
Piperine Reduces Neoplastic Progression in Cervical Cancer Cells by Downregulating the Cyclooxygenase 2 Pathway


 , and
, and
Abstract
1. Introduction
2. Results
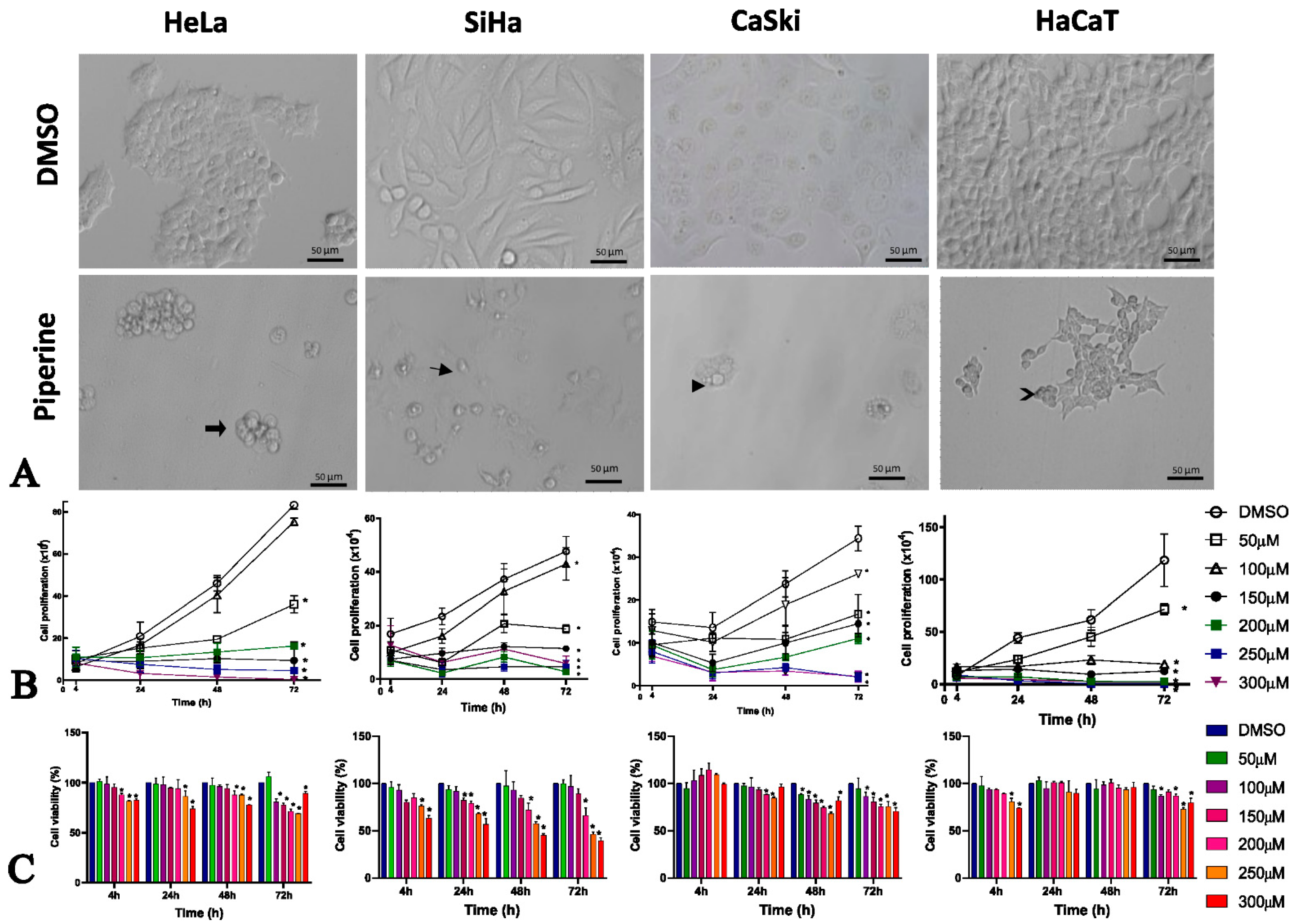
2.1. Piperine Alters Morphology, Viability, and Cell Proliferation
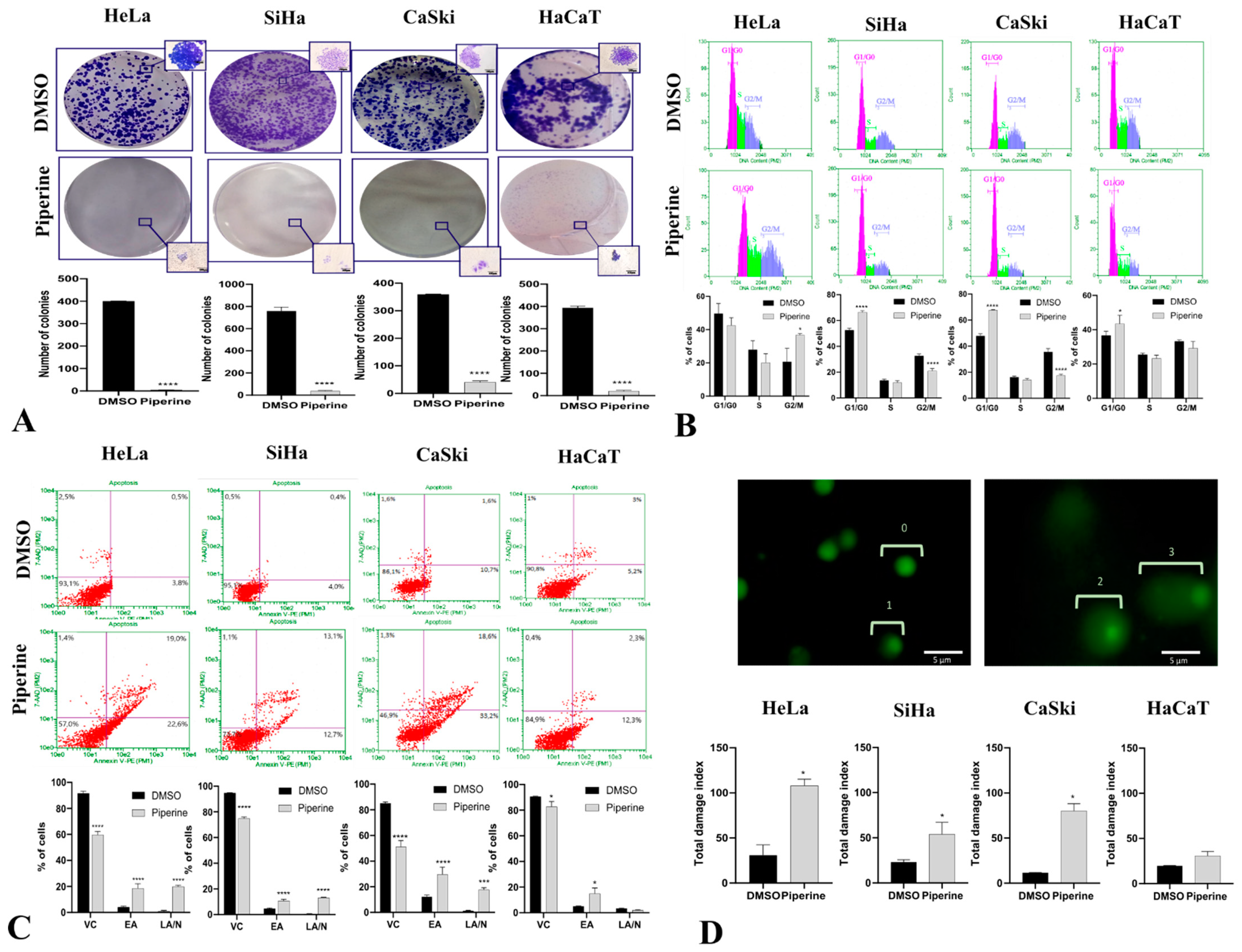
2.2. Piperine Inhibits Colony Formation, Impairs Cell Cycle Progression, and Induces Apoptosis by DNA Fragmentation
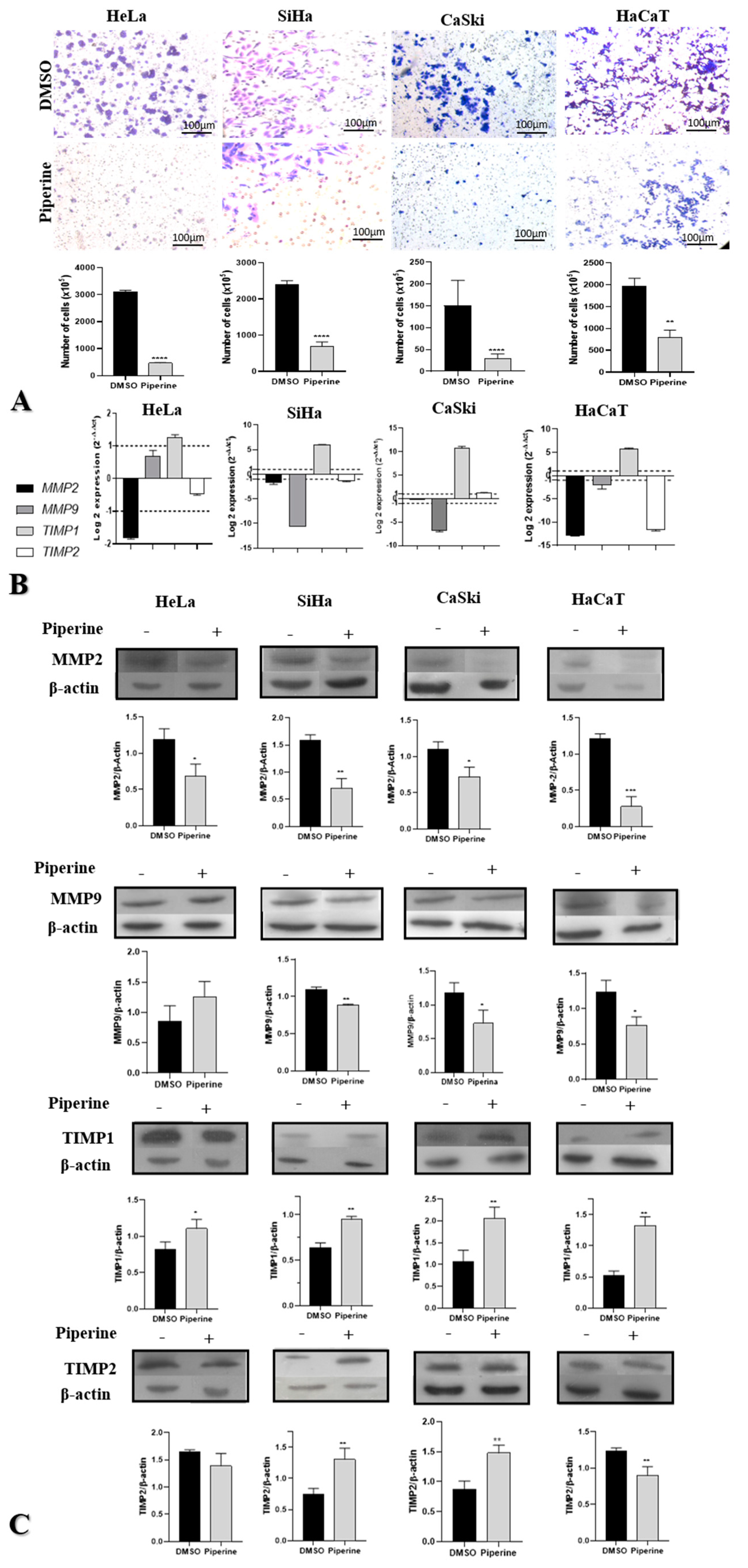
2.3. Piperine Reduces Cell Migration in Cervical Cancer through Regulation of Matrix Metalloproteinases and Their Antagonists
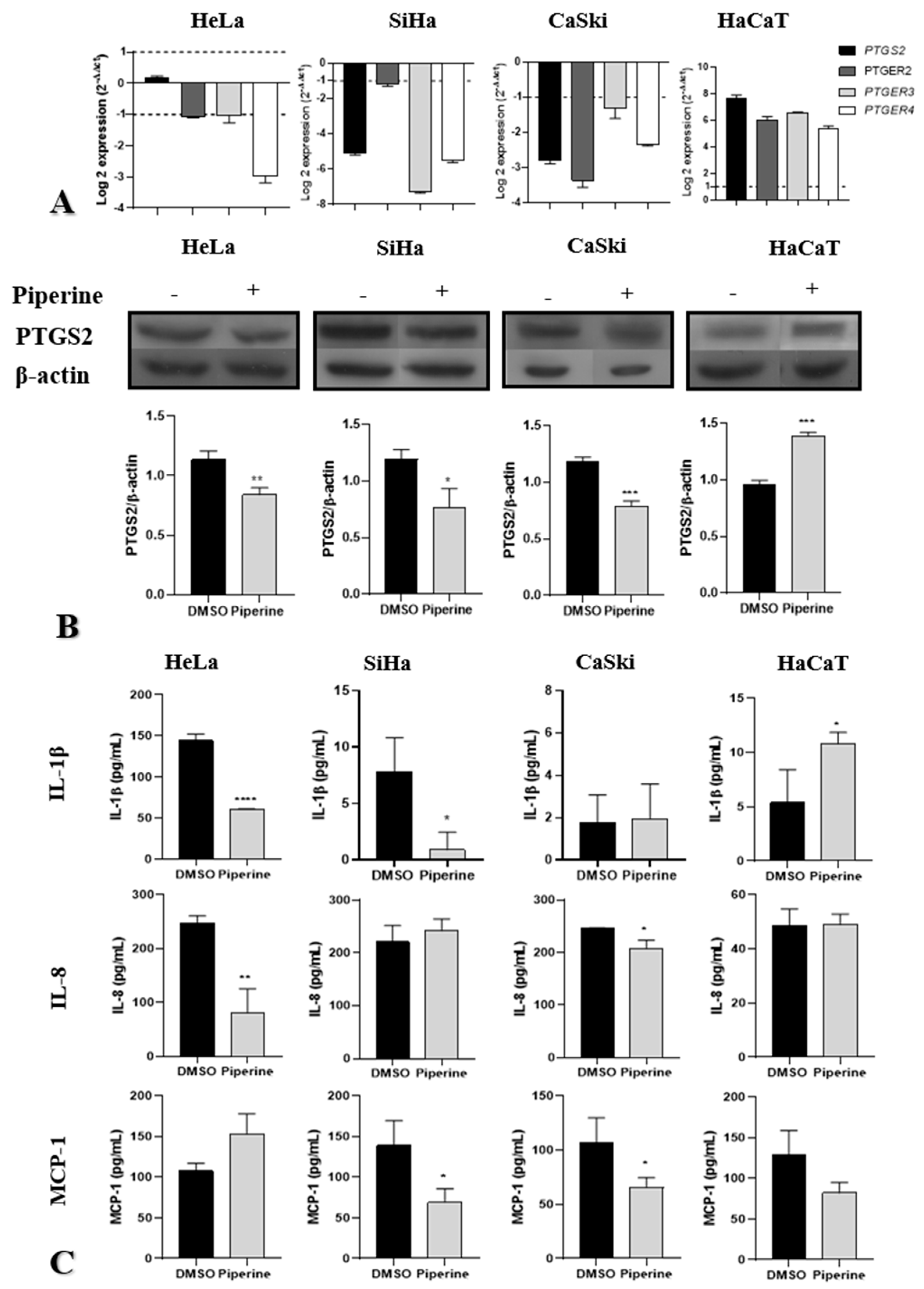
2.4. Piperine Reduces Tumorigenesis by Regulating the PTGS2 Inflammatory Pathway and Cytokine Secretion
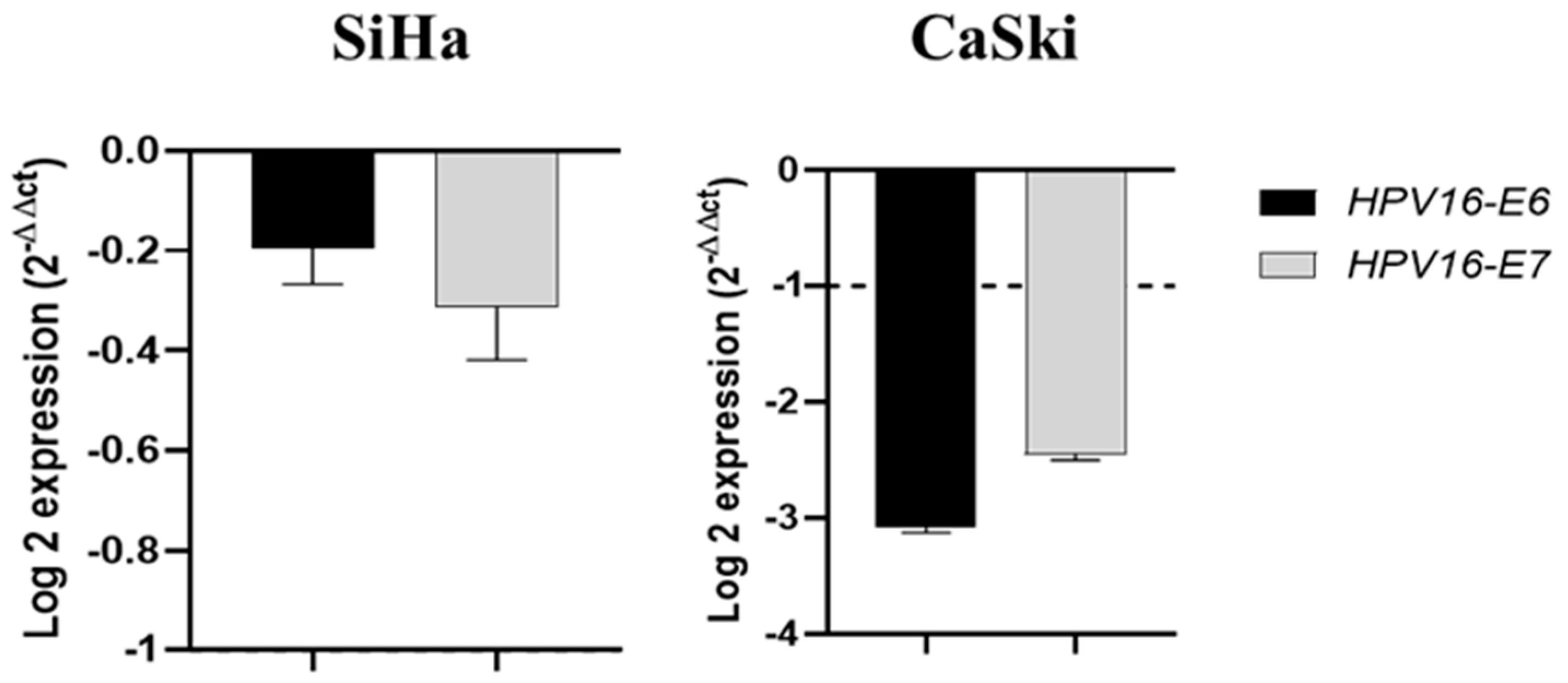
2.5. Piperine Reduces the Expression of HPV16 Oncogenes in CaSki Cells
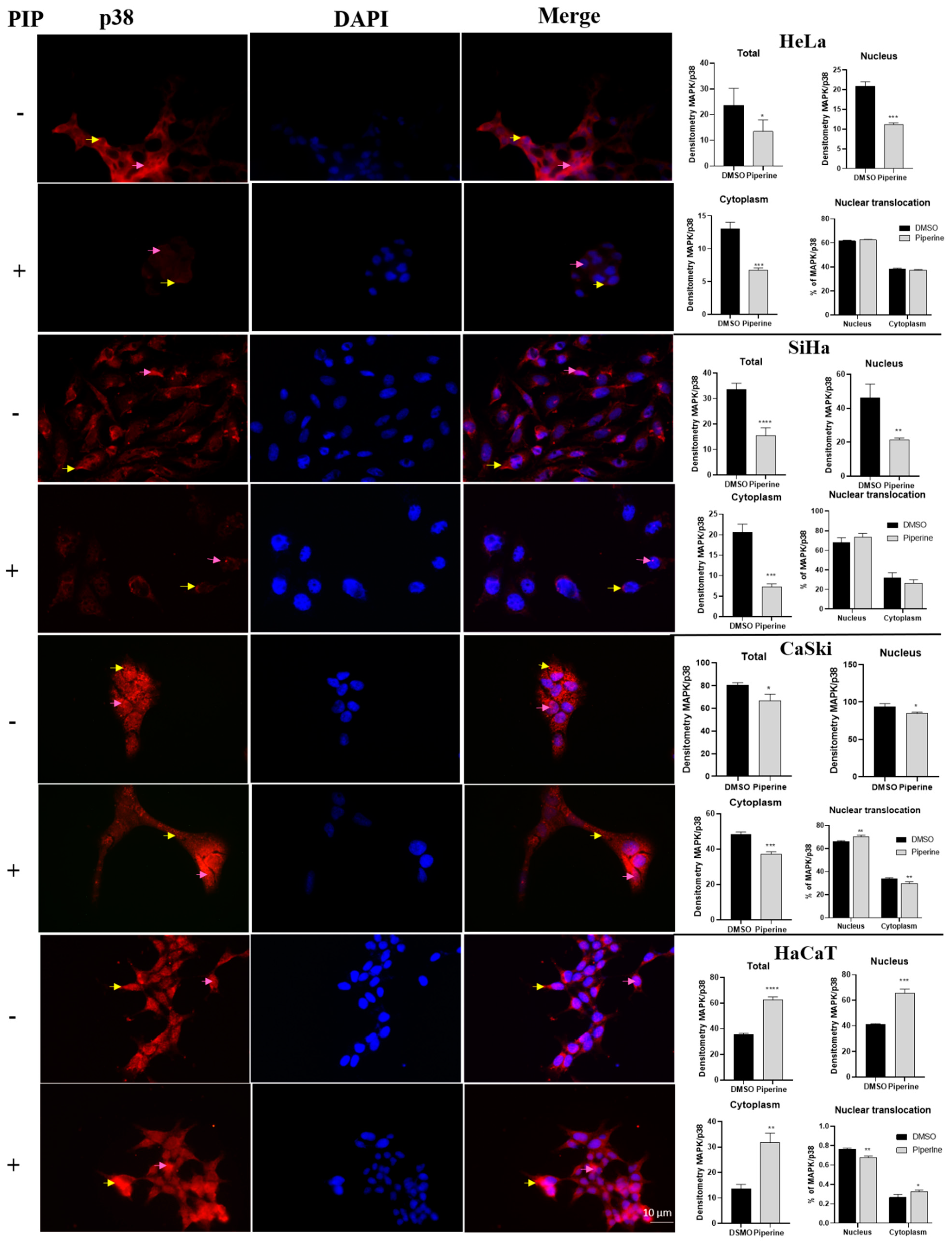
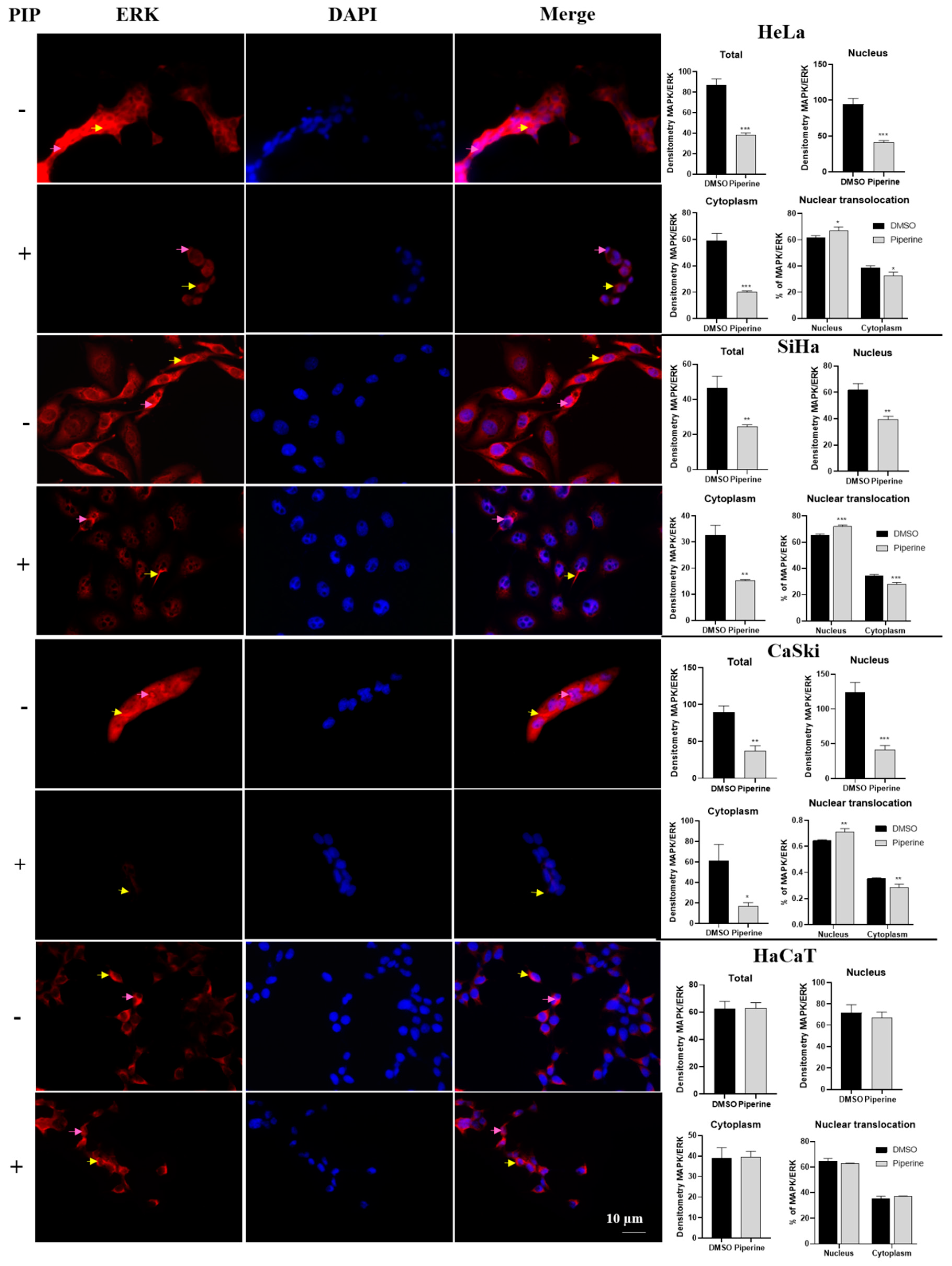
2.6. Piperine Modulates the Expression of p38 and ERK (MAPKs)
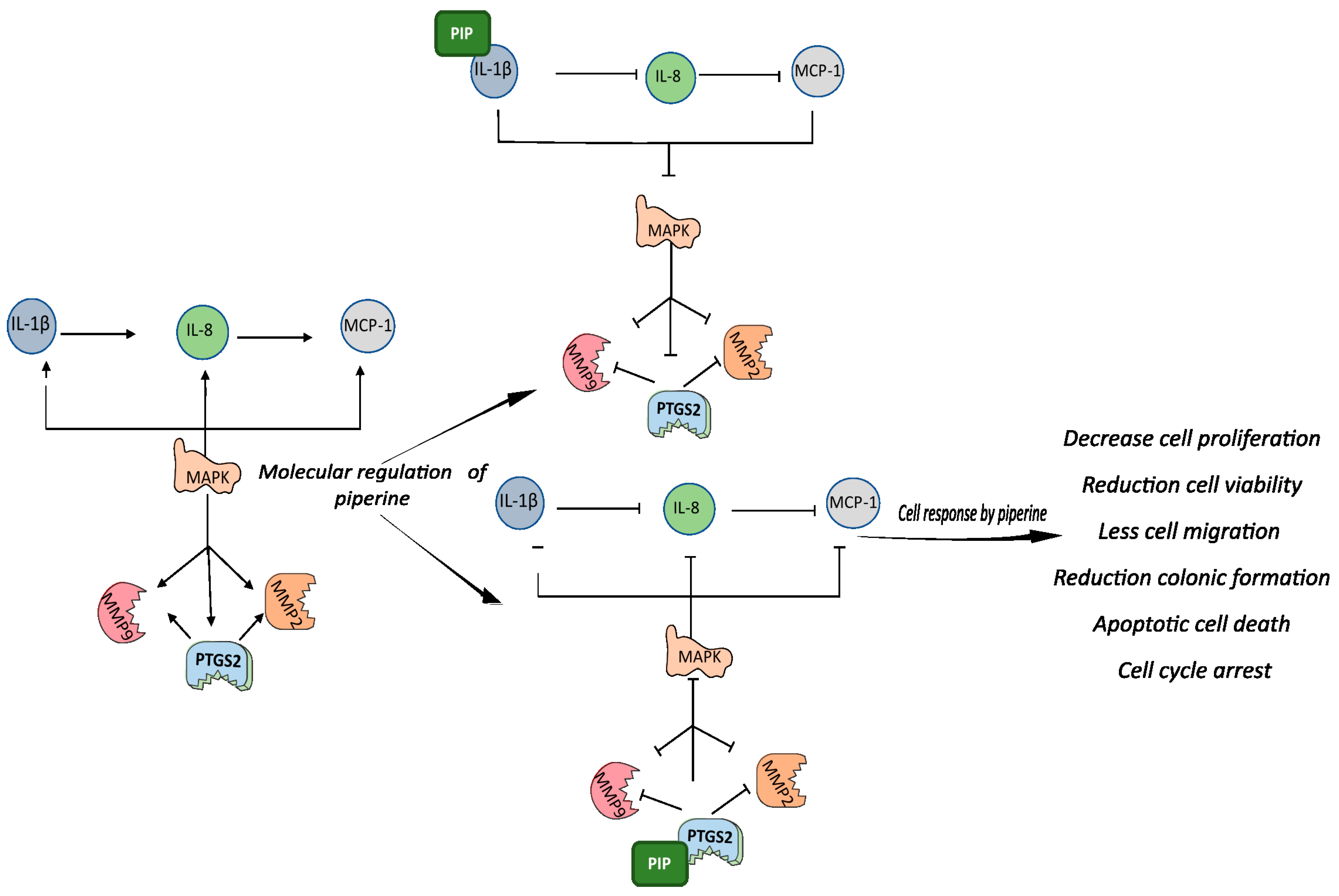
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Treatment
4.3. Cultivation and Analysis of Cell Morphology
4.4. Cell Proliferation
4.5. Cytotoxicity and Cell Viability Assay
4.6. Cell Migration Assay
4.7. Colony Formation Assay in Liquid Medium
4.8. Alkaline Comet Assay
4.9. Apoptosis and Cell Cycle Analysis
4.10. RNA Extraction and Quantification
4.11. Real-Time PCR Analysis
4.12. Protein Extraction and Quantification
4.13. Western Blotting
4.14. ELISA
4.15. Immunocytochemistry
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Werness, B.A.; Levine, A.J.; Howley, P.M. Association of Human Papillomavirus Types 16 and 18 E6 Proteins with P53. Science 1990, 248, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Ghittoni, R.; Accardi, R.; Hasan, U.; Gheit, T.; Sylla, B.; Tommasino, M. The Biological Properties of E6 and E7 Oncoproteins from Human Papillomaviruses. Virus Genes 2010, 40, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Senba, M.; Mori, N. Mechanisms of Virus Immune Evasion Lead to Development from Chronic Inflammation to Cancer Formation Associated with Human Papillomavirus Infection. Oncol. Rev. 2012, 6, 135–144. [Google Scholar] [CrossRef]
- Liu, J.; Cao, X. Cellular and Molecular Regulation of Innate Inflammatory Responses. Cell. Mol. Immunol. 2016, 13, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Baby, D.; Rajguru, J.; Patil, P.; Thakkannavar, S.; Pujari, V. Inflammation and Cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Ye, Y.; Peng, L.; Vattai, A.; Deuster, E.; Kuhn, C.; Dannecker, C.; Mahner, S.; Jeschke, U.; von Schönfeldt, V.; Heidegger, H.H. Prostaglandin E2 Receptor 3 (EP3) Signaling Promotes Migration of Cervical Cancer via Urokinase-Type Plasminogen Activator Receptor (UPAR). J. Cancer Res. Clin. Oncol. 2020, 146, 2189–2203. [Google Scholar] [CrossRef]
- Haglund, K.; Rusten, T.E.; Stenmark, H. Aberrant Receptor Signaling and Trafficking as Mechanisms in Oncogenesis. Crit. Rev. Oncog. 2007, 13, 39–74. [Google Scholar] [CrossRef]
- Subbaramaiah, K.; Dannenberg, A.J. Cyclooxygenase-2 Transcription Is Regulated by Human Papillomavirus 16 E6 and E7 Oncoproteins: Evidence of a Corepressor/Coactivator Exchange. Cancer Res. 2007, 67, 3976–3985. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, T.; Kim, M.-K.; Suh, D.H.; Chung, H.H.; Song, Y.S. Cyclooxygenase-1 and -2: Molecular Targets for Cervical Neoplasia. J. Cancer Prev. 2013, 18, 123–134. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, G.; Houston, A. Prostaglandin E2 and the EP Receptors in Malignancy: Possible Therapeutic Targets? Br. J. Pharmacol. 2015, 172, 5239–5250. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Pal, I.; Parekh, A.; Thakur, B.; Bharti, R.; Das, S.; Mandal, M. GW627368X Inhibits Proliferation and Induces Apoptosis in Cervical Cancer by Interfering with EP4/EGFR Interactive Signaling. Cell Death Dis. 2016, 7, e2154-13. [Google Scholar] [CrossRef] [PubMed]
- Schmoeckel, E.; Fraungruber, P.; Kuhn, C.; Jeschke, U.; Mahner, S.; Kolben, T.M.; Kolben, T.; Vilsmaier, T.; Hester, A.; Heidegger, H.H. The Role of EP-2 Receptor Expression in Cervical Intraepithelial Neoplasia. Histochem. Cell Biol. 2020, 154, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Sales, K.J.; Katz, A.A.; Davis, M.; Hinz, S.; Soeters, R.P.; Hofmeyr, M.D.; Millar, R.P.; Jabbour, H.N. Cyclooxygenase-2 Expression and Prostaglandin E2synthesis Are up-Regulated in Carcinomas of the Cervix: A Possible Autocrine/Paracrine Regulation of Neoplastic Cell Function via EP2/EP4 Receptors. J. Clin. Endocrinol. Metab. 2001, 86, 2243–2249. [Google Scholar] [CrossRef] [PubMed]
- Sales, K.J.; Katz, A.A.; Millar, R.P.; Jabbour, H.N. Seminal Plasma Activates Cyclooxygenase-2 and Prostaglandin E2 Receptor Expression and Signalling in Cervical Adenocarcinoma Cells. Mol. Hum. Reprod. 2002, 8, 1065–1070. [Google Scholar] [CrossRef]
- Oh, J.M.; Kim, S.H.; Lee, Y.I.; Seo, M.; Kim, S.Y.; Song, Y.S.; Kim, W.H.; Juhnn, Y.S. Human Papillomavirus E5 Protein Induces Expression of the EP4 Subtype of Prostaglandin E2 Receptor in Cyclic AMP Response Element-Dependent Pathways in Cervical Cancer Cells. Carcinogenesis 2009, 30, 141–149. [Google Scholar] [CrossRef]
- Adefuye, A.; Sales, K. Regulation of Inflammatory Pathways in Cancer and Infectious Disease of the Cervix. Scientifica 2012, 2012, 548150. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, X.; Jeschke, U.; von Schönfeldt, V. COX-2-PGE2-EPs in Gynecological Cancers. Arch. Gynecol. Obstet. 2020, 301, 1365–1375. [Google Scholar] [CrossRef]
- Braicu, E.I.; Gasimli, K.; Richter, R.; Nassir, M.; Kümmel, S.; Blohmer, J.U.; Yalcinkaya, I.; Chekerov, R.; Ignat, I.; Ionescu, A.; et al. Role of Serum VEGFA, TIMP2, MMP2 and MMP9 in Monitoring Response to Adjuvant Radiochemotherapy in Patients with Primary Cervical Cancer—Results of a Companion Protocol of the Randomized NOGGO-AGO Phase III Clinical Trial. Anticancer Res. 2014, 34, 385–391. [Google Scholar]
- Caldeira, J.; Laronha, H. Structure and function of human matrix metalloproteinases. Cell 2020, 9, 1076. [Google Scholar]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 147. [Google Scholar]
- Tasleem, F.; Azhar, I.; Ali, S.N.; Perveen, S.; Mahmood, Z.A. Analgesic and Anti-Inflammatory Activities of Piper Nigrum L. Asian Pac. J. Trop. Med. 2014, 7, S461–S468. [Google Scholar] [CrossRef] [PubMed]
- Friel, G.; Liu, C.S.; Kolomeyevskaya, N.V.; Hampras, S.S.; Kruszka, B.; Schmitt, K.; Cannioto, R.A.; Shashikant, B.; Odunsi, K.O.; Moysich, K.B. Aspirin and acetaminophen use and the risk of cervical cancer. J. Low. Genit. Tract Dis. 2015, 19, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Joharatnam-Hogan, N.; Cafferty, F.H.; Macnair, A.; Ring, A.; Langley, R.E. The Role of Aspirin in the Prevention of Ovarian, Endometrial and Cervical Cancers. Women’s Health 2020, 16, 1745506520961710. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Khoi, P.N.; Yoon, H.J.; Lian, S.; Joo, Y.E.; Chay, K.O.; Kim, K.K.; Jung, Y. Do Piperine Inhibits IL-1β-Induced IL-6 Expression by Suppressing P38 MAPK and STAT3 Activation in Gastric Cancer Cells. Mol. Cell. Biochem. 2015, 398, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Chen, X.; Cheng, S.; Shen, Y.; Peng, L.; Xu, H. Piperine Inhibits IL-β Induced Expression of Inflammatory Mediators in Human Osteoarthritis Chondrocyte. Int. Immunopharmacol. 2013, 17, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Jaisin, Y.; Ratanachamnong, P.; Wongsawatkul, O.; Watthammawut, A.; Malaniyom, K.; Natewong, S. Antioxidant and Anti-Inflammatory Effects of Piperine on UV-B-Irradiated Human HaCaT Keratinocyte Cells. Life Sci. 2020, 263, 118607. [Google Scholar] [CrossRef] [PubMed]
- Zadorozhna, M.; Tataranni, T.; Mangieri, D. Piperine: Role in Prevention and Progression of Cancer. Mol. Biol. Rep. 2019, 46, 5617–5629. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010†. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Zhang, W.; Zheng, Q.; Song, M.; Xiao, J.; Cao, Y.; Huang, Q.; Ho, C.-T.; Lu, M. A Review on the Bioavailability, Bio-Efficacies and Novel Delivery Systems for Piperine. Food Funct. 2021, 12, 8867–8881. [Google Scholar] [CrossRef]
- Massagué, J. G1 cell-cycle control and cancer. Nature 2004, 432, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Kastan, M.B.; Bartek, J. Cell-Cycle Checkpoints and Cancer. Nature 2004, 432, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Marino, M.; Martinez-Chantar, M.L.; Nawroth, R.; Sanchez-Garcia, I.; et al. Sustained Proliferation in Cancer: Mechanisms and Novel Therapeutic Targets. Semin. Cancer Biol. 2015, 35, S25–S54. [Google Scholar] [CrossRef]
- Guo, Y.; Pan, W.; Liu, S.; Shen, Z.; Xu, Y.; Hu, L. ERK/MAPK Signalling Pathway and Tumorigenesis (Review). Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling Pathway of MAPK/ERK in Cell Proliferation, Differentiation, Migration, Senescence and Apoptosis. J. Recept. Signal Transduct. 2015, 35, 600–604. [Google Scholar] [CrossRef]
- Apte, R.N.; Dotan, S.; Elkabets, M.; White, M.R.; Reich, E.; Carmi, Y.; Song, X.; Dvozkin, T.; Krelin, Y.; Voronov, E. The Involvement of IL-1 in Tumorigenesis, Tumor Invasiveness, Metastasis and Tumor-Host Interactions. Cancer Metastasis Rev. 2006, 25, 387–408. [Google Scholar] [CrossRef]
- Vendramini-Costa, D.B.; Carvalho, E.J. Molecular Link Mechanisms between Inflammation and Cancer. Curr. Pharm. Des. 2012, 18, 3831–3852. [Google Scholar] [CrossRef]
- Rébé, C.; Ghiringhelli, F. Interleukin-1 β and Cancer. Cancers 2020, 12, 1791. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Li, F.; Shao, M.; Zhang, W.; Zhang, C.; Zhao, X.; Luan, H.; Qi, Y.; Zhang, P.; Liang, L.; et al. IL-8 Is Upregulated in Cervical Cancer Tissues and Is Associated with the Proliferation and Migration of HeLa Cervical Cancer Cells. Oncol. Lett. 2018, 15, 1350–1356. [Google Scholar] [CrossRef]
- Paradkar, P.H.; Joshi, J.V.; Mertia, P.N.; Agashe, S.V.; Vaidya, R.A. Role of Cytokines in Genesis, Progression and Prognosis of Cervical Cancer. Asian Pac. J. Cancer Prev. 2014, 15, 3851–3864. [Google Scholar] [CrossRef]
- Vahedpour, Z.; Abedzadeh-Kalahroudi, M.; Sehat, M.; Piroozmand, A.; Memar, M. Comparison of Cervical Levels of Interleukins-6 and -8 in Patients with and without Cervical Intraepithelial Neoplasia. Asian Pac. J. Cancer Prev. 2021, 22, 1225–1230. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2016, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Elbadawi, M.; Efferth, T. Multiple Cell Death Modalities and Their Key Features (Review). World Acad. Sci. J. 2020, 2, 39–48. [Google Scholar] [CrossRef]
- Nahand, J.S.; Moghoofei, M.; Salmaninejad, A.; Bahmanpour, Z.; Karimzadeh, M.; Nasiri, M.; Mirzaei, H.R.; Pourhanifeh, M.H.; Bokharaei-Salim, F.; Mirzaei, H.; et al. Pathogenic Role of Exosomes and MicroRNAs in HPV-Mediated Inflammation and Cervical Cancer: A Review. Int. J. Cancer 2020, 146, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, P.B.; Power Coombs, M.R.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine, an Alkaloid from Black Pepper, Inhibits Growth of Human Colon Cancer Cells via G1 Arrest and Apoptosis Triggered by Endoplasmic Reticulum Stress. Mol. Carcinog. 2015, 54, 1070–1085. [Google Scholar] [CrossRef]
- Yaffe, P.B.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine Impairs Cell Cycle Progression and Causes Reactive Oxygen Species-Dependent Apoptosis in Rectal Cancer Cells. Exp. Mol. Pathol. 2013, 94, 109–114. [Google Scholar] [CrossRef]
- de Almeida, G.C.; Oliveira, L.F.S.; Predes, D.; Fokoue, H.H.; Kuster, R.M.; Oliveira, F.L.; Mendes, F.A.; Abreu, J.G. Piperine Suppresses the Wnt/β-Catenin Pathway and Has Anti-Cancer Effects on Colorectal Cancer Cells. Sci. Rep. 2020, 10, 11681. [Google Scholar] [CrossRef]
- Do, M.T.; Kim, H.G.; Choi, J.H.; Khanal, T.; Park, B.H.; Tran, T.P.; Jeong, T.C.; Jeong, H.G. Antitumor Efficacy of Piperine in the Treatment of Human HER2-Overexpressing Breast Cancer Cells. Food Chem. 2013, 141, 2591–2599. [Google Scholar] [CrossRef]
- Doucette, C.D.; Hilchie, A.L.; Liwski, R.; Hoskin, D.W. Piperine, a Dietary Phytochemical, Inhibits Angiogenesis. J. Nutr. Biochem. 2013, 24, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Yoo, E.S.; Choo, G.S.; Kim, S.H.; Woo, J.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.O.O.; Kim, S.K.; Park, B.K.; Cho, S.D.; et al. Antitumor and Apoptosis-Inducing Effects of Piperine on Human Melanoma Cells. Anticancer Res. 2019, 39, 1883–1892. [Google Scholar] [CrossRef]
- Fofaria, N.M.; Kim, S.H.; Srivastava, S.K. Piperine Causes G1 Phase Cell Cycle Arrest and Apoptosis in Melanoma Cells through Checkpoint Kinase-1 Activation. PLoS ONE 2014, 9, e94298. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wen, S.; Chen, G.; Wang, S. Antiproliferative Potential of Piperine and Curcumin in Drug-Resistant Human Leukemia Cancer Cells Are Mediated via Autophagy and Apoptosis Induction, S-Phase Cell Cycle Arrest and Inhibition of Cell Invasion and Migration. J. BUON 2020, 25, 401–406. [Google Scholar] [PubMed]
- Siddiqui, S.; Ahamad, M.S.; Jafri, A.; Afzal, M.; Arshad, M. Piperine Triggers Apoptosis of Human Oral Squamous Carcinoma Through Cell Cycle Arrest and Mitochondrial Oxidative Stress. Nutr. Cancer 2017, 69, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, X.; Li, H.; Li, B.; Sun, L.; Xie, T.; Zhu, T.; Zhou, H.; Ye, Z. Piperine Inhibits Proliferation of Human Osteosarcoma Cells via G2/M Phase Arrest and Metastasis by Suppressing MMP-2/-9 Expression. Int. Immunopharmacol. 2015, 24, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Valastyan, S.; Weinberg, R.A. Tumor Metastasis: Molecular Insights and Evolving Paradigms The Invasion-Metastasis Cascade. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef]
- Jing, Y.; Han, Z.; Zhang, S.; Liu, Y.; Wei, L. Epithelial-Mesenchymal Transition in Tumor Microenvironment. Cell Biosci. 2011, 1, 29. [Google Scholar] [CrossRef]
- Zeeshan, R.; Mutahir, Z. Cancer Metastasis—Tricks of the Trade. Bosn. J. Basic Med. Sci. 2017, 17, 172–182. [Google Scholar] [CrossRef]
- LIBRA, M.; SCALIS, A.; VELLA, N.; CLEMENTI, S.; SORIO, R.; STIVALA, F.; SPANDIDOS, D.A.; MAZZARINO, C. Uterine Cervical Carcinoma: Role of Matrix Metalloproteinases (Review). Int. J. Oncol. 2009, 34, 897–903. [Google Scholar] [CrossRef]
- Sidorkiewicz, I.; Piskór, B.; Dąbrowska, E.; Guzińska-Ustymowicz, K.; Pryczynicz, A.; Zbucka-Krętowska, M.; Ławicki, S. Plasma Levels and Tissue Expression of Selected Cytokines, Metalloproteinases and Tissue Inhibitors in Patients with Cervical Cancer. Anticancer Res. 2019, 39, 6403–6412. [Google Scholar] [CrossRef]
- Shiau, M.Y.; Fan, L.C.; Yang, S.C.; Tsao, C.H.; Lee, H.; Cheng, Y.W.; Lai, L.C.; Chang, Y.H. Human Papillomavirus Up-Regulates MMP-2 and MMP-9 Expression and Activity by Inducing Interleukin-8 in Lung Adenocarcinomas. PLoS ONE 2013, 8, e54423. [Google Scholar] [CrossRef]
- Yang, C.; Luo, L.; Bai, X.; Shen, K.; Liu, K.; Wang, J.; Hu, D. Highly-Expressed MicoRNA-21 in Adipose Derived Stem Cell Exosomes Can Enhance the Migration and Proliferation of the HaCaT Cells by Increasing the MMP-9 Expression through the PI3K/AKT Pathway. Arch. Biochem. Biophys. 2020, 681, 108259. [Google Scholar] [CrossRef]
- Greenshields, A.L.; Doucette, C.D.; Sutton, K.M.; Madera, L.; Annan, H.; Yaffe, P.B.; Knickle, A.F.; Dong, Z.; Hoskin, D.W. Piperine Inhibits the Growth and Motility of Triple-Negative Breast Cancer Cells. Cancer Lett. 2015, 357, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Yang, Y. Piperine Depresses the Migration Progression via Downregulating the Akt/MTOR/MMP-9 Signaling Pathway in DU145 Cells. Mol. Med. Rep. 2018, 17, 6363–6370. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Huang, Y.; Porta, R.; Yanagisawa, K.; Gonzalez, A.; Segi, E.; Johnson, D.H.; Narumiya, S.; Carbone, D.P. Host and Direct Antitumor Effects and Profound Reduction in Tumor Metastasis with Selective EP4 Receptor Antagonism. Cancer Res. 2006, 66, 9665–9672. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Qiu, Z.; Huang, J.; Li, Y.; Huang, H.; Xiang, T.; Wan, J.; Hui, T.; Lin, Y.; Li, H.; et al. Cyclooxygenase-2 in Tumor-Associated Macrophages Promotes Metastatic Potential of Breast Cancer Cells through Akt Pathway. Int. J. Biol. Sci. 2016, 12, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Liu, S.; Xiong, J.; Yang, H.; Wu, Y.; Xu, A.; Gong, Y. IL-1β Promotes Cervical Cancer through Activating NF-ΚB/CCL-2. Int. J. Clin. Exp. Pathol. 2021, 14, 426–433. [Google Scholar] [PubMed]
- Futagami, S.; Tatsuguchi, A.; Hiratsuka, T.; Shindo, T.; Horie, A.; Hamamoto, T.; Ueki, N.; Kusunoki, M.; Miyake, K.; Gudis, K.; et al. Monocyte Chemoattractant Protein 1 and CD40 Ligation Have a Synergistic Effect on Vascular Endothelial Growth Factor Production through Cyclooxygenase 2 Upregulation in Gastric Cancer. J. Gastroenterol. 2008, 43, 216–224. [Google Scholar] [CrossRef]
- Gonzalez Maglio, D.H.; Gonzalez Maglio, D.H.G.; Paz, M.L.; Cela, E.M.; Leoni, J. Cyclooxygenase-2 Overexpression in Non-Melanoma Skin Cancer: Molecular Pathways Involved as Targets for Prevention and Treatment. In Skin Cancers-Risk Factors, Prevention and Therapy; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef]
- Huang, Q.; Lan, F.; Wang, X.; Yu, Y.; Ouyang, X.; Zheng, F.; Han, J.; Lin, Y.; Xie, Y.; Xie, F.; et al. IL-1β-Induced Activation of P38 Promotes Metastasis in Gastric Adenocarcinoma via Upregulation of AP-1/c-Fos, MMP2 and MMP9. Mol. Cancer 2014, 13, 18. [Google Scholar] [CrossRef]
- Ponnulakshmi, R. Molecular Docking Data of Piperine with Bax, Caspase 3, Cox 2 and Caspase 9. Bioinformation 2020, 16, 458–461. [Google Scholar] [CrossRef]
- Zazeri, G.; Povinelli, A.P.R.; de Freitas Lima, M.; Cornélio, M.L. The Cytokine Il-1β and Piperine Complex Surveyed by Experimental and Computational Molecular Biophysics. Biomolecules 2020, 10, 1337. [Google Scholar] [CrossRef]
- Lee, S.; Rauch, J.; Kolch, W. Targeting MAPK Signaling in Cancer: Mechanisms of Drug Resistance and Sensitivity. Int. J. Mol. Sci. 2020, 21, 1102. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and Physiological Roles of Inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Vaibhav, K.; Shrivastava, P.; Javed, H.; Khan, A.; Ahmed, M.E.; Tabassum, R.; Khan, M.M.; Khuwaja, G.; Islam, F.; Saeed Siddiqui, M.; et al. Piperine Suppresses Cerebral Ischemia-Reperfusion-Induced Inflammation through the Repression of COX-2, NOS-2, and NF-ΚB in Middle Cerebral Artery Occlusion Rat Model. Mol. Cell. Biochem. 2012, 367, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Tharmalingam, N.; Park, M.; Lee, M.H.; Woo, H.J.; Kim, H.W.; Yang, J.Y.; Rhee, K.J.; Kim, J.B. Piperine Treatment Suppresses Helicobacter Pylori Toxin Entry in to Gastric Epithelium and Minimizes β-Catenin Mediated Oncogenesis and IL-8 Secretion in Vitro. Am. J. Transl. Res. 2016, 8, 885–898. [Google Scholar] [PubMed]
- Hu, D.; Wang, Y.; Chen, Z.; Ma, Z.; You, Q.; Zhang, X.; Liang, Q.; Tan, H.; Xiao, C.; Tang, X.; et al. The Protective Effect of Piperine on Dextran Sulfate Sodium Induced Inflammatory Bowel Disease and Its Relation with Pregnane X Receptor Activation. J. Ethnopharmacol. 2015, 169, 109–123. [Google Scholar] [CrossRef]
- Sieniawska, E.; Świątek, Ł.; Wota, M.; Rajtar, B.; Polz-Dacewicz, M. Microemulsions of Essentials Oils—Increase of Solubility and Antioxidant Activity or Cytotoxicity? Food Chem. Toxicol. 2019, 129, 115–124. [Google Scholar] [CrossRef]
- Hemmat, N.; Baghi, H.B. Human Papillomavirus E5 Protein, the Undercover Culprit of Tumorigenesis. Infect. Agents Cancer 2018, 13, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24 h | 48 h | 72 h | |
|---|---|---|---|
| HeLa | 208.0 | 151.0 | 69.9 |
| SiHa | 182.3 | 190.7 | 187.7 |
| CaSki | 167.8 | 83.3 | 104.2 |
| HaCaT | 218.4 | 162.5 | 181.7 |
| Oligonucleotides | Sequency |
|---|---|
| HPV16-E6 anti-sense | 5′ CTACGTGTTCTTGATGATCTG 3′ |
| HPV16-E6 sense | 5′ CTTACCACAGTTATGCACAGAG 3′ |
| HPV16-E7 anti-sense | 5′ TGCCCATTAACAGGTCTTCC 3′ |
| HPV16-E7 sense | 5′ ACAAGCAGAACCGGACAGAG 3′ |
| PTGS2 anti-sense | 5′ AGAAGGCTTCCCAGCTTTTG 3′ |
| PTGS2 sense | 5′ ATTCCCTTCCTTCGAAATGC 3′ |
| PTGER2 anti-sense | 5′ AGGTCCCATTTTTCCTTTCG 3′ |
| PTGER2 sense | 5′ CCACCTCATTCTCCTGGCTA 3′ |
| PTGER3 anti-sense | 5′ TCTCCGTGTGTGTCTTGCAG 3′ |
| PTGER3 sense | 5′ AGCTTATGGGGATCATGTGC 3′ |
| PTGER4 anti-sense | 5′ CCAAACTTGGCTGATATAACTGG 3′ |
| PTGER4 sense | 5′ CGAGATCCAGATGGTCATCTTAC 3′ |
| MMP2 anti-sense | 5′ CCGTCAAAGGGGTATCCATC 3′ |
| MMP2 sense | 5′ AAGTCTGGAGCGATGTGACC 3′ |
| MMP9 anti-sense | 5′ ATTTCGACTCTCCACGCATC 3′ |
| MMP9 sense | 5′ TTGTGCTCTTCCCTGGAGAC 3′ |
| TIMP1 anti-sense | 5′ TTTTCAGAGCCTTGGAGGAG 3′ |
| TIMP1 sense | 5′ ACTGTTGGCTGTGAGGAATG 3′ |
| TIMP2 anti-sense | 5′ CTATATCCTTCTCAGGCCCTTTG 3′ |
| TIMP2 sense | 5′ AGAAGGAAGTGGACTCTGGAAAC 3 |
| GAPDH anti-sense | 5′-ACCCACTCCTCCACCTTTGA-3 |
| GAPDH sente | 5′-CTGTTGCTGTAGCCAAATTCGT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, L.P.; de Sousa, S.O.; Gusson-Zanetoni, J.P.; de Melo Moreira Silva, L.L.; Frigieri, B.M.; Henrique, T.; Tajara, E.H.; Oliani, S.M.; Rodrigues-Lisoni, F.C. Piperine Reduces Neoplastic Progression in Cervical Cancer Cells by Downregulating the Cyclooxygenase 2 Pathway. Pharmaceuticals 2023, 16, 103. https://doi.org/10.3390/ph16010103
Cardoso LP, de Sousa SO, Gusson-Zanetoni JP, de Melo Moreira Silva LL, Frigieri BM, Henrique T, Tajara EH, Oliani SM, Rodrigues-Lisoni FC. Piperine Reduces Neoplastic Progression in Cervical Cancer Cells by Downregulating the Cyclooxygenase 2 Pathway. Pharmaceuticals. 2023; 16(1):103. https://doi.org/10.3390/ph16010103
Chicago/Turabian StyleCardoso, Luana Pereira, Stefanie Oliveira de Sousa, Juliana Prado Gusson-Zanetoni, Laura Luciana de Melo Moreira Silva, Barbara Maria Frigieri, Tiago Henrique, Eloiza Helena Tajara, Sonia Maria Oliani, and Flávia Cristina Rodrigues-Lisoni. 2023. "Piperine Reduces Neoplastic Progression in Cervical Cancer Cells by Downregulating the Cyclooxygenase 2 Pathway" Pharmaceuticals 16, no. 1: 103. https://doi.org/10.3390/ph16010103
APA StyleCardoso, L. P., de Sousa, S. O., Gusson-Zanetoni, J. P., de Melo Moreira Silva, L. L., Frigieri, B. M., Henrique, T., Tajara, E. H., Oliani, S. M., & Rodrigues-Lisoni, F. C. (2023). Piperine Reduces Neoplastic Progression in Cervical Cancer Cells by Downregulating the Cyclooxygenase 2 Pathway. Pharmaceuticals, 16(1), 103. https://doi.org/10.3390/ph16010103