
Carpaine Promotes Proliferation and Repair of H9c2 Cardiomyocytes after Oxidative Insults

 ,
,  , , and
, , and 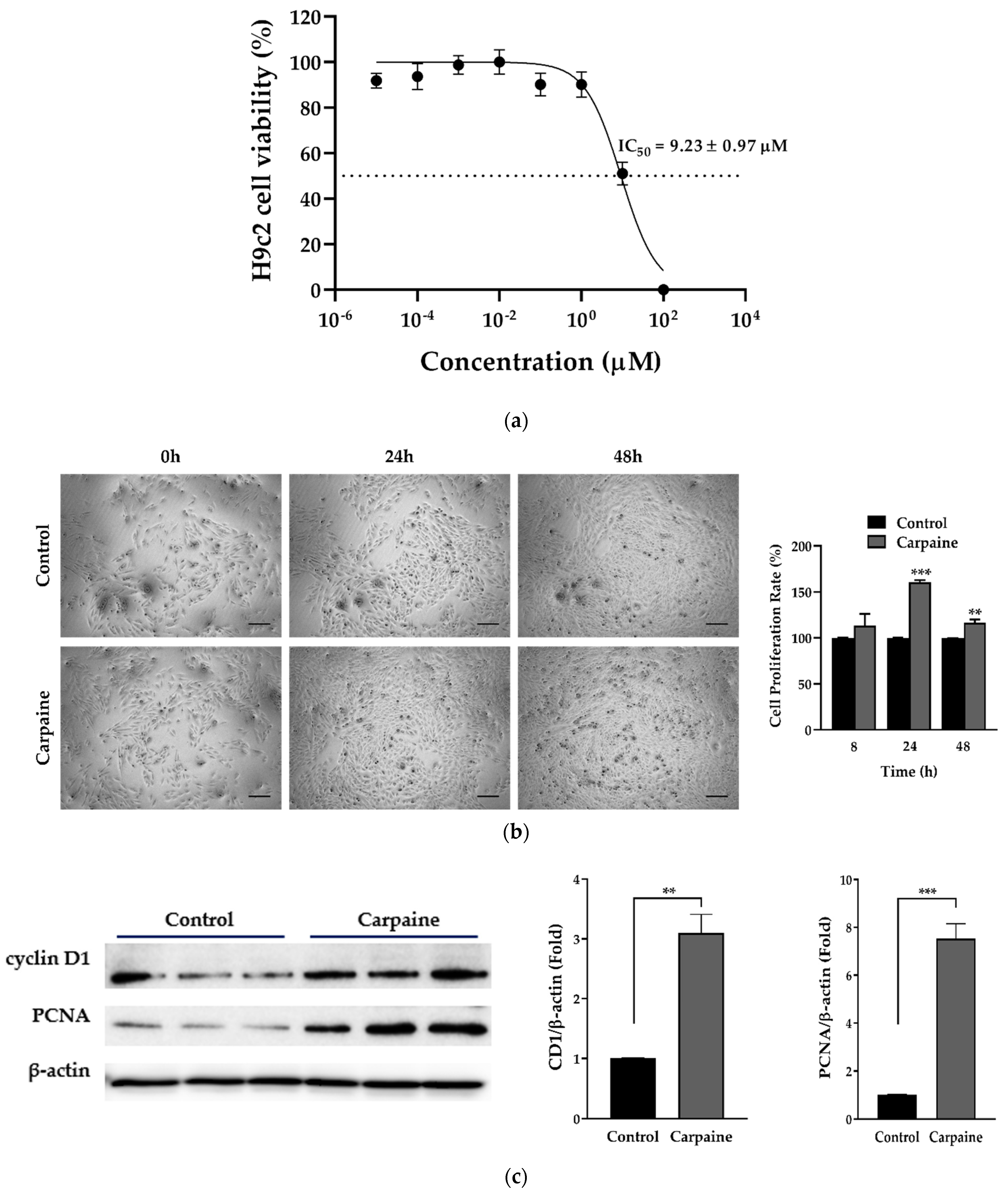{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
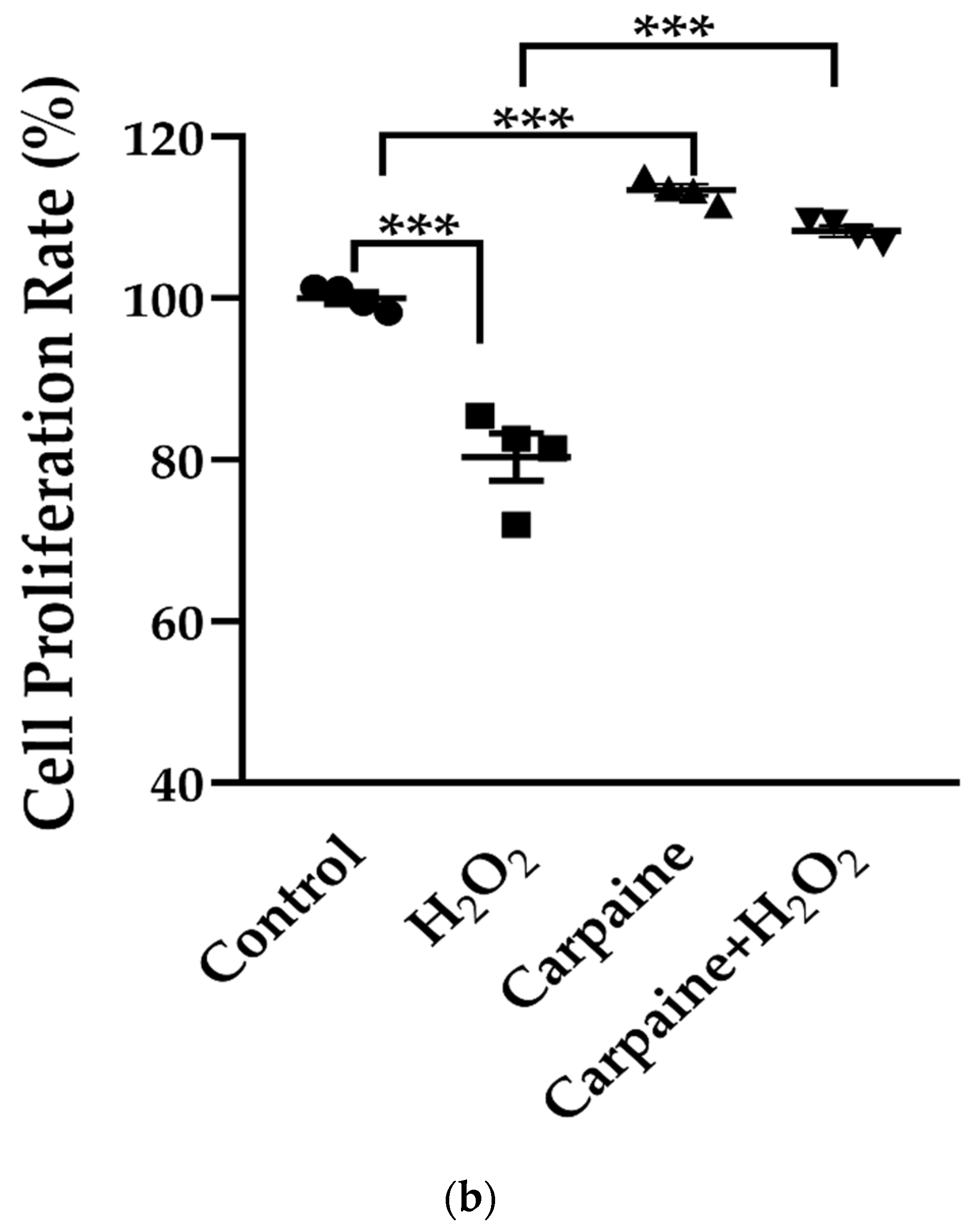
2.1. Carpaine Promotes Proliferation in H9c2 Cells
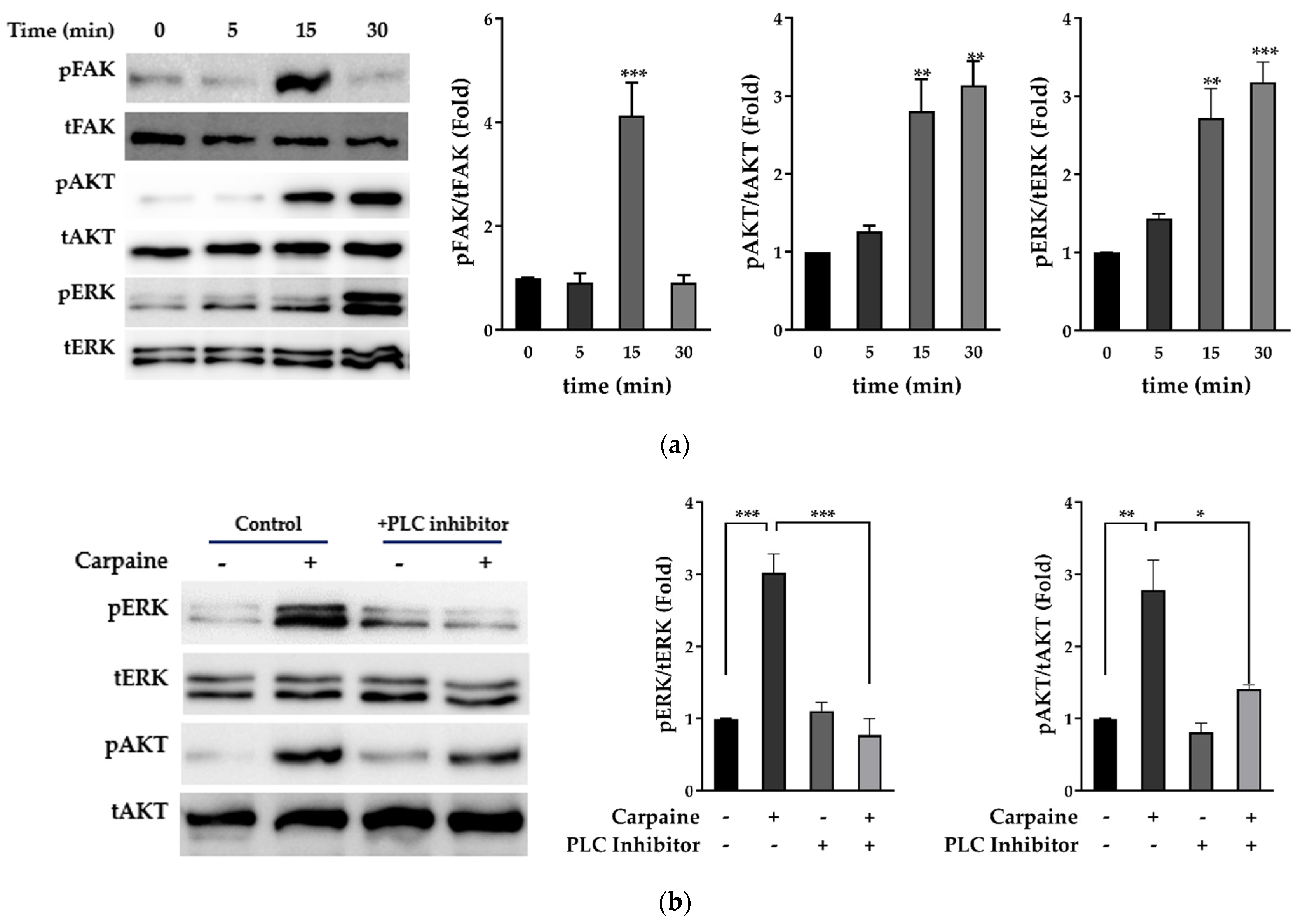
2.2. Carpaine Triggers Upstream FAK-Induced ERK1/2 and AKT Activation in H9c2 Cells
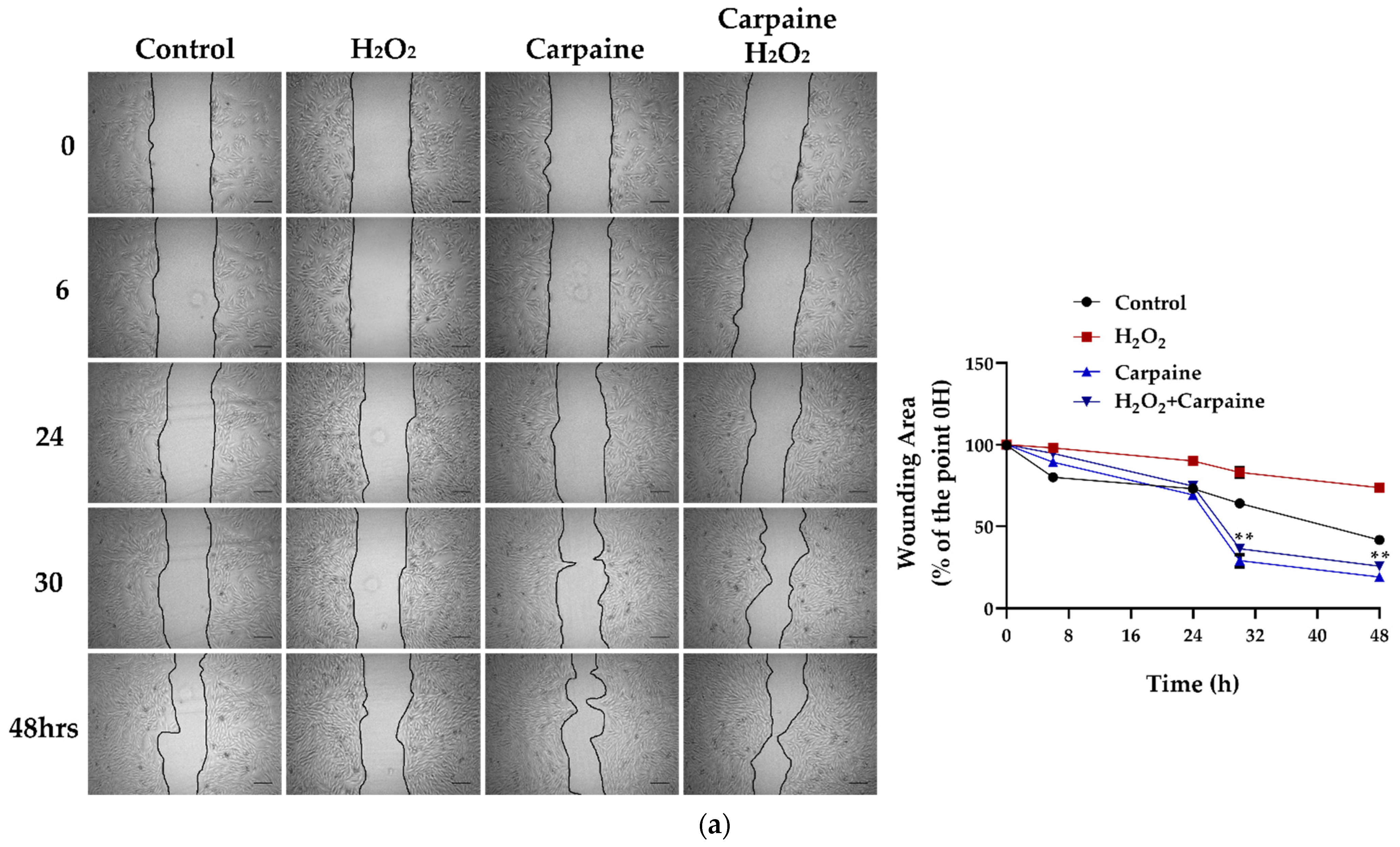
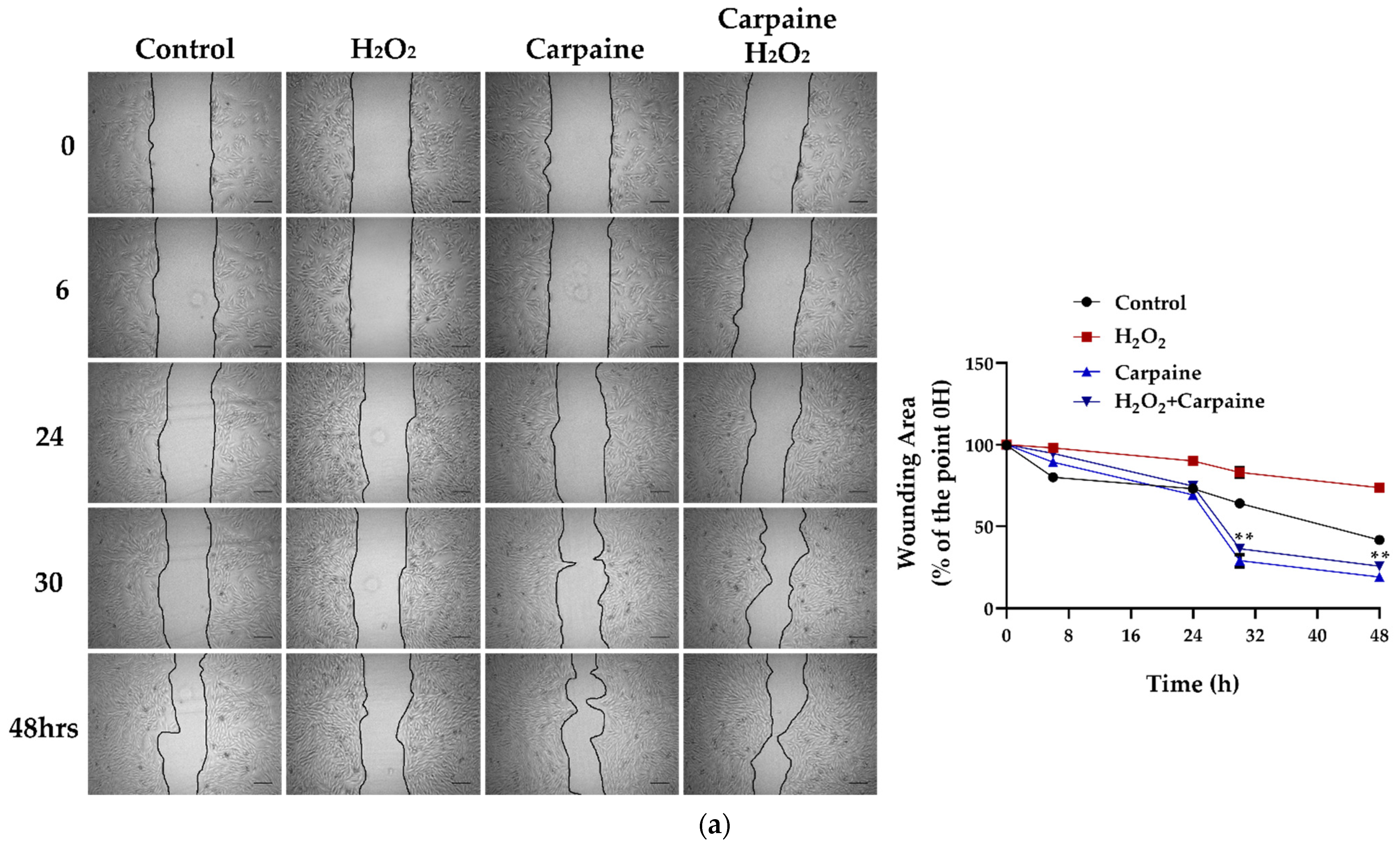
2.3. Carpaine Modulates the Wound-Healing Response by Stimulating Cardiomyocyte Proliferation in the Ischemic/Reperfusion Model
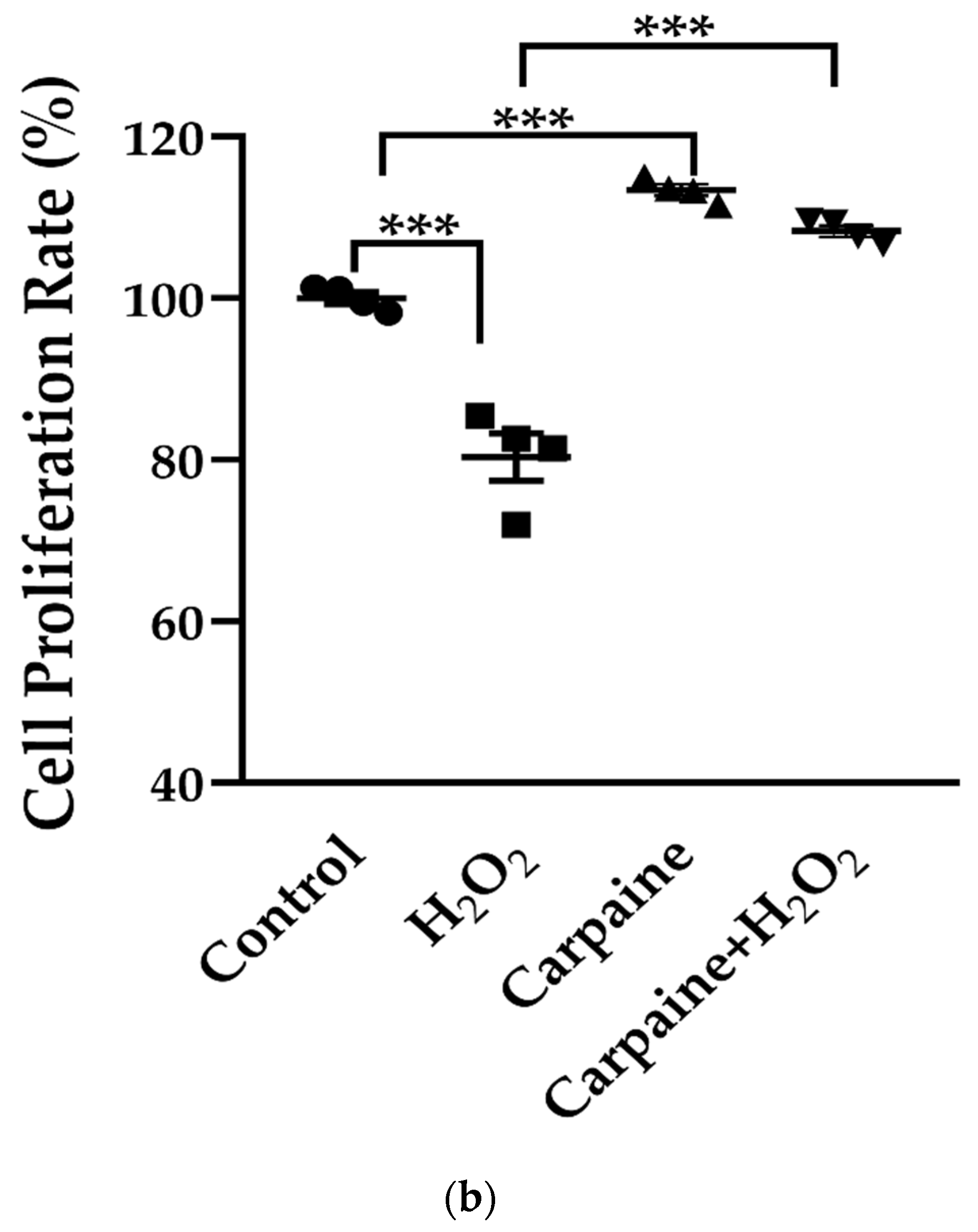
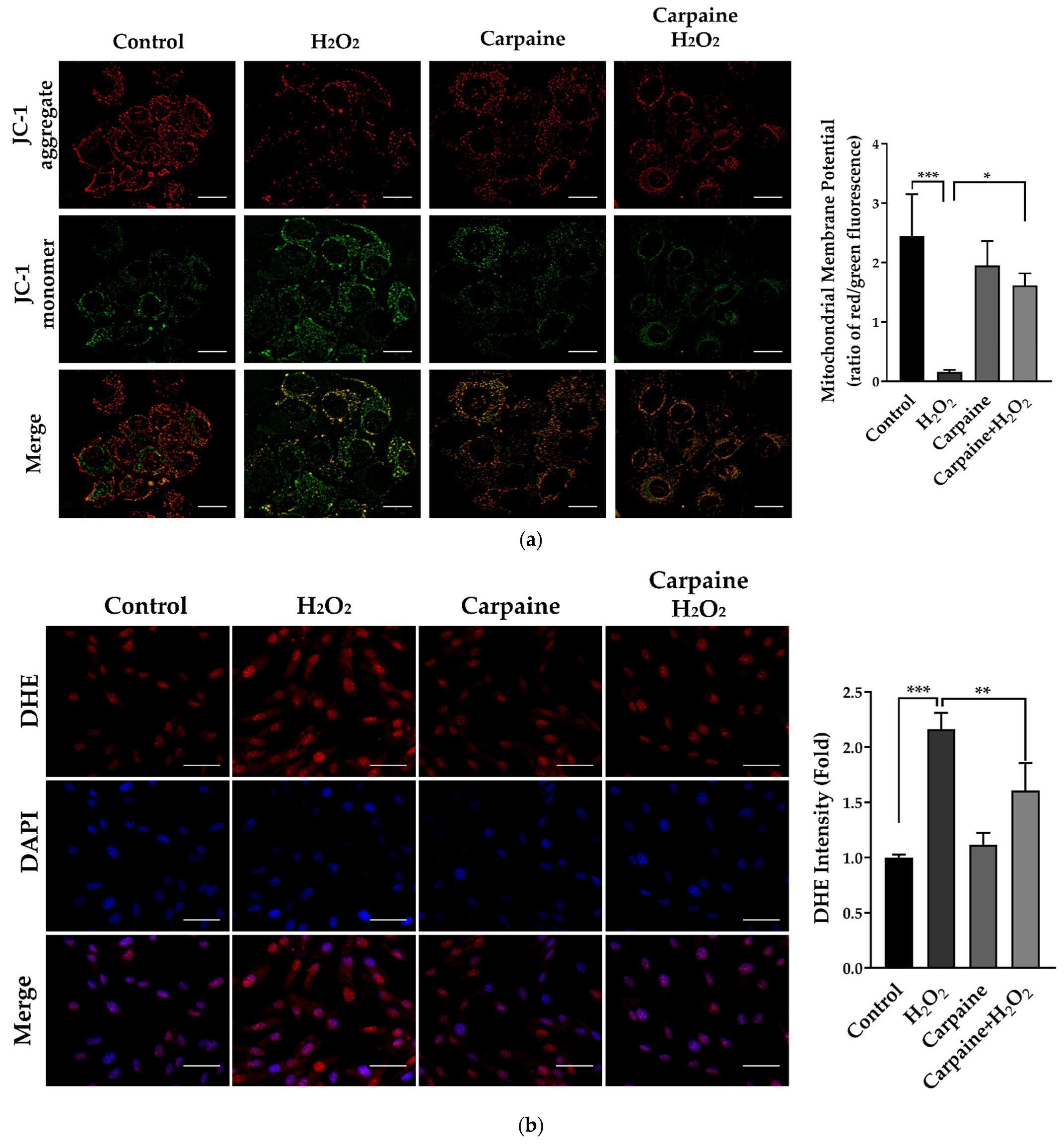
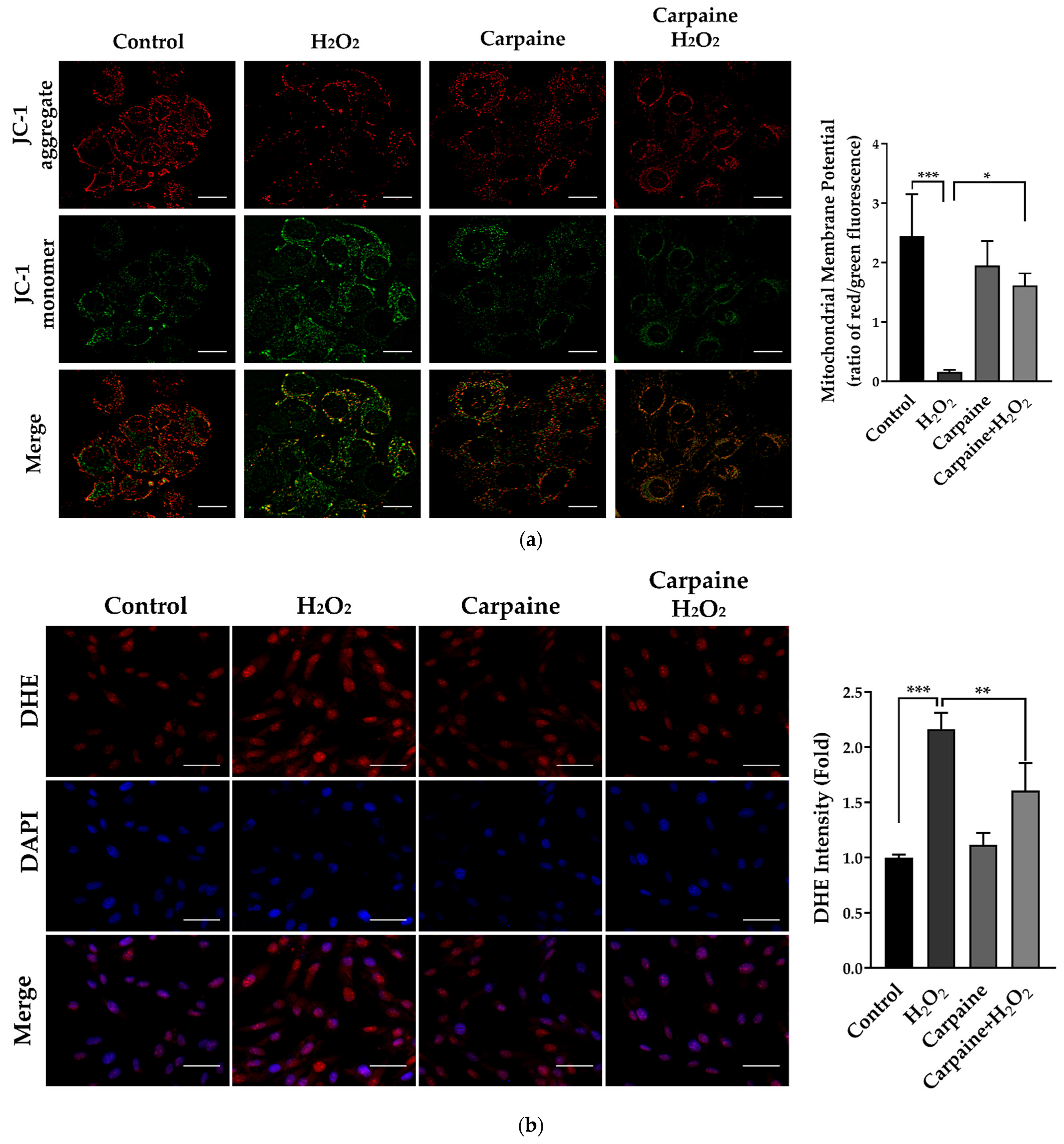
2.4. Carpaine Provides Protection against H2O2-Induced Oxidative Stress in H9c2 Cells
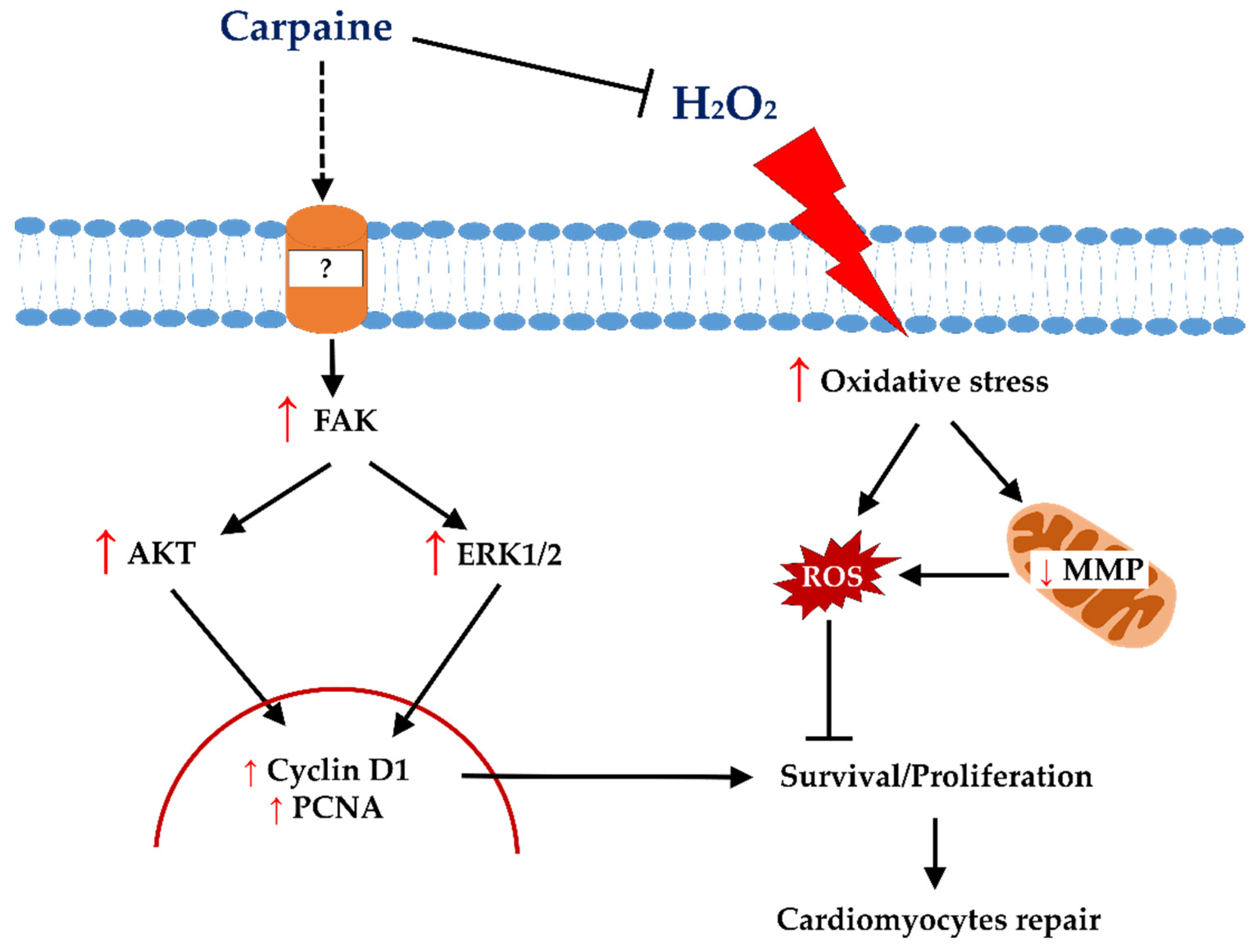
3. Discussion
4. Materials and Methods
4.1. Cell Line, Cell Culture, and Cell Treatment
4.2. Cytotoxicity and Proliferation Assay
4.3. Immunoblot Analysis
4.4. Wound Healing Assay
4.5. Assessment of the Mitochondrial Membrane Potential (MMP)
4.6. ROS Production Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yutzey, K.E. Cardiomyocyte Proliferation: Teaching an Old Dogma New Tricks. Circ. Res. 2017, 120, 627–629. [Google Scholar] [CrossRef]
- Soonpaa, M.H.; Kim, K.K.; Pajak, L.; Franklin, M.; Field, L.J. Cardiomyocyte DNA Synthesis and Binucleation during Murine Development. Am. J. Physiol. 1996, 271, H2183–H2189. [Google Scholar] [CrossRef] [PubMed]
- Neri, M.; Riezzo, I.; Pascale, N.; Pomara, C.; Turillazzi, E. Ischemia/Reperfusion Injury Following Acute Myocardial Infarction: A Critical Issue for Clinicians and Forensic Pathologists. Mediat. Inflamm. 2017, 2017, 7018393. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-W.; Chen, B.; Yang, X.; Fugate, J.A.; Kalucki, F.A.; Futakuchi-Tsuchida, A.; Couture, L.; Vogel, K.W.; Astley, C.A.; Baldessari, A.; et al. Human Embryonic Stem Cell-Derived Cardiomyocytes Restore Function in Infarcted Hearts of Non-Human Primates. Nat. Biotechnol. 2018, 36, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Senyo, S.E.; Lee, R.T.; Kühn, B. Cardiac Regeneration Based on Mechanisms of Cardiomyocyte Proliferation and Differentiation. Stem Cell Res. 2014, 13, 532–541. [Google Scholar] [CrossRef] [Green Version]
- Chien, K.R.; Frisén, J.; Fritsche-Danielson, R.; Melton, D.A.; Murry, C.E.; Weissman, I.L. Regenerating the Field of Cardiovascular Cell Therapy. Nat. Biotechnol. 2019, 37, 232–237. [Google Scholar] [CrossRef]
- Rajala, K.; Pekkanen-Mattila, M.; Aalto-Setälä, K. Cardiac Differentiation of Pluripotent Stem Cells. Stem Cells Int. 2011, 2011, 383709. [Google Scholar] [CrossRef] [Green Version]
- Fan, R.; Kim, N.-G.; Gumbiner, B.M. Regulation of Hippo Pathway by Mitogenic Growth Factors via Phosphoinositide 3-Kinase and Phosphoinositide-Dependent Kinase-1. Proc. Natl. Acad. Sci. USA 2013, 110, 2569–2574. [Google Scholar] [CrossRef] [Green Version]
- Zhen, L.; Zhao, Q.; Lü, J.; Deng, S.; Xu, Z.; Zhang, L.; Zhang, Y.; Fan, H.; Chen, X.; Liu, Z.; et al. MiR-301a-PTEN-AKT Signaling Induces Cardiomyocyte Proliferation and Promotes Cardiac Repair Post-MI. Mol. Ther.-Nucleic Acids 2020, 22, 251–262. [Google Scholar] [CrossRef]
- Liu, P.; Zhong, T.P. MAPK/ERK Signalling Is Required for Zebrafish Cardiac Regeneration. Biotechnol. Lett. 2017, 39, 1069–1077. [Google Scholar] [CrossRef]
- Uosaki, H.; Magadum, A.; Seo, K.; Fukushima, H.; Takeuchi, A.; Nakagawa, Y.; Moyes, K.W.; Narazaki, G.; Kuwahara, K.; Laflamme, M.; et al. Identification of Chemicals Inducing Cardiomyocyte Proliferation in Developmental Stage–Specific Manner With Pluripotent Stem Cells. Circ. Cardiovasc. Genet. 2013, 6, 624–633. [Google Scholar] [CrossRef] [Green Version]
- Carter, E.P.; Fearon, A.E.; Grose, R.P. Careless Talk Costs Lives: Fibroblast Growth Factor Receptor Signalling and the Consequences of Pathway Malfunction. Trends Cell Biol. 2015, 25, 221–233. [Google Scholar] [CrossRef]
- Leach, J.P.; Martin, J.F. Cardiomyocyte Proliferation for Therapeutic Regeneration. Curr. Cardiol. Rep. 2018, 20, 63. [Google Scholar] [CrossRef]
- Allijn, I.E.; Czarny, B.M.S.; Wang, X.; Chong, S.Y.; Weiler, M.; da Silva, A.E.; Metselaar, J.M.; Lam, C.S.P.; Pastorin, G.; de Kleijn, D.P.V.; et al. Liposome Encapsulated Berberine Treatment Attenuates Cardiac Dysfunction after Myocardial Infarction. J. Control. Release 2017, 247, 127–133. [Google Scholar] [CrossRef]
- Gogna, N.; Hamid, N.; Dorai, K. Metabolomic Profiling of the Phytomedicinal Constituents of Carica Papaya L. Leaves and Seeds by 1H NMR Spectroscopy and Multivariate Statistical Analysis. J. Pharm. Biomed. Anal. 2015, 115, 74–85. [Google Scholar] [CrossRef]
- Runnie, I.; Salleh, M.N.; Mohamed, S.; Head, R.J.; Abeywardena, M.Y. Vasorelaxation Induced by Common Edible Tropical Plant Extracts in Isolated Rat Aorta and Mesenteric Vascular Bed. J. Ethnopharmacol. 2004, 92, 311–316. [Google Scholar] [CrossRef]
- Julianti, T.; Oufir, M.; Hamburger, M. Quantification of the Antiplasmodial Alkaloid Carpaine in Papaya (Carica Papaya) Leaves. Planta Med. 2014, 80, 1138–1142. [Google Scholar] [CrossRef] [Green Version]
- Yap, J.Y.; Hii, C.L.; Ong, S.P.; Lim, K.H.; Abas, F.; Pin, K.Y. Effects of Drying on Total Polyphenols Content and Antioxidant Properties of Carica Papaya Leaves. J. Sci. Food Agric. 2020, 100, 2932–2937. [Google Scholar] [CrossRef]
- Nugroho, A.; Heryani, H.; Choi, J.S.; Park, H.-J. Identification and Quantification of Flavonoids in Carica Papaya Leaf and Peroxynitrite-Scavenging Activity. Asian Pac. J. Trop. Biomed. 2017, 7, 208–213. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Chou, H.-C.; Lin, S.-T.; Chen, Y.-H.; Chang, Y.-J.; Chen, L.; Chan, H.-L. Cardioprotective Effects of Quercetin in Cardiomyocyte under Ischemia/Reperfusion Injury. Evid. Based. Complement. Alternat. Med. 2013, 2013, 364519. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Liao, Z.; Huang, L.; Liu, D.; Yin, D.; He, M. Kaempferol Protects Cardiomyocytes against Anoxia/Reoxygenation Injury via Mitochondrial Pathway Mediated by SIRT1. Eur. J. Pharmacol. 2015, 761, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Burdick, E.M. Carpaine: An Alkaloid of Carica Papaya—Its Chemistry and Pharmacology. Econ. Bot. 1971, 25, 363–365. [Google Scholar] [CrossRef]
- Hornick, C.A.; Sanders, L.I.; Lin, Y.C. Effect of Carpaine, a Papaya Alkaloid, on the Circulatory Function in the Rat. Res. Commun. Chem. Pathol. Pharmacol. 1978, 22, 277–289. [Google Scholar]
- Umesh, H.R.; Ramesh, K.V.; Devaraju, K.S. Molecular Docking Studies of Phytochemicals against Trehalose–6–Phosphate Phosphatases of Pathogenic Microbes. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- DiMichele, L.A.; Hakim, Z.S.; Sayers, R.L.; Rojas, M.; Schwartz, R.J.; Mack, C.P.; Taylor, J.M. Transient Expression of FRNK Reveals Stage-Specific Requirement for Focal Adhesion Kinase Activity in Cardiac Growth. Circ. Res. 2009, 104, 1201–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, J.T.; Conlon, F.L.; Mack, C.P.; Taylor, J.M. Focal Adhesion Kinase Is Essential for Cardiac Looping and Multichamber Heart Formation. Genesis 2010, 48, 492–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yellon, D.M.; Hausenloy, D.J. Myocardial Reperfusion Injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef]
- Li, J.-M.; Zhou, H.; Cai, Q.; Xiao, G.-X. Role of Mitochondrial Dysfunction in Hydrogen Peroxide-Induced Apoptosis of Intestinal Epithelial Cells. World J. Gastroenterol. 2003, 9, 562–567. [Google Scholar] [CrossRef]
- Puente, B.N.; Kimura, W.; Muralidhar, S.A.; Moon, J.; Amatruda, J.F.; Phelps, K.L.; Grinsfelder, D.; Rothermel, B.A.; Chen, R.; Garcia, J.A.; et al. The Oxygen-Rich Postnatal Environment Induces Cardiomyocyte Cell-Cycle Arrest through DNA Damage Response. Cell 2014, 157, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, X.; Capasso, J.M.; Gerdes, A.M. Rapid Transition of Cardiac Myocytes from Hyperplasia to Hypertrophy During Postnatal Development. J. Mol. Cell. Cardiol. 1996, 28, 1737–1746. [Google Scholar] [CrossRef]
- Deng, S.; Zhao, Q.; Zhen, L.; Zhang, C.; Liu, C.; Wang, G.; Zhang, L.; Bao, L.; Lu, Y.; Meng, L.; et al. Neonatal Heart-Enriched MiR-708 Promotes Proliferation and Stress Resistance of Cardiomyocytes in Rodents. Theranostics 2017, 7, 1953–1965. [Google Scholar] [CrossRef]
- Wang, W.E.; Li, L.; Xia, X.; Fu, W.; Liao, Q.; Lan, C.; Yang, D.; Chen, H.; Yue, R.; Zeng, C.; et al. Dedifferentiation, Proliferation, and Redifferentiation of Adult Mammalian Cardiomyocytes After Ischemic Injury. Circulation 2017, 136, 834–848. [Google Scholar] [CrossRef]
- Yap, J.Y.; Hii, C.L.; Ong, S.P.; Lim, K.H.; Abas, F.; Pin, K.Y. Quantification of Carpaine and Antioxidant Properties of Extracts from Carica Papaya Plant Leaves and Stalks. J. Bioresour. Bioprod. 2021, 6, 350–358. [Google Scholar] [CrossRef]
- Schnabel, K.; Wu, C.-C.; Kurth, T.; Weidinger, G. Regeneration of Cryoinjury Induced Necrotic Heart Lesions in Zebrafish Is Associated with Epicardial Activation and Cardiomyocyte Proliferation. PLoS ONE 2011, 6, e18503. [Google Scholar] [CrossRef]
- Jopling, C.; Sleep, E.; Raya, M.; Martí, M.; Raya, A.; Izpisúa Belmonte, J.C. Zebrafish Heart Regeneration Occurs by Cardiomyocyte Dedifferentiation and Proliferation. Nature 2010, 464, 606–609. [Google Scholar] [CrossRef]
- Hashmi, S.; Ahmad, H.R. Molecular Switch Model for Cardiomyocyte Proliferation. Cell Regen. 2019, 8, 12–20. [Google Scholar] [CrossRef]
- Blankesteijn, W.M. Interventions in WNT Signaling to Induce Cardiomyocyte Proliferation: Crosstalk with Other Pathways. Mol. Pharmacol. 2020, 97, 90–101. [Google Scholar] [CrossRef]
- Parekh, P.; Motiwale, L.; Naik, N.; Rao, K.V.K. Downregulation of Cyclin D1 Is Associated with Decreased Levels of P38 MAP Kinases, Akt/PKB and Pak1 during Chemopreventive Effects of Resveratrol in Liver Cancer Cells. Exp. Toxicol. Pathol. 2011, 63, 167–173. [Google Scholar] [CrossRef]
- Beigi, F.; Schmeckpeper, J.; Pow-Anpongkul, P.; Payne, J.A.; Zhang, L.; Zhang, Z.; Huang, J.; Mirotsou, M.; Dzau, V.J. C3orf58, a Novel Paracrine Protein, Stimulates Cardiomyocyte Cell-Cycle Progression through the PI3K-AKT-CDK7 Pathway. Circ. Res. 2013, 113, 372–380. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zhang, H.; Beach, D. D Type Cyclins Associate with Multiple Protein Kinases and the DNA Replication and Repair Factor PCNA. Cell 1992, 71, 505–514. [Google Scholar] [CrossRef]
- Engel, F.B.; Schebesta, M.; Duong, M.T.; Lu, G.; Ren, S.; Madwed, J.B.; Jiang, H.; Wang, Y.; Keating, M.T. P38 MAP Kinase Inhibition Enables Proliferation of Adult Mammalian Cardiomyocytes. Genes Dev. 2005, 19, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saurin, A.T.; Martin, J.L.; Heads, R.J.; Foley, C.; Mockridge, J.W.; Wright, M.J.; Wang, Y.; Marber, M.S. The Role of Differential Activation of P38-Mitogen-Activated Protein Kinase in Preconditioned Ventricular Myocytes. FASEB J. 2000, 14, 2237–2246. [Google Scholar] [CrossRef] [PubMed]
- Ieda, M.; Tsuchihashi, T.; Ivey, K.N.; Ross, R.S.; Hong, T.-T.; Shaw, R.M.; Srivastava, D. Cardiac Fibroblasts Regulate Myocardial Proliferation through Beta1 Integrin Signaling. Dev. Cell 2009, 16, 233–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Uva, G.; Aharonov, A.; Lauriola, M.; Kain, D.; Yahalom-Ronen, Y.; Carvalho, S.; Weisinger, K.; Bassat, E.; Rajchman, D.; Yifa, O.; et al. ERBB2 Triggers Mammalian Heart Regeneration by Promoting Cardiomyocyte Dedifferentiation and Proliferation. Nat. Cell Biol. 2015, 17, 627–638. [Google Scholar] [CrossRef]
- Cheng, Z.; DiMichele, L.A.; Hakim, Z.S.; Rojas, M.; Mack, C.P.; Taylor, J.M. Targeted Focal Adhesion Kinase Activation in Cardiomyocytes Protects the Heart from Ischemia/Reperfusion Injury. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 924–933. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Zhang, D.; Liu, F.; Yang, Y.; Xu, H. Artesunate Alleviates Myocardial Ischemia/Reperfusion-Induced Myocardial Necrosis in Rats and Hypoxia/Reoxygenation-Induced Apoptosis in H9C2 Cells via Regulating the FAK/PI3K/Akt Pathway. Ann. Transl. Med. 2020, 8, 1291. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Tsang, A.; Yellon, D.M. The Reperfusion Injury Salvage Kinase Pathway: A Common Target for Both Ischemic Preconditioning and Postconditioning. Trends Cardiovasc. Med. 2005, 15, 69–75. [Google Scholar] [CrossRef]
- Cadenas, S. ROS and Redox Signaling in Myocardial Ischemia-Reperfusion Injury and Cardioprotection. Free Radic. Biol. Med. 2018, 117, 76–89. [Google Scholar] [CrossRef]
- Brand, M.D. Mitochondrial Generation of Superoxide and Hydrogen Peroxide as the Source of Mitochondrial Redox Signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef]
- Andrienko, T.; Pasdois, P.; Rossbach, A.; Halestrap, A.P. Real-Time Fluorescence Measurements of ROS and [Ca2+] in Ischemic/Reperfused Rat Hearts: Detectable Increases Occur Only after Mitochondrial Pore Opening and Are Attenuated by Ischemic Preconditioning. PLoS ONE 2016, 11, e0167300. [Google Scholar] [CrossRef] [Green Version]
- Bugger, H.; Pfeil, K. Mitochondrial ROS in Myocardial Ischemia Reperfusion and Remodeling. Biochim. Biophys. Acta-Mol. Basis Dis. 2020, 1866, 165768. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudi, S.; Chin, Y.-Z.; Wasli, N.S.; Fong, S.-Y.; Shimmi, S.C.; How, S.-E.; Sunggip, C. Carpaine Promotes Proliferation and Repair of H9c2 Cardiomyocytes after Oxidative Insults. Pharmaceuticals 2022, 15, 230. https://doi.org/10.3390/ph15020230
Sudi S, Chin Y-Z, Wasli NS, Fong S-Y, Shimmi SC, How S-E, Sunggip C. Carpaine Promotes Proliferation and Repair of H9c2 Cardiomyocytes after Oxidative Insults. Pharmaceuticals. 2022; 15(2):230. https://doi.org/10.3390/ph15020230
Chicago/Turabian StyleSudi, Suhaini, Yee-Zheng Chin, Nur Syafinaz Wasli, Siat-Yee Fong, Sadia Choudhury Shimmi, Siew-Eng How, and Caroline Sunggip. 2022. "Carpaine Promotes Proliferation and Repair of H9c2 Cardiomyocytes after Oxidative Insults" Pharmaceuticals 15, no. 2: 230. https://doi.org/10.3390/ph15020230
APA StyleSudi, S., Chin, Y.-Z., Wasli, N. S., Fong, S.-Y., Shimmi, S. C., How, S.-E., & Sunggip, C. (2022). Carpaine Promotes Proliferation and Repair of H9c2 Cardiomyocytes after Oxidative Insults. Pharmaceuticals, 15(2), 230. https://doi.org/10.3390/ph15020230