
Targeting Transient Receptor Potential (TRP) Channels, Mas-Related G-Protein-Coupled Receptors (Mrgprs), and Protease-Activated Receptors (PARs) to Relieve Itch

Abstract
:1. Introduction
2. The Transient Receptor Potential (TRP) Channels and Itch
2.1. TRP Cation Channel, Subfamily A, Member 1 (TRPA1) in Acute Itch
TRPA1 in Chronic Itch
2.2. TRP Cation Channel, Subfamily V (Vanilloid), Member 1 (TRPV1) in Acute Itch
TRPV1 in Chronic Itch
2.3. TRP Cation Channel, Subfamily V (Vanilloid), Member 2 (TRPV2)
2.4. TRP Cation Channel, Subfamily V (Vanilloid), Member 3 (TRPV3)
2.5. TRP Cation Channel, Subfamily V (Vanilloid), Member 4 (TRPV4)
2.6. TRP Cation Channel, Subfamily C (Canonical), Members 3,4 (TRPC3 and TRPC4)
2.7. TRP Cation Channel, Subfamily M (Melastatin), Member 8 (TRPM8)
3. G-Protein-Coupled Receptors (GPCRs) and Itch Sensation
4. Protease Activated Receptors (PARs) and Itch Sensation
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | atopic dermatitis |
| ACD | allergic contact dermatitis |
| AITC | allyl isothiocyanate |
| BAM8-22 | bovine adrenal medulla peptide |
| BNP | B-type natriuretic peptide |
| CQ | chloroquine |
| CA | cinnamaldehyde |
| DRG | dorsal root ganglion |
| GS | glucosylsphingosine |
| GPCRs | G-protein-coupled receptors |
| HR1, HR4 | histamine receptors |
| HDM | house dust mite |
| HMC | human mast cell (line) |
| ILs | interleukins |
| IMQ | imiquimod |
| IP3 | inositol triphosphate |
| KLK | kallikreins |
| KO | knockout (mice) |
| MOR | morphine |
| Mrgpr | Mas-related G protein-coupled receptor |
| mROS | mitochondrial reactive oxygen species |
| NGF | nerve growth factor |
| NK1R | neurokinin receptor 1 |
| NP | nonpeptidergic (receptors) |
| NPR1 | natriuretic pepdide receptor |
| OS | Olmsted syndrome |
| PAR | protease-activated receptor |
| PIP2 | phosphatidyl-inositol biphosphate |
| PLC | phospholipase C |
| PGE | prostaglandin E |
| SADBE | squaric acid dibutyl ester |
| SLIGRL | hexapeptide (Ser-Leu-Ile-Gly-Arg-Leu-NH2) |
| S1P | sphingosin 1-phosphate |
| S1PR3 | S1P receptor 3 |
| SP | substance P |
| TSLP | thymic stromal lymphopoietin |
| TRP | transient receptor potential (channels family) |
| TRPA | TRP ankyrin (subfamily) |
| TRPC | TRP canonical (subfamily) |
| TRPM | TRP melastatin (subfamily) |
| TRPV | TRP vanilloid (subfamily) |
| TrkA | tropomyosin receptor kinase A |
| TG | trigeminal ganglia |
| TNF | tumor necrosis factor |
References
- Halvorsen, J.A.; Dalgard, F.; Thoresen, M.; Bjertness, E.; Lien, L. Itch and pain in adolescents are associated with suicidal ideation: A population-based cross-sectional study. Acta Derm.-Venereol. 2012, 92, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, J.I.; Hanifin, J.M. Adult eczema prevalence and associations with asthma and other health and demographic factors: A US population–based study. J. Allergy Clin. Immunol. 2013, 132, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Drucker, A.M.; Wang, A.R.; Li, W.Q.; Sevetson, E.; Block, J.K.; Qureshi, A.A. The burden of atopic dermatitis: Summary of a report for the National Eczema Association. J. Investig. Dermatol. 2017, 137, 26–30. [Google Scholar] [CrossRef]
- Michalek, I.M.; Loring, B.; John, S.M. A systematic review of worldwide epidemiology of psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef]
- Krueger, G.G.; Bergstresser, P.R.; Lowe, N.J.; Voorhees, J.J.; Weinstein, G.D. Psoriasis. J. Am. Acad. Dermatol. 1984, 11, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Bickers, D.R.; Lim, H.W.; Margolis, D.; Weinstock, M.A.; Goodman, C.; Faulkner, E.; Gold, C.; Gemmen, E.; Dall, T. The burden of skin diseases: 2004: A joint project of the American Academy of Dermatology Association and the Society for Investigative Dermatology. J. Am. Acad. Dermatol. 2005, 55, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, K.E.; Florence, C.S.; Joski, P. Which medical conditions account for the rise in health care spending? Health Aff. 2004, 23 (Suppl. 1), Suppl Web Exclusives. W4–437–45. [Google Scholar] [CrossRef]
- Akiyama, T.; Carstens, E. Neural processing of itch. Neuroscience 2013, 250, 697–714. [Google Scholar] [CrossRef]
- Martins, M.S.; Almeida, I.F.; Cruz, M.T.; Sousa, E. Chronic pruritus: From pathophysiology to drug design. Biochem. Pharmacol. 2023, 212, 115568. [Google Scholar] [CrossRef]
- Tsagareli, M.G.; Carstens, E.E. Allodynia and hyperalgesia in pruritus: Involvement of TRP channels. In Histaminergic and Non-Histaminergic Itch; Tsagareli, M.G., Follansbee, T., Eds.; Nova Science: New York, NY, USA, 2023; Chapter 1; pp. 1–23. [Google Scholar]
- Vander Does, A.; Ju, T.; Mohsin, N.; Chopra, D.; Yosipovitch, G. How to get rid of itching. Pharmacol. Ther. 2023, 243, 108355. [Google Scholar] [CrossRef]
- Ständer, S.; Weisshaar, E.; Mettang, T.; Szepietowski, J.C.; Carstens, E.; Ikoma, A.; Bergasa, N.V.; Gieler, U.; Misery, L.; Wallengren, J.; et al. Clinical classification of itch: A position paper of the International Forum for the Study of Itch. Acta Derm.-Venereol. 2007, 87, 291–294. [Google Scholar] [CrossRef]
- Sun, S.; Dong, X. TRP channels and itch. Semin. Immunopathol. 2016, 38, 293–307. [Google Scholar] [CrossRef]
- Misery, L.; Pierre, O.; Le Gall-Ianotto, C.; Lebonvallet, N.; Chernyshov, P.V.; Le Garrec, R.; Talagas, M. Basic mechanisms of itch. J. Allergy Clin. Immunol. 2023, 152, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, O.; Oladipo, O.; Mahmoud, R.H.; Yosipovitch, G. Itch: From the skin to the brain—Peripheral and central neural sensitization in chronic itch. Front. Mol. Neurosci. 2023, 16, 1272230. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, O.; Soares, G.B.; Yosipovitch, G. Transient receptor potential channels and itch. Int. J. Mol. Sci. 2023, 24, 420. [Google Scholar] [CrossRef]
- Tsagareli, M.G.; Nozadze, I. An overview on transient receptor potential channels superfamily. Behav. Pharmacol. 2020, 31, 413–434. [Google Scholar] [CrossRef]
- Sanjel, B.; Shim, W.-S. Molecules that Channel stimulus into pruritus. In Histaminergic and Non-Histaminergic Itch; Tsagareli, M.G., Follansbee, T., Eds.; Nova Science: New York, NY, USA, 2023; Chapter 4; pp. 59–92. [Google Scholar]
- Denda, M.; Fuziwara, S.; Inoue, K.; Denda, S.; Akamatsu, H.; Tomitaka, A.; Matsunaga, K. Immunoreactivity of VR1 on Epidermal Keratinocyte of Human Skin. Biochem. Biophys. Res. Commun. 2001, 285, 1250–1252. [Google Scholar] [CrossRef]
- Southall, M.D.; Li, T.; Gharibova, L.S.; Pei, Y.; Nicol, G.D.; Travers, J.B. Activation of Epidermal Vanilloid Receptor-1 Induces Release of Proinflammatory Mediators in Human Keratinocytes. J. Pharmacol. Exp. Ther. 2003, 304, 217–222. [Google Scholar] [CrossRef]
- Staender, S.; Moormann, C.; Schumacher, M.; Buddenkotte, J.; Artuc, M.; Shpacovitch, V.; Brzoska, T.; Lippert, U.; Henz, B.M.; Luger, T.A.; et al. Expression of vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendage structures. Exp. Dermatol. 2004, 13, 129–139. [Google Scholar] [CrossRef]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The genetics of chronic itch: Gene expression in the skin of patients with atopic dermatitis and psoriasis with severe itch. J. Investig. Dermatol. 2018, 138, 1311–1317. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, Y.; Yu, W.; Li, J.; Yao, J.; Zhang, J.; Wang, J.; Wang, C. Transient receptor potential ankyrin 1 (TRPA1) modulators: Recent update and future perspective. Eur. J. Med. Chem. 2023, 257, 115392. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Gerhold, K.A.; Bifolck-Fisher, A.; Liu, Q.; Patel, K.N.; Dong, X.; Bautista, D.M. TRPA1 is required for histamine-independent, Mas-related G protein-coupled receptor-mediated itch. Nat. Neurosci. 2011, 14, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.Z.; Morita, T.; Brem, R.B.; Bautista, D.M. S1PR3 mediates itch and pain via distinct TRP channel-dependent pathways. J. Neurosci. 2018, 38, 7833–7843. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Nelson, A.M.; Batia, L.; Morita, T.; Estandian, D.; Owens, D.M.; Lumpkin, E.A.; Bautista, D.M. The ion channel TRPA1 is required for chronic itch. J. Neurosci. 2013, 33, 9283–9294. [Google Scholar] [CrossRef] [PubMed]
- Wilzopolski, J.; Kietzmann, M.; Mishra, S.K.; Stark, H.; Bäumer, W.; Rossbach, K. TRPV1 and TRPA1 channels are both involved downstream of histamine-induced itch. Biomolecules 2021, 11, 1166. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Li, X.Y. Inflammatory mediators causing cutaneous chronic itch in some diseases via transient receptor potential channel subfamily V member 1 and subfamily A member 1. J. Dermatol. 2019, 46, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Usoskin, D.; Nagi, S.S.; Hu, Y.; Kupari, J.; Bouchatta, O.; Cranfill, S.L.; Gautam, M.; Su, Y.; Lu, Y.; et al. Single-Soma Deep RNA Sequencing of Human Dorsal Root Ganglion Neurons Reveals Novel Molecular and Cellular Mechanisms Underlying Somatosensation. bioRxiv 2023. [Google Scholar] [CrossRef]
- Tsagareli, M.G.; Nozadze, I.; Tsiklauri, N.; Carstens, M.I.; Gurtskaia, G.; Carstens, E. Thermal hyperalgesia and mechanical allodynia elicited by histamine and non-histaminergic itch mediators: Respective involvement of TRPV1 and TRPA1. Neuroscience 2020, 449, 35–45. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.T.; Hu, Y.; Shan, W.Q.; Wang, Z.H.; Fu, Q.Y.; Fu, D.N.; Ji, J.; Liu, T. Formalin itch test: Low-dose formalin induces histamine-independent, TRPA1-mediated itch in mice. Front. Med. 2021, 8, 627725. [Google Scholar] [CrossRef]
- Lyons, J.J.; Milner, J.D.; Stone, K.D. Atopic dermatitis in children: Clinical features, pathophysiology, and treatment. Immunol. Allergy Clin. N. Am. 2015, 35, 161–183. [Google Scholar] [CrossRef]
- Shahwan, K.T.; Kimball, A.B. Itch intensity in moderate-to-severe plaque psoriasis versus atopic dermatitis: A meta-analysis. J. Am. Acad. Dermatol. 2017, 76, 1198–1200.e1. [Google Scholar] [CrossRef]
- Huet, F.; Faffa, M.S.; Poizeau, F.; Merhand, S.; Misery, L.; Brenaut, E. Characteristics of pruritus in relation to self-assessed severity of atopic dermatitis. Acta. Derm. Venereol. 2019, 99, 279–283. [Google Scholar] [CrossRef]
- Ziegler, S.F.; Roan, F.; Bell, B.D.; Stoklasek, T.A.; Kitajima, M.; Han, H. The biology of thymic stromal lymphopoietin (TSLP). Adv. Pharmacol. 2013, 66, 129–155. [Google Scholar] [CrossRef]
- Moore, C.; Gupta, R.; Jordt, S.E.; Chen, Y.; Liedtke, W.B. Regulation of pain and itch by TRP channels. Neurosci. Bull. 2017, 34, 120–142. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Thé, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hener, P.; Zhang, Z.; Kato, S.; Metzger, D.; Chambon, P. Topical vitamin D3 and low-calcemic analogs induce thymic stromal lymphopoietin in mouse keratinocytes and trigger an atopic dermatitis. Proc. Natl. Acad. Sci. USA 2006, 103, 11736–11741. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; McClain, S.P.; Batia, L.M.; Pellegrino, M.; Wilson, S.R.; Kienzler, M.A.; Lyman, K.; Olsen, A.S.B.; Wong, J.F.; Stucky, C.L.; et al. HTR7 mediates serotonergic acute and chronic itch. Neuron 2015, 87, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.-H.; Oh, S.Y.; Lu, J.; Lou, H.; Myers, A.C.; Zhu, Z.; Zheng, T. TRPA1-dependent pruritus in IL-13–induced chronic atopic dermatitis. J. Immunol. 2013, 191, 5371–5382. [Google Scholar] [CrossRef]
- Zeng, D.; Chen, C.; Zhou, W.; Ma, X.; Pu, X.; Zeng, Y.; Zhou, W.; Lv, F. TRPA1 deficiency alleviates inflammation of atopic dermatitis by reducing macrophage infiltration. Life Sci. 2021, 266, 118906. [Google Scholar] [CrossRef]
- Sanjel, B.; Kim, B.H.; Song, M.H.; Carstens, E.; Shim, W.S. Glucosylsphingosine evokes pruritus via activation of 5-HT2A receptor and TRPV4 in sensory neurons. Br. J. Pharmacol. 2022, 179, 2193–2207. [Google Scholar] [CrossRef] [PubMed]
- Afzal, R.; Shim, W.S. Activation of serotonin receptor 2 by glucosylsphingosine can be enhanced by TRPA1 but not TRPV1: Implication of a novel glucosylsphingosine-mediated itch pathway. Biochim. Biophys. Acta Biomembr. 2022, 1864, 184014. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Escalera, J.; Balakrishna, S.; Fan, L.; Caceres, A.I.; Robinson, E.; Sui, A.; McKay, M.C.; McAlexander, M.A.; Herrick, C.A.; et al. TRPA1 controls inflammation and pruritogen responses in allergic contact dermatitis. FASEB J. 2013, 27, 3549–3563. [Google Scholar] [CrossRef]
- Feng, J.; Yang, P.; Mack, M.R.; Dryn, D.; Luo, J.; Gong, X.; Liu, S.; Oetjen, L.K.; Zholos, A.V.; Mei, Z.; et al. Sensory TRP channels contribute differentially to skin inflammation and persistent itch. Nat. Comm. 2017, 8, 980. [Google Scholar] [CrossRef] [PubMed]
- Szepietowski, J.C.; Reich, A. Pruritus in psoriasis: An update. Eur. J. Pain 2016, 20, 41–46. [Google Scholar] [CrossRef]
- Calautti, E.; Avalle, L.; Poli, V. Psoriasis: A STAT3-centric view. Int. J. Mol. Sci. 2018, 19, 171. [Google Scholar] [CrossRef]
- Flutter, B.; Nestle, F.O. TLRs to Cytokines: Mechanistic insights from the imiquimod mouse model of psoriasis. Eur. J. Immunol. 2013, 43, 3138–3146. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, D.; Follansbee, T.; Wu, X.; Yu, S.; Wang, B.; Shi, Z.; Domocos, D.T.D.T.; Carstens, M.; Carstens, E.; et al. Transient receptor potential ankyrin 1 (TRPA1) positively regulates imiquimod-induced, psoriasiform dermal inflammation in mice. J. Cell. Mol. Med. 2019, 23, 4819–4828. [Google Scholar] [CrossRef]
- Follansbee, T.; Zhou, Y.; Wu, X.; Delahanty, J.; Nguyen, A.; Domocos, D.; Carstens, M.I.; Hwang, S.T.; Carstens, E. Signs of chronic itch in the mouse imiquimod model of psoriasiform dermatitis. Itch 2019, 4, e25. [Google Scholar] [CrossRef]
- Petrus, M.; Peier, A.M.; Bandell, M.; Hwang, S.W.; Huynh, T.; Olney, N.; Jegla, T.; Patapoutian, A. A role of TRPA1 in mechanical hyperalgesia is revealed by pharmacological inhibition. Mol. Pain 2007, 3, 40. [Google Scholar] [CrossRef]
- Wong, L.S.; Otsuka, A.; Yamamoto, Y.; Nonomura, Y.; Nakashima, C.; Kitayama, N.; Usui, K.; Honda, T.; Kabashima, K. TRPA1 channel participates in tacrolimus-induced pruritus in a chronic contact hypersensitivity murine model. J. Dermatol. Sci. 2018, 89, 207–209. [Google Scholar] [CrossRef]
- Ju, Y.; Luo, M.; Yan, T.; Zhou, Z.; Zhang, M.; Zhao, Z.; Liu, X.; Mei, Z.; Xiong, H. TRPA1 is involved in the inhibitory effect of Ke-Teng-Zi on allergic contact dermatitis via MAPK and JAK/STAT3 signaling pathways. J. Ethnopharmacol. 2023, 307, 116182. [Google Scholar] [CrossRef]
- Yang, N.; Shao, H.; Deng, J.; Yang, Y.; Tang, Z.; Wu, G.; Liu, Y. Dictamnine ameliorates chronic itch in DNFB-induced atopic dermatitis mice via inhibiting MrgprA3. Biochem. Pharmacol. 2023, 208, 115368. [Google Scholar] [CrossRef]
- Caterina, M.J. How do you feel? A warm and touching 2021 Nobel tribute. J. Clin. Investig. 2021, 31, e156587. [Google Scholar] [CrossRef]
- Kim, B.M.; Lee, S.H.; Shim, W.S.; Oh, U. Histamine-induced Ca(2+) influx via the PLA(2)/lipoxygenase/TRPV1 pathway in rat sensory neurons. Neurosci. Lett. 2004, 361, 159–162. [Google Scholar] [CrossRef]
- Imamachi, N.; Park, G.H.; Lee, H.; Anderson, D.J.; Simon, M.I.; Basbaum, A.I.; Han, S.K. TRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11330–11335. [Google Scholar] [CrossRef]
- Belghiti, M.; Estévez-Herrera, J.; Giménez-Garzó, C.; González-Usano, A.; Montoliu, C.; Ferrer-Montiel, A.; Felipo, V.; Planells-Cases, R. Potentiation of the transient receptor potential vanilloid 1 channel contributes to pruritogenesis in a rat model of liver disease. J. Biol. Chem. 2013, 288, 9675–9685. [Google Scholar] [CrossRef] [PubMed]
- Shirolkar, P.; Mishra, S.K. Role of TRP ion channels in pruritus. Neurosci. Lett. 2022, 768, 136379. [Google Scholar] [CrossRef] [PubMed]
- Hui-Beckman, J.; Goleva, E.; Leung, D.Y.M.; Kim, B.E. The impact of temperature on the skin barrier and atopic dermatitis. Ann. Allergy Asthma Immunol. 2023, 131, P713–P719. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.W.; Seo, J.A.; Jang, W.H.; Koh, H.J.; Bae, I.H.; Park, Y.H.; Lim, K.M. Antipruritic effects of TRPV1 antagonist in murine atopic dermatitis and itching models. J. Investig. Dermatol. 2011, 131, 1576–1579. [Google Scholar] [CrossRef]
- Yun, J.W.; Seo, J.A.; Jeong, Y.S.; Bae, I.H.; Jang, W.H.; Lee, J.; Kim, S.Y.; Shin, S.S.; Woo, B.Y.; Lee, K.W.; et al. TRPV1 antagonist can suppress the atopic dermatitis-like symptoms by accelerating skin barrier recovery. J. Dermatol. Sci. 2011, 62, 8–15. [Google Scholar] [CrossRef]
- Metz, M.; Krause, K.; Maurer, M.; Magerl, M. Treatment of notalgia paraesthetica with an 8% capsaicin patch. Br. J. Dermatol. 2011, 165, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Ansari, A.; Weinstein, D.; Sami, N. Notalgia paresthetica: Treatment review and algorithmic approach. J. Dermatol. Treat. 2019, 31, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Carvajal, A.; Fernández-Ballester, G.; Ferrer-Montiel, A. TRPV1 in chronic pruritus and pain: Soft modulation as a therapeutic strategy. Front. Mol. Neurosci. 2022, 15, 930964. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Choi, C.S.; Bae, I.H.; Choi, J.K.; Park, Y.H.; Park, M. A novel, topical, nonsteroidal, TRPV1 antagonist, PAC-14028 cream improves skin barrier function and exerts anti-inflammatory action through modulating epidermal differentiation markers and suppressing Th2 cytokines in atopic dermatitis. J. Dermatol. Sci. 2018, 91, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Won, C.H.; Jung, K.; Nam, H.J.; Choi, G.; Park, Y.H.; Park, M.; Kim, B. Efficacy and safety of PAC-14028 cream–a novel, topical, nonsteroidal, selective TRPV 1 antagonist in patients with mild-to-moderate atopic dermatitis: A phase II b randomized trial. Br. J. Dermatol. 2019, 180, 1030–1038. [Google Scholar] [CrossRef]
- Park, C.W.; Kim, B.J.; Lee, Y.W.; Won, C.; Park, C.O.; Chung, B.Y.; Lee, D.H.; Jung, K.; Nam, H.-J.; Choi, G. Asivatrep, a TRPV1 antagonist, for the topical treatment of atopic dermatitis: Phase 3, randomized, vehicle-controlled study (CAPTAIN-AD). J. Allergy Clin. Immunol. 2022, 149, 1340–1347.e4. [Google Scholar] [CrossRef]
- Sideris, N.; Paschou, E.; Bakirtzi, K.; Kiritsi, D.; Papadimitriou, I.; Tsentemeidou, A.; Sotiriou, E.; Vakirlis, E. New and upcoming topical treatments for atopic dermatitis: A review of the literature. J. Clin. Med. 2022, 11, 4974. [Google Scholar] [CrossRef]
- Gunthorpe, M.J.; Hannan, S.L.; Smart, D.; Jerman, J.C.; Arpino, S.; Smith, G.D.; Brough, S.; Wright, J.; Egerton, J.; Lappin, S.C.; et al. Characterization of SB-705498, a potent and selective vanilloid receptor-1 (VR1/TRPV1) antagonist that inhibits the capsaicin-, acid-, and heat-mediated activation of the receptor. J. Pharmacol. Exp. Ther. 2007, 321, 1183–1192. [Google Scholar] [CrossRef]
- Gibson, R.A.; Robertson, J.; Mistry, H.; McCallum, S.; Fernando, D.; Wyres, M.; Yosipovitch, G. A randomized trial evaluating the effects of the TRPV1 antagonist SB705498 on pruritus induced by histamine, and cowhage challenge in healthy volunteers. PLoS ONE 2014, 9, e100610. [Google Scholar] [CrossRef]
- Zhang, D.; Spielmann, A.; Wang, L.; Ding, G.; Huang, F.; Gu, Q.; Schwarz, W. Mast-cell degranulation induced by physical stimuli involves the activation of transient-receptor-potential channel TRPV2. Physiol. Res. 2012, 61, 113–124. [Google Scholar] [CrossRef]
- Guo, Y.; Song, Y.; Liu, W.; Wang, T.; Ma, X.; Yu, Z. Novel insights into the role of keratinocytes-expressed TRPV3 in the skin. Biomolecules 2023, 13, 513. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Bíró, T.A. TR(i)P to pruritus research: Role of TRPV3 in inflammation and itch. J. Investig. Dermatol. 2009, 129, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Larkin, C.; Chen, W.; Szabó, I.L.; Shan, C.; Dajnoki, Z.; Szegedi, A.; Buhl, T.; Fan, Y.; O’Neill, S.; Walls, D.; et al. Novel insights into the TRPV3-mediated itch in atopic dermatitis. J. Allergy Clin. Immunol. 2021, 147, 1110–1114.e5. [Google Scholar] [CrossRef] [PubMed]
- Um, J.Y.; Kim, H.B.; Kim, J.C.; Park, J.S.; Lee, S.Y.; Chung, B.Y.; Park, C.W.; Kim, H.O. TRPV3 and itch: The role of TRPV3 in chronic pruritus according to clinical and experimental evidence. Int. J. Mol. Sci. 2022, 23, 14962. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Luo, A.; Kamau, P.M.; Takomthong, P.; Hu, J.; Boonyarat, C.; Luo, L.; Lai, R. A plant-derived TRPV3 inhibitor suppresses pain and itch. Br. J. Pharmacol. 2021, 178, 1669–1683. [Google Scholar] [CrossRef]
- Seo, S.H.; Kim, S.; Kim, S.E.; Chung, S.; Lee, S.E. Enhanced thermal sensitivity of TRPV3 in keratinocytes underlies heat-induced pruritogen release and pruritus in atopic dermatitis. J. Investig. Dermatol. 2020, 140, 2199–2209.e6. [Google Scholar] [CrossRef]
- Agarwala, M.K.; Georgs, R.; Pramanik, R.; McGrath, J.A. Olmsted syndrome in an Indian male with new de novo mutation in TRPV3. Br. J. Dermatol. 2016, 174, 209–211. [Google Scholar] [CrossRef]
- Liedtke, W. TRPV4 plays an evolutionary conserved role in the transduction of osmotic and mechanical stimuli in live animals. J. Physiol. 2005, 567 Pt 1, 53–58. [Google Scholar] [CrossRef]
- Chen, Y.; Fang, Q.; Wang, Z.; Zhang, J.Y.; MacLeod, A.; Hall, R.P.; Liedtke, W.B. Transient receptor potential vanilloid 4 ion channel functions as a pruriceptor in epidermal keratinocytes to evoke histaminergic itch. J. Biol. Chem. 2016, 291, 10252–10262. [Google Scholar] [CrossRef]
- Kim, S.; Barry, D.M.; Liu, X.Y.; Yin, S.; Munanairi, A.; Meng, Q.T.; Cheng, W.; Mo, P.; Wan, L.; Liu, S.B.; et al. Facilitation of TRPV4 by TRPV1 is required for itch transmission in some sensory neuron populations. Sci. Signal. 2016, 9, ra71. [Google Scholar] [CrossRef]
- Akiyama, T.; Ivanov, M.; Nagamine, M.; Davoodi, A.; Carstens, M.I.; Ikoma, A.; Cevikbas, F.; Kempkes, C.; Buddenkotte, J.; Steinhoff, M.; et al. Involvement of TRPV4 in serotonin-evoked scratching. J. Investig. Dermatol. 2016, 136, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Henry, G.; Chen, Y. Emerging role of transient receptor potential vanilloid 4 (TRPV4) ion channel in acute and chronic itch. Int. J. Mol. Sci. 2021, 22, 7591. [Google Scholar] [CrossRef]
- Domocos, D.; Follansbee, T.; Nguyen, A.; Nguyen, T.; Carstens, M.I.; Carstens, E. Cinnamaldehyde elicits itch behavior via TRPV1 and TRPV4 but not TRPA1. Itch 2020, 5, e36. [Google Scholar] [CrossRef]
- Luo, J.; Feng, J.; Yu, G.; Yang, P.; Mack, M.R.; Du, J.; Yu, W.; Qian, A.; Zhang, Y.; Liu, S.; et al. Transient receptor potential vanilloid 4-expressing macrophages and keratinocytes contribute differentially to allergic and nonallergic chronic itch. J. Allergy Clin. Immunol. 2018, 141, 608–619.e7. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Xiang, L.; Zheng, S.; Zhao, Y.; Qin, Y.; Zhang, L.; Zhou, L. Vitexin inhibits pain and itch behavior via modulating TRPV4 activity in mice. Biomed. Pharmacother. 2023, 165, 115101. [Google Scholar] [CrossRef]
- Yan, J.; Ye, F.; Ju, Y.; Wang, D.; Chen, J.; Zhang, X.; Yin, Z.; Wang, C.; Yang, Y.; Zhu, C.; et al. Cimifugin relieves pruritus in psoriasis by inhibiting TRPV4. Cell Calcium 2021, 97, 102429. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Limjunyawong, N.; Narang, C.; Jamaldeen, H.; Yu, S.; Patiram, S.; Nie, H.; Caterina, M.J.; Dong, X.; et al. Sensory neuron-expressed TRPC3 mediates acute and chronic itch. Pain 2023, 164, 98–110. [Google Scholar] [CrossRef]
- Lee, S.H.; Cho, P.S.; Tonello, R.; Lee, H.K.; Jang, J.H.; Park, G.Y.; Hwang, S.W.; Park, C.K.; Jung, S.J.; Berta, T. Peripheral serotonin receptor 2B and transient receptor potential channel 4 mediate pruritus to serotonergic antidepressants in mice. J. Allergy Clin. Immunol. 2018, 142, 1349–1352.e16. [Google Scholar] [CrossRef]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- Liu, Y.; Mikrani, R.; He, Y.; Faran Ashraf Baig, M.M.; Abbas, M.; Naveed, M.; Tang, M.; Zhang, Q.; Lim, C.; Zhou, X. TRPM8 channels: A review of distribution and clinical role. Eur. J. Pharmacol. 2020, 882, 173312. [Google Scholar] [CrossRef] [PubMed]
- Palkar, R.; Ongun, S.; Catich, E.; Li, N.; Borad, N.; Sarkisian, A.; McKemy, D.D. Cooling relief of acute and chronic itch requires TRPM8 channels and neurons. J. Investig. Dermatol. 2018, 138, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Kardon, A.P.; Polgár, E.; Hachisuka, J.; Snyder, L.M.; Cameron, D.; Savage, S.; Cai, X.; Karnup, S.; Fan, C.R.; Hemenway, G.M.; et al. Dynorphin acts as a neuromodulator to inhibit itch in the dorsal horn of the spinal cord. Neuron 2014, 82, 573–586. [Google Scholar] [CrossRef]
- Kang, S.Y.; Choi, M.G.; Wei, E.T.; Selescu, T.; Lee, S.Y.; Kim, J.C.; Chung, B.Y.; Park, C.W.; Kim, H.O. TRPM8 agonist (cryosim-1) gel for scalp itch: A randomised, vehicle-controlled clinical trial. J. Eur. Acad. Dermatol. Venereol. 2022, 36, e588–e589. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.J.; Kim, J.C.; Wei, E.T.; Selescu, T.; Chung, B.Y.; Park, C.W.; Kim, H.O. A randomized, vehicle-controlled clinical trial of a synthetic TRPM8 agonist (Cryosim-1) gel for itch. J. Am. Acad. Dermatol. 2021, 84, 869–871. [Google Scholar] [CrossRef]
- McNeil, B.D. Minireview: Mas-related G protein-coupled receptor X2 activation by therapeutic drugs. Neurosci. Lett. 2021, 751, 135746. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Han, S.; Zylka, M.J.; Simon, M.I.; Anderson, D.J. A diverse family of GPCRs expressed in specific subsets of nociceptive sensory neurons. Cell 2001, 106, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Dong, X. The role of the Mrgpr receptor family in itch. In Pharmacology of Itch; Cowan, A., Yosipovitch, G., Eds.; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 226, pp. 71–88. [Google Scholar] [CrossRef]
- McNeil, B.; Dong, X. Mrgprs as itch receptors. In Itch: Mechanisms and Treatment; Carstens, E., Akiyama, T., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014; Chapter 12; pp. 213–236. [Google Scholar]
- Serhan, N.; Cenac, N.; Basso, L.; Gaudenzio, N. Mas-related G protein-coupled receptors (Mrgprs)—Key regulators of neuroimmune interactions. Neurosci Lett. 2021, 749, 135724. [Google Scholar] [CrossRef]
- Liu, Q.; Tang, Z.; Surdenikova, L.; Kim, S.; Patel, K.N.; Kim, A.; Ru, F.; Guan, Y.; Weng, H.J.; Geng, Y.; et al. Sensory neuron-specific GPCR Mrgprs are itch receptors mediating chloroquine-induced pruritus. Cell 2009, 139, 1353–1365. [Google Scholar] [CrossRef]
- Steele, H.R.; Han, L. The signaling pathway and polymorphisms of Mrgprs. Neurosci Lett. 2021, 744, 135562. [Google Scholar] [CrossRef]
- Inclan-Rico, J.M.; Kim, B.S.; Abdus-Saboor, I. Beyond somatosensation: Mrgprs in mucosal tissues. Neurosci. Lett. 2021, 748, 135689. [Google Scholar] [CrossRef]
- Dvorak, A.M. New aspects of mast cell biology. Int. Arch. Allergy Immunol. 1997, 114, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Holgate, S.T. The role of mast cells and basophils in inflammation. Clin. Exp. Allergy 2000, 30 (Suppl. 1), 28–32. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Alysandratos, K.-D.; Angelidou, A.; Delivanis, D.-A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Wen, Z.; Miniati, A.; et al. Mast cells and inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 21–33. [Google Scholar] [CrossRef] [PubMed]
- McNeil, B.D.; Pundir, P.; Meeker, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature 2015, 519, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Meixiong, J.; Vasavda, C.; Green, D.; Zheng, Q.; Qi, L.; Kwatra, S.G.; Hamilton, J.P.; Snyder, S.H.; Dong, X. Identification of a bilirubin receptor that may mediate a component of cholestatic itch. eLife 2019, 8, e44116. [Google Scholar] [CrossRef] [PubMed]
- Meixiong, J.; Vasavda, C.; Snyder, S.H.; Dong, X. MRGPRX4 is a G protein-coupled receptor activated by bile acids that may contribute to cholestatic pruritus. Proc. Natl. Acad. Sci. USA 2019, 116, 10525–10530. [Google Scholar] [CrossRef] [PubMed]
- Meixiong, J.; Anderson, M.; Limjunyawong, N.; Sabbagh, M.F.; Hu, E.; Mack, M.R.; Oetjen, L.K.; Wang, F.; Kim, B.S.; Dong, X. Activation of mast-cell-expressed Mas-related G-protein-coupled receptors drives non-histaminergic itch. Immunity 2019, 50, 1163–1171.e5. [Google Scholar] [CrossRef]
- Fujisawa, D.; Kashiwakura, J.-I.; Kita, H.; Kikukawa, Y.; Fujitani, Y.; Sasaki-Sakamoto, T.; Kuroda, K.; Nunomura, S.; Hayama, K.; Terui, T.; et al. Expression of Mas-related gene X2 on mast cells is upregulated in the skin of patients with severe chronic urticaria. J. Allergy Clin. Immunol. 2014, 134, 622–633.e9. [Google Scholar] [CrossRef]
- Shtessel, M.; Limjunyawong, N.; Oliver, E.T.; Chichester, K.; Gao, L.; Dong, X.; Saini, S.S. MRGPRX2 Activation Causes Increased Skin Reactivity in Patients with Chronic Spontaneous Urticaria. J. Investig. Dermatol. 2021, 141, 678–681.e2. [Google Scholar] [CrossRef]
- Ryu, K.; Heo, Y.; Lee, Y.; Jeon, H.; Namkung, W. Berbamine reduces chloroquine-induced itch in mice through inhibition of MrgprX1. Int. J. Mol. Sci. 2022, 23, 14321. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.B.; Iuga, A.O.; Shimada, S.G.; LaMotte, R.H.; Lerner, E.A. Cowhage-evoked itch is mediated by a novel cysteine protease: A ligand of protease-activated receptors. J. Neurosci. 2018, 28, 4331–4335. [Google Scholar] [CrossRef]
- Shelley, W.B.; Arthur, R.P. Mucunain, the active pruritogenic proteinase of cowhage. Science 1955, 122, 469–470. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Lerner, E.A.; Carstens, E. Protease-activated receptors and itch. Handb. Exp. Pharmacol. 2015, 226, 219–235. [Google Scholar] [CrossRef]
- Kempkes, C.; Buddenkotte, J.; Cevikbas, F.; Buhl, T.; Steinhoff, M. Role of PAR-2 in neuroimmune communication and itch. In Itch: Mechanisms and Treatment; Carstens, E., Akiyama, T., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014; Chapter 11; pp. 193–212. [Google Scholar]
- Buhl, T.; Ikoma, A.; Kempkes, C.; Cevikbas, F.; Sulk, M.; Buddenkotte, J.; Akiyama, T.; Crumrine, D.; Camerer, E.; Carstens, E.; et al. Protease-activated receptor-2 regulates neuro-epidermal communication in atopic dermatitis. Front. Immunol. 2020, 11, 1740. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.; Pitcher, T.; Grant, A.D.; Hewitt, E.; Lindstrom, E.; Malcangio, M. Cathepsin S acts via protease-activated receptor 2 to activate sensory neurons and induce itch-like behavior. Neurobiol. Pain 2019, 6, 100032. [Google Scholar] [CrossRef]
- Kim, H.I.; Lee, G.B.; Song, D.E.; Sanjel, B.; Lee, W.J.; Shim, W.S. FSLLRY-NH2, a protease-activated receptor 2 (PAR2) antagonist, activates mas-related G protein-coupled receptor C11 (MrgprC11) to induce scratching behaviors in mice. Life Sci. 2023, 325, 121786. [Google Scholar] [CrossRef]
- Cao, T.; Tan, E.S.; Chan, Y.H.; Yosipovitch, G.; Tey, H.L. Anti-pruritic efficacies of doxycycline and erythromycin in the treatment of acne vulgaris: A randomized single-blinded pilot study. Indian J. Dermatol. Venereol. Leprol. 2018, 84, 458–460. [Google Scholar] [CrossRef]
- Ishikawa, C.; Tsuda, T.; Konishi, H.; Nakagawa, N.; Yamanishi, K. Tetracyclines modulate protease-activated receptor 2-mediated proinflammatory reactions in epidermal keratinocytes. Antimicrob. Agents Chemother. 2009, 53, 1760–1765. [Google Scholar] [CrossRef]
- Cao, T.; Tan, W.D.; Kim, H.; Tey, H.L. Efficacy of a topical proteinase-activated receptor-2 inhibitor on cowhage-induced pruritus: A randomized placebo-controlled double-blind study. J. Eur. Acad. Dermatol. Venereol. 2017, 31, e191–e192. [Google Scholar] [CrossRef]
- Ruppenstein, A.; Limberg, M.M.; Loser, K.; Kremer, A.E.; Homey, B.; Raap, U. Involvement of neuro-immune interactions in pruritus with special focus on receptor expressions. Front. Med. 2021, 8, 627985. [Google Scholar] [CrossRef] [PubMed]
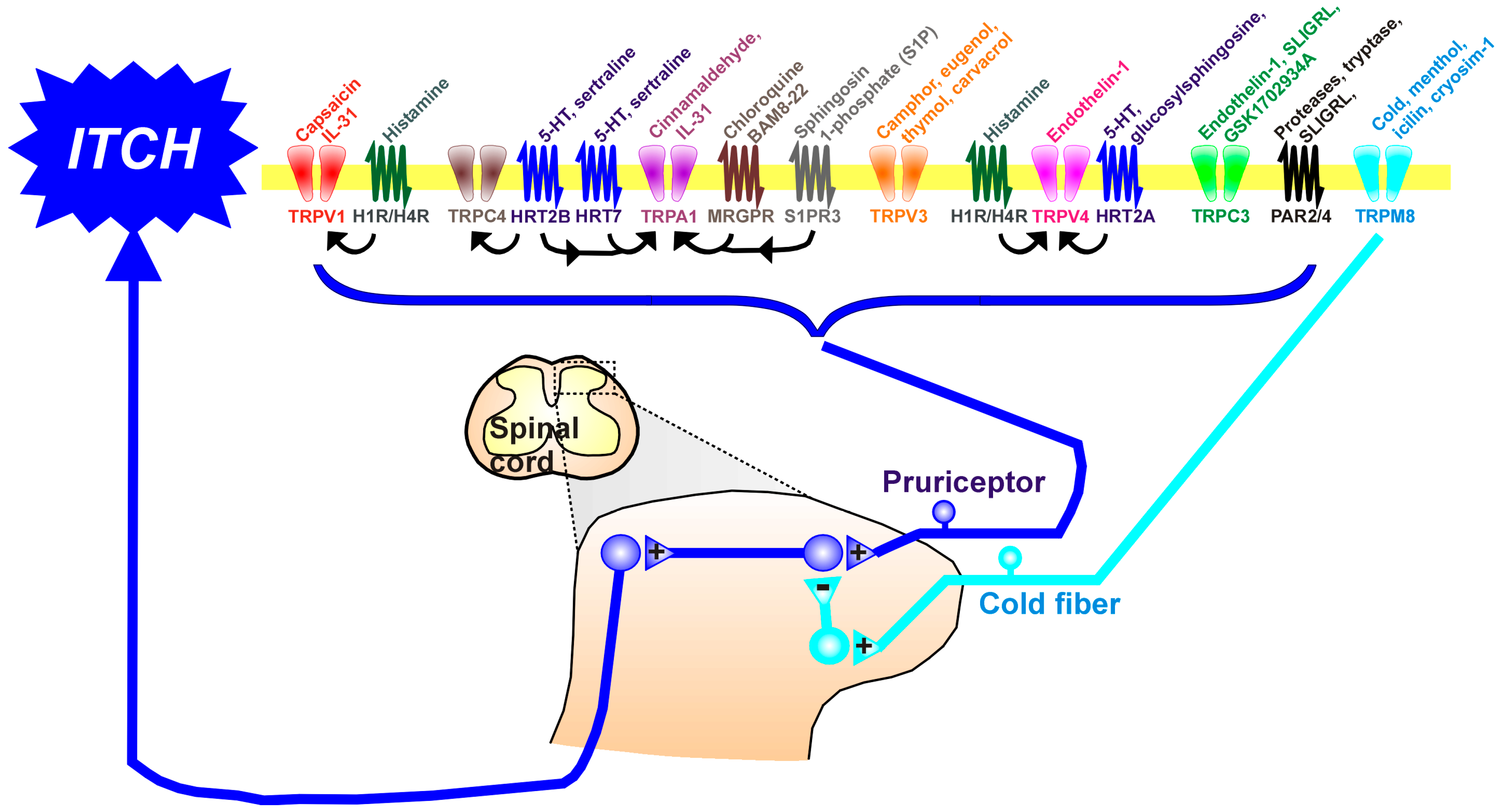
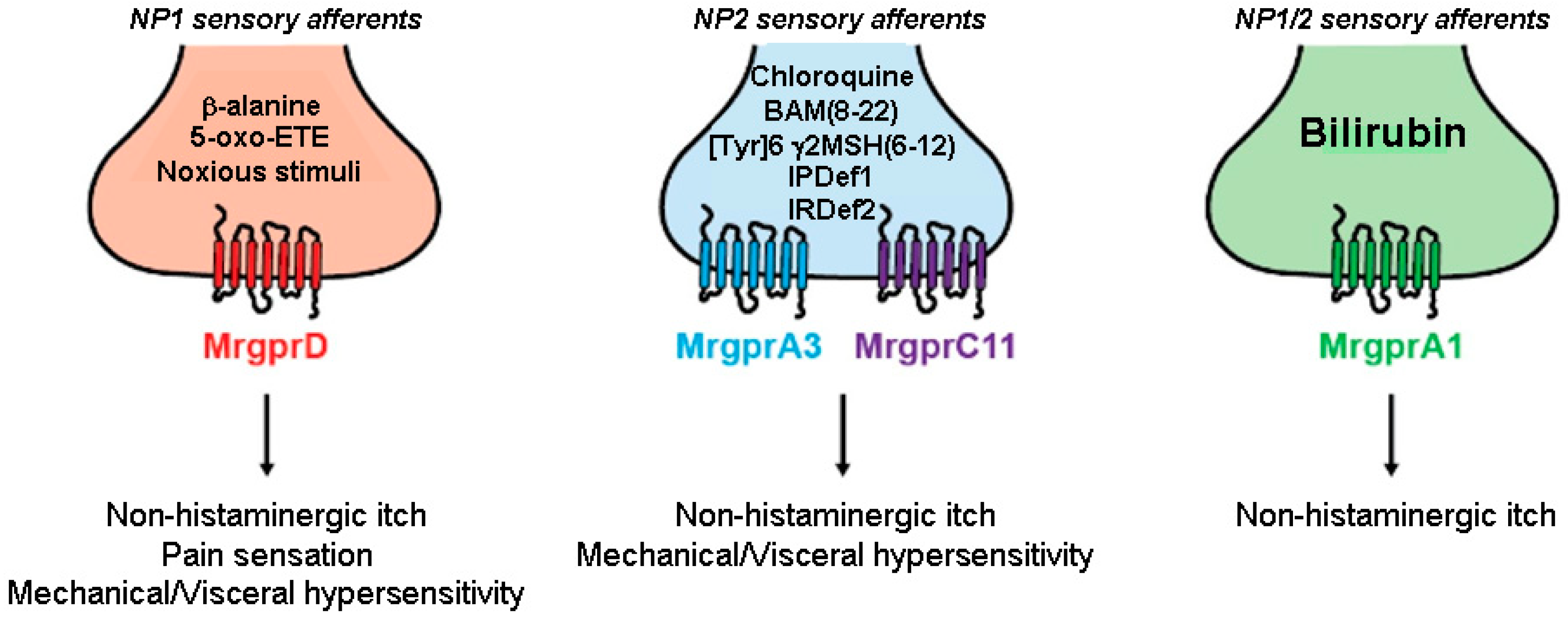
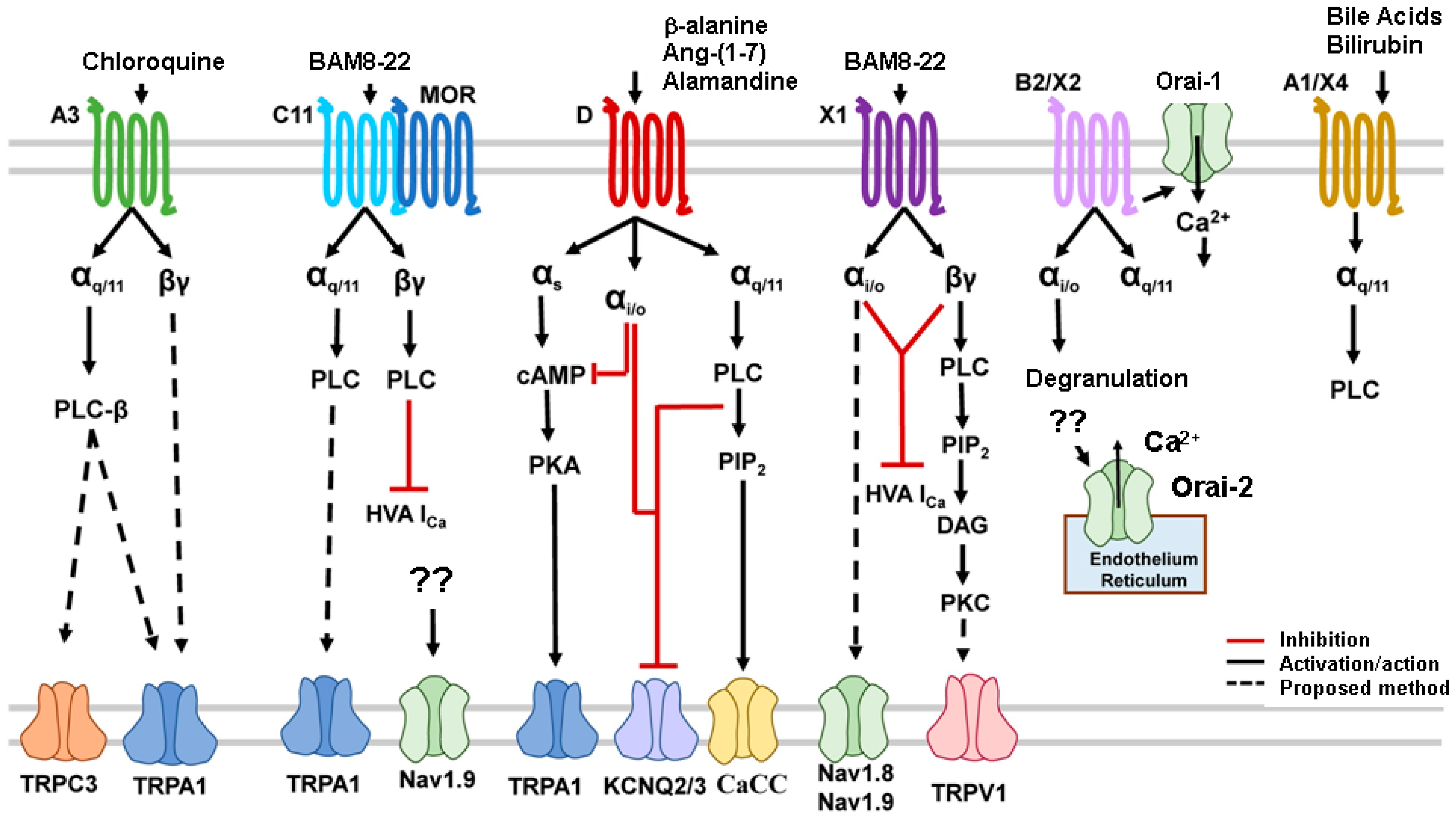

{kind=link}
{kind=link}
{kind=link}
| Itch Receptor | Agonists/ Activators | Antagonists/ Inhibitors | Disease Implications | Itch-Related Clinical Trial |
|---|---|---|---|---|
| TRPA1 | AITC, eugenol, cinnamaldehyde | HC-030031, A967079 | Non-histaminergic itch, Atopic dermatitis, allergic contact dermatitis, psoriasis | A967079-Completed |
| TRPV1 | Capsaicin, resiniferatoxin | AMG-517, (Asivatrep) PAC-14028, SB705498 | Histaminergic itch, atopic dermatitis, allergic contact dermatitis, | SB705498-Phase I, PAC-14028-Completed |
| TRPV2 | NA | SKF96365 | Atopic dermatitis | No ongoing trials |
| TRPV3 | Camphor, eugenol | Citrusinine-II, KM001 | Atopic dermatitis, Olmsted syndrome, Lichen simplex chronicus | KM-001-Phase I |
| TRPV4 | NA | Vitexin, cimifugin | Serotonergic itch, dry skin itch, contact dermatitis | No ongoing trials |
| TRPC3 | NA | NA | Non-histaminergic itch, | No ongoing trials |
| TRPC4 | Sertraline | ML204, HC-070 | Sertraline induced itch | No ongoing trials |
| TRPM8 | Menthol, cryosim-1 | Menthoxy-propanediol | Agonism functions as antipruritic, including scalp itch | No ongoing trials |
| MRGPRX2 | SP, PACAP | EP262 | Atopic dermatitis, chronic urticaria | EP262-Phase II |
| MRGPRX4 | Bile acids, Bilirubin | EP547 | Cholestatic itch, uremic pruritus | EP547-Phase II |
| MRGPRX1 | BAM8-22 | Berbamine | Chloroquine-induced itch | No ongoing trials |
| PAR2/4 | Mucunain, tryptase, hexapeptide cathepsin S, FSLLRY-NH2 | MMP, doxycycline | Atopic dermatitis, psoriasis, dry skin itch, contact dermatitis | Cowage-evoked pruritis, pruritic acne |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsagareli, M.G.; Follansbee, T.; Iodi Carstens, M.; Carstens, E. Targeting Transient Receptor Potential (TRP) Channels, Mas-Related G-Protein-Coupled Receptors (Mrgprs), and Protease-Activated Receptors (PARs) to Relieve Itch. Pharmaceuticals 2023, 16, 1707. https://doi.org/10.3390/ph16121707
Tsagareli MG, Follansbee T, Iodi Carstens M, Carstens E. Targeting Transient Receptor Potential (TRP) Channels, Mas-Related G-Protein-Coupled Receptors (Mrgprs), and Protease-Activated Receptors (PARs) to Relieve Itch. Pharmaceuticals. 2023; 16(12):1707. https://doi.org/10.3390/ph16121707
Chicago/Turabian StyleTsagareli, Merab G., Taylor Follansbee, Mirela Iodi Carstens, and Earl Carstens. 2023. "Targeting Transient Receptor Potential (TRP) Channels, Mas-Related G-Protein-Coupled Receptors (Mrgprs), and Protease-Activated Receptors (PARs) to Relieve Itch" Pharmaceuticals 16, no. 12: 1707. https://doi.org/10.3390/ph16121707
APA StyleTsagareli, M. G., Follansbee, T., Iodi Carstens, M., & Carstens, E. (2023). Targeting Transient Receptor Potential (TRP) Channels, Mas-Related G-Protein-Coupled Receptors (Mrgprs), and Protease-Activated Receptors (PARs) to Relieve Itch. Pharmaceuticals, 16(12), 1707. https://doi.org/10.3390/ph16121707