

Topiramate Reprofiling for the Attenuation of Cadmium-Induced Testicular Impairment in Rats: Role of NLRP3 Inflammasome and AMPK/mTOR-Linked Autophagy
 , , , and
, , , and
Abstract
1. Introduction
2. Results
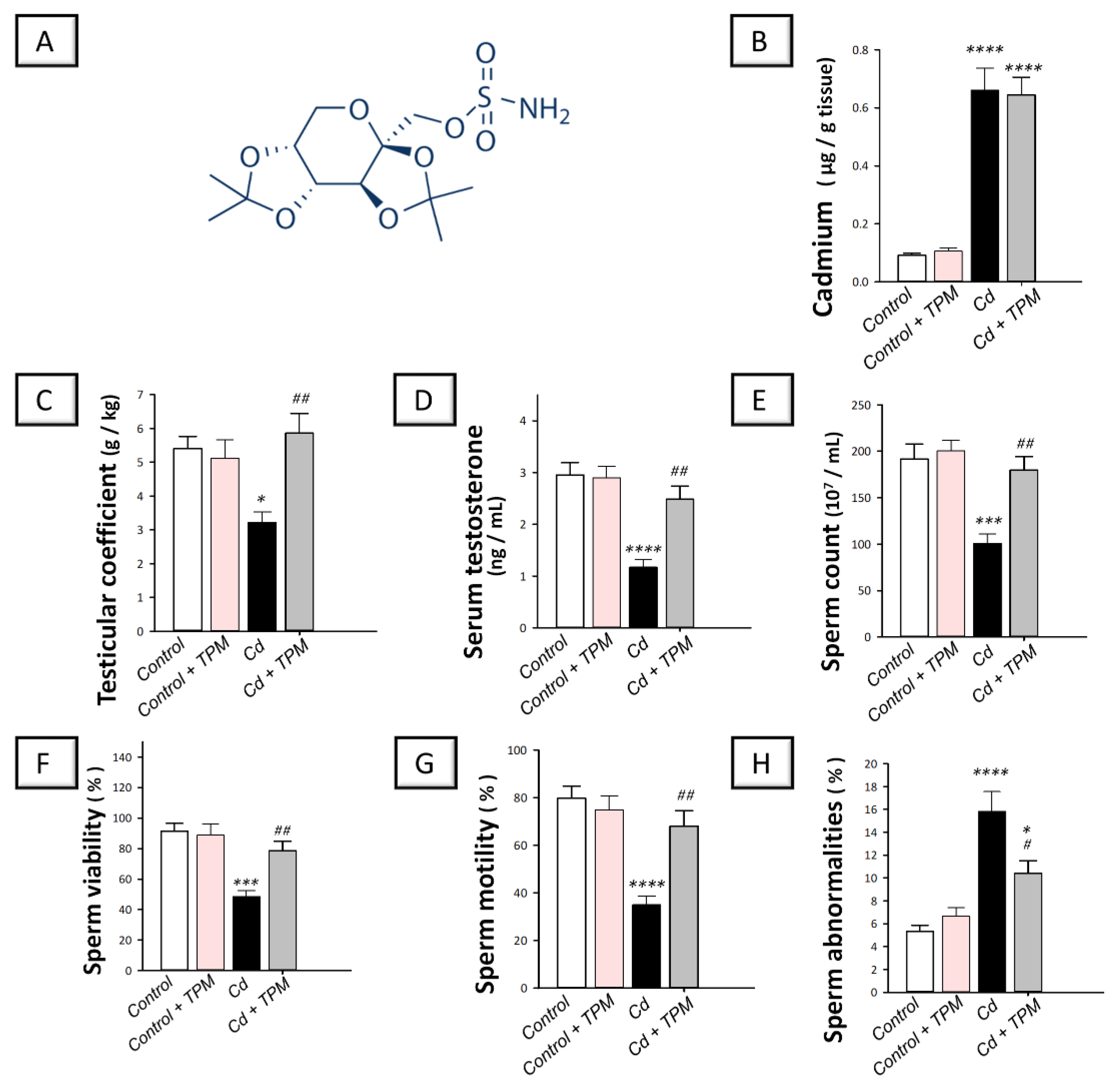
2.1. Effect of Topiramate on Testicular Dysfunction and Impaired Spermatogenesis in Cadmium-Induced Testicular Injury in Rats
2.2. Effect of Topiramate on the Histopathological Aberrations in Cadmium-Induced Testicular Injury in Rats
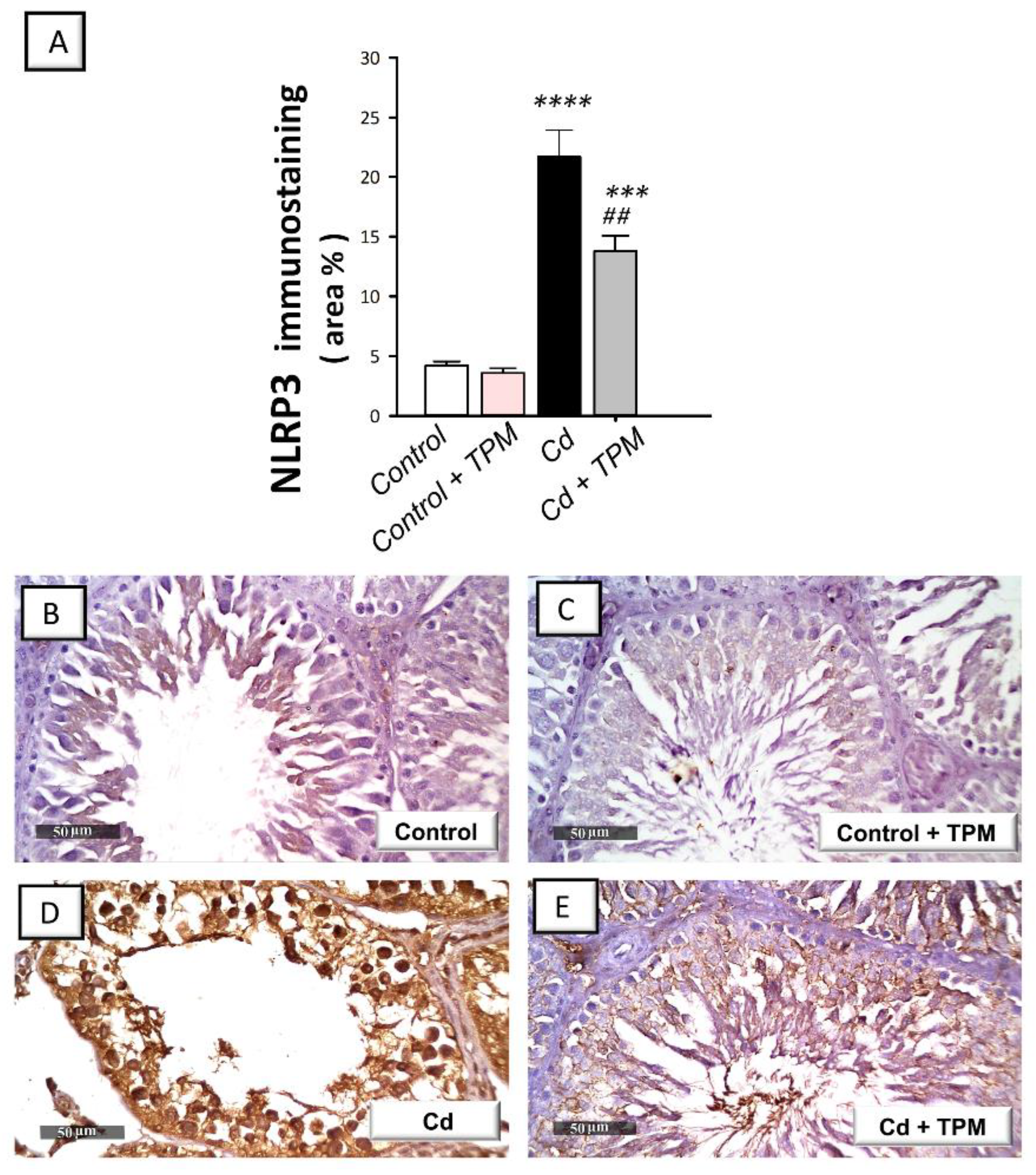
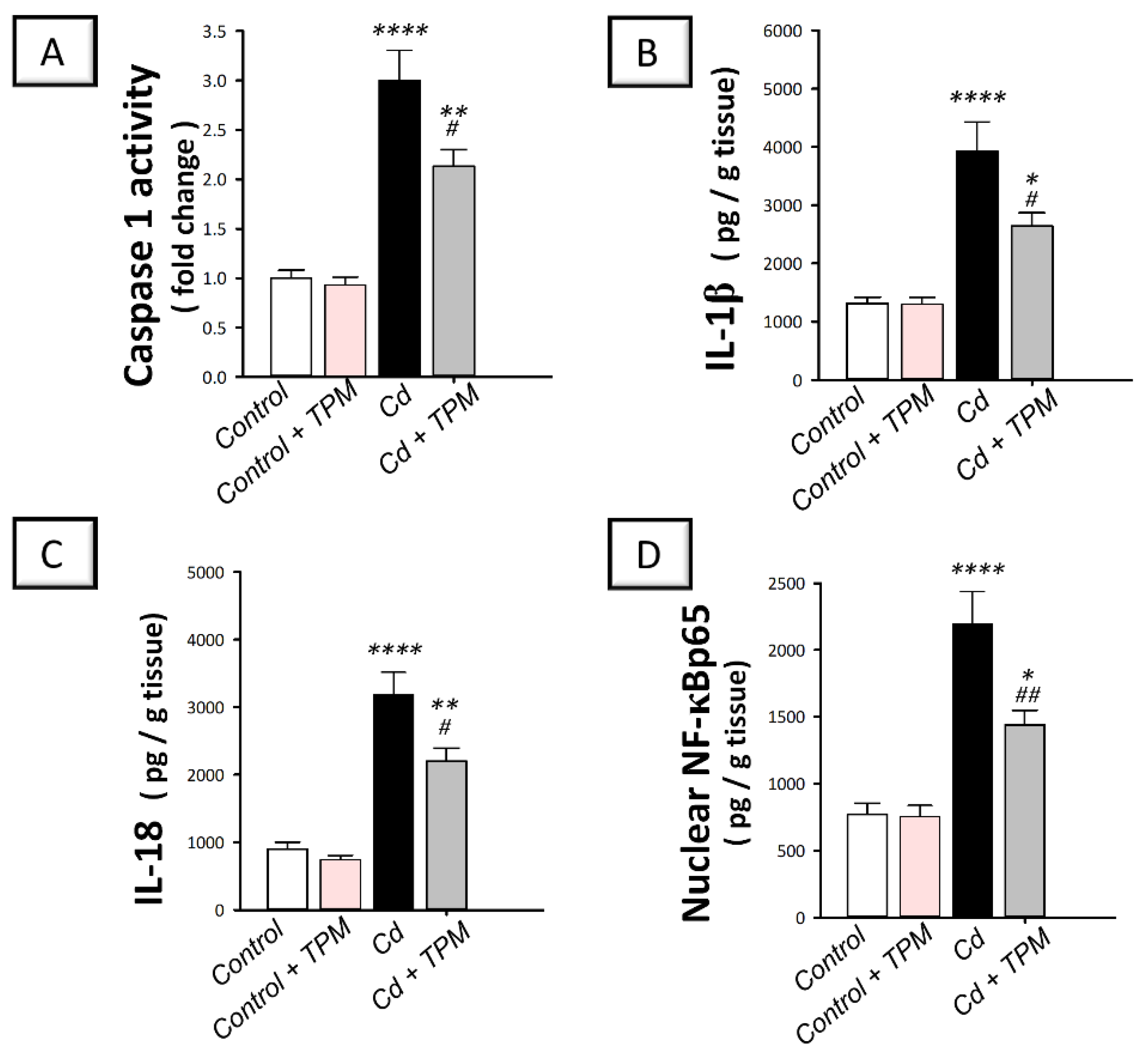
2.3. Effect of Topiramate on Testicular NLRP3 Inflammasome Activation in Cadmium-Induced Testicular Injury in Rats
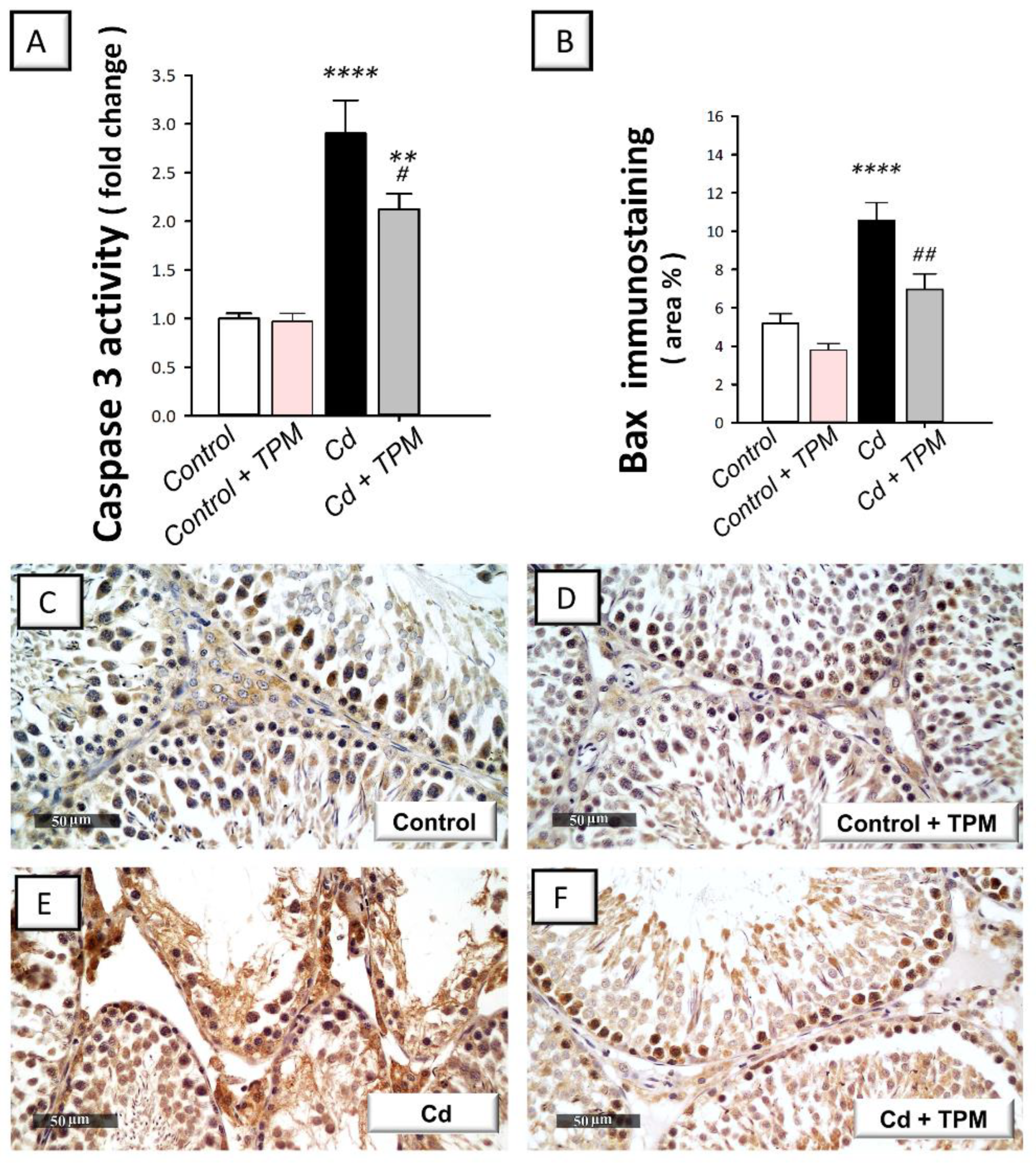
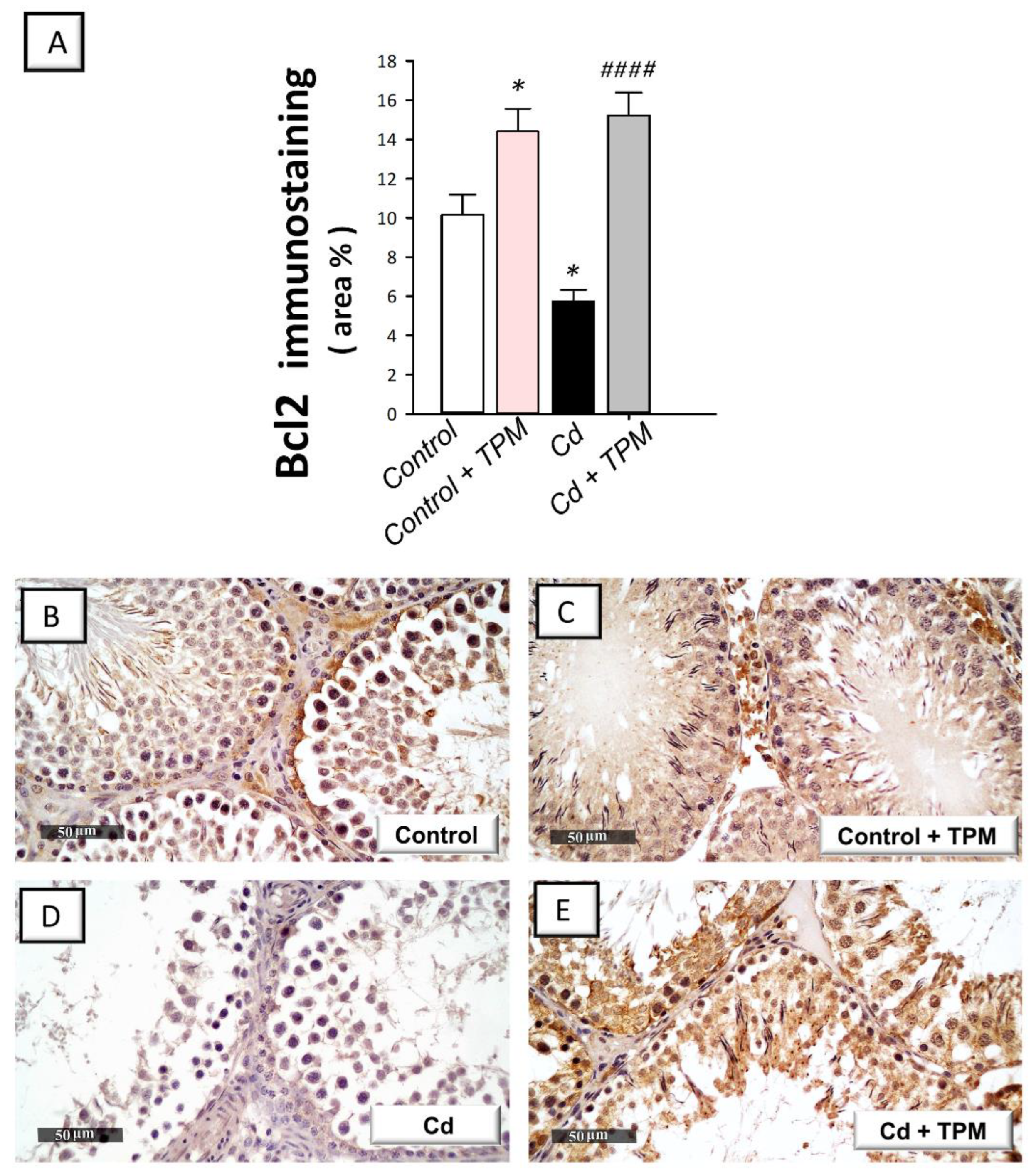
2.4. Effect of Topiramate on Testicular Apoptosis Markers in Cadmium-Induced Testicular Injury in Rats
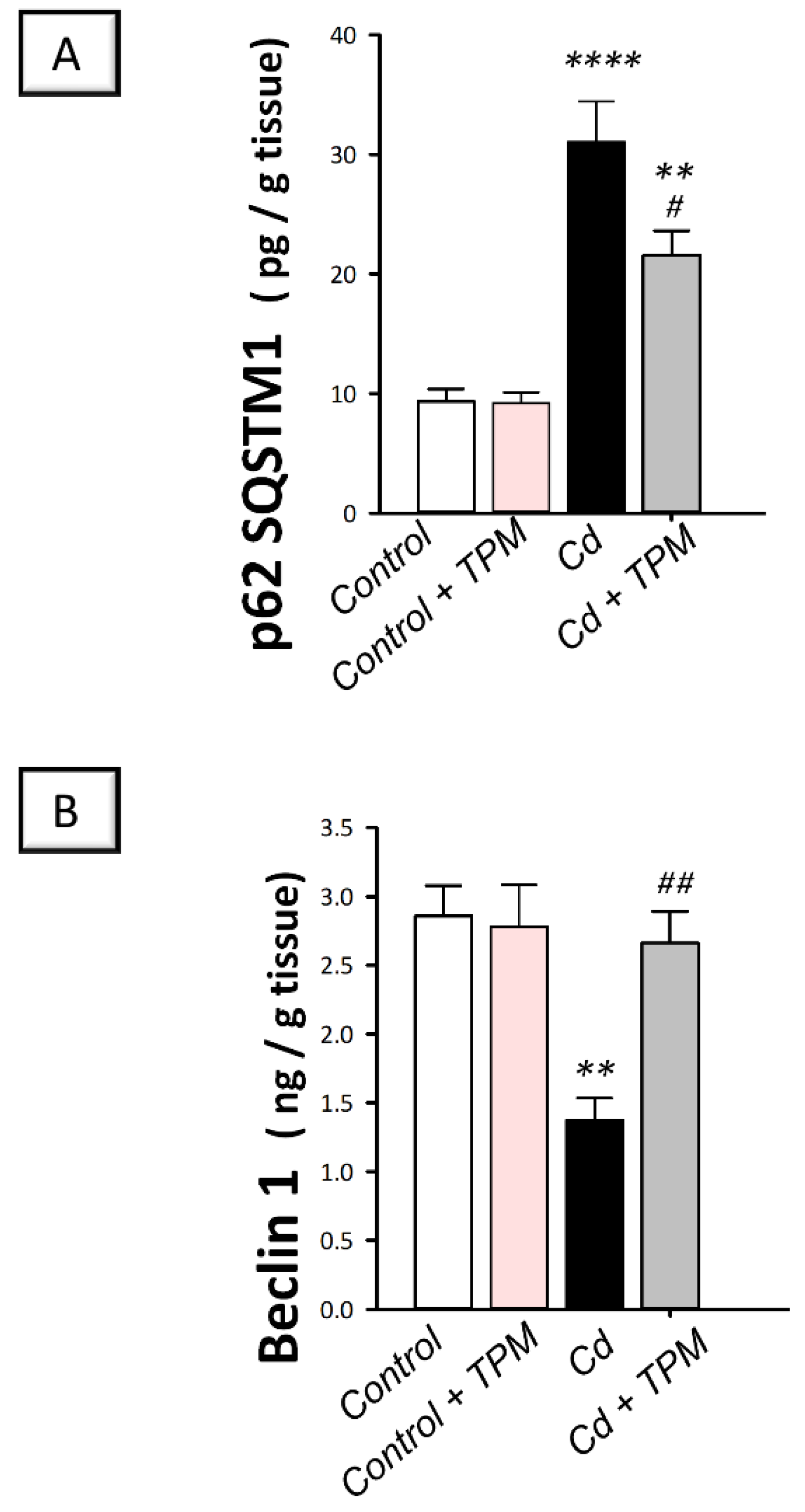
2.5. Effect of Topiramate on Defective Autophagy in Cadmium-Induced Testicular Injury in Rats
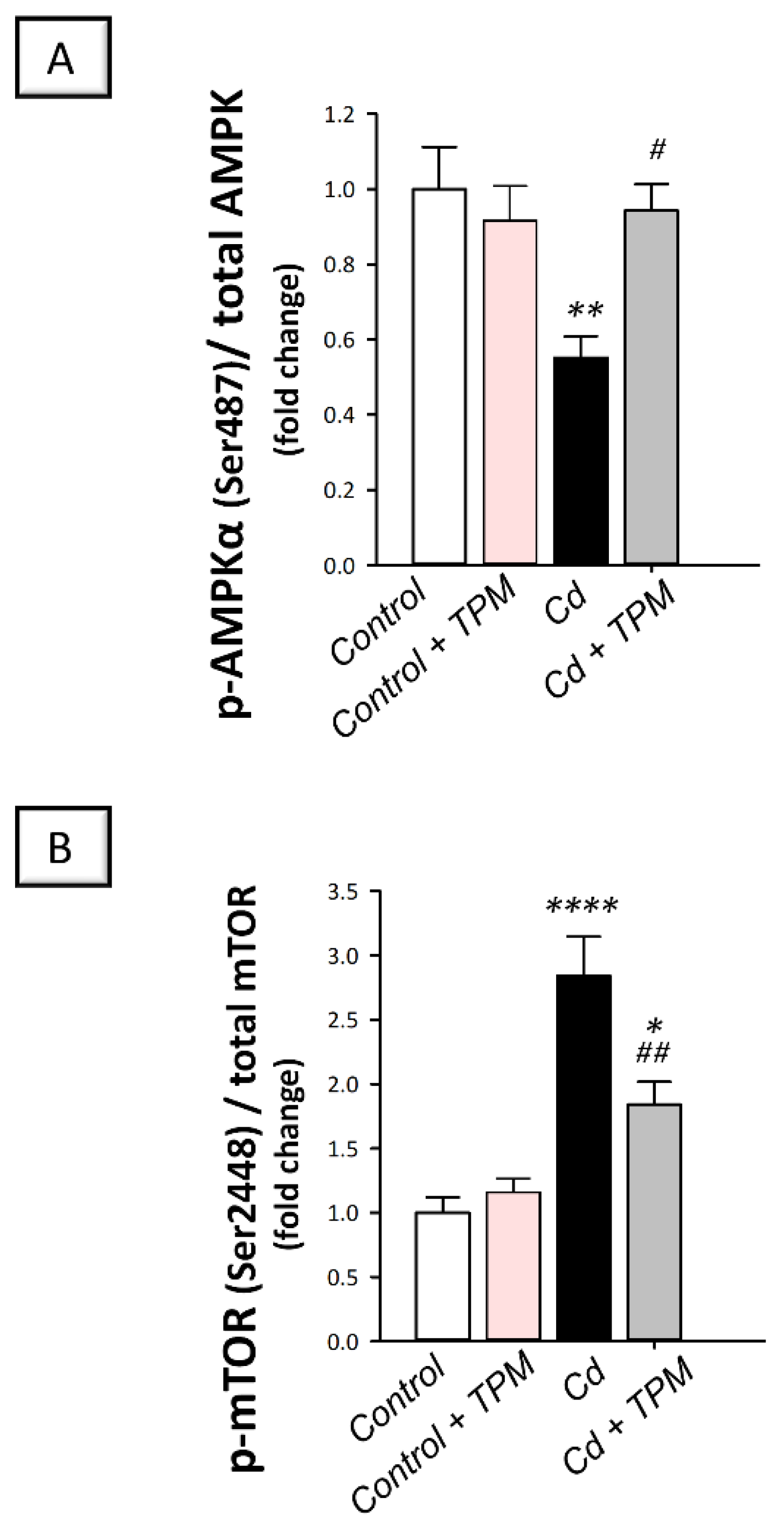
2.6. Effect of Topiramate on AMPK/mTOR Pathway in Cadmium-Induced Testicular Damage in Rats
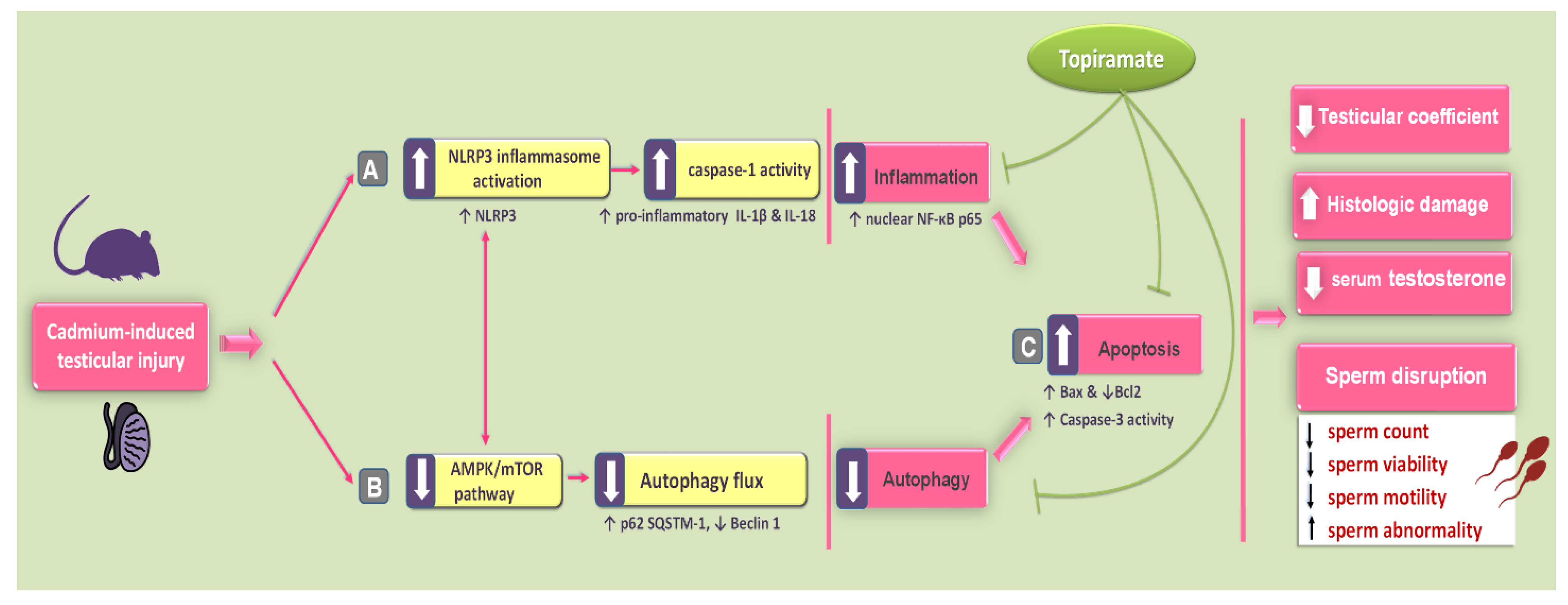
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Experimental Design and Protocol
4.4. Evaluation of Semen Parameters
4.5. Serum Testosterone Determination
4.6. Estimation of Inflammatory Markers
4.7. Determination of the Activity of Caspase-1 and Caspase-3
4.8. Estimation of Autophagy Markers
4.9. Estimation of Cadmium Metal Content in Testis
4.10. Histopathology
4.11. Immunohistochemistry
4.12. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Y.; He, X.; Qi, K.; Wang, T.; Qi, Y.; Cui, L.; Wang, F.; Song, M. Effects of environmental contaminants on fertility and reproductive health. J. Environ. Sci. 2019, 77, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Ola-Mudathir, K.F.; Suru, S.M.; Fafunso, M.A.; Obioha, U.E.; Faremi, T.Y. Protective roles of onion and garlic extracts on cadmium-induced changes in sperm characteristics and testicular oxidative damage in rats. Food Chem. Toxicol. 2008, 46, 3604–3611. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, C.; Galdiero, M.; Pivonello, C.; Salzano, C.; Gianfrilli, D.; Piscitelli, P.; Lenzi, A.; Colao, A.; Pivonello, R. The environment and male reproduction: The effect of cadmium exposure on reproductive function and its implication in fertility. Reprod. Toxicol. 2017, 73, 105–127. [Google Scholar] [CrossRef] [PubMed]
- Habib, R.; Wahdan, S.A.; Gad, A.M.; Azab, S.S. Infliximab abrogates cadmium-induced testicular damage and spermiotoxicity via enhancement of steroidogenesis and suppression of inflammation and apoptosis mediators. Ecotoxicol. Environ. Saf. 2019, 182, 109398. [Google Scholar] [CrossRef]
- Mead, M.N. Cadmium confusion: Do consumers need protection? Environ. Health Perspect. 2010, 118, A528–A534. [Google Scholar] [CrossRef]
- Minutoli, L.; Puzzolo, D.; Rinaldi, M.; Irrera, N.; Marini, H.; Arcoraci, V.; Bitto, A.; Crea, G.; Pisani, A.; Squadrito, F.; et al. ROS-Mediated NLRP3 Inflammasome Activation in Brain, Heart, Kidney, and Testis Ischemia/Reperfusion Injury. Oxidative Med. Cell. Longev. 2016, 2016, 2183026. [Google Scholar] [CrossRef]
- Hong, Y.; Zhou, Y.; Shen, L.; Wei, Y.; Long, C.; Fu, Y.; Wu, H.; Wang, J.; Wu, Y.; Wu, S.; et al. Exposure to DEHP induces testis toxicity and injury through the ROS/mTOR/NLRP3 signaling pathway in immature rats. Ecotoxicol. Environ. Saf. 2021, 227, 112889. [Google Scholar] [CrossRef]
- Zhou, G.X.; Zhu, H.L.; Shi, X.T.; Nan, Y.; Liu, W.B.; Dai, L.M.; Xiong, Y.W.; Yi, S.J.; Cao, X.L.; Xu, D.X.; et al. Autophagy in Sertoli cell protects against environmental cadmium-induced germ cell apoptosis in mouse testes. Environ. Pollut. 2021, 270, 116241. [Google Scholar] [CrossRef]
- Yu, W.; Xu, Z.; Gao, Q.; Xu, Y.; Wang, B.; Dai, Y. Protective role of wogonin against cadmium induced testicular toxicity: Involvement of antioxidant, anti-inflammatory and anti-apoptotic pathways. Life Sci. 2020, 258, 118192. [Google Scholar] [CrossRef]
- Bao, R.-K.; Zheng, S.-F.; Wang, X.-Y. Selenium protects against cadmium-induced kidney apoptosis in chickens by activating the PI3K/AKT/Bcl-2 signaling pathway. Environ. Sci. Pollut. Res. Int. 2017, 24, 20342–20353. [Google Scholar] [CrossRef]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Song, W.; Xu, D.; Chen, X.; Li, X.; Zhao, Y. Autophagy Induced by ROS Aggravates Testis Oxidative Damage in Diabetes via Breaking the Feedforward Loop Linking p62 and Nrf2. Oxidative Med. Cell. Longev. 2020, 2020, 7156579. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, H.; Wang, K.; Yang, Z.; Liu, Z. Protective effect of quercetin on rat testes against cadmium toxicity by alleviating oxidative stress and autophagy. Environ. Sci. Pollut. Res. 2020, 27, 25278–25286. [Google Scholar] [CrossRef]
- Chen, X.; Li, C.; Chen, Y.; Ni, C.; Chen, X.; Zhang, L.; Xu, X.; Chen, M.; Ma, X.; Zhan, H.; et al. Aflatoxin B1 impairs leydig cells through inhibiting AMPK/mTOR-mediated autophagy flux pathway. Chemosphere 2019, 233, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-Y.; Zhu, X.-L.; Zhao, B.-X.; Shi, L.; Wang, W.; Hu, W.; Qin, S.-L.; Chen, B.-H.; Zhou, P.-H.; Qiu, B.; et al. Adrenomedullin alleviates the pyroptosis of Leydig cells by promoting autophagy via the ROS–AMPK–mTOR axis. Cell Death Dis. 2019, 10, 489. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Li, Z.X.; Zhao, Y.T.; Liu, M.; Wang, Y.; Lian, G.C.; Zhao, Q.; Wang, H.L. PCPA protects against monocrotaline-induced pulmonary arterial remodeling in rats: Potential roles of connective tissue growth factor. Oncotarget 2017, 8, 111642–111655. [Google Scholar] [CrossRef]
- Liu, F.; Wang, X.-Y.; Zhou, X.-P.; Liu, Z.-P.; Song, X.-B.; Wang, Z.-Y.; Wang, L. Cadmium disrupts autophagic flux by inhibiting cytosolic Ca2+-dependent autophagosome-lysosome fusion in primary rat proximal tubular cells. Toxicology 2017, 383, 13–23. [Google Scholar] [CrossRef]
- Messner, B.; Ploner, C.; Laufer, G.; Bernhard, D. Cadmium activates a programmed, lysosomal membrane permeabilization-dependent necrosis pathway. Toxicol. Lett. 2012, 212, 268–275. [Google Scholar] [CrossRef]
- Marino, G.; Niso-Santano, M.; Baehrecke, E.H.; Kroemer, G. Self-consumption: The interplay of autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2014, 15, 81–94. [Google Scholar] [CrossRef]
- Arab, H.H.; Gad, A.M.; Reda, E.; Yahia, R.; Eid, A.H. Activation of autophagy by sitagliptin attenuates cadmium-induced testicular impairment in rats: Targeting AMPK/mTOR and Nrf2/HO-1 pathways. Life Sci. 2021, 269, 119031. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, C.-Q.; Zeng, L.; Cheng, L.; Ma, L.; Zhang, M.; Zhang, Y.-Z. Melatonin regulates the cross-talk between autophagy and apoptosis by SIRT3 in testicular Leydig cells. Biochem. Biophys. Res. Commun. 2021, 555, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Manzini, S.; Busnelli, M.; Parolini, C.; Minoli, L.; Ossoli, A.; Brambilla, E.; Simonelli, S.; Lekka, E.; Persidis, A.; Scanziani, E.; et al. Topiramate protects apoE-deficient mice from kidney damage without affecting plasma lipids. Pharmacol. Res. 2019, 141, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Chrościńska-Krawczyk, M.; Wałek, M.; Tylus, B.; Czuczwar, S.J. Selected classical and novel antiepileptic drugs–mechanisms of action, neuroprotection, and effectiveness in epileptic and non-epileptic conditions. J. Epileptol. 2014, 22, 37–50. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, S.; Sheng, Y.; Peng, X.; Liu, H.; Jin, N.; Cai, J.; Shu, Y.; Li, T.; Li, P.; et al. Topiramate modulates post-infarction inflammation primarily by targeting monocytes or macrophages. Cardiovasc. Res. 2017, 113, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Y.; Xiao, J.; Zhang, H.; Yang, C.; Wei, Z.; Chen, W.; Du, X.; Liu, J. Modulating Neuro-Immune-Induced Macrophage Polarization with Topiramate Attenuates Experimental Abdominal Aortic Aneurysm. Front. Pharmacol. 2020, 11, 565461. [Google Scholar] [CrossRef]
- Jara, C.P.; Bobbo, V.C.D.; Carraro, R.S.; de Araujo, T.M.F.; Lima, M.H.M.; Velloso, L.A.; Araujo, E.P. Effects of topical topiramate in wound healing in mice. Arch. Dermatol. Res. 2018, 310, 363–373. [Google Scholar] [CrossRef]
- Shalaby, H.N.; El-Tanbouly, D.M.; Zaki, H.F. Topiramate mitigates 3-nitropropionic acid-induced striatal neurotoxicity via modulation of AMPA receptors. Food Chem. Toxicol. 2018, 118, 227–234. [Google Scholar] [CrossRef]
- Cheng, X.L.; Li, M.K. Effect of topiramate on apoptosis-related protein expression of hippocampus in model rats with Alzheimers disease. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 761–768. [Google Scholar]
- Motaghinejad, M.; Motevalian, M.; Abdollahi, M.; Heidari, M.; Madjd, Z. Topiramate Confers Neuroprotection against Methylphenidate-Induced Neurodegeneration in Dentate Gyrus and CA1 Regions of Hippocampus via CREB/BDNF Pathway in Rats. Neurotox. Res. 2017, 31, 373–399. [Google Scholar] [CrossRef]
- Kadasah, S.; Al Eid, A.S.; Alawad, S.S.; Al Shahrani, A.S.; Alruwaihi, A.S.; Elfaki, I.; Arshaduddin, M. Gastro protecting influence of Topiramate in ethanol produced gastric ulcers in rats. Toxicol. Rep. 2021, 8, 1031–1039. [Google Scholar] [CrossRef]
- Arab, H.H.; Gad, A.M.; Fikry, E.M.; Eid, A.H. Ellagic acid attenuates testicular disruption in rheumatoid arthritis via targeting inflammatory signals, oxidative perturbations and apoptosis. Life Sci. 2019, 239, 117012. [Google Scholar] [CrossRef]
- Pergel, A.; Demiral, G.; Tumkaya, L.; Mercantepe, T.; Ozdemir, A.; Kalcan, S.; Colakoglu, M.K.; Yilmaz, A.; Bedir, R.; Karakaya, A. The protective effects of topiramate on intestinal injury induced with infrarenal aortic occlusion via oxidative stress and apoptosis. Clin. Exp. Hypertens. 2021, 43, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Xie, M.; Li, Y.; Gong, X.; Li, J. Topiramate Reverses Physiological and Behavioral Alterations by Postoperative Cognitive Dysfunction in Rat Model through Inhibiting TNF Signaling Pathway. Neuromol. Med. 2020, 22, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, X.-F.; Li, Y.-M.; Chen, N.; Fan, Y.; Huang, W.-K.; Hu, S.-F.; Rao, M.; Zhang, Y.-Z.; Su, P. Cross-talk between autophagy and apoptosis regulates testicular injury/recovery induced by cadmium via PI3K with mTOR-independent pathway. Cell Death Dis 2020, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Ni, B.; Yang, Y.-D.; Tang, Z.-W.; Gao, Z.-Q.; Feng, L.; Liao, W.-G.; Gao, Y.-Q. Elevation of autophagy rescues spermatogenesis by inhibiting apoptosis of mouse spermatocytes. Reprod. Nutr. Dev. 2018, 156, 545–558. [Google Scholar] [CrossRef]
- Venditti, M.; Rhouma, M.B.; Romano, M.Z.; Messaoudi, I.; Reiter, R.J.; Minucci, S. Evidence of melatonin ameliorative effects on the blood-testis barrier and sperm quality alterations induced by cadmium in the rat testis. Ecotoxicol. Environ. Saf. 2021, 226, 112878. [Google Scholar] [CrossRef]
- Wirawan, E.; Walle, L.V.; Kersse, K.; Cornelis, S.; Claerhout, S.; Vanoverberghe, I.; Roelandt, R.; de Rycke, R.; Verspurten, J.; Declercq, W. Caspase-mediated cleavage of Beclin-1 inactivates Beclin-1-induced autophagy and enhances apoptosis by promoting the release of proapoptotic factors from mitochondria. Cell Death Dis. 2010, 1, e18. [Google Scholar] [CrossRef]
- Shi, J.Q.; Wang, B.R.; Tian, Y.Y.; Xu, J.; Gao, L.; Zhao, S.L.; Jiang, T.; Xie, H.G.; Zhang, Y.D. Antiepileptics topiramate and levetiracetam alleviate behavioral deficits and reduce neuropathology in APPswe/PS1dE9 transgenic mice. CNS Neurosci. Ther. 2013, 19, 871–881. [Google Scholar] [CrossRef]
- Bassi, G.; Sidhu, S.K.; Mishra, S. The Expanding Role of Mitochondria, Autophagy and Lipophagy in Steroidogenesis. Cells 2021, 10, 1851. [Google Scholar] [CrossRef]
- Texada, M.J.; Malita, A.; Christensen, C.F.; Dall, K.B.; Faergeman, N.J.; Nagy, S.; Halberg, K.A.; Rewitz, K. Autophagy-Mediated Cholesterol Trafficking Controls Steroid Production. Dev. Cell 2019, 48, 659–671.e4. [Google Scholar] [CrossRef]
- Buzun, K.; Gornowicz, A.; Lesyk, R.; Bielawski, K.; Bielawska, A. Autophagy Modulators in Cancer Therapy. Int. J. Mol. Sci. 2021, 22, 5804. [Google Scholar] [CrossRef] [PubMed]
- Condello, M.; Pellegrini, E.; Caraglia, M.; Meschini, S. Targeting Autophagy to Overcome Human Diseases. Int. J. Mol. Sci. 2019, 20, 725. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Pattingre, S.; Sinha, S.; Bassik, M.; Levine, B. JNK1-mediated phosphorylation of Bcl-2 regulates starvation-induced autophagy. Mol. Cell 2008, 30, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Shi, M.; Maique, J.; Shaffer, J.; Yan, S.; Moe, O.W.; Hu, M.C. Beclin 1/Bcl-2 complex-dependent autophagy activity modulates renal susceptibility to ischemia-reperfusion injury and mediates renoprotection by Klotho. Am. J. Physiol. Renal. Physiol. 2020, 318, F772–F792. [Google Scholar] [CrossRef]
- Wu, H.; Song, A.; Hu, W.; Dai, M. The anti-atherosclerotic effect of paeonol against vascular smooth muscle cell proliferation by up-regulation of autophagy via the AMPK/mTOR signaling pathway. Front. Pharmacol. 2018, 8, 948. [Google Scholar] [CrossRef]
- Arab, H.H.; Ashour, A.M.; Eid, A.H.; Arafa, E.A.; Al Khabbaz, H.J.; El-Aal, S.A.A. Targeting oxidative stress, apoptosis, and autophagy by galangin mitigates cadmium-induced renal damage: Role of SIRT1/Nrf2 and AMPK/mTOR pathways. Life Sci. 2022, 291, 120300. [Google Scholar] [CrossRef]
- Bashir, N.; Shagirtha, K.; Manoharan, V.; Miltonprabu, S. The molecular and biochemical insight view of grape seed proanthocyanidins in ameliorating cadmium-induced testes-toxicity in rat model: Implication of PI3K/Akt/Nrf-2 signaling. Biosci. Rep. 2019, 39, BSR20180515. [Google Scholar] [CrossRef]
- Yilmaz, M.; Naziroglu, M.; Kutluhan, S.; Yilmaz, N.; Yurekli, V.A.; Vural, H. Topiramate modulates hippocampus NMDA receptors via brain Ca(2+) homeostasis in pentylentetrazol-induced epilepsy of rats. J. Recept. Signal Transduct Res. 2011, 31, 173–179. [Google Scholar] [CrossRef]
- Arab, H.H.; Eid, A.H.; Mahmoud, A.M.; Senousy, M.A. Linagliptin mitigates experimental inflammatory bowel disease in rats by targeting inflammatory and redox signaling. Life Sci. 2021, 273, 119295. [Google Scholar] [CrossRef]
- Yan, W.-J.; Mu, Y.; Yu, N.; Yi, T.-L.; Zhang, Y.; Pang, X.-L.; Cheng, D.; Yang, J. Protective effects of metformin on reproductive function in obese male rats induced by high-fat diet. J. Assist. Reprod. Genet. 2015, 32, 1097–1104. [Google Scholar] [CrossRef]
- Bearden, H.J.; Fuquay, J.W. Applied Animal Reproduction; Reston Publishing Company, Inc.: Reston, VA, USA, 1980. [Google Scholar]
- Elkhadragy, M.F.; Al-Olayan, E.M.; Al-Amiery, A.A.; Moneim, A.E.A. Protective Effects of Fragaria ananassa Extract against Cadmium Chloride-Induced Acute Renal Toxicity in Rats. Biol. Trace Elem. Res. 2018, 181, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Saad, M.A.; El-Sahhar, A.E.; Al-Shorbagy, M.Y. Mechanistic perspective of morin protection against ketoprofen-induced gastric mucosal injury: Targeting HMGB1/RAGE/NF-κB, DJ-1/Nrf2/HO-1 and PI3K/mTOR pathways. Arch. Biochem. Biophys. 2020, 693, 108552. [Google Scholar] [CrossRef] [PubMed]
- Salama, S.A.; Arab, H.H.; Hassan, M.H.; Maghrabi, I.A. Cadmium-induced hepatocellular injury: Modulatory effects of γ-glutamyl cysteine on the biomarkers of inflammation, DNA damage, and apoptotic cell death. J. Trace Elem. Med. Biol. 2019, 52, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Ashour, A.M.; Alqarni, A.M.; Arafa, E.-S.A.; Kabel, A.M. Camel Milk Mitigates Cyclosporine-Induced Renal Damage in Rats: Targeting p38/ERK/JNK MAPKs, NF-κB, and Matrix Metalloproteinases. Biology 2021, 10, 442. [Google Scholar] [CrossRef] [PubMed]
- Fikry, E.M.; Gad, A.M.; Eid, A.H.; Arab, H.H. Caffeic acid and ellagic acid ameliorate adjuvant-induced arthritis in rats via targeting inflammatory signals, chitinase-3-like protein-1 and angiogenesis. Biomed. Pharmacother. 2019, 110, 878–886. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Description |
|---|---|---|
| Control group | 6 | The oral vehicle of topiramate was received by gavage (0.5% carboxymethyl cellulose; CMC; 10 mL/kg/day). Meanwhile, normal saline vehicle (10 mL/kg/day) was given by gavage. The treatments lasted for 2 months. |
| Control + TPM group | 6 | Topiramate suspended in CMC was received by gavage (50 mg/kg/day, 10 mL/kg). Meanwhile, normal saline (10 mL/kg/day) was given by gavage. The treatments lasted for 2 months. |
| Cd group | 6 | 0.5% CMC (10 mL/kg/day) was received by gavage. Meanwhile, a gavage of cadmium chloride solution (5 mg/kg/day; 10 mL/kg) was given daily. The treatments lasted for 2 months. The dose of cadmium chloride is compliant with previous studies [20,47]. |
| Cd + TPM group | 6 | Topiramate (50 mg/kg/day, 10 mL/kg) was received by gavage. Meanwhile, a gavage of cadmium chloride solution (5 mg/kg/day; 10 mL/kg) was given daily. The treatments lasted for 2 months. The dose of topiramate was based on the previously reported efficacy against several pathological models [27,29,48]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arab, H.H.; Abd El Aal, H.A.; Alsufyani, S.E.; El-Sheikh, A.A.K.; Arafa, E.-S.A.; Ashour, A.M.; Kabel, A.M.; Eid, A.H. Topiramate Reprofiling for the Attenuation of Cadmium-Induced Testicular Impairment in Rats: Role of NLRP3 Inflammasome and AMPK/mTOR-Linked Autophagy. Pharmaceuticals 2022, 15, 1402. https://doi.org/10.3390/ph15111402
Arab HH, Abd El Aal HA, Alsufyani SE, El-Sheikh AAK, Arafa E-SA, Ashour AM, Kabel AM, Eid AH. Topiramate Reprofiling for the Attenuation of Cadmium-Induced Testicular Impairment in Rats: Role of NLRP3 Inflammasome and AMPK/mTOR-Linked Autophagy. Pharmaceuticals. 2022; 15(11):1402. https://doi.org/10.3390/ph15111402
Chicago/Turabian StyleArab, Hany H., Hayat A. Abd El Aal, Shuruq E. Alsufyani, Azza A. K. El-Sheikh, El-Shaimaa A. Arafa, Ahmed M. Ashour, Ahmed M. Kabel, and Ahmed H. Eid. 2022. "Topiramate Reprofiling for the Attenuation of Cadmium-Induced Testicular Impairment in Rats: Role of NLRP3 Inflammasome and AMPK/mTOR-Linked Autophagy" Pharmaceuticals 15, no. 11: 1402. https://doi.org/10.3390/ph15111402
APA StyleArab, H. H., Abd El Aal, H. A., Alsufyani, S. E., El-Sheikh, A. A. K., Arafa, E.-S. A., Ashour, A. M., Kabel, A. M., & Eid, A. H. (2022). Topiramate Reprofiling for the Attenuation of Cadmium-Induced Testicular Impairment in Rats: Role of NLRP3 Inflammasome and AMPK/mTOR-Linked Autophagy. Pharmaceuticals, 15(11), 1402. https://doi.org/10.3390/ph15111402