
Chelation of Zinc with Biogenic Amino Acids: Description of Properties Using Balaban Index, Assessment of Biological Activity on Spirostomum Ambiguum Cellular Biosensor, Influence on Biofilms and Direct Antibacterial Action

 , , ,
, , , 
Abstract
:1. Introduction
2. Results and Discussion
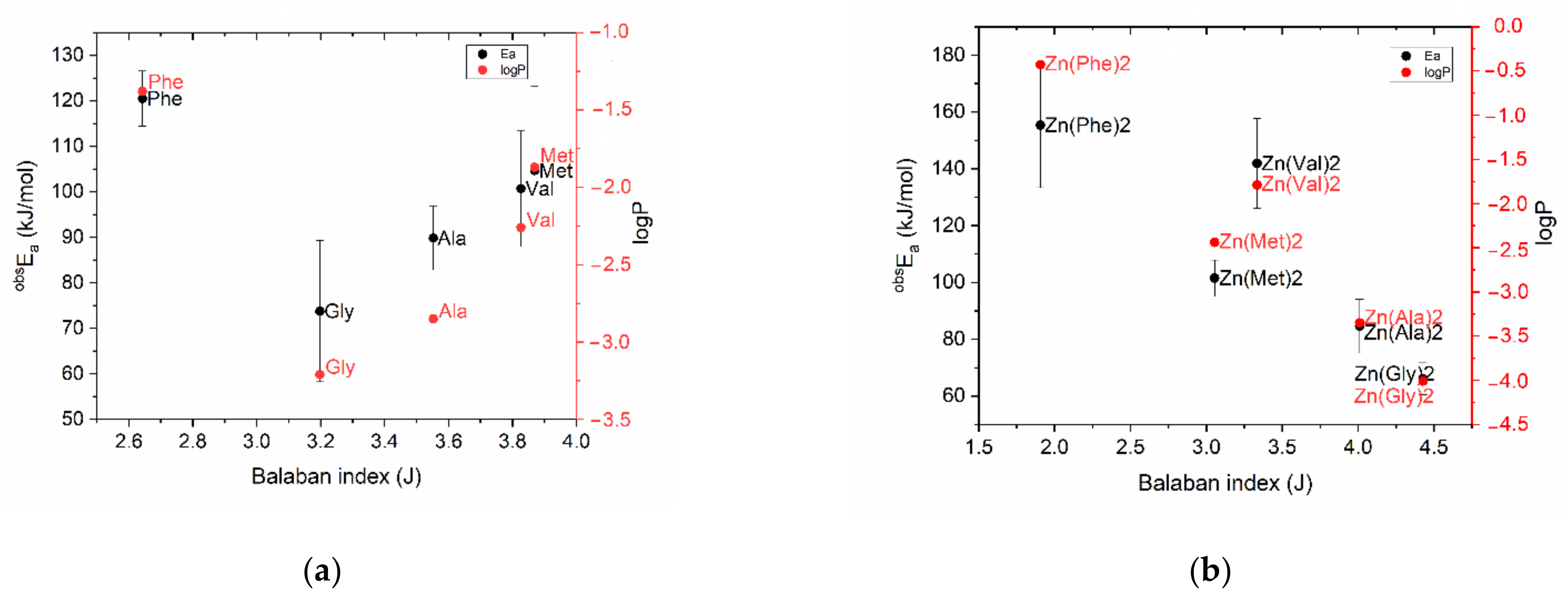
2.1. Prediction of Zn(AA)2 Properties Using Topological Descriptors
2.2. Experimental Assessment of Zn(AA)2 Biological Activity Using S. Ambiguum Cellular Biosensor
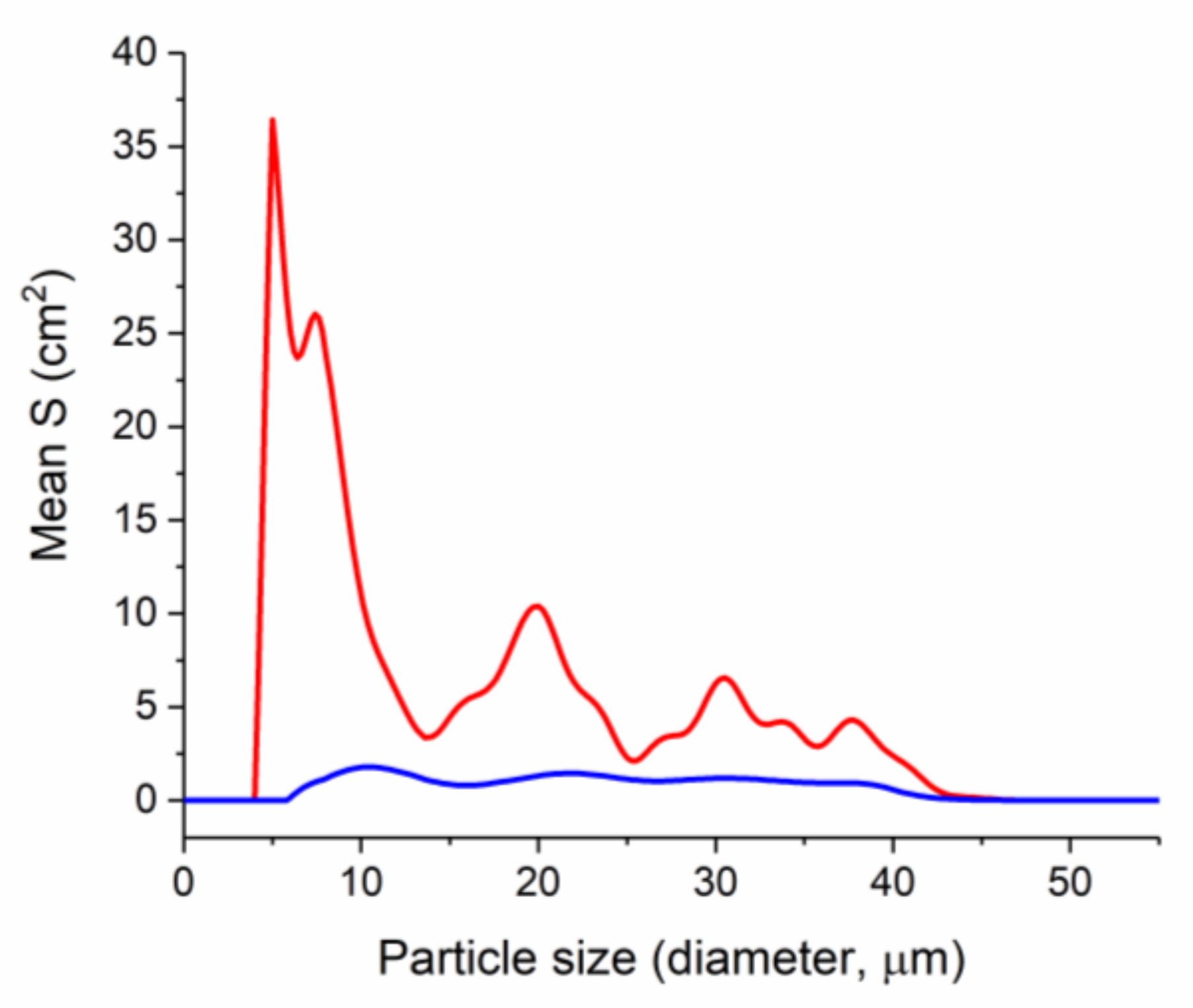
2.3. Microbiological Screening: Comparative Study of the Effect of Zinc Amino Acid Complexes on Biofilm Formation in E. coli Culture
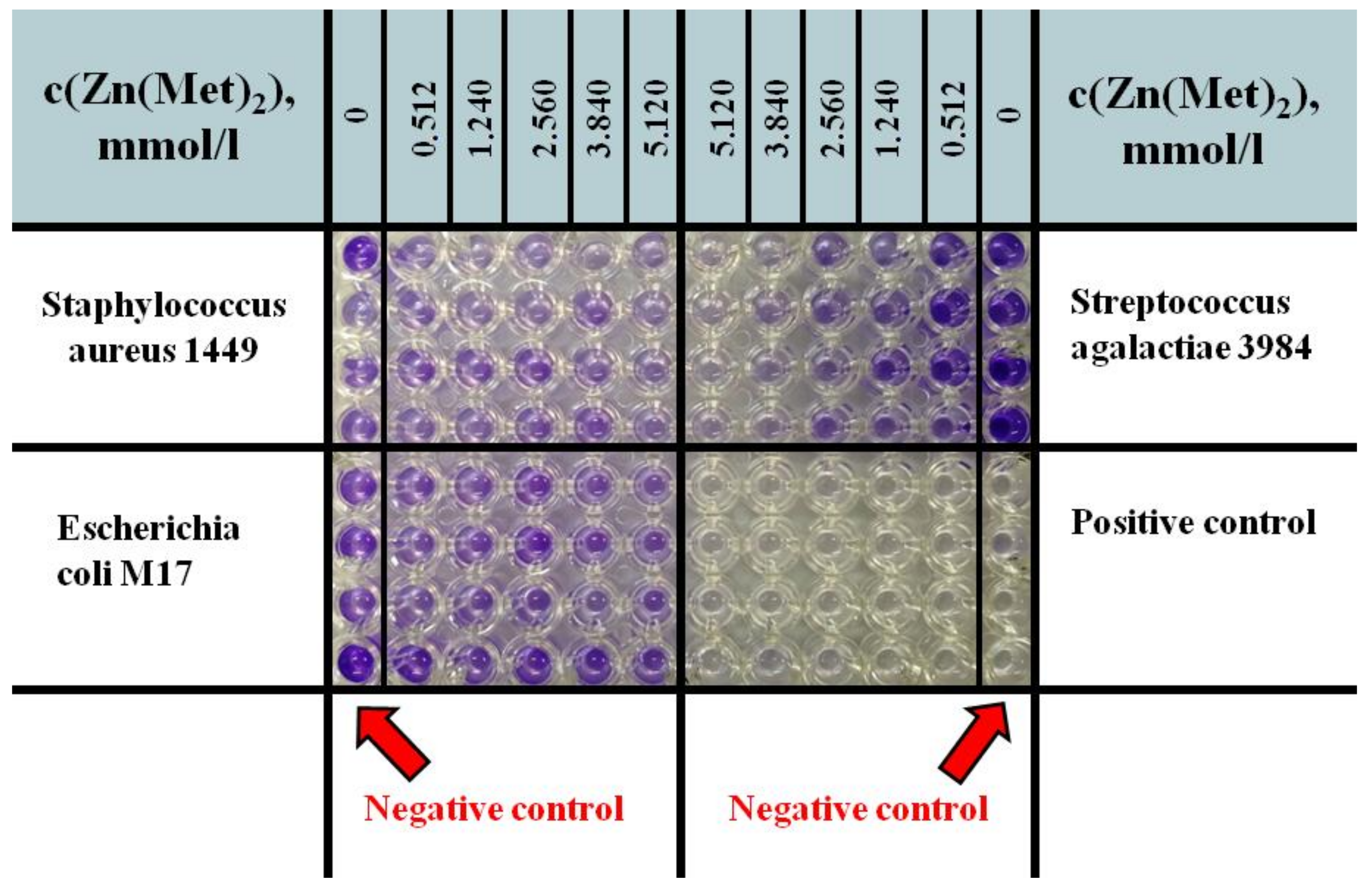
2.4. Assessment of Antibacterial Activity of Zinc Methioninate Complex
Determination of Minimum Inhibitory Concentrations (MIC) for Zinc Methioninate Complex
2.5. Zn(Met)2 Impact on Biofilm Formation of Pathogenic and Conditionally Pathogenic Strains
3. Materials and Methods
3.1. Chemicals and Media
3.2. Construction of Molecular Graphs and Balaban Index Calculation
- -
- The values of the diagonal elements Dii of the topological distance matrix:where Zi—the number of all electrons in atom iDii = 1 − 6/ZiDii (Zn-Zn) = 1 − 6/30 = 0.8.
- -
- The values of the parameter Pij for the bonds of various atoms used in the calculation of non-diagonal elements of the topological distance matrix:where Zi and Zj—the number of all electrons in atoms i and j, respectively, and br—the multiplicity of the bond between atoms i and j.Pij = 36/brZiZjPij (Zn-O) = 36/1 × 30 × 8 = 0.15Pij (Zn-N) = 36/1 × 30 × 7 = 0.171
3.3. Cellular Biosensor Spirostomum ambiguum for Testing the Biological Activity
3.3.1. Test Solutions
3.3.2. Cell Biosensor Spirostomum ambiguum and Research Technique
3.4. Comparative Study of the Zinc Amino Acid Complexes Effect on Biofilm Formation in E. coli Culture
3.4.1. Bacterial Strain and Test Solutions
3.4.2. Particle Size Analysis with Laser Diffraction Method
3.5. Determination of Antimicrobial Activity of Zinc Methioninate Complex
3.5.1. Bacterial Strains
3.5.2. Test Solution of Zn(Met)2
3.5.3. Inoculum Preparation
3.5.4. Assessment of Antimicrobial Activity Using Well Diffusion Method
3.5.5. Determination of Minimum Inhibitory Concentration (MIC)
3.6. Zinc Methioninate Complex Impact on Biofilm Formation
3.7. Data Processing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moriwa, Y.; Suzuki, N.; Shoji, A.; Yanagida, A. Analysis of complexation interactions between metal ions and drugs under pseudo-physiological pH conditions by a high-throughput screening method using a solid-phase extraction cartridge. Anal. Sci. 2020, 36, 709–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, L.; Csintalan, G.; Kálmán, E.; Sipos, P.; Szvetnik, A. Fémionok és fémkomplexek alkalmazása az orvostudományban I [Applications of metal ions and their complexes in medicine I]. Acta Pharm. Hung. 2003, 73, 221–236. [Google Scholar] [PubMed]
- Jurca, T.; Marian, E.; Vicaş, L.G.; Mureşan, M.; Fritea, L. Metal Complexes of Pharmaceutical Substances. In Spectroscopic Analyses—Developments and Applications; Sharmin, E., Zafar, F., Eds.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, K.; Sen, K. Some drugs in action: Metal ions do influence the activity! IJPSR 2015, 6, 1–13. [Google Scholar]
- Pattan, S.R.; Pawar, S.B.; Vetal, S.S.; Gharate, U.D.; Bhawar, S.B. The scope of metal complexes in drug design—A review. Indian Drugs 2012, 49, 5–12. [Google Scholar] [CrossRef]
- Boros, E.; Dyson, P.J.; Gasser, G. Classification of metal-based drugs according to their mechanisms of action. Chem 2020, 6, 41–60. [Google Scholar] [CrossRef]
- Yoshikawa, Y.; Ueda, E.; Suzuki, Y.; Yanagihara, N.; Sakurai, H.; Kojima, Y. New insulinomimetic zinc(II) complexes of alpha-amino acids and their derivatives with Zn(N2O2) coordination mode. Chem. Pharm. Bull. 2001, 49, 652–654. [Google Scholar] [CrossRef] [Green Version]
- Sukul, A.; Haque, S.; Poddar, K.S.; Hossain, M.S.; Niloy, K.K.; Saha, S.K. Comparative physicochemical, anti-inflammatory, and analgesic activity assay of synthesized chromium and nickel complexes of indomethacin. Cogent Chem. 2017, 3, 1302312. [Google Scholar] [CrossRef]
- Kartina, D.; Wahab, A.W.; Ahmad, A.; Irfandi, R.; Raya, I. In vitro antibacterial and anticancer activity of Zn(II)Valinedithiocarbamate complexes. J. Phys. Conf. Ser. 2019, 1341, 032042. [Google Scholar] [CrossRef]
- Borthagaray, G.; Mondelli, M.; Torre, M.H. Essential transition metal ion complexation as a strategy to improve the antimicrobial activity of organic drugs. J. Infect Dis. Epidemiol. 2016, 2, 2–14. [Google Scholar] [CrossRef]
- Nunes, W.D.G.; do Nascimento, A.L.C.S.; Moura, A.; Gaglieri, C.; Vallim, G.B.; Nascimento, L.C.; Mendes, R.A.; Ionashiro, M.; Caires, F.J. Thermal, spectroscopic and antimicrobial activity characterization of some norfloxacin complexes. J. Therm. Anal. Calorim. 2018, 132, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Kumar, Y.; Puri, P.; Sharma, C.; Aneja, K.R. Metal-based biologically active compounds: Synthesis, spectral, and antimicrobial studies of cobalt, nickel, copper, and zinc complexes of triazole-derived schiff bases. Bioinorg. Chem. Appl. 2011, 2011, 901716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X.; Li, J.; Khan, M.; Hossain, Z.; Ma, F.; Liu, X. A novel zinc complex with antibacterial and antioxidant activity. BMC Chem. 2021, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Rahmouni, N.T.; Bensiradj, N.H.; Megatli, S.A.; Djebbar, S.; Baitich, O.B. New mixed amino acids complexes of iron(III) and zinc(II) with isonitrosoacetophenone: Synthesis, spectral characterization, DFT study and anticancer activity. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 213, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.A.; Omar, S.N.; Darawsheh, M.D.; Fares, H. Synthesis, characterization and antimicrobial activity of zinc(II) ibuprofen complexes with nitrogen-based ligands. J. Coord. Chem. 2016, 69, 1110–1122. [Google Scholar] [CrossRef]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef]
- Sauer, A.K.; Pfaender, S.; Hagmeyer, S.; Tarana, L.; Mattes, A.K.; Briel, F.; Küry, S.; Boeckers, T.M.; Grabrucker, A.M. Characterization of zinc amino acid complexes for zinc delivery in vitro using Caco-2 cells and enterocytes from hiPSC. Biometals 2017, 30, 643–661. [Google Scholar] [CrossRef] [Green Version]
- Chien, X.X.; Zafra-Stone, S.; Bagchi, M.; Bagchi, D. Bioavailability, antioxidant and immune-enhancing properties of zinc methionine. Biofactors 2006, 27, 231–244. [Google Scholar] [CrossRef]
- Sapota, A.; Daragó, A.; Skrzypińska-Gawrysiak, M.; Nasiadek, M.; Klimczak, M.; Kilanowicz, A. The bioavailability of different zinc compounds used as human dietary supplements in rat prostate: A comparative study. Biometals 2014, 27, 495–505. [Google Scholar] [CrossRef]
- Abendrot, M.; Chęcińska, L.; Kusz, J.; Lisowska, K.; Zawadzka, K.; Felczak, A.; Kalinowska-Lis, U. Zinc(II) complexes with amino acids for potential use in dermatology: Synthesis, crystal structures, and antibacterial activity. Molecules 2020, 25, 951. [Google Scholar] [CrossRef] [Green Version]
- Krężel, A.; Maret, W. The biological inorganic chemistry of zinc ions. Arch. Biochem. Biophys. 2016, 611, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Sakr, S.H.; Elshafie, H.S.; Camele, I.; Sadeek, S.A. Synthesis, spectroscopic, and biological studies of mixed ligand complexes of gemifloxacin and glycine with Zn(II), Sn(II), and Ce(III). Molecules 2018, 23, 1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.N.; Hansch, C.; Ames, M.M. Selection of a reference partitioning system for drug design work. J. Pharm. Sci. 1975, 64, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, O.; Elmes, R.; O’Sullivan, F.; Farragher, J.; Robinson, S.; Walker, G. Investigating structural property relationships to enable repurposing of pharmaceuticals as zinc ionophores. Pharmaceutics 2021, 13, 2032. [Google Scholar] [CrossRef] [PubMed]
- Ting, Y.; Cui, Z.; Shicheng, C.; Lei, G.; Hang, L.; Ruowei, F.; Qingfeng, Z.; Jinsong, X.; Zhuang, C.; Zongyong, J. Dietary high zinc oxide modulates the microbiome of ileum and colon in weaned piglets. Front. Microbiol. 2017, 8, 825. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.S. The antibacterial activity of dimethyl sulfoxide (DMSO) with and without of some ligand complexes of the transitional metal ions of ethyl coumarin against bacteria isolate from burn and wound infection. J. Nar. Sci. Res. 2014, 4, 106–109. [Google Scholar]
- Qin, Z.; Balasubramanian, S.K.; Wolkers, W.F.; Pearce, J.A.; Bischof, J.C. Correlated parameter fit of Arrhenius model for thermal denaturation of proteins and cells. Ann. Biomed. Eng. 2014, 42, 2392–2404. [Google Scholar] [CrossRef] [Green Version]
- Poddar, R.; Jain, A.; Kidwai, M. Bis[(l)prolinate-N,O]Zn: A water-soluble and recycle catalyst for various organic transformations. J. Adv. Res. 2017, 8, 245–270. [Google Scholar] [CrossRef]
- Wedekind, K.J.; Hortin, A.E.; Baker, D.H. Methodology for assessing zinc bioavailability: Efficacy estimates for zinc-methionine, zinc sulfate, and zinc oxide. J. Anim. Sci. 1992, 70, 178–187. [Google Scholar] [CrossRef]
- Chang, Y.; Tang, H.; Zhang, Z.; Yang, T.; Wu, B.; Zhao, H.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; et al. Zinc methionine improves the growth performance of meat ducks by enhancing the antioxidant capacity and intestinal barrier function. Front. Vet. Sci. 2022, 9, 774160. [Google Scholar] [CrossRef]
- Mofokeng, T.P.; Moloto, M.J.; Shumbula, P.M.; Nyamukamba, P.; Mubiayi, P.K.; Takaidza, S.; Marais, L. Antimicrobial activity of amino acid-capped zinc and copper sulphide nanoparticles. J. Nanotechnol. 2018, 2018, 4902675. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.S.; Khurshid, S.J.; Iqbal, M.Z. Antibacterial activity of copper-amino acid complexes. J. Pak. Med. Assoc. 1990, 40, 221–222. [Google Scholar] [PubMed]
- Zhou, B.; Trinajstić, N. Bounds on the Balaban index. Croat. Chem. Acta 2008, 81, 319–323. [Google Scholar]
- Raos, N.; Milicević, A. Estimation of stability constants of coordination compounds using models based on topological indices. Arh. Hig. Rada. Toksikol. 2009, 60, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Balaban, A.T.; Khadikar, P.V.; Supuran, C.T.; Thakur, A.; Thakur, M. Study on supramolecular complexing ability vis-à-vis estimation of pKa of substituted sulfonamides: Dominating role of Balaban index (J). Bioorganic Med. Chem. Lett. 2005, 15, 3966–3973. [Google Scholar] [CrossRef]
- Kujawski, J.; Popielarska, H.; Myka, A.; Drabińska, B.; Bernard, M. The log P parameter as a molecular descriptor in the computer-aided drug design—An overview. CMST 2012, 18, 81–88. [Google Scholar] [CrossRef]
- Remko, M.; Boháč, A.; Kováčiková, L. Molecular structure, pKa, lipophilicity, solubility, absorption, polar surface area, and blood brain barrier penetration of some antiangiogenic agents. Struct. Chem. 2011, 22, 635–648. [Google Scholar] [CrossRef]
- Hansch, C.; Kurup, A.; Garg, R.; Gao, H. Chem-bioinformatics and QSAR: A review of QSAR lacking positive hydrophobic terms. Chem. Rev. 2001, 101, 619–672. [Google Scholar] [CrossRef]
- Jhanwar, B.; Sharma, V.; Singla, R.K.; Shrivastava, B. QSAR—Hansch analysis and related approaches in drug design. Pharmacologyonline 2011, 1, 306–344. [Google Scholar]
- Uspenskaya, E.V.; Pleteneva, T.V.; Hanh, P.M.; Kazimova, I.V. Assesment of biology activity of the peeling substances by the physicochemical approaches on the Spirostomum ambiguum cell model. Int. J. Pharm. Pharm. Sci. 2021, 13, 82–86. [Google Scholar] [CrossRef]
- Gudasi, K.B.; Patil, M.S.; Vadavi, R.S. Synthesis, characterization of copper(II), cobalt(II), nickel(II), zinc(II) and cadmium(II) complexes of [7-hydroxy-4-methyl-8-coumarinyl]glycine and a comparitive study of their microbial activities. Eur. J. Med. Chem. 2008, 43, 2436–2441. [Google Scholar] [CrossRef]
- Rao, N.S.; Reddy, M.G. Studies on the synthesis, characterisation and antimicrobial activity of new Co(II), Ni(II) and Zn(II) complexes of Schiff base derived from ninhydrin and glycine. Biol. Metals 1990, 3, 19–23. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Sakr, S.H.; Sadeek, S.A.; Camele, I. Biological investigations and spectroscopic studies of new moxifloxacin/glycine—metal vomplexes. Chem. Biodivers. 2019, 16, e1800633. [Google Scholar] [CrossRef] [PubMed]
- Syroyeshkin, A.V.; Buyanova, N.I.; Sokolova, S.L.; Uspenskaya, E.V.; Shchipkov, V.P. Role of plasmids in the formation of E. Coli polycellular forms. Bull. Exp. Biol. Med. 2011, 150, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Katongole, P.; Nalubega, F.; Florence, N.C.; Asiimwe, B.; Andia, I. Biofilm formation, antimicrobial susceptibility and virulence genes of Uropathogenic Escherichia coli isolated from clinical isolates in Uganda. BMC Infect. Dis. 2020, 20, 453. [Google Scholar] [CrossRef] [PubMed]
- Stoitsova, S.; Vacheva, A.; Paunova-Krasteva, T.; Ivanova, R.; Danova, S.; Manasiev, J. The multicellular behavior of Escherichia coli: A target intferences. In Proceedings of the New Trends in Microbiology 65th Anniversary of the Stephan Angeloff; Institute of Microbilogy, Bulgarian Academy of Sciences: Sofia, Bulgary, 2012; pp. 147–156. [Google Scholar]
- Syroeshkin, A.V.; Popov, P.I.; Grebennikova, T.V.; Frolov, V.A.; Pleteneva, T.V. Laser diffraction for standardization of heterogeneous pharmaceutical preparations. J. Pharm. Biomed. Anal. 2005, 37, 927–930. [Google Scholar] [CrossRef]
- Syroeshkin, A.V.; Buyanova, N.I.; Sokolova, S.L.; Shchipkov, V.P. The influence of Escherichia coli specific surface cell structures on the process of polycellular forms creation in liquid medium. RUDN J. Med. 2009, 4, 154–157. [Google Scholar]
- Abendrot, M.; Płuciennik, E.; Felczak, A.; Zawadzka, K.; Piatczak, E.; Nowaczyk, P.; Kalinowska-Lis, U. Zinc(II) complexes of amino acids as new active ingredients for anti-acne dermatological preparations. Int. J. Mol. Sci. 2021, 22, 1641. [Google Scholar] [CrossRef]
- Harrison, J.J.; Turner, R.J.; Marques, L.L.R.; Ceri, H. Biofilms: A new understanding of these microbial communities is driving a revolution that may transform the science of microbiology. Am. Sci. 2005, 93, 508–515. [Google Scholar] [CrossRef]
- Lopez, D.; Vlamakis, H.; Kolter, R. Biofilms. Cold Spring Harb. Perspect. Biol. 2010, 2, a000398. [Google Scholar] [CrossRef]
- Rolfe, R.D. The role of probiotic cultures in the control of gastrointestinal health. J. Nutr. 2000, 130, 396S–402S. [Google Scholar] [CrossRef]
- Kruis, W.; Schütz, E.; Fric, P.; Fixa, B.; Judmaier, G.; Stolte, M. Double-blind comparison of an oral Escherichia coli preparation and mesalazine in maintaining remission of ulcerative colitis. Aliment. Pharmacol. Ther. 1997, 11, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, L.R.; Small, J.; Hoerr, R.A.; Bostwick, E.F.; Maines, L.; Koltun, W.A. In vitro and in vivo effects of the probiotic Escherichia coli strain M-17: Immunomodulation and attenuation of murine colitis. Br. J. Nutr. 2008, 100, 530–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popov, V.I. ChemicPen. Certificate of official registration of a software No. 2005612073. Rospatent. 2005. [Google Scholar]
- Popov, V.I. ChemicDescript. Certificate of official registration of a software No. 2003612305. Rospatent. 2003. [Google Scholar]
- Cetramax ChemicPen Cetramax ChemicPen 2.6. Available online: https://cetramax-chemicpen.software.informer.com (accessed on 1 July 2022).
- Hadda, T.B.; Rastija, V.; AlMalki, F.; Titi, A.; Touzani, R.; Mabkhot, Y.N.; Khalid, S.; Zarrouk, A.; Siddiqui, B.S. Petra/Osiris/Molinspiration and molecular docking analyses of 3-hydroxy-indolin-2-one derivatives as potential antiviral agents. Curr. Comput.-Aided. Drug Des. 2021, 17, 123–133. [Google Scholar] [CrossRef]
- Tetko, I.V.; Gasteiger, J.; Todeschini, R.; Mauri, A.; Livingstone, D.; Ertl, P.; Palyulin, V.A.; Radchenko, E.V.; Zefirov, N.S.; Makarenko, A.S.; et al. Virtual computational chemistry laboratory—Design and description. J. Comput. Aid. Mol. Des. 2005, 19, 453–463. [Google Scholar] [CrossRef]
- Hallman, P.S.; Perrin, D.D.; Watt, A.E. The computed distribution of copper (II) and zinc (II) ions among seventeen amino acids present in human blood plasma. Biochem. J. 1971, 121, 549–555. [Google Scholar] [CrossRef]
- Goncharuk, V.V.; Syroeshkin, A.V.; Zlatskiy, I.A.; Uspenskaya, E.V.; Orekhova, A.V.; Levitskaya, O.V.; Dobrovolskiy, V.I.; Pleteneva, T.V. Quasichemical description of the cell death kinetics of cellular biosensor Spirostomum ambiguum for testing the biological activity of aqueous solutions. J. Water Chem. Technol. 2017, 39, 178–187. [Google Scholar] [CrossRef]
- Levitskaya, O.V.; Syroeshkin, A.V.; Pleteneva, T.V. Arrhenius kinetics as a bioactivity assessment criterion for drug substances and excipients. Pharm. Chem. J. 2016, 49, 779–781. [Google Scholar] [CrossRef]
- Ashby, M.J.; Neale, J.E.; Knott, S.J.; Critchley, I.A. Effect of antibiotics on non-growing planktonic cells and biofilms of Escherichia coli. J. Antimicrob. Chemother. 1994, 33, 443–452. [Google Scholar] [CrossRef]
- Berezinskaya, T.L. The Study of the Subpopulation Structure of Bacterial Cultures and Ways of Controlling Interpopulation Cell Transitions: Dissertation of the Candidate of Biological Sciences: 03.00.07; Russian Federation, RUDN University: Moscow, Russia, 2005; 187p. [Google Scholar]
- Buyanova, N.I.; Sokolova, S.L.; Uspenskaya, E.V.; Shchipkov, V.P.; Syroyeshkin, A.V. Policelluljuljarnye formy Escherichia coli. [Polycellular forms of Escherichia coli]. Zdorov’e i obrazovanie v XXI veke 2008, 10, 647–648. [Google Scholar]
- Mbarga, M.J.A.; Podoprigora, I.; Davares, A.K.L.; Esther, N.; Senyagin, A. Urinary tract infections: Virulence factors, resistance to antibiotics, and management of uropathogenic bacteria with medicinal plants—A review. J. Appl. Pharm. Sci. 2021, 11, 1–12. [Google Scholar]
- Mbarga, M.J.A.; Podoprigora, I.V.; Volina, E.G.; Davares, A.K.L.; Das, M.S.; Sharova, I.N. Prolonged exposure to antimicrobials induces changes in susceptibility to antibiotics, biofilm formation and pathogenicity in Staphylococcus aureus. J. Pharm. Res. Int. 2021, 33, 140–151. [Google Scholar]
- The European Pharmacopoeia, 8th ed.; 01/2008:1027; Council of Europe: Strasbourg, France, 2013; Volume 2.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | AA | Zn(AA)2 | ||
|---|---|---|---|---|
| J | logP | J | logP | |
| Gly | 3.1974 | −3.21 | 4.4268 | −4.01 |
| Ala | 3.5523 | −2.85 | 4.0092 | −3.35 |
| Val | 3.8267 | −2.26 | 3.3345 | −1.79 |
| Leu | 3.6362 | −1.52 | 2.9205 | −0.73 |
| Ile | 3.8679 | −1.72 | 3.0964 | −0.79 |
| Met | 3.8694 | −1.87 | 3.0553 | −2.44 |
| Thr | 3.9231 | −2.94 | 3.3977 | −4.45 |
| Lys | 3.3287 | −3.05 | 2.5290 | −4.29 |
| Trp | 2.2250 | −1.06 | 1.4497 | −0.13 |
| Phe | 2.6425 | −1.38 | 1.9064 | −0.43 |
| N Atom i/j | 1 N | 2 C | 3C | 4 O | 5 O | 6 Zn | 7 N | 8 C | 9 C | 10 O | 11 O | ∑Sij |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 N | 0.143 | 0.857 | 1.857 | 2.232 | 0.321 | 0.171 | 0.342 | 1.199 | 1.071 | 1.446 | 0.321 | 9.96 |
| 2 C | 0.857 | 0 | 1 | 1.375 | 1.75 | 1.028 | 1.199 | 2.056 | 1.928 | 2.303 | 1.178 | 14.674 |
| 3C | 1.857 | 1 | 0 | 0.375 | 0.75 | 0.9 | 1.071 | 1.928 | 1.8 | 2.175 | 1.05 | 12.906 |
| 4 O | 2.232 | 1.375 | 0.375 | 0.25 | 1.125 | 1.275 | 1.446 | 2.303 | 2.175 | 2.55 | 1.425 | 16.531 |
| 5 O | 0.321 | 1.75 | 0.75 | 1.125 | 0.25 | 0.15 | 0.321 | 1.178 | 1.05 | 1.425 | 0.3 | 8.62 |
| 6 Zn | 0.171 | 1.028 | 0.9 | 1.275 | 0.15 | 0.8 | 0.171 | 1.028 | 0.9 | 1.275 | 0.15 | 7.848 |
| 7 N | 0.342 | 1.199 | 1.071 | 1.446 | 0.321 | 0.171 | 0.143 | 0.857 | 1.857 | 2.232 | 0.321 | 9.96 |
| 8 C | 1.199 | 2.056 | 1.928 | 2.303 | 1.178 | 1.028 | 0.857 | 0 | 1 | 1.375 | 1.75 | 14.674 |
| 9 C | 1.071 | 1.928 | 1.8 | 2.175 | 1.05 | 0.9 | 1.857 | 1 | 0 | 0.375 | 0.75 | 12.906 |
| 10 O | 1.446 | 2.303 | 2.175 | 2.55 | 1.425 | 1.275 | 2.232 | 1.375 | 0.375 | 0.25 | 1.125 | 16.531 |
| 11 O | 0.321 | 1.178 | 1.05 | 1.425 | 0.3 | 0.15 | 0.321 | 1.75 | 0.75 | 1.125 | 0.25 | 8.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marukhlenko, A.V.; Morozova, M.A.; Mbarga, A.M.J.; Antipova, N.V.; Syroeshkin, A.V.; Podoprigora, I.V.; Maksimova, T.V. Chelation of Zinc with Biogenic Amino Acids: Description of Properties Using Balaban Index, Assessment of Biological Activity on Spirostomum Ambiguum Cellular Biosensor, Influence on Biofilms and Direct Antibacterial Action. Pharmaceuticals 2022, 15, 979. https://doi.org/10.3390/ph15080979
Marukhlenko AV, Morozova MA, Mbarga AMJ, Antipova NV, Syroeshkin AV, Podoprigora IV, Maksimova TV. Chelation of Zinc with Biogenic Amino Acids: Description of Properties Using Balaban Index, Assessment of Biological Activity on Spirostomum Ambiguum Cellular Biosensor, Influence on Biofilms and Direct Antibacterial Action. Pharmaceuticals. 2022; 15(8):979. https://doi.org/10.3390/ph15080979
Chicago/Turabian StyleMarukhlenko, Alla V., Mariya A. Morozova, Arsène M. J. Mbarga, Nadezhda V. Antipova, Anton V. Syroeshkin, Irina V. Podoprigora, and Tatiana V. Maksimova. 2022. "Chelation of Zinc with Biogenic Amino Acids: Description of Properties Using Balaban Index, Assessment of Biological Activity on Spirostomum Ambiguum Cellular Biosensor, Influence on Biofilms and Direct Antibacterial Action" Pharmaceuticals 15, no. 8: 979. https://doi.org/10.3390/ph15080979
APA StyleMarukhlenko, A. V., Morozova, M. A., Mbarga, A. M. J., Antipova, N. V., Syroeshkin, A. V., Podoprigora, I. V., & Maksimova, T. V. (2022). Chelation of Zinc with Biogenic Amino Acids: Description of Properties Using Balaban Index, Assessment of Biological Activity on Spirostomum Ambiguum Cellular Biosensor, Influence on Biofilms and Direct Antibacterial Action. Pharmaceuticals, 15(8), 979. https://doi.org/10.3390/ph15080979