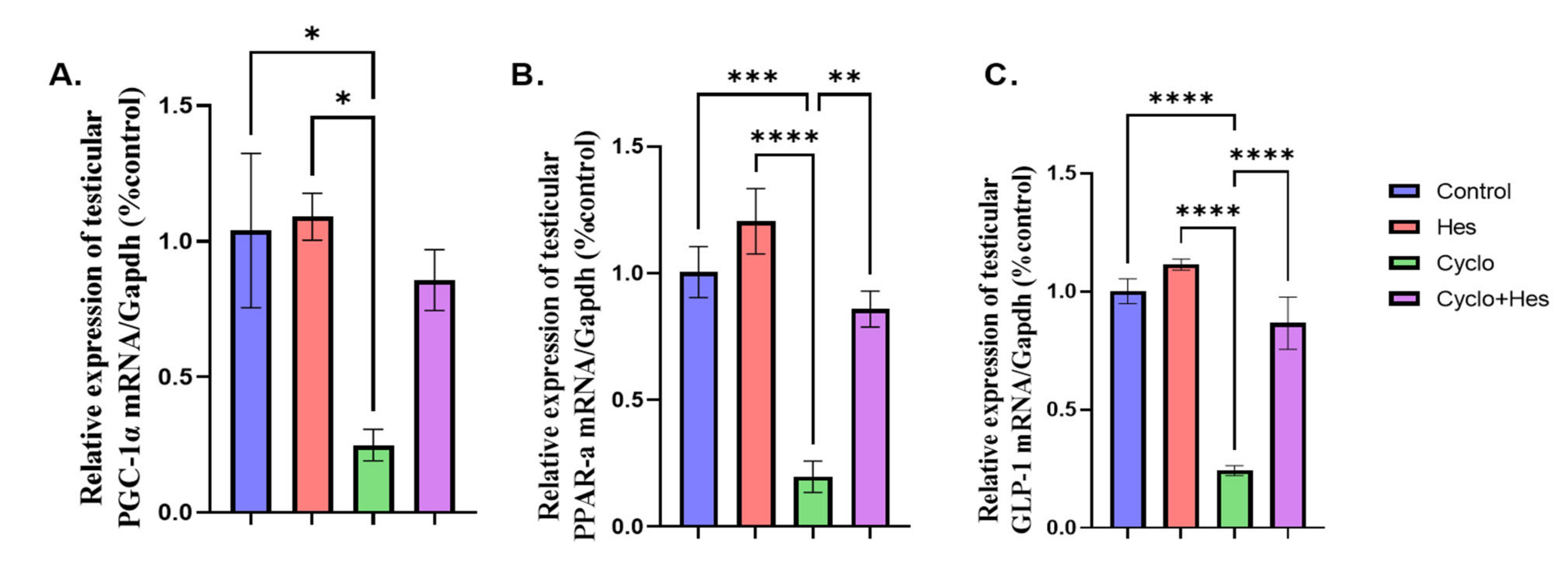
2.6. Effect on Testicular mRNA Expression of GLP-1, PGC-1, and PPAR-α
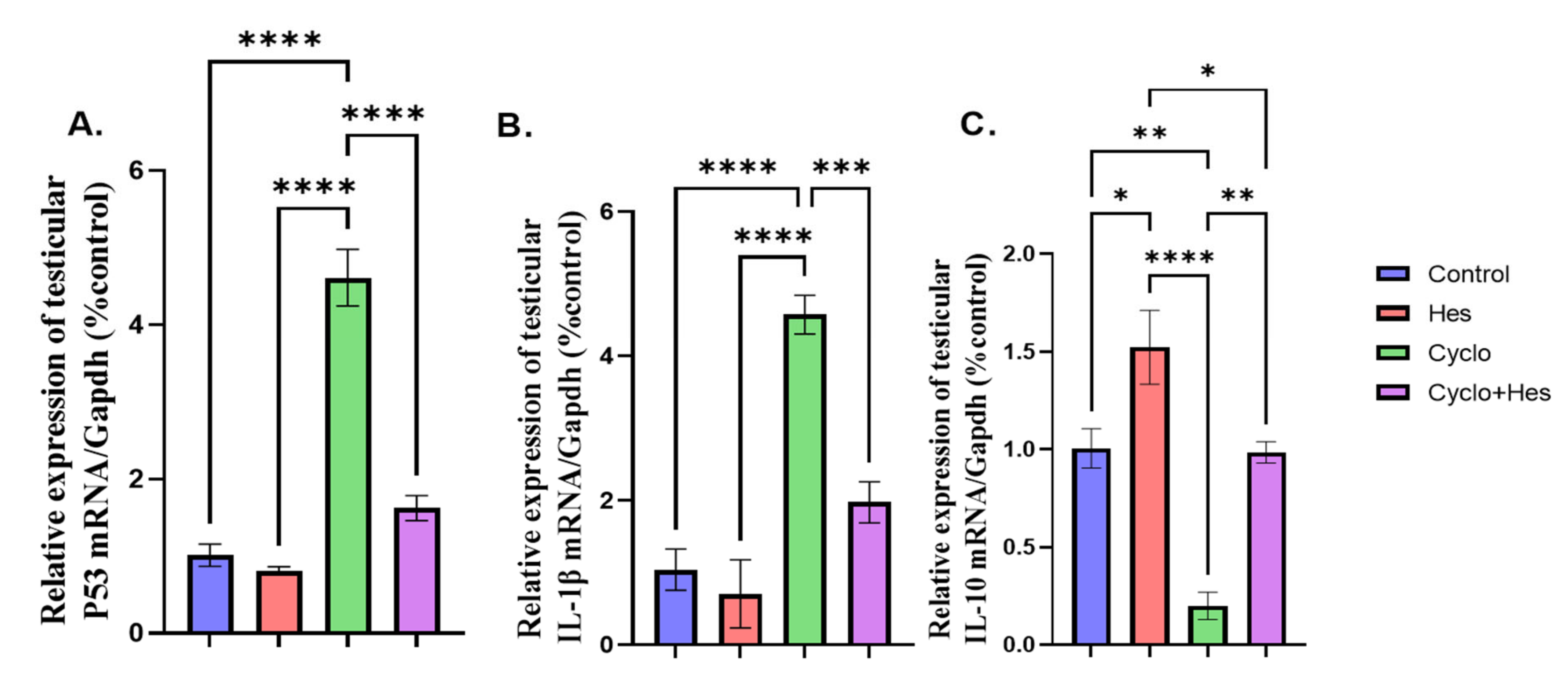
2.7. Effect on Testicular Apoptotic and Inflammatory Marker
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Experimental Animals
4.3. Experimental Design and Sample Collection
4.4. Hormonal and Biochemical Analysis
4.5. Real-Time Quantitative RT-PCR (qRT-PCR) Analysis
4.6. Histopathological and Immunohistochemical Examination
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yilmaz, E.; Coskun, E.I.; Sahin, N.; Ciplak, B.; Ekici, K. MPV, NLR, and platelet count: New hematologic markers in diagnosis of malignant ovarian tumor. Eur. J. Gynaecol. Oncol. 2017, 38, 346–349. [Google Scholar] [PubMed]
- Klareskog, L.; van der Heijde, D.; de Jager, J.P.; Gough, A.; Kalden, J.; Malaise, M.; Mola, E.M.; Pavelka, K.; Sany, J.; Settas, L.; et al. Therapeutic effect of the combination of etanercept and methotrexate compared with each treatment alone in patients with rheumatoid arthritis: Double-blind randomised controlled trial. Lancet 2004, 363, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Veal, G.J.; Cole, M.; Chinnaswamy, G.; Sludden, J.; Jamieson, D.; Errington, J.; Malik, G.; Hill, C.R.; Chamberlain, T.; Boddy, A.V. Cyclophosphamide pharmacokinetics and pharmacogenetics in children with B-cell non-Hodgkin’s lymphoma. Eur. J. Cancer 2016, 55, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Boisseaux, P.; Noury, P.; Thomas, H.; Garric, J. Immune responses in the aquatic gastropod Lymnaea stagnalis under short-term exposure to pharmaceuticals of concern for immune systems: Diclofenac, cyclophosphamide and cyclosporine A. Ecotoxicol. Environ. Saf. 2017, 139, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Grzesiuk, M.; Mielecki, D.; Pilżys, T.; Garbicz, D.; Marcinkowski, M.; Grzesiuk, E. How cyclophosphamide at environmentally relevant concentration influences Daphnia magna life history and its proteome. PLoS ONE 2018, 13, e0195366. [Google Scholar] [CrossRef]
- Temel, Y.; Çağlayan, C.; Ahmed, B.M.; Kandemir, F.M.; Çiftci, M. The effects of chrysin and naringin on cyclophosphamide-induced erythrocyte damage in rats: Biochemical evaluation of some enzyme activities in vivo and in vitro. Naunyn Schmiedebergs Arch Pharm. 2021, 394, 645–654. [Google Scholar] [CrossRef]
- Spears, N.; Lopes, F.; Stefansdottir, A.; Rossi, V.; De Felici, M.; Anderson, R.A.; Klinger, F.G. Ovarian damage from chemotherapy and current approaches to its protection. Hum. Reprod. Update 2019, 25, 673–693. [Google Scholar] [CrossRef]
- Akomolafe, S.F.; Aluko, B.T. Protective effect of curcumin on fertility in cyclophosphamide exposed rats: Involvement of multiple pathways. J. Food Biochem. 2020, 44, e13095. [Google Scholar] [CrossRef]
- Khamis, T.; Abdelalim, A.F.; Saeed, A.A.; Edress, N.M.; Nafea, A.; Ebian, H.F.; Algendy, R.; Hendawy, D.M.; Arisha, A.H.; Abdallah, S.H. Breast milk MSCs upregulated β-cells PDX1, Ngn3, and PCNA expression via remodeling ER stress/inflammatory/apoptotic signaling pathways in type 1 diabetic rats. Eur. J. Pharmacol. 2021, 905, 174188. [Google Scholar] [CrossRef]
- Khamis, T.; Abdelalim, A.F.; Abdallah, S.H.; Saeed, A.A.; Edress, N.M.; Arisha, A.H. Early intervention with breast milk mesenchymal stem cells attenuates the development of diabetic-induced testicular dysfunction via hypothalamic Kisspeptin/Kiss1r-GnRH/GnIH system in male rats. Biochim. Et Biophys. Acta-Mol. Basis Dis. 2020, 1866, 165577. [Google Scholar] [CrossRef]
- Ebokaiwe, A.P.; Obasi, D.O.; Njoku, R.C.; Osawe, S. Cyclophosphamide-induced testicular oxidative-inflammatory injury is accompanied by altered immunosuppressive indoleamine 2, 3-dioxygenase in Wister rats: Influence of dietary quercetin. Andrologia 2022, 54, e14341. [Google Scholar] [CrossRef]
- Van den Boogaard, W.M.C.; Komninos, D.S.J.; Vermeij, W.P. Chemotherapy Side-Effects: Not All DNA Damage Is Equal. Cancers 2022, 14, 627. [Google Scholar] [CrossRef]
- Naz, S.; Imran, M.; Rauf, A.; Orhan, I.E.; Shariati, M.A.; Iahtisham Ul, H.; IqraYasmin; Shahbaz, M.; Qaisrani, T.B.; Shah, Z.A.; et al. Chrysin: Pharmacological and therapeutic properties. Life Sci. 2019, 235, 116797. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef]
- Shoorei, H.; Banimohammad, M.; Kebria, M.M.; Afshar, M.; Taheri, M.M.; Shokoohi, M.; Farashah, M.S.; Eftekharzadeh, M.; Akhiani, O.; Gaspar, R.; et al. Hesperidin improves the follicular development in 3D culture of isolated preantral ovarian follicles of mice. Exp. Biol. Med. 2019, 244, 352–361. [Google Scholar] [CrossRef]
- Lehmann, V.; Chemaitilly, W.; Lu, L.; Green, D.M.; Kutteh, W.H.; Brinkman, T.M.; Srivastava, D.K.; Robison, L.L.; Hudson, M.M.; Klosky, J.L. Gonadal Functioning and Perceptions of Infertility Risk Among Adult Survivors of Childhood Cancer: A Report From the St Jude Lifetime Cohort Study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2019, 37, 893–902. [Google Scholar] [CrossRef]
- Fusco, R.; Salinaro, A.T.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Crea, R.; Cordaro, M.; Cuzzocrea, S.; et al. Hidrox(®) Counteracts Cyclophosphamide-Induced Male Infertility through NRF2 Pathways in a Mouse Model. Antioxidants 2021, 10, 778. [Google Scholar] [CrossRef]
- Ghobadi, E.; Moloudizargari, M.; Asghari, M.H.; Abdollahi, M. The mechanisms of cyclophosphamide-induced testicular toxicity and the protective agents. Expert Opin. Drug Metab. Toxicol. 2017, 13, 525–536. [Google Scholar] [CrossRef]
- Yasueda, A.; Urushima, H.; Ito, T. Efficacy and Interaction of Antioxidant Supplements as Adjuvant Therapy in Cancer Treatment: A Systematic Review. Integr. Cancer Ther. 2016, 15, 17–39. [Google Scholar] [CrossRef]
- Zouchoune, B. How the ascorbic acid and hesperidin do improve the biological activities of the cinnamon: Theoretical investigation. Struct. Chem. 2020, 31, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Afzal, S.M.; Vafa, A.; Rashid, S.; Barnwal, P.; Shahid, A.; Shree, A.; Islam, J.; Ali, N.; Sultana, S. Protective effect of hesperidin against N,N’-dimethylhydrazine induced oxidative stress, inflammation, and apoptotic response in the colon of Wistar rats. Environ. Toxicol. 2020, 36, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Khorwal, G.; Chauhan, R.; Nagar, M.; Khorwal, G. Effect of cyclophosphamide on liver in albino rats: A comparative dose dependent histomorphological study. Int. J. Biomed. Adv. Res. 2017, 8, 102–107. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, Y.; Wang, Y. Protective roles and mechanisms of rosmarinic acid in cyclophosphamide-induced premature ovarian failure. J. Biochem. Mol. Toxicol. 2020, 34, e22591. [Google Scholar] [CrossRef]
- Omole, J.G.; Ayoka, O.A.; Alabi, Q.K.; Adefisayo, M.A.; Asafa, M.A.; Olubunmi, B.O.; Fadeyi, B.A. Protective Effect of Kolaviron on Cyclophosphamide-Induced Cardiac Toxicity in Rats. J. Evid.-Based Integr. Med. 2018, 23, 2156587218757649. [Google Scholar] [CrossRef]
- Razak, R.; Ismail, F.; Isa, M.L.M.; Wahab, A.Y.A.; Muhammad, H.; Ramli, R.; Ismail, R. Ameliorative Effects of Aquilaria malaccensis Leaves Aqueous Extract on Reproductive Toxicity Induced by Cyclophosphamide in Male Rats. Malays. J. Med. Sci.: MJMS 2019, 26, 44–57. [Google Scholar] [CrossRef]
- Afolabi, O.K.; Wusu, A.D.; Ugbaja, R.; Fatoki, J.O. Aluminium phosphide-induced testicular toxicity through oxidative stress in Wistar rats: Ameliorative role of hesperidin. Toxicol. Res. Appl. 2018, 2, 2397847318812794. [Google Scholar] [CrossRef]
- Hamdy, S.M.; Sayed, O.N.; Abdel Latif, A.K.M.; Abd-Elazeez, A.M.; Amin, A.M. Protective Effect Of Hesperidin And Tiger Nut Against DMBA Carcinogenicity In Female Rats. Biochem. Lett. 2016, 12, 150–167. [Google Scholar] [CrossRef]
- Rezaei, S.; Hosseinimehr, S.J.; Zargari, M.; Karimpour Malekshah, A.; Mirzaei, M.; Talebpour Amiri, F. Protective effects of sinapic acid against cyclophosphamide-induced testicular toxicity via inhibiting oxidative stress, caspase-3 and NF-kB activity in BALB/c mice. Andrologia 2021, 53, e14196. [Google Scholar] [CrossRef]
- Mehraban, Z.; Ghaffari Novin, M.; Golmohammadi, M.G.; Sagha, M.; Ziai, S.A.; Abdollahifar, M.A.; Nazarian, H. Protective Effect of Gallic Acid on Testicular Tissue, Sperm Parameters, and DNA Fragmentation against Toxicity Induced by Cyclophosphamide in Adult NMRI Mice. Urol. J. 2020, 17, 78–85. [Google Scholar] [CrossRef]
- Mohammadi, F.; Nikzad, H.; Taghizadeh, M.; Taherian, A.; Azami-Tameh, A.; Hosseini, S.M.; Moravveji, A. Protective effect of Zingiber officinale extract on rat testis after cyclophosphamide treatment. Andrologia 2014, 46, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Chen, L.; Wu, H.; Chen, T.; Wang, F.; Chen, X.; Sun, H.; Li, X. GnRH Antagonist Improves Pubertal Cyclophosphamide-Induced Long-Term Testicular Injury in Adult Rats. Int. J. Endocrinol. 2018, 2018, 4272575. [Google Scholar] [CrossRef] [PubMed]
- Helmy, H.S.; Senousy, M.A.; El-Sahar, A.E.; Sayed, R.H.; Saad, M.A.; Elbaz, E.M. Aberrations of miR-126-3p, miR-181a and sirtuin1 network mediate Di-(2-ethylhexyl) phthalate-induced testicular damage in rats: The protective role of hesperidin. Toxicology 2020, 433–434, 152406. [Google Scholar] [CrossRef] [PubMed]
- Shabaan, S.; Madi, N.; Elgharib, M.; Nasif, E. Study the Effect of Silymarin on Cyclophosphamide Induced Testicular Damage in Adult Albino Rats. Bull. Egypt. Soc. Physiol. Sci. 2021, 41, 553–564. [Google Scholar] [CrossRef]
- Hassan, M.A.; El Bohy, K.M.; El Sharkawy, N.I.; Imam, T.S.; El-Metwally, A.E.; Hamed Arisha, A.; Mohammed, H.A.; Abd-Elhakim, Y.M. Iprodione and chlorpyrifos induce testicular damage, oxidative stress, apoptosis and suppression of steroidogenic- and spermatogenic-related genes in immature male albino rats. Andrologia 2021, 53, e13978. [Google Scholar] [CrossRef]
- Kasem, S.E.; Abdelnaby, A.A.; Mohammed, P.A.; Hemdan, S.B.; Abd El-Fattah, R.M.Z.; Elsayed, R.M. Protective Effect of Hesperidin on Kidneys and Testes of Adult Male Rats Exposed to Bisphenol A. Egypt. J. Hosp. Med. 2022, 88, 3005–3013. [Google Scholar] [CrossRef]
- Arisha, A.H.; Ahmed, M.M.; Kamel, M.A.; Attia, Y.A.; Hussein, M.M.A. Morin ameliorates the testicular apoptosis, oxidative stress, and impact on blood–testis barrier induced by photo-extracellularly synthesized silver nanoparticles. Environ. Sci. Pollut. Res. 2019, 26, 28749–28762. [Google Scholar] [CrossRef]
- Hussein, M.M.A.; Gad, E.; Ahmed, M.M.; Arisha, A.H.; Mahdy, H.F.; Swelum, A.A.A.; Tukur, H.A.; Saadeldin, I.M. Amelioration of titanium dioxide nanoparticle reprotoxicity by the antioxidants morin and rutin. Environ. Sci. Pollut. Res. 2019, 26, 29074–29084. [Google Scholar] [CrossRef]
- Salimnejad, R.; Soleimani Rad, J.; Nejad, D. Protective Effect of Ghrelin on Oxidative Stress and Tissue Damages of Mice Testes Followed By Chemotherapy With Cyclophosphamide. Crescent J. Med. Biol. Sci. 2018, 5, 138–143. [Google Scholar]
- Ekeleme-Egedigwe, C.A.; Famurewa, A.C.; David, E.E.; Eleazu, C.O.; Egedigwe, U.O. Antioxidant potential of garlic oil supplementation prevents cyclophosphamide-induced oxidative testicular damage and endocrine depletion in rats. J. Nutr. Intermed. Metab. 2019, 18, 100109. [Google Scholar] [CrossRef]
- Alkhalaf, M.I.; Alansari, W.S.; Alshubaily, F.A.; Alnajeebi, A.M.; Eskandrani, A.A.; Tashkandi, M.A.; Babteen, N.A. Chemoprotective effects of inositol hexaphosphate against cyclophosphamide-induced testicular damage in rats. Sci. Rep. 2020, 10, 12599. [Google Scholar] [CrossRef]
- Shokoohi, M.; Khaki, A.; Shoorei, H.; Khaki, A.A.; Moghimian, M.; Abtahi-Eivary, S.H. Hesperidin attenuated apoptotic-related genes in testicle of a male rat model of varicocoele. Andrology 2020, 8, 249–258. [Google Scholar] [CrossRef]
- Saber, T.M.; Arisha, A.H.; Abo-Elmaaty, A.M.A.; Abdelgawad, F.E.; Metwally, M.M.M.; Saber, T.; Mansour, M.F. Thymol alleviates imidacloprid-induced testicular toxicity by modulating oxidative stress and expression of steroidogenesis and apoptosis-related genes in adult male rats. Ecotoxicol. Environ. Saf. 2021, 221, 112435. [Google Scholar] [CrossRef]
- Saber, T.M.; Mansour, M.F.; Abdelaziz, A.S.; Mohamed, R.M.S.; Fouad, R.A.; Arisha, A.H. Argan oil ameliorates sodium fluoride-induced renal damage via inhibiting oxidative damage, inflammation, and intermediate filament protein expression in male rats. Environ. Sci. Pollut. Res. Int. 2020, 27, 30426–30436. [Google Scholar] [CrossRef]
- Iqubal, A.; Syed, M.A.; Najmi, A.K.; Ali, J.; Haque, S.E. Ameliorative effect of nerolidol on cyclophosphamide-induced gonadal toxicity in Swiss Albino mice: Biochemical-, histological- and immunohistochemical-based evidences. Andrologia 2020, 52, e13535. [Google Scholar] [CrossRef]
- Hamzeh, M.; Hosseinimehr, S.J.; Karimpour, A.; Mohammadi, H.R.; Khalatbary, A.R.; Talebpour Amiri, F. Cerium Oxide Nanoparticles Protect Cyclophosphamide-induced Testicular Toxicity in Mice. Int. J. Prev. Med. 2019, 10, 5. [Google Scholar] [CrossRef]
- Anan, H.H.; Zidan, R.A.; Abd El-Baset, S.A.; Ali, M.M. Ameliorative effect of zinc oxide nanoparticles on cyclophosphamide induced testicular injury in adult rat. Tissue Cell 2018, 54, 80–93. [Google Scholar] [CrossRef]
- Afkhami-Ardakani, M.; Hasanzadeh, S.; Shahrooz, R.; Delirezh, N.; Malekinejad, H. Antioxidant effects of Spirulina platensis (Arthrospira platensis) on cyclophosphamide-induced testicular injury in rats. Vet. Res. Forum: Int. Q. J. 2018, 9, 35–41. [Google Scholar]
- Cengiz, M.; Sahinturk, V.; Yildiz, S.C.; Şahin, İ.K.; Bilici, N.; Yaman, S.O.; Altuner, Y.; Appak-Baskoy, S.; Ayhanci, A. Cyclophosphamide induced oxidative stress, lipid per oxidation, apoptosis and histopathological changes in rats: Protective role of boron. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. (GMS) 2020, 62, 126574. [Google Scholar] [CrossRef]
- Torabi, F.; Malekzadeh Shafaroudi, M.; Rezaei, N. Combined protective effect of zinc oxide nanoparticles and melatonin on cyclophosphamide-induced toxicity in testicular histology and sperm parameters in adult Wistar rats. Int. J. Reprod. Biomed. 2017, 15, 403–412. [Google Scholar] [CrossRef]
- Nakazawa, H.; Chang, K.; Shinozaki, S.; Yasukawa, T.; Ishimaru, K.; Yasuhara, S.; Yu, Y.M.; Martyn, J.A.; Tompkins, R.G.; Shimokado, K.; et al. iNOS as a Driver of Inflammation and Apoptosis in Mouse Skeletal Muscle after Burn Injury: Possible Involvement of Sirt1 S-Nitrosylation-Mediated Acetylation of p65 NF-κB and p53. PLoS ONE 2017, 12, e0170391. [Google Scholar] [CrossRef] [PubMed]
- Salama, R.M.; Abd Elwahab, A.H.; Abd-Elgalil, M.M.; Elmongy, N.F.; Schaalan, M.F. LCZ696 (sacubitril/valsartan) protects against cyclophosphamide-induced testicular toxicity in rats: Role of neprilysin inhibition and lncRNA TUG1 in ameliorating apoptosis. Toxicology 2020, 437, 152439. [Google Scholar] [CrossRef] [PubMed]
- Hamza, A.A.; Elwy, H.M.; Badawi, A.M. Fenugreek seed extract attenuates cisplatin-induced testicular damage in Wistar rats. Andrologia 2016, 48, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, J.A.; Akinpade, T.G.; Olatunji, S.Y.; Owolabi, J.O.; Enya, J.I.; Adelodun, S.T.; Fabiyi, S.O.; Desalu, A.B. Observable Protective Activities of Quercetin on Aluminum Chloride-Induced Testicular Toxicity in Adult Male Wistar Rat. J. Hum. Reprod. Sci. 2021, 14, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Hassanein, E.H.M.; Ali, F.E.M.; Omar, Z.M.M.; Rashwan, E.K.; Mohammedsaleh, Z.M.; Abd El-Ghafar, O.A.M. Regulation of Keap-1/Nrf2/AKT and iNOS/NF-κB/TLR4 signals by apocynin abrogated methotrexate-induced testicular toxicity: Mechanistic insights and computational pharmacological analysis. Life Sci. 2021, 284, 119911. [Google Scholar] [CrossRef]
- Azouz, R.A.; Korany, R.M.S.; Noshy, P.A. Silica Nanoparticle-Induced Reproductive Toxicity in Male Albino Rats via Testicular Apoptosis and Oxidative Stress. Biol. Trace Elem. Res. 2022. [Google Scholar] [CrossRef]
- Famurewa, A.C.; Ekeleme-Egedigwe, C.A.; Onwe, C.S.; Egedigwe, U.O.; Okoro, C.O.; Egedigwe, U.J.; Asogwa, N.T. Ginger juice prevents cisplatin-induced oxidative stress, endocrine imbalance and NO/iNOS/NF-κB signalling via modulating testicular redox-inflammatory mechanism in rats. Andrologia 2020, 52, e13786. [Google Scholar] [CrossRef]
- Fouad, A.; Albuali, W.; Jresat, I. Protective Effect of Hesperidin against Cyclophosphamide Hepatotoxicity in Rats. Int. J. Bioeng. Life Sci. 2014, 8, 730–733. [Google Scholar]
- Siddiqi, A.; Hasan, S.K.; Nafees, S.; Rashid, S.; Saidullah, B.; Sultana, S. Chemopreventive efficacy of hesperidin against chemically induced nephrotoxicity and renal carcinogenesis via amelioration of oxidative stress and modulation of multiple molecular pathways. Exp. Mol. Pathol. 2015, 99, 641–653. [Google Scholar] [CrossRef]
- Filobbos, S.; Amin, N.; Yacoub, M.; Abd El_Hakim, K.R. Possible Protective Effect of Melatonin on Cisplatin-Induced Testicular Toxicity in Adult Albino Rats. A Histological and Immunohistochemical Study. Egypt. J. Histol. 2020, 43, 891–901. [Google Scholar] [CrossRef]
- Zickri, M.B.; Moustafa, M.H.; Fasseh, A.E.; Kamar, S.S. Antioxidant and antiapoptotic paracrine effects of mesenchymal stem cells on spermatogenic arrest in oligospermia rat model. Ann. Anat. Anat. Anz. Off. Organ Anat. Ges. 2021, 237, 151750. [Google Scholar] [CrossRef]
- Wang, Y.; Bai, L.; Zhang, J.; Li, H.; Yang, W.; Li, M. Lepidium draba L. leaves extract ameliorated cyclophosphamide-induced testicular toxicity by modulation of ROS-dependent Keap1/Nrf2/HO1, Bax/Bcl2/p53/caspase-3, and inflammatory signaling pathways. J. Food Biochem. 2021, 45, e13987. [Google Scholar] [CrossRef]
- Bashandy, S.A.E.; Ebaid, H.; Al-Tamimi, J.; Ahmed-Farid, O.A.; Omara, E.A.; Alhazza, I.M. Melatonin Alleviated Potassium Dichromate-Induced Oxidative Stress and Reprotoxicity in Male Rats. BioMed. Res. Int. 2021, 2021, 3565360. [Google Scholar] [CrossRef]
- Li, L.; Chen, B.; An, T.; Zhang, H.; Xia, B.; Li, R.; Zhu, R.; Tian, Y.; Wang, L.; Zhao, D.; et al. BaZiBuShen alleviates altered testicular morphology and spermatogenesis and modulates Sirt6/P53 and Sirt6/NF-κB pathways in aging mice induced by D-galactose and NaNO(2). J. Ethnopharmacol. 2021, 271, 113810. [Google Scholar] [CrossRef]
- Moradi, M.; Goodarzi, N.; Faramarzi, A.; Cheraghi, H.; Hashemian, A.H.; Jalili, C. Melatonin protects rats testes against bleomycin, etoposide, and cisplatin-induced toxicity via mitigating nitro-oxidative stress and apoptosis. Biomed. Pharmacother. = Biomed. Pharmacother. 2021, 138, 111481. [Google Scholar] [CrossRef]
- Li, S.; Che, S.; Chen, S.; Ruan, Z.; Zhang, L. Hesperidin partly ameliorates the decabromodiphenyl ether-induced reproductive toxicity in pubertal mice. Environ. Sci. Pollut. Res. Int. 2022, 29, 90391–90403. [Google Scholar] [CrossRef]
- Kaya, K.; Ciftci, O.; Cetin, A.; Doğan, H.; Başak, N. Hesperidin protects testicular and spermatological damages induced by cisplatin in rats. Andrologia 2015, 47, 793–800. [Google Scholar] [CrossRef]
- Celik, E.; Oguzturk, H.; Sahin, N.; Turtay, M.G.; Oguz, F.; Ciftci, O. Protective effects of hesperidin in experimental testicular ischemia/reperfusion injury in rats. Arch. Med. Sci. 2016, 12, 928–934. [Google Scholar] [CrossRef]
- Elwan, W.M.; Kassab, A.A. The Potential Protective Role of Hesperidin Against Capecitabine-Induced Corneal Toxicity in Adult Male Albino Rat. Light and Electron Microscopic Study. Egypt. J. Histol. 2017, 40, 201–215. [Google Scholar] [CrossRef]
- Turk, E.; Kandemir, F.M.; Yildirim, S.; Caglayan, C.; Kucukler, S.; Kuzu, M. Protective Effect of Hesperidin on Sodium Arsenite-Induced Nephrotoxicity and Hepatotoxicity in Rats. Biol. Trace Elem. Res. 2019, 189, 95–108. [Google Scholar] [CrossRef]
- El-Diasty, H.H.; El-Sayyad, H.; Refaat, S.; El-Ghaweet, H.A. Efficacy of Quercetin-Sensitized Cisplatin against N-Nitroso-NMethylurea Induced Testicular Carcinogenesis in Wistar Rats. Asian Pac. J. Cancer Prev. 2021, 22, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Adana, M.Y.; Imam, A.; Bello, A.A.; Sunmonu, O.E.; Alege, E.P.; Onigbolabi, O.G.; Ajao, M.S. Oral thymoquinone modulates cyclophosphamide-induced testicular toxicity in adolescent Wistar rats. Andrologia 2022, 54, e14368. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, M.; Tahir, A.; Samad, A.; Ashraf, A.; Ameen, M.; Imran, M.; Yousaf, S.; Sarwar, N. Casticin Alleviates Testicular and Spermatological Damage Induced by Cisplatin in Rats. Pak. Vet. J. 2020, 40, 234–238. [Google Scholar] [CrossRef]
- Delgarm, N.; Morovati-Sharifabad, M.; Salehi, E.; Afkhami-Ardakani, M.; Heydarnejad, M.S. Exploring the main effects of phoenix dactylifera on destructive changes caused by cyclophosphamide in male reproductive system in mice. Vet. Res. Forum Int. Q. J. 2022, 13, 249–255. [Google Scholar] [CrossRef]
- Bakhtiary, Z.; Shahrooz, R.; Ahmadi, A.; Soltanalinejad, F. Ethyl Pyruvate Ameliorates The Damage Induced by Cyclophosphamide on Adult Mice Testes. Int. J. Fertil. Steril. 2016, 10, 79–86. [Google Scholar] [CrossRef]
- Samie, A.; Sedaghat, R.; Baluchnejadmojarad, T.; Roghani, M. Hesperetin, a citrus flavonoid, attenuates testicular damage in diabetic rats via inhibition of oxidative stress, inflammation, and apoptosis. Life Sci. 2018, 210, 132–139. [Google Scholar] [CrossRef]
- Alanbaki, A.; Al-Mayali, H.; Al-Mayali, H. Ameliorative effect of Quercetin and Hesperidin on Antioxidant and Histological Changes in the Testis of Etoposide-Induced Adult Male Rats. Res. J. Pharm. Technol. 2018, 11, 564. [Google Scholar] [CrossRef]
- Jahan, S.; Iftikhar, N.; Ullah, H.; Rukh, G.; Hussain, I. Alleviative effect of quercetin on rat testis against arsenic: A histological and biochemical study. Syst. Biol. Reprod. Med. 2015, 61, 89–95. [Google Scholar] [CrossRef]
- Aboelwafa, H.R.; Ramadan, R.A.; El-Kott, A.F.; Abdelhamid, F.M. The protective effect of melatonin supplementation against taxol-induced testicular cytotoxicity in adult rats. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. Biol. 2022, 55, e11614. [Google Scholar] [CrossRef]
- Arisha, A.H.; Moustafa, A. Potential inhibitory effect of swimming exercise on the Kisspeptin–GnRH signaling pathway in male rats. Theriogenology 2019, 133, 87–96. [Google Scholar] [CrossRef]
- Osawe, S.O.; Farombi, E.O. Quercetin and rutin ameliorates sulphasalazine-induced spermiotoxicity, alterations in reproductive hormones and steroidogenic enzyme imbalance in rats. Andrologia 2018, 50, e12981. [Google Scholar] [CrossRef]
- Sharma, P.; Aslam Khan, I.; Singh, R. Curcumin and Quercetin Ameliorated Cypermethrin and Deltamethrin-Induced Reproductive System Impairment in Male Wistar Rats by Upregulating The Activity of Pituitary-Gonadal Hormones and Steroidogenic Enzymes. Int. J. Fertil. Steril. 2018, 12, 72–80. [Google Scholar] [CrossRef]
- Nayak, G.; Rao, A.; Mullick, P.; Mutalik, S.; Kalthur, S.G.; Adiga, S.K.; Kalthur, G. Ethanolic extract of Moringa oleifera leaves alleviate cyclophosphamide-induced testicular toxicity by improving endocrine function and modulating cell specific gene expression in mouse testis. J. Ethnopharmacol. 2020, 259, 112922. [Google Scholar] [CrossRef]
- Nna, V.U.; Ujah, G.A.; Suleiman, J.B.; Mohamed, M.; Nwokocha, C.; Akpan, T.J.; Ekuma, H.C.; Fubara, V.V.; Kekung-Asu, C.B.; Osim, E.E. Tert-butylhydroquinone preserve testicular steroidogenesis and spermatogenesis in cisplatin-intoxicated rats by targeting oxidative stress, inflammation and apoptosis. Toxicology 2020, 441, 152528. [Google Scholar] [CrossRef]
- Adelakun, S.A.; Akintunde, O.W.; Jeje, S.O.; Alao, O.A. Ameliorating and protective potential of 1-isothiocyanato-4-methyl sulfonyl butane on cisplatin induced oligozoospermia and testicular dysfunction via redox-inflammatory pathway: Histomorphometric and immunohistochemical evaluation using proliferating cell nuclear antigen. Phytomedicine Plus 2022, 2, 100268. [Google Scholar] [CrossRef]
- Noshy, P.A.; Khalaf, A.A.A.; Ibrahim, M.A.; Mekkawy, A.M.; Abdelrahman, R.E.; Farghali, A.; Tammam, A.A.; Zaki, A.R. Alterations in reproductive parameters and steroid biosynthesis induced by nickel oxide nanoparticles in male rats: The ameliorative effect of hesperidin. Toxicology 2022, 473, 153208. [Google Scholar] [CrossRef]
- Can, S.; Çetik Yıldız, S.; Keskin, C.; Şahintürk, V.; Cengiz, M.; Appak Başköy, S.; Ayhanci, A.; Akıncı, G. Investigation into the protective effects of Hypericum Triquetrifolium Turra seed against cyclophosphamide-induced testicular injury in Sprague Dawley rats. Drug Chem. Toxicol. 2022, 45, 1679–1686. [Google Scholar] [CrossRef]
- Ye, H.; Ng, H.W.; Sakkiah, S.; Ge, W.; Perkins, R.; Tong, W.; Hong, H. Pathway Analysis Revealed Potential Diverse Health Impacts of Flavonoids that Bind Estrogen Receptors. Int. J. Environ. Res. Public Health 2016, 13, 373. [Google Scholar] [CrossRef]
- Huang, Z.; Fang, F.; Wang, J.; Wong, C.-W. Structural activity relationship of flavonoids with estrogen-related receptor gamma. FEBS Lett. 2010, 584, 22–26. [Google Scholar] [CrossRef]
- Cannarella, R.; Calogero, A.E.; Condorelli, R.A.; Greco, E.A.; Aversa, A.; La Vignera, S. Is there a role for glucagon-like peptide-1 receptor agonists in the treatment of male infertility? Andrology 2021, 9, 1499–1503. [Google Scholar] [CrossRef]
- Starovlah, I.M.; Radovic Pletikosic, S.M.; Kostic, T.S.; Andric, S.A. Reduced spermatozoa functionality during stress is the consequence of adrenergic-mediated disturbance of mitochondrial dynamics markers. Sci. Rep. 2020, 10, 16813. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, D.M.; Alsemeh, A.E.; Khamis, T. Semaglutide early intervention attenuated testicular dysfunction by targeting the GLP-1-PPAR-α-Kisspeptin-Steroidogenesis signaling pathway in a testicular ischemia-reperfusion rat model. Peptides 2022, 149, 170711. [Google Scholar] [CrossRef] [PubMed]
- Almeer, R.S.; Abdel Moneim, A.E. Evaluation of the Protective Effect of Olive Leaf Extract on Cisplatin-Induced Testicular Damage in Rats. Oxid Med. Cell Longev. 2018, 2018, 8487248. [Google Scholar] [CrossRef] [PubMed]
- Gur, C.; Kandemir, O.; Kandemir, F.M. Investigation of the effects of hesperidin administration on abamectin-induced testicular toxicity in rats through oxidative stress, endoplasmic reticulum stress, inflammation, apoptosis, autophagy, and JAK2/STAT3 pathways. Environ. Toxicol. 2022, 37, 401–412. [Google Scholar] [CrossRef]
- Negm, W.A.; El-Kadem, A.H.; Hussein, I.A.; Alqahtani, M.J. The Mechanistic Perspective of Bilobetin Protective Effects against Cisplatin-Induced Testicular Toxicity: Role of Nrf-2/Keap-1 Signaling, Inflammation, and Apoptosis. Biomedicines 2022, 10, 1134. [Google Scholar] [CrossRef]
- Selvaraj, P.; Pugalendi, K.V. Efficacy of hesperidin on plasma, heart and liver tissue lipids in rats subjected to isoproterenol-induced cardiotoxicity. Exp. Toxicol. Pathol. Off. J. Ges. Fur Toxikol. Pathol. 2012, 64, 449–452. [Google Scholar] [CrossRef]
- Al-Shahat, A.; Hulail, M.A.E.; Soliman, N.M.M.; Khamis, T.; Fericean, L.M.; Arisha, A.H.; Moawad, R.S. Melatonin Mitigates Cisplatin-Induced Ovarian Dysfunction via Altering Steroidogenesis, Inflammation, Apoptosis, Oxidative Stress, and PTEN/PI3K/Akt/mTOR/AMPK Signaling Pathway in Female Rats. Pharmaceutics. 2022, 14, 2769. [Google Scholar] [CrossRef]
- Zirkin, B.R.; Chen, H. Regulation of Leydig cell steroidogenic function during aging. Biol. Reprod. 2000, 63, 977–981. [Google Scholar] [CrossRef]
- Moustafa, A. Effect of Light-Dark Cycle Misalignment on the Hypothalamic-Pituitary-Gonadal Axis, Testicular Oxidative Stress, and Expression of Clock Genes in Adult Male Rats. Int. J. Endocrinol. 2020, 2020, 1426846. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Layton, C. 12—Connective and other mesenchymal tissues with their stains. In Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Suvarna, S.K., Layton, C., Bancroft, J.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 153–175. [Google Scholar]
- Johnston, D.S.; Russell, L.D.; Friel, P.J.; Griswold, M.D. Murine germ cells do not require functional androgen receptors to complete spermatogenesis following spermatogonial stem cell transplantation. Endocrinology 2001, 142, 2405–2408. [Google Scholar] [CrossRef]
- Sharpe, R.M. Sperm counts and fertility in men: A rocky road ahead. Science & Society Series on Sex and Science. EMBO Rep. 2012, 13, 398–403. [Google Scholar] [CrossRef]
- Jensen, E.C. Quantitative analysis of histological staining and fluorescence using ImageJ. Anat. Rec. 2013, 296, 378–381. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′ to 3′) | Reverse Primer Sequence (5′ to 3′) | Product Size | Accession No. |
|---|---|---|---|---|
| GLP1 | CACCTCCTCTCAGCTCAGTC | CGTTCTCCTCCGTGTCTTGA | 128 | NM_012707.2 |
| Pparα | GTCCTCTGGTTGTCCCCTTG | GTCAGTTCACAGGGAAGGCA | 176 | NM_013196.2 |
| PGC1α | TTCAGGAGCTGGATGGCTTG | GGGCAGCACACTCTATGTCA | 70 | NM_031347.1 |
| Gapdh | GCATCTTCTTGTGCAGTGCC | GGTAACCAGGCGTCCGATAC | 91 | NM_017008.4 |
| Kiss-1 | TGCTGCTTCTCCTCTGTGTGG | ATTAACGAGTTCCTGGGGTCC | 110 | NM_181692.1 |
| Kiss-1r | CTTTCCTTCTGTGCTGCGTA | CCTGCTGGATGTAGTTGACG | 102 | NM_023992.1 |
| GnRH1 | AGGAGCTCTGGAACGTCTGAT | AGCGTCAATGTCACACTCGG | 100 | NM_012767.2 |
| GnRHr | TCAGGACCCACGCAAACTAC | CTGGCTCTGACACCCTGTTT | 182 | NM_031038.3 |
| StAr | CCCAAATGTCAAGGAAATCA | AGGCATCTCCCCAAAGTG | 187 | NM_031558.3 |
| CYP11A1 | AAGTATCCGTGATGTGGG | TCATACAGTGTCGCCTTTTCT | 127 | NM_017286.3 |
| CYP17A1 | TGGCTTTCCTGGTGCACAATC | TGAAAGTTGGTGTTCGGCTGAAG | 90 | NM_012753.2 |
| HSD17B3 | AGTGTGTGAGGTTCTCCCGGTACCT | TACAACATTGAGTCCATGTCTGGCCAG | 161 | NM_054007.1 |
| CYP19A1 | GCTGAGAGACGTGGAGACCTG | CTCTGTCACCAACAACAGTGTGG | 178 | NM_017085.2 |
| IL10 | GTAGAAGTGATGCCCCAGGC | AGAAATCGATGACAGCGTCG | 116 | NM_012854.2 |
| IL1β | CACCTCTCAAGCAGAGCACAGA | ACGGGTTCCATGGTGAAGTC | 81 | NM_031512.2 |
| P53 | GTTCGTGTTTGTGCCTGTCC | TGCTCTCTTTGCACTCCCTG | 108 | NM_030989.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khamis, T.; Hegazy, A.A.; El-Fatah, S.S.A.; Abdelfattah, E.R.; Abdelfattah, M.M.M.; Fericean, L.M.; Arisha, A.H. Hesperidin Mitigates Cyclophosphamide-Induced Testicular Dysfunction via Altering the Hypothalamic Pituitary Gonadal Axis and Testicular Steroidogenesis, Inflammation, and Apoptosis in Male Rats. Pharmaceuticals 2023, 16, 301. https://doi.org/10.3390/ph16020301
Khamis T, Hegazy AA, El-Fatah SSA, Abdelfattah ER, Abdelfattah MMM, Fericean LM, Arisha AH. Hesperidin Mitigates Cyclophosphamide-Induced Testicular Dysfunction via Altering the Hypothalamic Pituitary Gonadal Axis and Testicular Steroidogenesis, Inflammation, and Apoptosis in Male Rats. Pharmaceuticals. 2023; 16(2):301. https://doi.org/10.3390/ph16020301
Chicago/Turabian StyleKhamis, Tarek, Abdelmonem Awad Hegazy, Samaa Salah Abd El-Fatah, Eman Ramadan Abdelfattah, Marwa Mohamed Mahmoud Abdelfattah, Liana Mihaela Fericean, and Ahmed Hamed Arisha. 2023. "Hesperidin Mitigates Cyclophosphamide-Induced Testicular Dysfunction via Altering the Hypothalamic Pituitary Gonadal Axis and Testicular Steroidogenesis, Inflammation, and Apoptosis in Male Rats" Pharmaceuticals 16, no. 2: 301. https://doi.org/10.3390/ph16020301
APA StyleKhamis, T., Hegazy, A. A., El-Fatah, S. S. A., Abdelfattah, E. R., Abdelfattah, M. M. M., Fericean, L. M., & Arisha, A. H. (2023). Hesperidin Mitigates Cyclophosphamide-Induced Testicular Dysfunction via Altering the Hypothalamic Pituitary Gonadal Axis and Testicular Steroidogenesis, Inflammation, and Apoptosis in Male Rats. Pharmaceuticals, 16(2), 301. https://doi.org/10.3390/ph16020301