
First In Silico Screening of Insect Molecules for Identification of Novel Anti-Parasitic Compounds

 , ,
, ,
Abstract
:
1. Introduction
2. Results
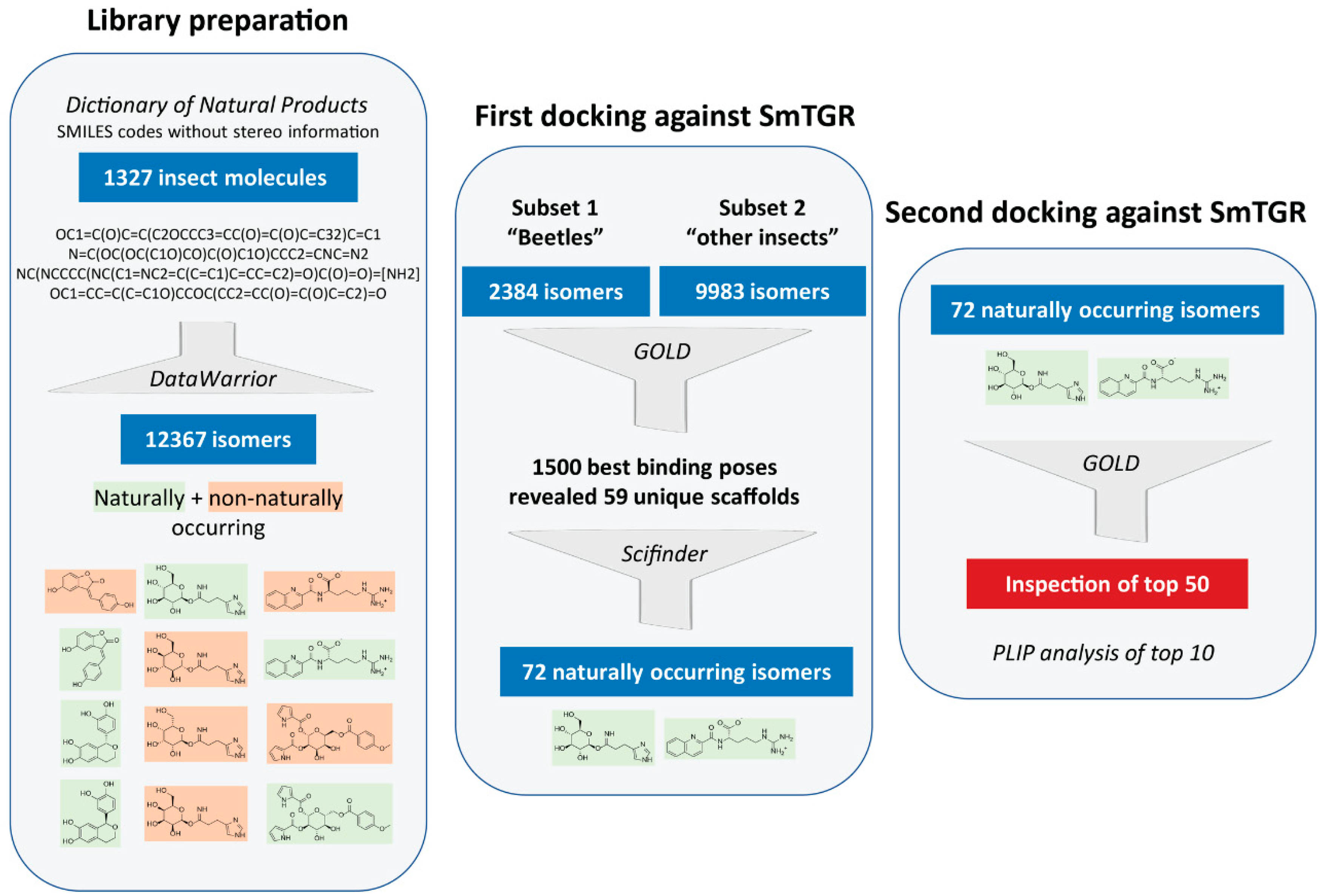
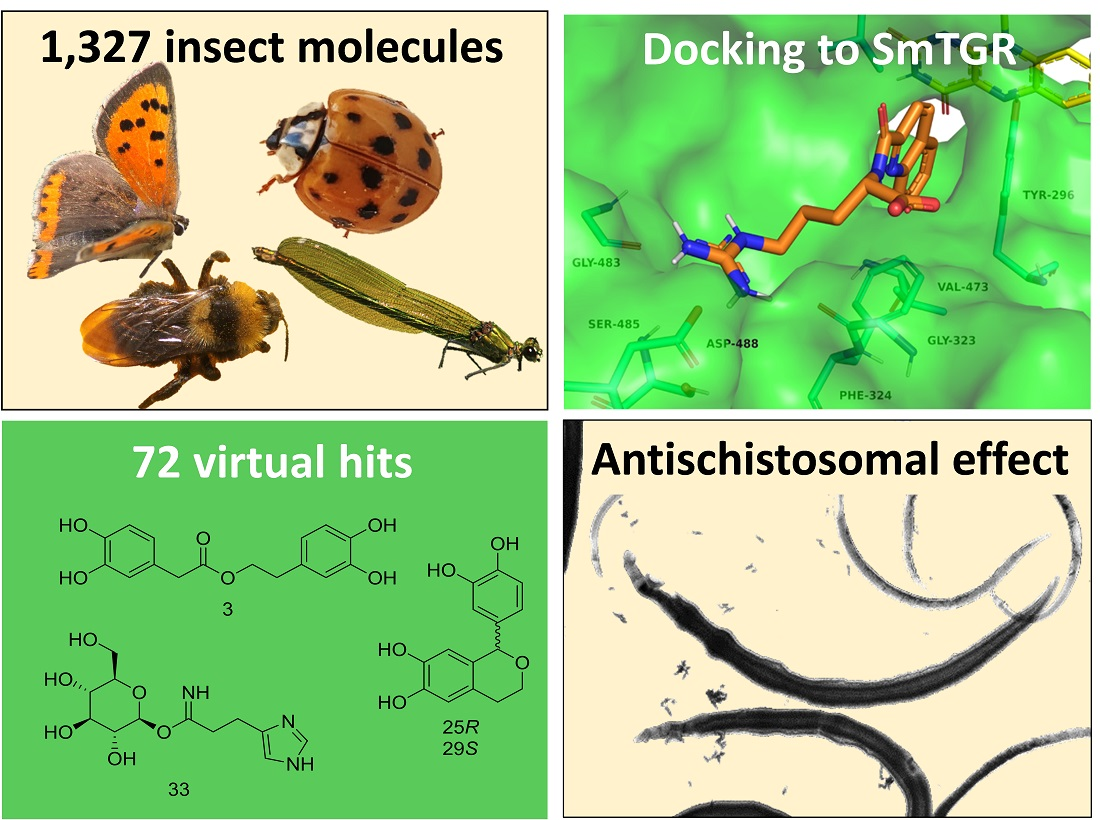
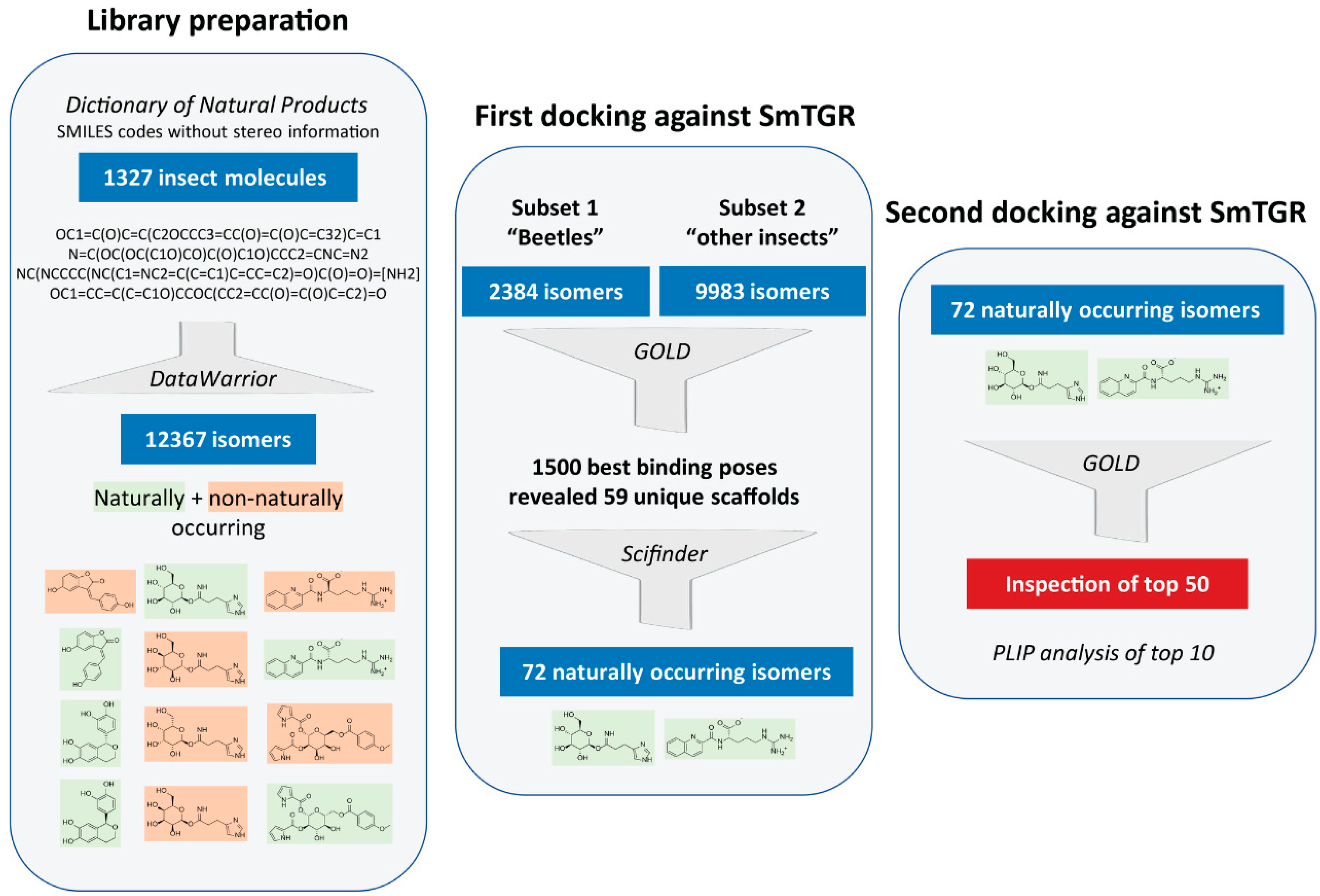
2.1. Creation of a Library of Insect Compounds Suitable for Docking Studies
2.2. Prioritization of SmTGR Ligand-Binding Pockets
2.3. Docking Results
2.3.1. Docking Predicts SmTGR-Inhibiting Insect Compounds
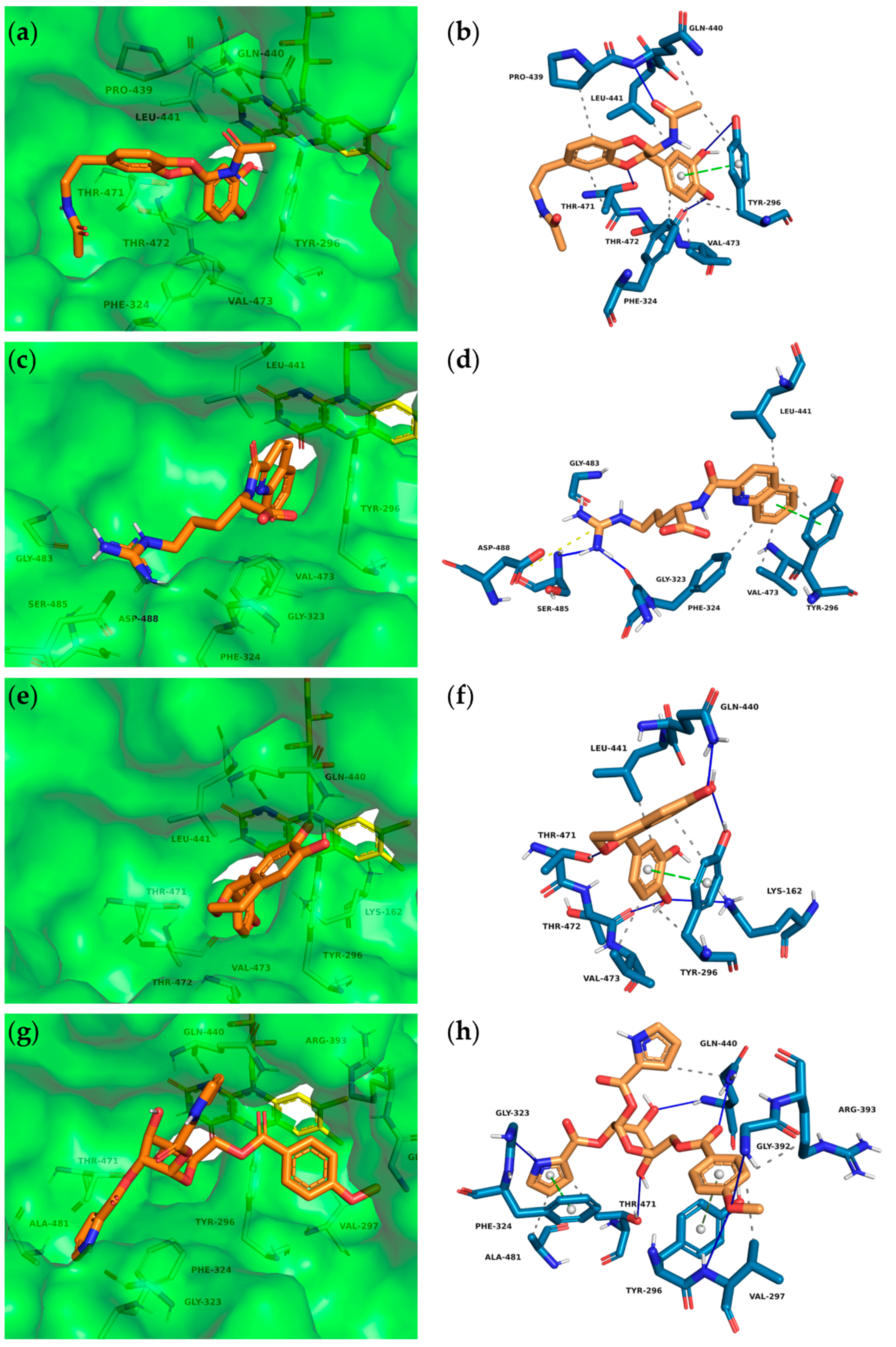
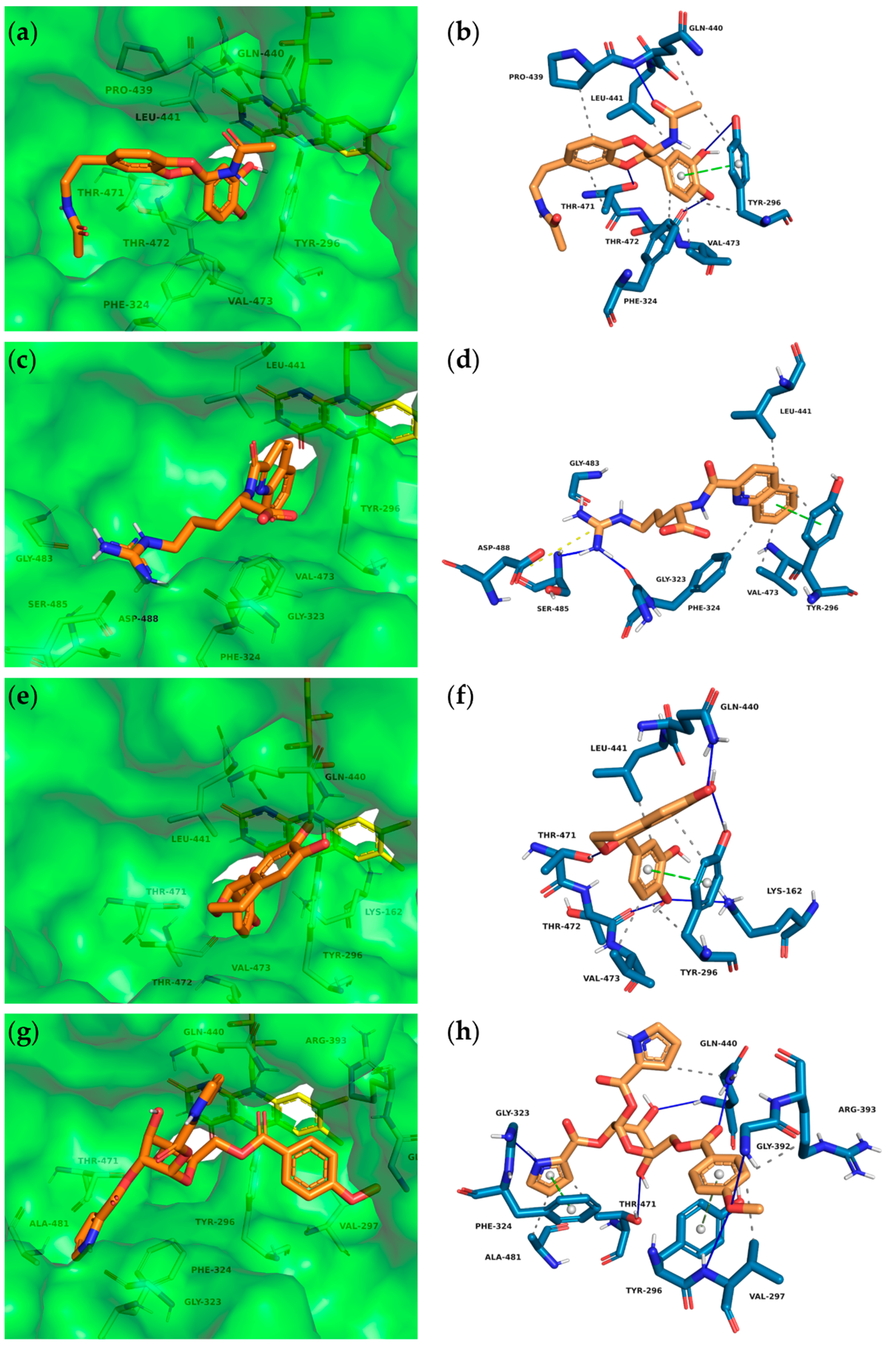
2.3.2. Predicted Interactions within the SmTGR Cavity
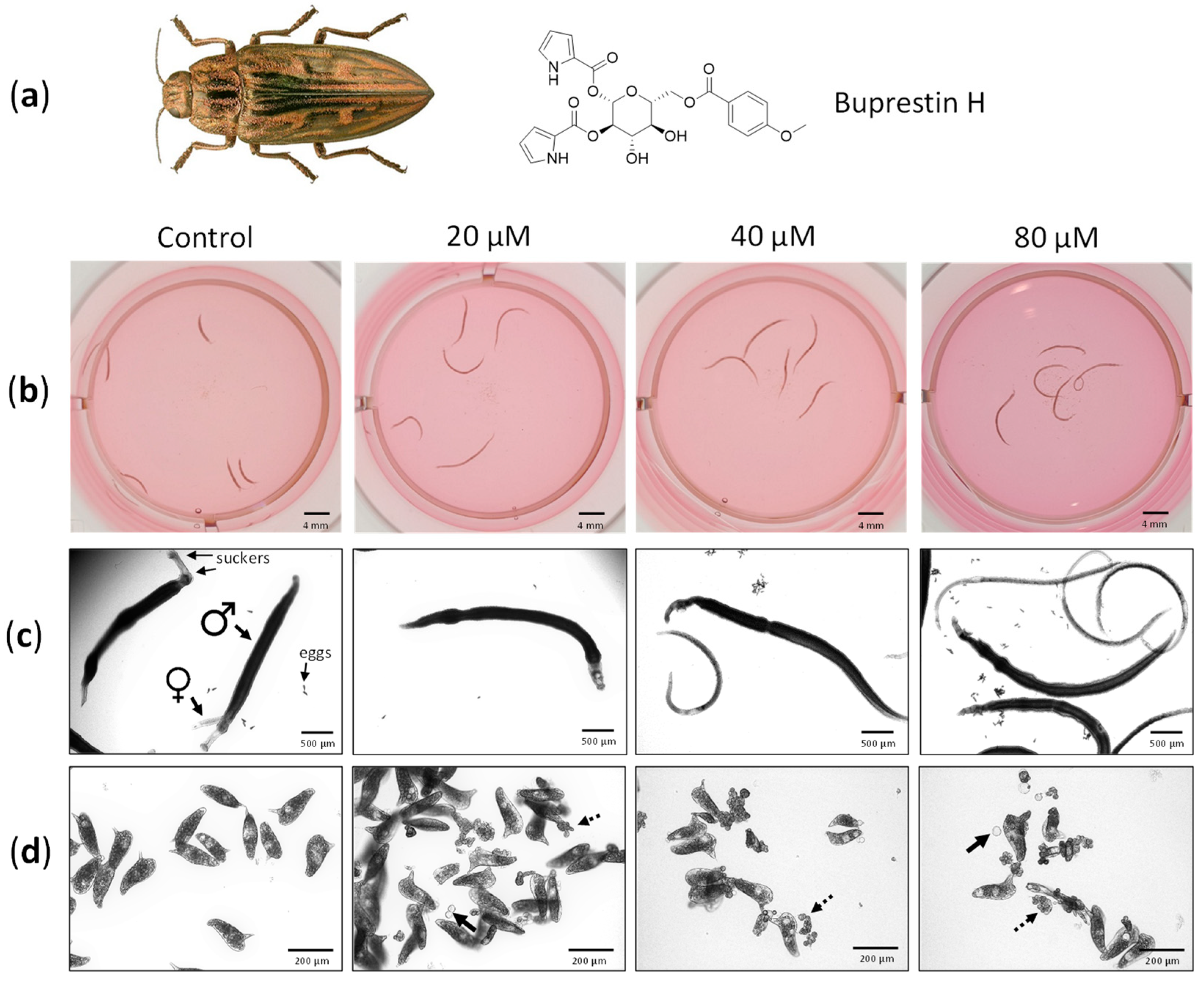
2.4. Validation of Buprestin H Activity against Schistosoma mansoni In Vitro
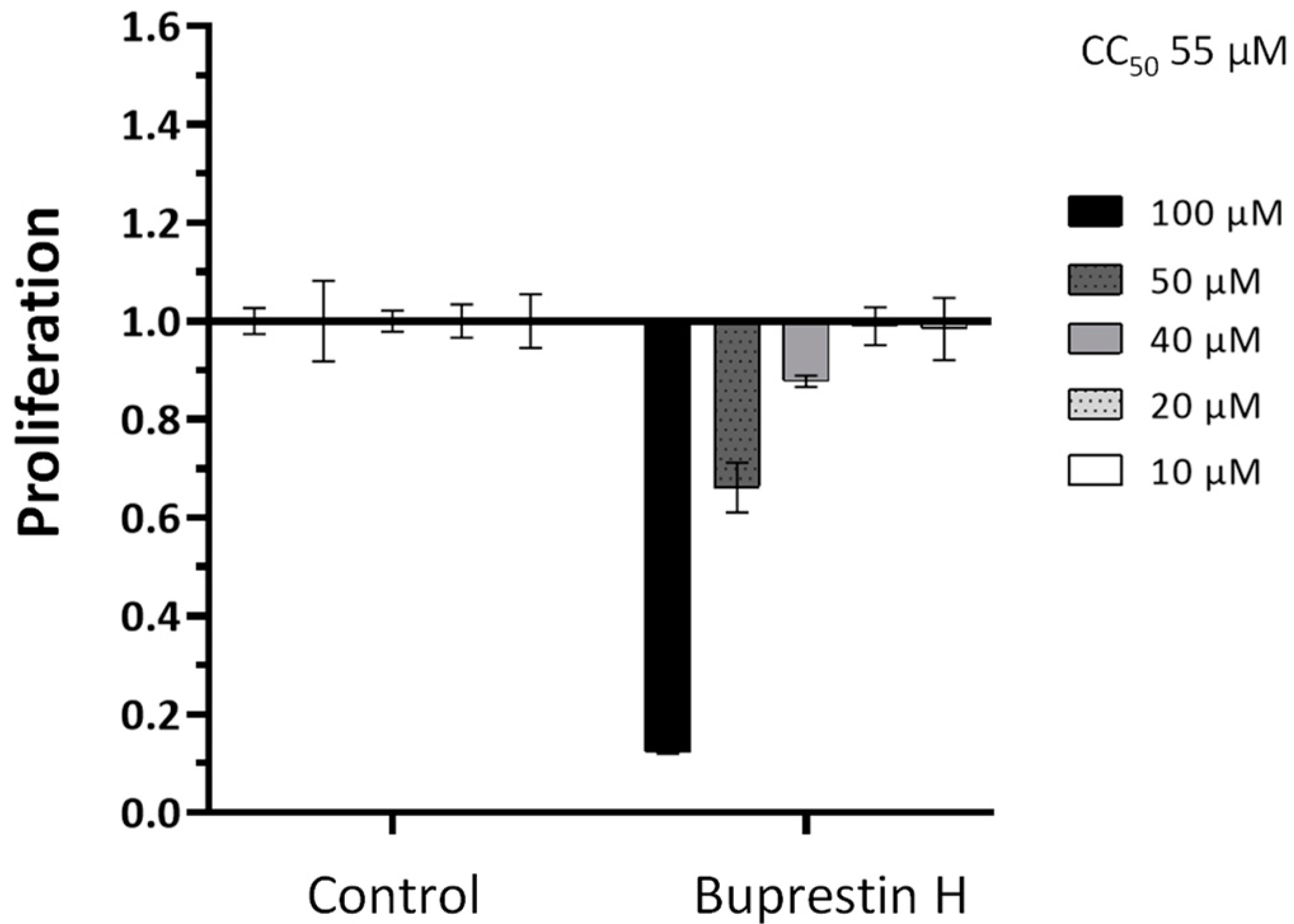
2.4.1. Activity against Adult Worms
2.4.2. Activity against Juvenile Worms and Cytotoxic Potential
3. Discussion
3.1. Advances Achieved in Virtual Screening of Insect Molecules
3.2. Predicted Molecular Interactions between Insect Molecules and SmTGR
3.3. Buprestin H as Antischistosomal Compound
3.4. Hurdles to Overcome for In Vitro Screens of Insect Molecules
4. Materials and Methods
4.1. Data Set Preparation
4.2. Docking Analyses
4.3. Visual Inspection
4.4. PLIP Analyses
4.5. Buprestin H
4.6. Maintenance of Schistosoma mansoni
4.7. Biological Evaluation on Schistosoma mansoni
4.8. Cytotoxic Measurements
4.9. Ethics Statement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Ndjonka, D.; Rapado, L.N.; Silber, A.M.; Liebau, E.; Wrenger, C. Natural products as a source for treating neglected parasitic diseases. Int. J. Mol. Sci. 2013, 14, 3395–3439. [Google Scholar] [CrossRef] [Green Version]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dossey, A.T. Insects and their chemical weaponry: New potential for drug discovery. Nat. Prod. Rep. 2010, 27, 1737–1757. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Chernysh, S.; Kim, S.I.; Bekker, G.; Pleskach, V.A.; Filatova, N.A.; Anikin, V.B.; Platonov, V.G.; Bulet, P. Antiviral and antitumor peptides from insects. Proc. Natl. Acad. Sci. USA 2002, 99, 12628–12632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibley, C.H.; Hunt, S.Y. Drug resistance in parasites: Can we stay ahead of the evolutionary curve? Trends Parasitol. 2003, 19, 532–537. [Google Scholar] [CrossRef]
- World Health Organization. Fact Sheet Schistosomiasis. Available online: https://www.who.int/news-room/fact-sheets/detail/schistosomiasis (accessed on 1 December 2021).
- King, C.H.; Dickman, K.; Tisch, D.J. Reassessment of the cost of chronic helmintic infection: A meta-analysis of disability-related outcomes in endemic schistosomiasis. Lancet 2005, 365, 1561–1569. [Google Scholar] [CrossRef]
- Fukushige, M.; Chase-Topping, M.; Woolhouse, M.E.J.; Mutapi, F. Efficacy of praziquantel has been maintained over four decades (from 1977 to 2018): A systematic review and meta-analysis of factors influence its efficacy. PLoS Neglected Trop. Dis. 2021, 15, e0009189. [Google Scholar] [CrossRef] [PubMed]
- Fallon, P.G.; Doenhoff, M.J. Drug-resistant schistosomiasis: Resistance to praziquantel and oxamniquine induced in Schistosoma mansoni in mice is drug specific. Am. J. Trop. Med. Hyg. 1994, 51, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.; Bennett, J.L.; Tao, L.-F.; Farghaly, A.; Bruce, J.; Metwally, A. Characterization of isolates of Schistosoma mansoni from Egyptian villagers that tolerate high doses of praziquantel. Am. J. Trop. Med. Hyg. 1996, 55, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Melman, S.D.; Steinauer, M.L.; Cunningham, C.; Kubatko, L.; Mwangi, I.N.; Wynn, N.B.; Mutuku, M.W.; Karanja, D.M.S.; Colley, D.; Black, C.L.; et al. Reduced susceptibility to praziquantel among naturally occurring Kenyan isolates of Schistosoma mansoni. PLoS Negl. Trop. Dis. 2009, 3, e504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LoVerde, P. Do antioxidants play a role in schistosome host–parasite interactions? Parasitol. Today 1998, 14, 284–289. [Google Scholar] [CrossRef]
- Alger, H.M.; Williams, D.L. The disulfide redox system of Schistosoma mansoni and the importance of a multifunctional enzyme, thioredoxin glutathione reductase. Mol. Biochem. Parasitol. 2002, 121, 129–139. [Google Scholar] [CrossRef]
- Kuntz, A.N.; Davioud-Charvet, E.; Sayed, A.A.; Califf, L.L.; Dessolin, J.; Arnér, E.S.; Williams, D.L. Thioredoxin glutathione reductase from Schistosoma mansoni: An essential parasite enzyme and a key drug target. PLoS Med. 2007, 4, e206. [Google Scholar]
- Salinas, G.; Selkirk, M.E.; Chalar, C.; Maizels, R.M.; Fernández, C. Linked thioredoxin-glutathione systems in platyhelminths. Trends Parasitol. 2004, 20, 340–346. [Google Scholar] [CrossRef]
- Simeonov, A.; Jadhav, A.; Sayed, A.A.; Wang, Y.; Nelson, M.E.; Thomas, C.J.; Inglese, J.; Williams, D.L.; Austin, C.P. Quantitative high-throughput screen identifies inhibitors of the Schistosoma mansoni redox cascade. PLoS Negl. Trop. Dis. 2008, 2, e127. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Ziniel, P.D.; He, P.-Q.; Kommer, V.P.; Crowther, G.J.; He, M.; Liu, Q.; Van Voorhis, W.C.; Williams, D.L.; Wang, M.-W. High-throughput screening against thioredoxin glutathione reductase identifies novel inhibitors with potential therapeutic value for schistosomiasis. Infect. Dis. Poverty 2015, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Braga, R.; Alves, V.; Silva, A.D.C.E.; Nascimento, M.N.; Silva, F.C.; Lião, L.M.; Andrade, C.H. Virtual screening strategies in medicinal chemistry: The state of the art and current challenges. Curr. Top. Med. Chem. 2014, 14, 1899–1912. [Google Scholar] [CrossRef]
- Lyu, H.; Petukhov, P.A.; Banta, P.R.; Jadhav, A.; Lea, W.A.; Cheng, Q.; Arnér, E.S.J.; Simeonov, A.; Thatcher, G.R.J.; Angelucci, F.; et al. Characterization of lead compounds targeting the selenoprotein thioredoxin glutathione reductase for treatment of schistosomiasis. ACS Infect. Dis. 2020, 6, 393–405. [Google Scholar] [CrossRef]
- Neves, B.J.; Dantas, R.F.; Senger, M.R.; Melo-Filho, C.C.; Valente, W.C.G.; de Almeida, A.C.M.; Rezende-Neto, J.M.; Lima, E.F.C.; Paveley, R.; Furnham, N.; et al. Discovery of new anti-schistosomal hits by integration of QSAR-based virtual screening and high content screening. J. Med. Chem. 2016, 59, 7075–7088. [Google Scholar] [CrossRef] [Green Version]
- Fata, F.; Silvestri, I.; Ardini, M.; Ippoliti, R.; Di Leandro, L.; Demitri, N.; Polentarutti, M.; Di Matteo, A.; Lyu, H.; Thatcher, G.R.; et al. Probing the surface of a parasite drug target thioredoxin glutathione reductase using small molecule fragments. ACS Infect. Dis. 2021, 7, 1932–1944. [Google Scholar] [CrossRef]
- Neves, B.J.; Andrade, C.H.; Cravo, P.V.L. Natural products as leads in schistosome drug discovery. Molecules 2015, 20, 1872–1903. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque, R.D.D.G.; Mahomoodally, M.F.; Lobine, D.; Suroowan, S.; Rengasamy, K.R. Botanical products in the treatment and control of schistosomiasis: Recent studies and distribution of active plant resources according to affected regions. Biology 2020, 9, 223. [Google Scholar] [CrossRef]
- Kellershohn, J.; Thomas, L.; Hahnel, S.R.; Grünweller, A.; Hartmann, R.K.; Hardt, M.; Vilcinskas, A.; Grevelding, C.G.; Haeberlein, S. Insects in anthelminthics research: Lady beetle-derived harmonine affects survival, reproduction and stem cell proliferation of Schistosoma mansoni. PLoS Negl. Trop. Dis. 2019, 13, e0007240. [Google Scholar] [CrossRef] [Green Version]
- Tonk, M.; Vilcinskas, A.; Grevelding, C.G.; Haeberlein, S. Anthelminthic activity of assassin bug venom against the blood fluke Schistosoma mansoni. Antibiotics 2020, 9, 664. [Google Scholar] [CrossRef] [PubMed]
- Dictionary of Natural Products 28. Available online: https://dnp.chemnetbase.com/faces/chemical/ChemicalSearch.xhtml (accessed on 25 November 2020).
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein-ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, I.; Lyu, H.; Fata, F.; Boumis, G.; Miele, A.E.; Ardini, M.; Ippoliti, R.; Bellelli, A.; Jadhav, A.; Lea, W.A.; et al. Fragment-based discovery of a regulatory site in thioredoxin gutathione reductase acting as “doorstop” for NADPH entry. ACS Chem. Biol. 2018, 13, 2190–2202. [Google Scholar] [CrossRef] [PubMed]
- SciFinder. Available online: https://www.cas.org/solutions/cas-scifinder-discovery-platform/cas-scifinder (accessed on 20 August 2021).
- PLIP Protein-Ligand Interaction Profiler. Available online: https://plip-tool.biotec.tu-dresden.de/plip-web/plip/index (accessed on 27 November 2021).
- Noda, N.; Kubota, S.; Miyata, Y.; Miyahara, K. Optically active N-acetyldopamine dimer of the crude drug “zentai”, the cast-off shell of the cicada, Cryptotympana sp. Chem. Pharm. Bull. 2000, 48, 1749–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.-N.; Tu, Z.-C.; Wang, X.-L.; Yan, Y.-M.; Fang, P.; Zuo, Z.-L.; Hou, B.; Yang, T.-H.; Cheng, Y.-X. Bioactive compounds from the insect Aspongopus chinensis. Bioorganic Med. Chem. Lett. 2014, 24, 5164–5169. [Google Scholar] [CrossRef] [PubMed]
- Tada, T.; Ohnishi, K.; Suzuki, K.; Tomita, H.; Okamori, M.; Katuzaki, H.; Komiya, T.; Imai, K. Potential cosmetic whitening agents from insect cuticle: Tyrosinase inhibitory activity of N-acetyldopamine dimers from exuviae of cicada, Cryptotympana tustulata FABR. J. Oleo Sci. 2002, 51, 355–358. [Google Scholar] [CrossRef]
- Yan, Y.-M.; Li, L.-J.; Qin, X.-C.; Lu, Q.; Tu, Z.-C.; Cheng, Y.-X. Compounds from the insect Blaps japanensis with COX-1 and COX-2 inhibitory activities. Bioorganic Med. Chem. Lett. 2015, 25, 2469–2472. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Cheng, L.; Xiao, S.; Hou, X. A New dopamine derivative from Cotidius chinensis. Rec. Nat. Prod. 2020, 14, 361–366. [Google Scholar] [CrossRef]
- Wang, S.F.; Braekman, J.C.; Daloze, D.; Pasteels, J.; Soetens, P.; Handjieva, N.V.; Kalushkov, P. Nα-quinaldyl-L-arginine·HCl, a new defensive alkaloid from Subcoccinella 24-punctata (Coleoptera, Coccinellidae). Cell. Mol. Life Sci. 1996, 52, 628–630. [Google Scholar] [CrossRef]
- Yan, Y.-M.; Dai, H.-Q.; Du, Y.; Schneider, B.; Guo, H.; Li, D.-P.; Zhang, L.-X.; Fu, H.; Dong, X.-P.; Cheng, Y.-X. Identification of blapsins A and B as potent small-molecule 14-3-3 inhibitors from the insect Blaps japanensis. Bioorganic Med. Chem. Lett. 2012, 22, 4179–4181. [Google Scholar] [CrossRef] [PubMed]
- Ryczek, S.; Dettner, K.; Unverzagt, C. Synthesis of buprestins D, E, F, G and H; structural confirmation and biological testing of acyl glucoses from jewel beetles (Coleoptera: Buprestidae). Bioorganic Med. Chem. 2009, 17, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Saidemberg, D.M.; Da Silva-Filho, L.C.; Tognoli, L.M.M.C.; Tormena, C.F.; Palma, M.S. Polybioside, a neuroactive compound from the venom of the social wasp Polybia paulista. J. Nat. Prod. 2010, 73, 527–531. [Google Scholar] [CrossRef]
- Barbier, M. Synthesis of Z-Marginalin and identification of the natural product as the E isomer. Eur. J. Org. Chem. 1987, 6, 545–546. [Google Scholar] [CrossRef]
- Biodiversity Map, National Biodiversity Information Network. 2021. Available online: https://baza.biomap.pl (accessed on 1 December 2021).
- Liu, J.; Dyer, D.H.; Cheng, J.; Wang, J.; Wang, S.; Yang, Z.; Wang, X.; Hu, W. Aldose reductase from Schistosoma japonicum: Crystallization and structure-based inhibitor screening for discovering antischistosomal lead compounds. Parasites Vectors 2013, 6, 162. [Google Scholar] [CrossRef] [Green Version]
- Kannan, S.; Melesina, J.; Hauser, A.-T.; Chakrabarti, A.; Heimburg, T.; Schmidtkunz, K.; Walter, A.; Marek, M.; Pierce, R.J.; Romier, C.; et al. Discovery of inhibitors of Schistosoma mansoni HDAC8 by combining homology modeling, virtual screening, and in vitro validation. J. Chem. Inf. Model. 2014, 54, 3005–3019. [Google Scholar] [CrossRef]
- Padalino, G.; Chalmers, I.W.; Brancale, A.; Hoffmann, K.F. Identification of 6-(piperazin-1-yl)-1,3,5-triazine as a chemical scaffold with broad anti-schistosomal activities. Wellcome Open Res. 2020, 5, 169. [Google Scholar] [CrossRef]
- Saccoccia, F.; Brindisi, M.; Gimmelli, R.; Relitti, N.; Guidi, A.; Saraswati, A.P.; Cavella, C.; Brogi, S.; Chemi, G.; Butini, S.; et al. Screening and phenotypical characterization of Schistosoma mansoni histone deacetylase 8 (SmHDAC8) inhibitors as multistage antischistosomal agents. ACS Infect. Dis. 2020, 6, 100–113. [Google Scholar] [CrossRef]
- Melo-Filho, C.C.; Dantas, R.F.; Braga, R.C.; Neves, B.J.; Senger, M.R.; Valente, W.C.; Rezende-Neto, J.M.; Chaves, W.T.; Muratov, E.N.; Paveley, R.A.; et al. QSAR-driven discovery of novel chemical scaffolds active against Schistosoma mansoni. J. Chem. Inf. Model 2016, 56, 1357–1372. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.; Shukla, H.; Kalita, P.; Tripathi, T. Structural insights into natural compounds as inhibitors of Fasciola gigantica thioredoxin glutathione reductase. J. Cell. Biochem. 2018, 119, 3067–3080. [Google Scholar] [CrossRef]
- Hilder, T.A.; Chung, S.-H. Conduction and block of inward rectifier K+channels: Predicted structure of a potent blocker of Kir2. Biochemistry 2013, 52, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, N.; Wang, W.; Wang, J.; Zhu, Z.; Li, X. Molecular mechanisms of novel peptides from silkworm pupae that inhibit α-glucosidase. Peptides 2016, 76, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Ehrman, T.M.; Barlow, D.; Hylands, P.J. Phytochemical informatics of traditional Chinese medicine and therapeutic relevance. J. Chem. Inf. Model. 2007, 47, 2316–2334. [Google Scholar] [CrossRef] [PubMed]
- Sterling, T.; Irwin, J.J. ZINC 15--ligand discovery for everyone. J. Chem. Inf. Model 2015, 55, 2324–2337. [Google Scholar] [CrossRef]
- Sorokina, M.; Steinbeck, C. Review on natural products databases: Where to find data in 2020. J. Cheminform. 2020, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Sander, T.; Freyss, J.; von Korff, M.; Rufener, C. DataWarrior: An open-source program for chemistry aware data visualization and analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef]
- Lucas, A.J.; Sproston, J.L.; Barton, P.; Riley, R.J. Estimating human ADME properties, pharmacokinetic parameters and likely clinical dose in drug discovery. Expert Opin. Drug Discov. 2019, 14, 1313–1327. [Google Scholar] [CrossRef]
- Meyer, E.A.; Castellano, R.K.; Diederich, F. Interactions with aromatic rings in chemical and biological recognition. Angew. Chem. Int. Ed. 2003, 42, 1210–1250. [Google Scholar] [CrossRef] [PubMed]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A web server for target prediction of bioactive small molecules. Nucleic Acids Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef]
- Moore, B.P.; Brown, W.V. The buprestins: Bitter principles of jewel beetles (coleoptera: Buprestidae). Aust. J. Entomol. 1985, 24, 81–85. [Google Scholar] [CrossRef]
- Xiao, S.-H. Mefloquine, a new type of compound against schistosomes and other helminthes in experimental studies. Parasitol. Res. 2013, 112, 3723–3740. [Google Scholar] [CrossRef]
- Wong, W.; Bai, X.-C.; Sleebs, B.E.; Triglia, T.; Brown, A.; Thompson, J.K.; Jackson, K.E.; Hanssen, E.; Marapana, D.S.; Fernandez, I.S.; et al. Mefloquine targets the Plasmodium falciparum 80S ribosome to inhibit protein synthesis. Nat. Microbiol. 2017, 2, 17031. [Google Scholar] [CrossRef]
- Wang, J.; Paz, C.; Padalino, G.; Coghlan, A.; Lu, Z.; Gradinaru, I.; Collins, J.N.R.; Berriman, M.; Hoffmann, K.F.; Collins, J.J. Large-scale RNAi screening uncovers therapeutic targets in the parasite Schistosoma mansoni. Science 2020, 369, 1649–1653. [Google Scholar] [CrossRef] [PubMed]
- Schütte, L.D.; Baumeister, S.; Weis, B.; Hudemann, C.; Hanschmann, E.-M.; Lillig, C.H. Identification of potential protein dithiol-disulfide substrates of mammalian Grx2. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 4999–5005. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [Green Version]
- Bissantz, C.; Kuhn, B.; Stahl, M. A medicinal chemist’s guide to molecular interactions. J. Med. Chem. 2010, 53, 5061–5084. [Google Scholar] [CrossRef]
- Gönnert, R. Schistosomiasis-Studien. II. Über die Eibildung bei Schistosoma mansoni und das Schicksal der Eier im Wirtsorganismus. Z Tropenmed. Parasitol. 1955, 6, 33–52. [Google Scholar] [PubMed]
- Grevelding, C.G. The female-specific W1 sequence of the Puerto Rican strain of Schistosoma mansoni occurs in both genders of a Liberian strain. Mol. Biochem. Parasitol. 1995, 71, 269–272. [Google Scholar] [CrossRef]
- Dettman, C.D.; Higgins-Opitz, S.B.; Saikoolal, A. Enhanced efficacy of the paddling method for schistosome infection of rodents by a four-step pre-soaking procedure. Parasitol. Res. 1989, 76, 183–184. [Google Scholar] [CrossRef] [PubMed]
- Milligan, J.N.; Jolly, E.R. Cercarial transformation and in vitro cultivation of Schistosoma mansoni schistosomules. J. Vis. Exp. 2011, 54, e3191. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No., Name | Structure | Origin | Function/Activity + |
|---|---|---|---|
| 1, 2, 3 | 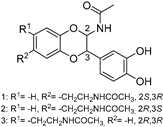 | 1: Cicada—Cryptotympana tustulata FABR. (Hemiptera), Stink bug—Cotidius chinensis/Aspongopus chinensis (Hemiptera), Stink beetle—Blaps japanensis, Dung beetle—Catharsius molossus (Coleoptera) [33,34,35,36,37] 2/3: Cicada—Cryptotympana tustulata FABR. (Hemiptera) [35] # | Crude drug “zentai”, COX-2 inhibitor, whitening agent |
| 4 |  | Ladybird—Subcoccinella vigintiquatuorpunctata (Coleoptera) [38] | Defensive alkaloid (ant deterrent) |
| 5 (R), 6 (S) Blapsin B |  | Stink beetle—Blaps japanensis (Coleoptera) [39] | Inhibitor of 14-3-3 protein-protein interactions, antioxidant |
| 7 Buprestin H | 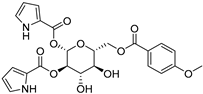 | Jewel beetle—Chalcophora mariana (Coleoptera) [40] | Unknown |
| 8 Blapsin A | 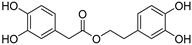 | Stink beetle—Blaps japanensis (Coleoptera) [39] | Inhibitor of 14-3-3 protein-protein interactions |
| 9 Polybioside | 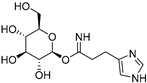 | Social wasp—Polybia paulista (Hymenoptera) [41] | Component in neuroactive venom |
| 10 Marginalin | 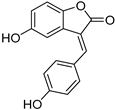 | Water beetle, Dytiscus marginalis (Coleoptera) [42] | Pygidial bladders coloring substance |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallinger, T.L.; Aboagye, S.Y.; Obermann, W.; Weiss, M.; Grünweller, A.; Unverzagt, C.; Williams, D.L.; Schlitzer, M.; Haeberlein, S. First In Silico Screening of Insect Molecules for Identification of Novel Anti-Parasitic Compounds. Pharmaceuticals 2022, 15, 119. https://doi.org/10.3390/ph15020119
Gallinger TL, Aboagye SY, Obermann W, Weiss M, Grünweller A, Unverzagt C, Williams DL, Schlitzer M, Haeberlein S. First In Silico Screening of Insect Molecules for Identification of Novel Anti-Parasitic Compounds. Pharmaceuticals. 2022; 15(2):119. https://doi.org/10.3390/ph15020119
Chicago/Turabian StyleGallinger, Tom L., Samuel Y. Aboagye, Wiebke Obermann, Michael Weiss, Arnold Grünweller, Carlo Unverzagt, David L. Williams, Martin Schlitzer, and Simone Haeberlein. 2022. "First In Silico Screening of Insect Molecules for Identification of Novel Anti-Parasitic Compounds" Pharmaceuticals 15, no. 2: 119. https://doi.org/10.3390/ph15020119
APA StyleGallinger, T. L., Aboagye, S. Y., Obermann, W., Weiss, M., Grünweller, A., Unverzagt, C., Williams, D. L., Schlitzer, M., & Haeberlein, S. (2022). First In Silico Screening of Insect Molecules for Identification of Novel Anti-Parasitic Compounds. Pharmaceuticals, 15(2), 119. https://doi.org/10.3390/ph15020119