
Design and Synthesis of a Mitochondria-Targeting Radioprotectant for Promoting Skin Wound Healing Combined with Ionizing Radiation Injury


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
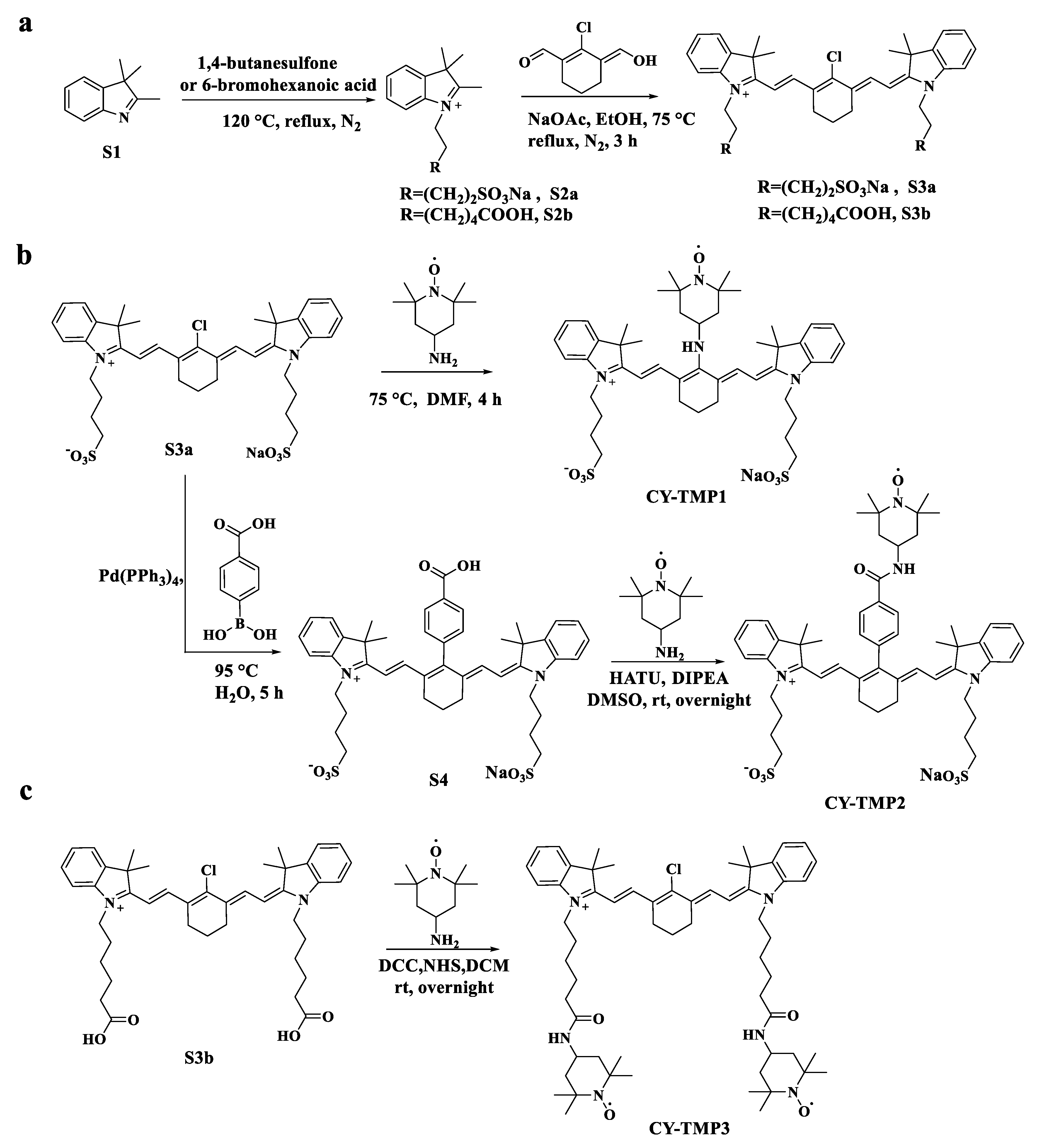
2.1. Synthesis and Characterization of TEMPO-Conjugated Heptamethine Cyanines
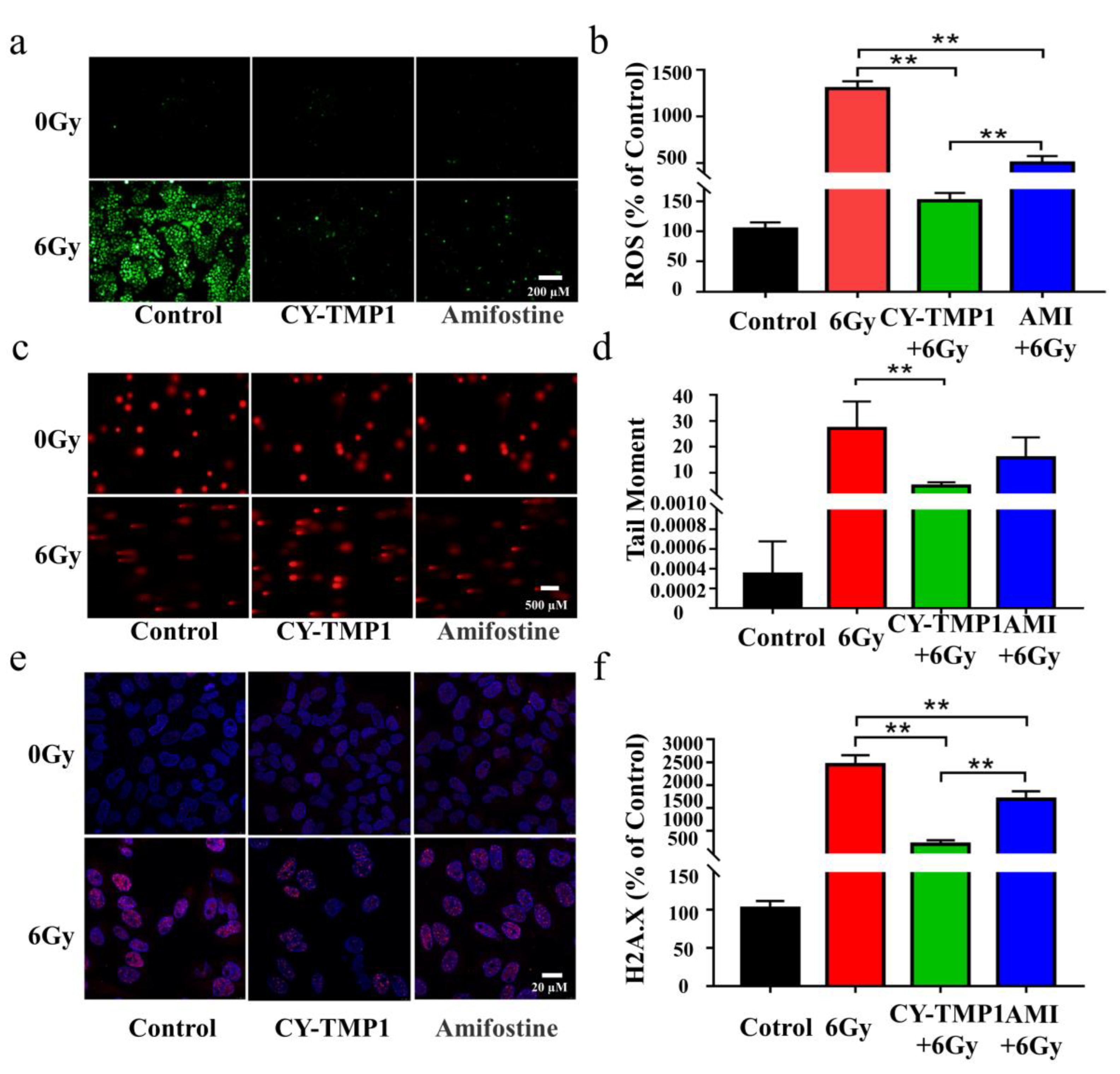
2.2. Evaluation of Radioprotective Effect In Vitro
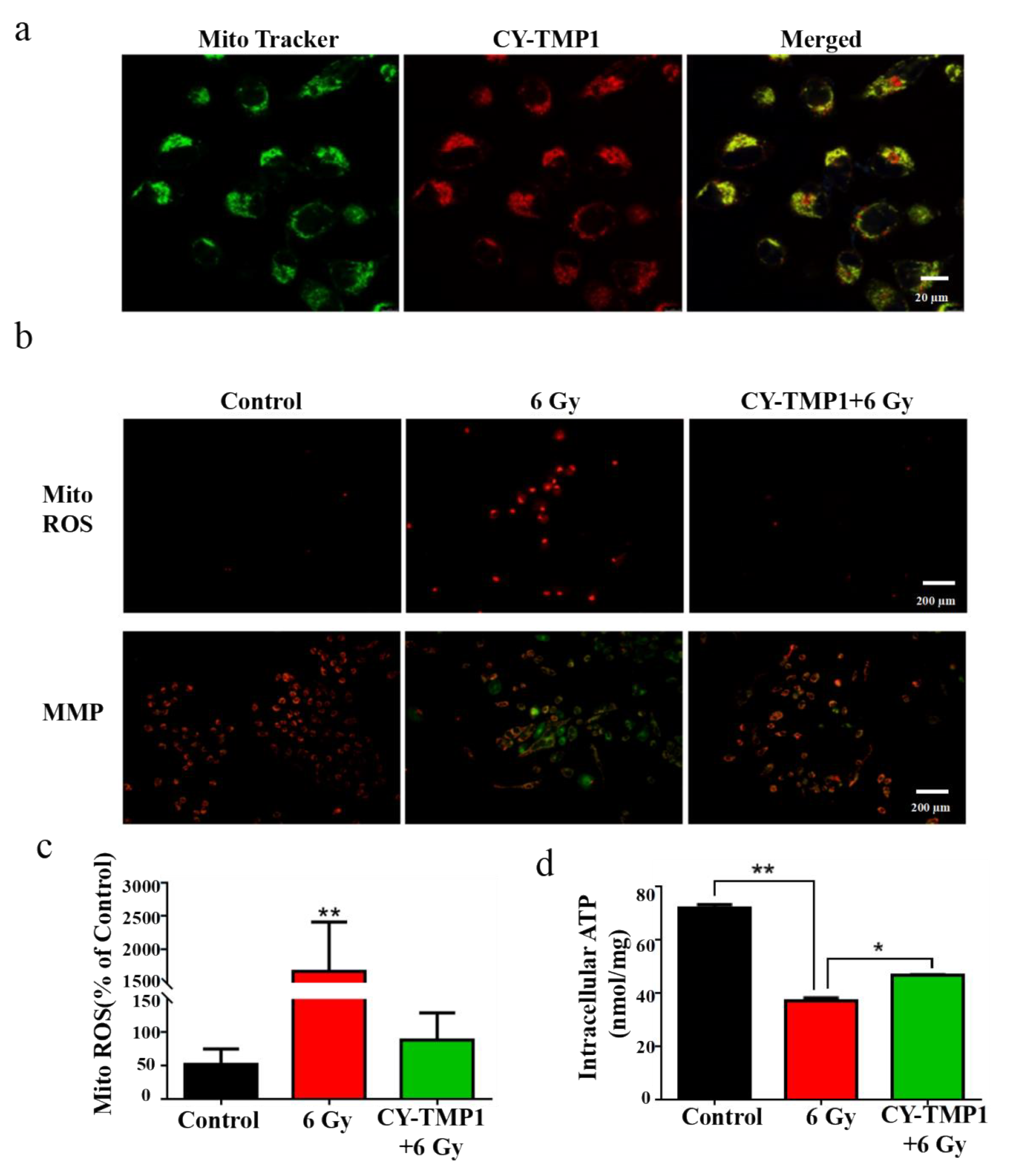
2.3. Mitochondria-Targeting ROS Scavenging and Mechanism Study
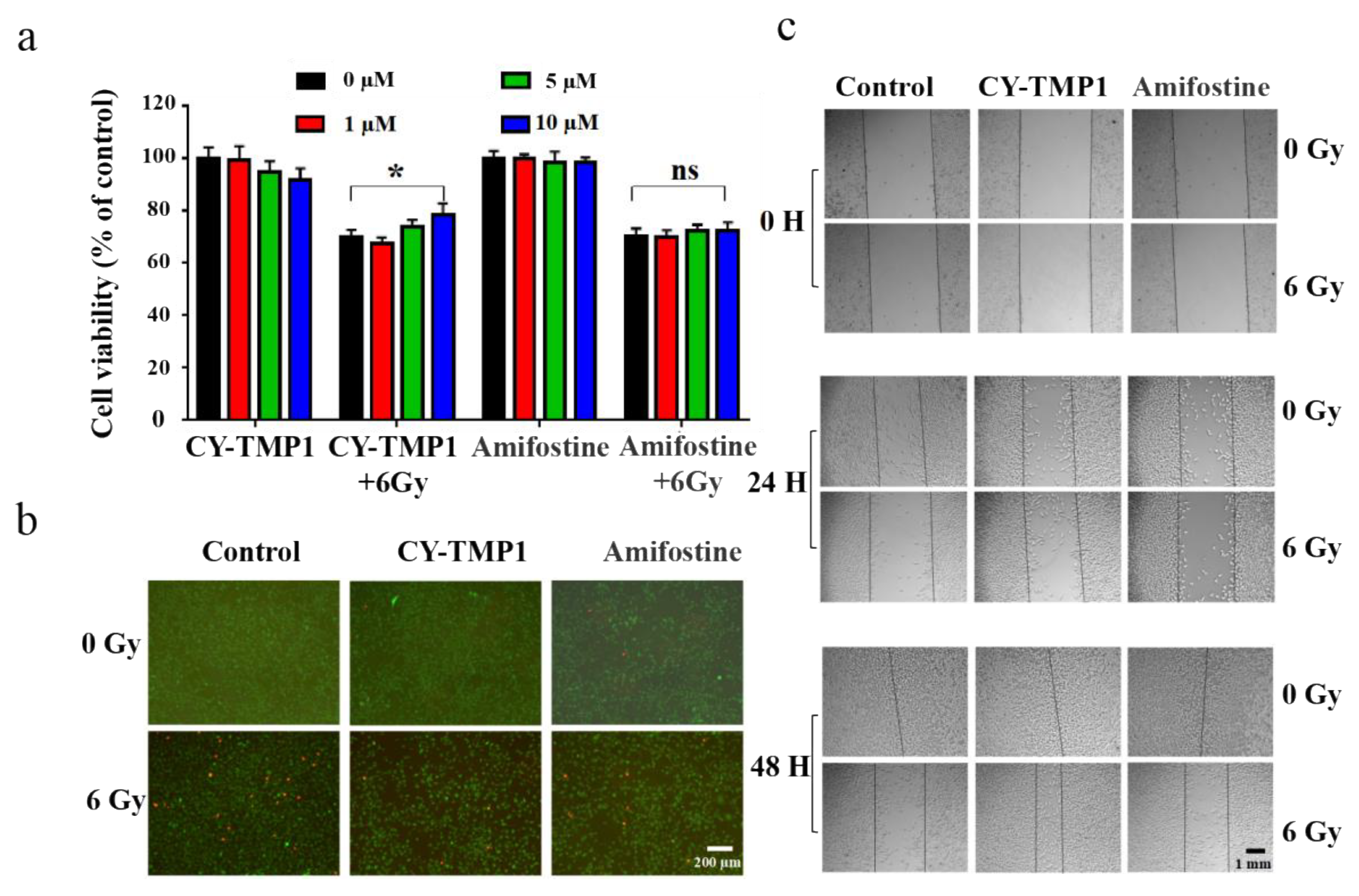
2.4. Radiation Protection and Wound Scratch Assay of CY-TMP1 on Human Fibroblast Cells
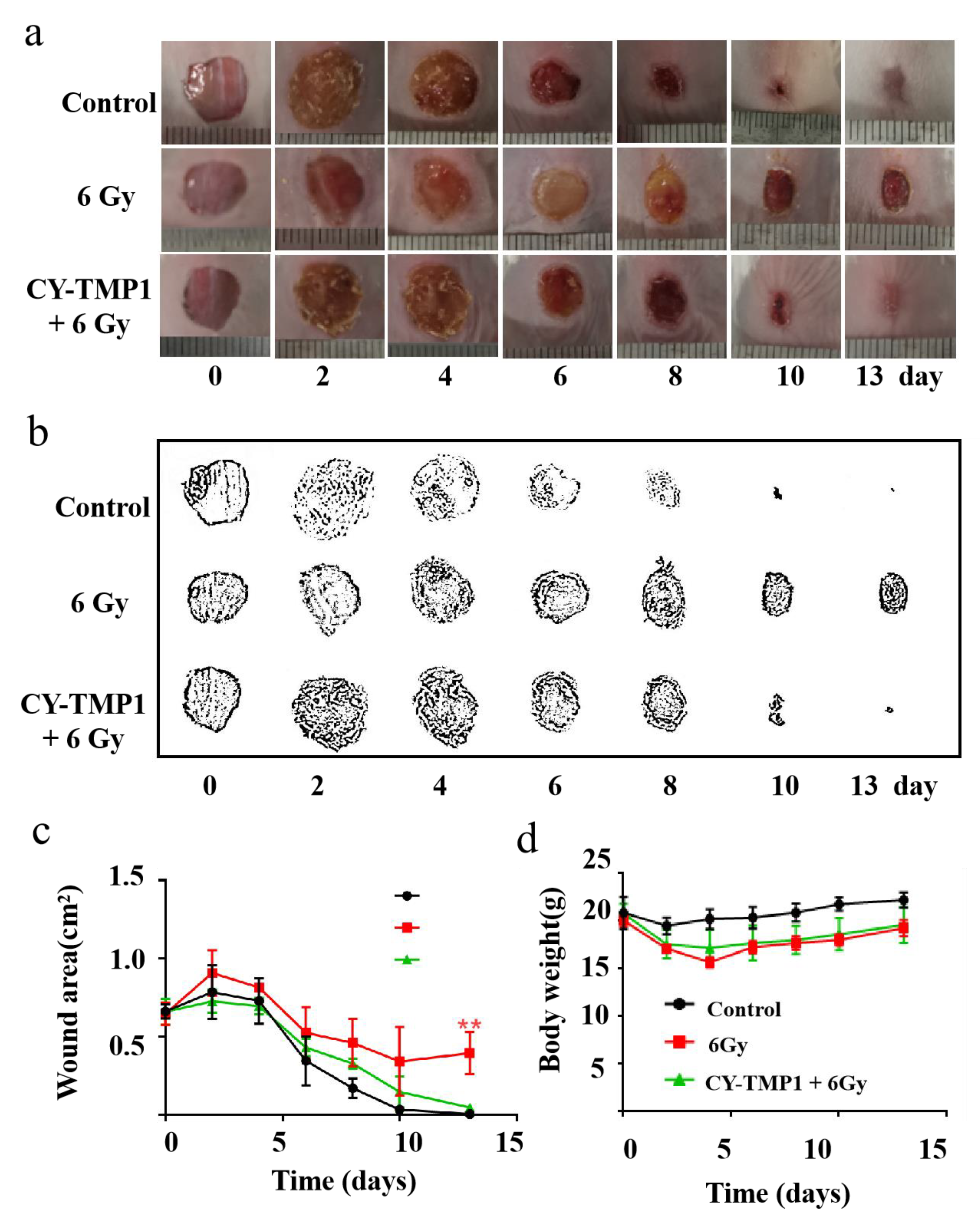
2.5. CY-TMP1 Facilitated Wound Healing Combined with Radiation Injury in Mice
3. Materials and Methods
3.1. Materials
3.2. Chemical Synthesis and Structure Characterization
3.2.1. Synthesis of CY-TMP1
3.2.2. Synthesis of CY-TMP2
3.2.3. Synthesis of CY-TMP3
3.3. In Vitro Experimental for Radioprotection Effect Evaluation
3.3.1. Cell Culture and Irradiation
3.3.2. CCK-8 Assay
3.3.3. Cell Colony-Forming Assay
3.3.4. Mitochondrial Localization, Mito-ROS and Membrane Potential Assay
3.3.5. Detection of Reactive Oxygen Species (ROS)
3.3.6. Comet Assay
3.3.7. Immunofluorescence
3.3.8. Calcein/PI staining
3.3.9. Wound Scratch Assay
3.4. In Vivo Experiments for Wound Closure Investigation
3.4.1. Animals
3.4.2. Histology and Immunofluorescence Staining
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hay, R.J.; Johns, N.E.; Williams, H.C.; Bolliger, I.W.; Dellavalle, R.P.; Margolis, D.J.; Marks, R.; Naldi, L.; Weinstock, M.A.; Wulf, S.K.; et al. The global burden of skin disease in 2010: An analysis of the prevalence and impact of skin conditions. J. Investig. Dermatol. 2014, 134, 1527–1534. [Google Scholar] [CrossRef] [Green Version]
- Devalia, H.L.; Mansfield, L. Radiotherapy and wound healing. Int. Wound J. 2008, 5, 40–44. [Google Scholar] [CrossRef]
- Gieringer, M.; Gosepath, J.; Naim, R. Radiotherapy and wound healing: Principles, management and prospects (review). Oncol. Rep. 2011, 26, 299–307. [Google Scholar] [CrossRef]
- Bourdais, R.; Achkar, S.; Honoré, C.; Faron, M.; Cavalcanti, A.; Auzac, G.; Ngo, C.; Haddag-Miliani, L.; Verret, B.; Dumont, S.; et al. Prospective evaluation of intensity-modulated radiotherapy toxicity in extremity soft tissue sarcomas patients: A role for irradiated healthy soft tissue volume? Clin. Transl. Radiat. Oncol. 2021, 29, 79–84. [Google Scholar] [CrossRef]
- Anderson, K.; Hamm, R.L. Factors That Impair Wound Healing. J. Am. Coll. Clin. Wound Spec. 2012, 4, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.W.; Carter, M.J.; Mashiach, E.; Guthrie, S.D. Vascular assessment of wound healing: A clinical review. Int. Wound J. 2017, 14, 460–469. [Google Scholar] [CrossRef]
- Ji, W.O.; Lee, M.H.; Kim, G.H.; Kim, E.H. Quantitation of the ROS production in plasma and radiation treatments of biotargets. Sci. Rep. 2019, 9, 19837. [Google Scholar] [CrossRef]
- Szumiel, I. Ionizing radiation-induced oxidative stress, epigenetic changes and genomic instability: The pivotal role of mitochondria. Int. J. Radiat. Biol. 2015, 91, 1–12. [Google Scholar] [CrossRef]
- Yoshida, T.; Goto, S.; Kawakatsu, M.; Urata, Y.; Li, T.S. Mitochondrial dysfunction, a probable cause of persistent oxidative stress after exposure to ionizing radiation. Free Radic. Res. 2012, 46, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Kam, W.W.; Banati, R.B. Effects of ionizing radiation on mitochondria. Free Radic. Biol. Med. 2013, 65, 607–619. [Google Scholar] [CrossRef]
- Kagan, V.E.; Tyurin, V.A.; Jiang, J.; Tyurina, Y.Y.; Ritov, V.B.; Amoscato, A.A.; Osipov, A.N.; Belikova, N.A.; Kapralov, A.A.; Kini, V.; et al. Cytochromec acts as a cardiolipin oxygenase required for release of proapoptotic factors. Nat. Chem. Biol. 2005, 1, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Jia, X. Polydatin alleviates radiation-induced testes injury by scavenging ROS and inhibiting apoptosis pathways. Med. Sci. Monit. 2018, 24, 8993–9000. [Google Scholar] [CrossRef]
- Li, H.; Yang, Z.Y.; Liu, C.; Zeng, Y.P.; Hao, Y.H.; Gu, Y.; Wang, W.D.; Li, R. PEGylated ceria nanoparticles used for radioprotection on human liver cells under γ-ray irradiation. Free Radic. Biol. Med. 2015, 87, 26–35. [Google Scholar] [CrossRef]
- Li, J.; Du, R.; Bian, Q.; Zhang, D.; Gao, S.; Yuan, A.; Ying, X.; Shen, Y.; Gao, J. Topical application of HA-g-TEMPO accelerates the acute wound healing via reducing reactive oxygen species (ROS) and promoting angiogenesis. Int. J. Pharm. 2021, 597, 120328. [Google Scholar] [CrossRef]
- Wu, H.; Li, F.; Wang, S.; Lu, J.; Li, J.; Du, Y.; Sun, X.; Chen, X.; Gao, J.; Ling, D. Ceria nanocrystals decorated mesoporous silica nanoparticle based ROS-scavenging tissue adhesive for highly efficient regenerative wound healing. Biomaterials 2018, 151, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.R.; Wang, J.Y.; Li, Y.Y.; Meng, Y.Y.; Zhang, Y.; Yang, F.J.; Xu, W.Q. Design and synthesis a mitochondria-targeted dihydronicotinamide as radioprotector. Free Radic. Biol. Med. 2019, 136, 45–51. [Google Scholar] [CrossRef]
- Jiang, J.; Belikova, N.A.; Hoye, A.T.; Zhao, Q.; Epperly, M.W.; Greenberger, J.S.; Wipf, P.; Kagan, V.E. A mitochondria-targeted nitroxide/hemigramicidin S conjugate protects mouse embryonic cells against gamma irradiation. Int. J. Radiat. Oncol. Biol. Phys. 2008, 70, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Jiang, J.; Belikova, N.A.; Stoyanovsky, D.A.; Kagan, V.E.; Mintz, A.H. Protection of normal brain cells from γ-irradiation-induced apoptosis by a mitochondria-targeted triphenyl-phosphonium-nitroxide: A possible utility in glioblastoma therapy. J. Neuro-Oncol. 2010, 100, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Tan, X.; Fang, S.; Wang, Y.; Liu, T.; Wang, X.; Yuan, Y.; Sun, H.; Qi, Q.; Shi, C. Mitochondria-targeted small-molecule fluorophores for dual modal cancer phototherapy. Adv. Funct. Mater. 2016, 26, 2826–2835. [Google Scholar] [CrossRef]
- Chen, S.; Yu, S.; Du, Z.; Huang, X.; He, M.; Long, S.; Liu, J.; Lan, Y.; Yang, D.; Wang, H.; et al. Synthesis of mitochondria-anchored nitroimidazoles with a versatile NIR fluorophore for hypoxic tumor-targeting imaging and chemoradiotherapy. J. Med. Chem. 2021, 64, 3381–3391. [Google Scholar] [CrossRef]
- Ménard, C.; Camphausen, K.; Muanza, T.; Sears-Crouse, N.; Smith, S.; Ben-Josef, E.; Coleman, C.N. Clinical trial of endorectal amifostine for radioprotection in patients with prostate cancer: Rationale and early results. Semin. Oncol. 2003, 30, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Luo, S.; Long, L.; Wang, Y.; Wang, D.; Fang, S.; Ouyang, Q.; Su, Y.; Cheng, T.; Shi, C. Structure-guided design and synthesis of a mitochondria-targeting near-infrared fluorophore with multimodal therapeutic activities. Adv. Mater. 2017, 29, 1704196. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Tan, X.; Qi, Q.; Guo, Q.; Ran, X.; Zhang, L.; Zhang, E.; Liang, Y.; Weng, L.; Zheng, H.; et al. A multifunctional heptamethine near-infrared dye for cancer theranosis. Biomaterials 2013, 34, 2244–2251. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Z.; Liu, H.; Huang, X.; Li, Y.; Wang, L.; Liu, J.; Long, S.; Li, R.; Xiang, Q.; Luo, S. Design and Synthesis of a Mitochondria-Targeting Radioprotectant for Promoting Skin Wound Healing Combined with Ionizing Radiation Injury. Pharmaceuticals 2022, 15, 721. https://doi.org/10.3390/ph15060721
Du Z, Liu H, Huang X, Li Y, Wang L, Liu J, Long S, Li R, Xiang Q, Luo S. Design and Synthesis of a Mitochondria-Targeting Radioprotectant for Promoting Skin Wound Healing Combined with Ionizing Radiation Injury. Pharmaceuticals. 2022; 15(6):721. https://doi.org/10.3390/ph15060721
Chicago/Turabian StyleDu, Zaizhi, Han Liu, Xie Huang, Yang Li, Liting Wang, Jing Liu, Shuang Long, Rong Li, Qiang Xiang, and Shenglin Luo. 2022. "Design and Synthesis of a Mitochondria-Targeting Radioprotectant for Promoting Skin Wound Healing Combined with Ionizing Radiation Injury" Pharmaceuticals 15, no. 6: 721. https://doi.org/10.3390/ph15060721
APA StyleDu, Z., Liu, H., Huang, X., Li, Y., Wang, L., Liu, J., Long, S., Li, R., Xiang, Q., & Luo, S. (2022). Design and Synthesis of a Mitochondria-Targeting Radioprotectant for Promoting Skin Wound Healing Combined with Ionizing Radiation Injury. Pharmaceuticals, 15(6), 721. https://doi.org/10.3390/ph15060721