
Preclinical Evaluation of the Multiple Tyrosine Kinases Inhibitor Anlotinib in Leukemia Stem Cells

 ,
,
Abstract
1. Introduction
2. Results
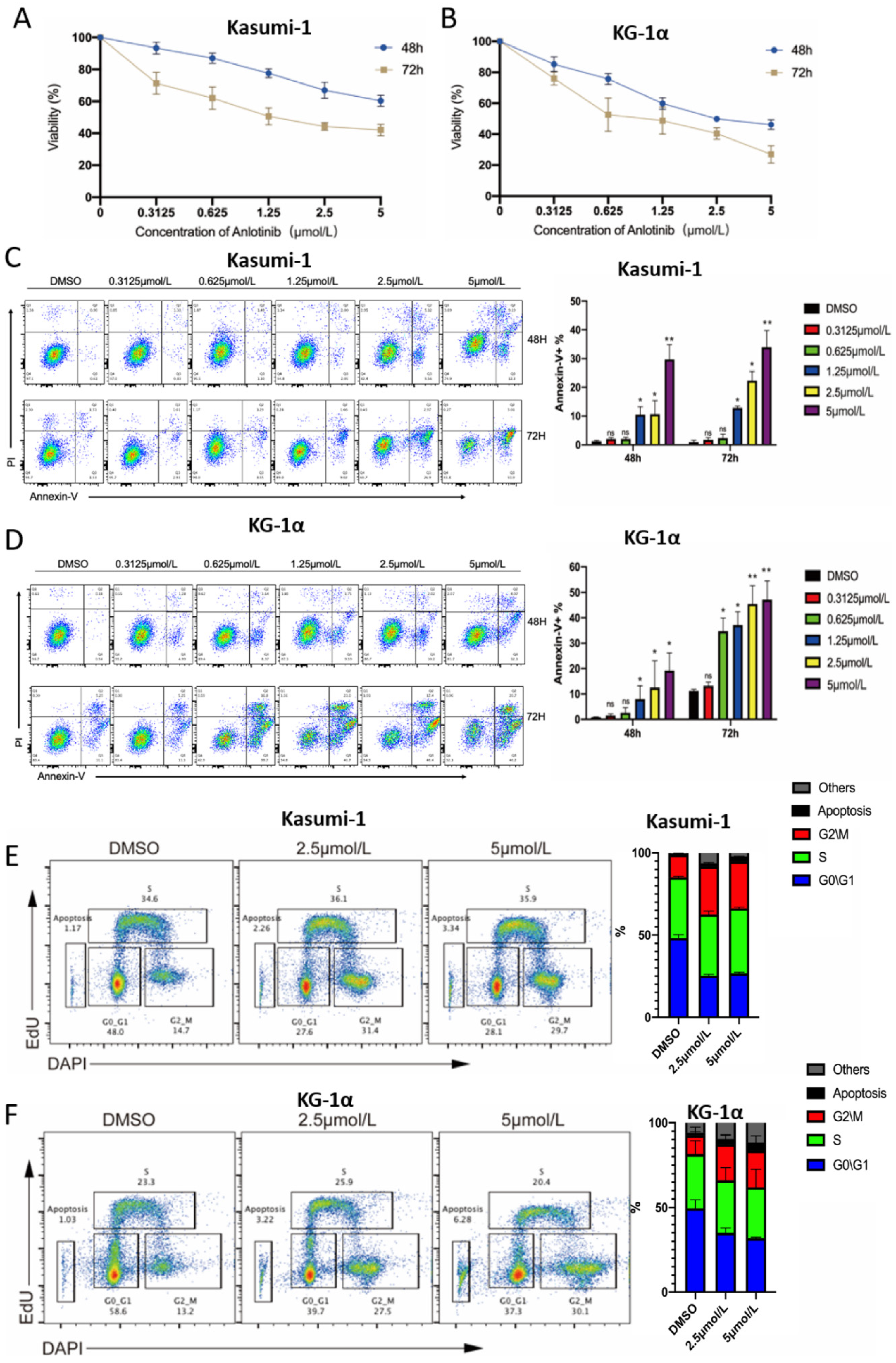
2.1. Anlotinib Exerts Potential Anti-LSCs Effects by Inhibiting Proliferation and Inducing Apoptosis of LSC-Like Cells In Vitro
2.2. Anlotinib Targets Primary LSCs While Sparing Normal Mononuclear Cells Ex Vivo
2.3. Anlotinib Impairs the Regeneration Capacity of LSCs in Patient-Derived Xenograft Mice Models
2.4. Anti-LSCs Activity of Anlotinib Might Be Associated with the Inhibition of the JAK-STAT Pathway
2.5. Anlotinib Enhances the Sensitivity of LSC-Like Cells to Idarubicin by Regulating Apoptosis-Related Proteins
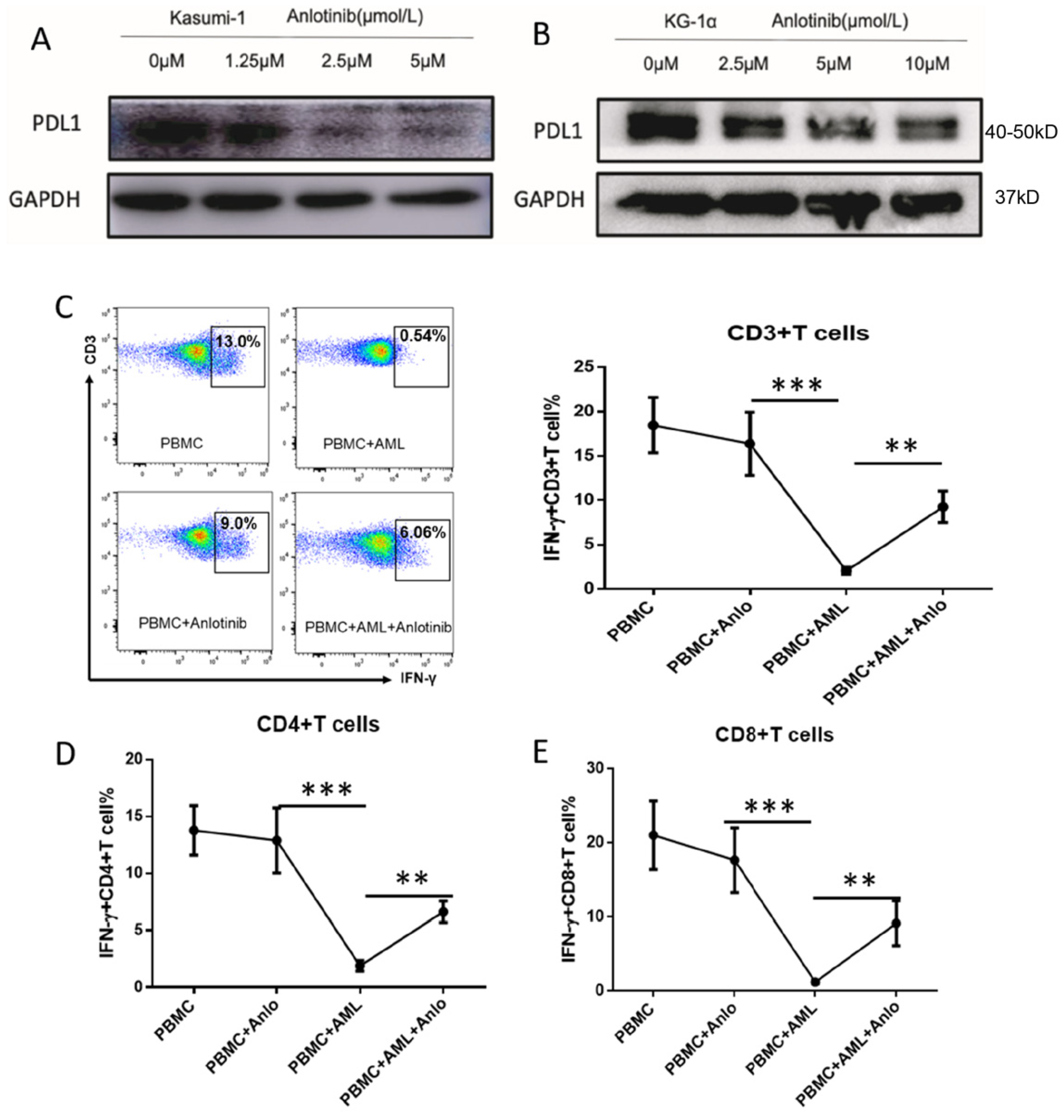
2.6. Anlotinib Disrupts the Immunosuppressive Effects of LSC-Like Cells on T Cells via Downregulation of PD-L1
3. Materials and Methods
3.1. Cell Line Culture
3.2. Patients and Donor Samples
3.3. Cell Viability Assay
3.4. Detection of Apoptosis
3.5. Ex-Vivo Evaluation of Cytokine Production of T Cells
3.6. Western Blot Analysis
3.7. AML Patient-Derived Xenograft (PDX) Mice Models
3.8. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sami, S.A.; Darwish, N.H.E.; Barile, A.N.M.; Mousa, S.A. Current and Future Molecular Targets for Acute Myeloid Leukemia Therapy. Curr. Treat. Options Oncol. 2020, 21, 3. [Google Scholar] [CrossRef]
- Chasov, V.; Mirgayazova, R.; Zmievskaya, E.; Khadiullina, R.; Valiullina, A.; Clarke, J.S.; Rizvanov, A.; Baud, M.G.J.; Bulatov, E. Key Players in the Mutant p53 Team: Small Molecules, Gene Editing, Immunotherapy. Front. Oncol. 2020, 10, 1460. [Google Scholar] [CrossRef]
- Short, N.J.; Konopleva, M.; Kadia, T.M.; Borthakur, G.; Ravandi, F.; DiNardo, C.D.; Daver, N. Advances in the Treatment of Acute Myeloid Leukemia: New Drugs and New Challenges. Cancer Discov. 2020, 10, 506–525. [Google Scholar] [CrossRef]
- Thomas, D.; Majeti, R. Biology and relevance of human acute myeloid leukemia stem cells. Blood 2017, 129, 1577–1585. [Google Scholar]
- Li, Y.; Wang, Y.; Zhou, Y.; Li, J.; Chen, K.; Zhang, L.; Deng, M.; Deng, S.; Li, P.; Xu, B. Cooperative effect of chidamide and chemotherapeutic drugs induce apoptosis by DNA damage accumulation and repair defects in acute myeloid leukemia stem and progenitor cells. Clin. Epigenetics 2017, 9, 83. [Google Scholar] [CrossRef]
- Xu, B.; Wang, S.; Li, R.; Chen, K.; He, L.; Deng, M.; Kannappan, V.; Zha, J.; Dong, H.; Wang, W. Disulfiram/copper selectively eradicates AML leukemia stem cells in vitro and in vivo by simultaneous induction of ROS-JNK and inhibition of NF-kappaB and Nrf2. Cell Death Dis. 2017, 8, e2797. [Google Scholar]
- Ding, Y.; Gao, H.; Zhang, Q. The biomarkers of leukemia stem cells in acute myeloid leukemia. Stem Cell Investig. 2017, 4, 19. [Google Scholar]
- Vetrie, D.; Helgason, G.V.; Copland, M. The leukaemia stem cell: Similarities, differences and clinical prospects in CML and AML. Nat. Rev. Cancer 2020, 20, 158–173. [Google Scholar] [CrossRef]
- Li, Y.; Chen, K.; Zhou, Y.; Xiao, Y.; Deng, M.; Jiang, Z.; Ye, W.; Wang, X.; Wei, X.; Li, J.; et al. A New Strategy to Target Acute Myeloid Leukemia Stem and Progenitor Cells Using Chidamide, a Histone Deacetylase Inhibitor. Curr. Cancer Drug Targets 2015, 15, 493–503. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Y.; Chen, K.; Shi, P.; Li, Y.; Deng, M.; Jiang, Z.; Wang, X.; Li, P.; Xu, B. The pan-Bcl2 Inhibitor AT101 Activates the Intrinsic Apoptotic Pathway and Causes DNA Damage in Acute Myeloid Leukemia Stem-Like Cells. Target. Oncol. 2017, 12, 677–687. [Google Scholar] [CrossRef]
- Cook, A.; Li, L.; Ho, Y.; Lin, A.; Stein, A.; Forman, S.; Perrotti, D.; Jove, R.; Bhatia, R. Role of altered growth factor receptor-mediated JAK2 signaling in growth and maintenance of human acute myeloid leukemia stem cells. Blood 2014, 123, 2826–2837. [Google Scholar] [CrossRef]
- Saito, Y.; Kitamura, H.; Hijikata, A.; Tomizawa-Murasawa, M.; Tanaka, S.; Takagi, S.; Uchida, N.; Suzuki, N.; Sone, A.; Najima, Y.; et al. Identification of Therapeutic Targets for Quiescent, Chemotherapy-Resistant Human Leukemia Stem Cells. Sci. Transl. Med. 2010, 2, 17ra9. [Google Scholar] [CrossRef]
- Schepers, K.; Campbell, T.B.; Passegué, E. Normal and Leukemic Stem Cell Niches: Insights and Therapeutic Opportunities. Cell Stem Cell 2015, 16, 254–267. [Google Scholar] [CrossRef]
- Berenstein, R. Class III Receptor Tyrosine Kinases in Acute Leukemia—Biological Functions and Modern Laboratory Analysis. Biomark. Insights 2015, 10, 1–14. [Google Scholar] [CrossRef]
- Wingelhofer, B.; Neubauer, H.A.; Valent, P.; Han, X.; Constantinescu, S.N.; Gunning, P.T.; Müller, M.; Moriggl, R. Implications of STAT3 and STAT5 signaling on gene regulation and chromatin remodeling in hematopoietic cancer. Leukemia 2018, 32, 1713–1726. [Google Scholar] [CrossRef]
- Halim, C.E.; Deng, S.; Ong, M.S.; Yap, C.T. Involvement of STAT5 in Oncogenesis. Biomedicines 2020, 8, 316. [Google Scholar] [CrossRef]
- Prestipino, A.; Emhardt, A.J.; Aumann, K.; Sullivan, D.O.; Gorantla, S.P.; Duquesne, S.; Melchinger, W.; Braun, L.; Vuckovic, S.; Boerries, M.; et al. Oncogenic JAK2 V617F causes PD-L1 expression, mediating immune escape in myeloproliferative neoplasms. Sci. Transl. Med. 2018, 10, eaam7729. [Google Scholar] [CrossRef]
- Shen, G.; Zheng, F.; Ren, D.; Du, F.; Dong, Q.; Wang, Z.; Zhao, F.; Ahmad, R.; Zhao, J. Anlotinib: A novel multi-targeting tyrosine kinase inhibitor in clinical development. J. Hematol. Oncol. 2018, 11, 120. [Google Scholar] [CrossRef]
- Han, B.; Li, K.; Wang, Q.; Zhang, L.; Shi, J.; Wang, Z.; Cheng, Y.; He, J.; Shi, Y.; Zhao, Y.; et al. Effect of Anlotinib as a Third-Line or Further Treatment on Overall Survival of Patients With Advanced Non-Small Cell Lung Cancer: The ALTER 0303 Phase 3 Randomized Clinical Trial. JAMA Oncol. 2018, 4, 1569–1575. [Google Scholar]
- Li, S. Anlotinib: A Novel Targeted Drug for Bone and Soft Tissue Sarcoma. Front. Oncol. 2021, 11, 664853. [Google Scholar] [CrossRef]
- Liang, L.; Hui, K.; Hu, C.; Wen, Y.; Yang, S.; Zhu, P.; Wang, L.; Xia, Y.; Qiao, Y.; Sun, W.; et al. Autophagy inhibition potentiates the anti-angiogenic property of multikinase inhibitor anlotinib through JAK2/STAT3/VEGFA signaling in non-small cell lung cancer cells. J. Exp. Clin. Cancer Res. 2019, 38, 71. [Google Scholar]
- Karajannis, M.A.; Vincent, L.; DiRenzo, R.; Shmelkov, S.V.; Zhang, F.; Feldman, E.J.; Bohlen, P.; Zhu, Z.; Sun, H.; Kussie, P.; et al. Activation of FGFR1β signaling pathway promotes survival, migration and resistance to chemotherapy in acute myeloid leukemia cells. Leukemia 2006, 20, 979–986. [Google Scholar] [CrossRef]
- Li, P.; Huang, T.; Zou, Q.; Liu, D.; Wang, Y.; Tan, X.; Wei, Y.; Qiu, H. FGFR2 Promotes Expression of PD-L1 in Colorectal Cancer via the JAK/STAT3 Signaling Pathway. J. Immunol. 2019, 202, 3065–3075. [Google Scholar] [CrossRef]
- Carter, J.L.; Hege, K.; Yang, J.; Kalpage, H.A.; Su, Y.; Edwards, H.; Hüttemann, M.; Taub, J.W.; Ge, Y. Targeting multiple signaling pathways: The new approach to acute myeloid leukemia therapy. Signal Transduct. Target. Ther. 2020, 5, 288. [Google Scholar] [CrossRef]
- Yumeen, S.; Mirza, F.N.; Lewis, J.M.; King, A.L.O.; Kim, S.R.; Carlson, K.R.; Umlauf, S.R.; Surovtseva, Y.V.; Foss, F.M.; Girardi, M. JAK inhibition synergistically potentiates BCL2, BET, HDAC, and proteasome inhibition in advanced CTCL. Blood Adv. 2020, 4, 2213–2226. [Google Scholar] [CrossRef]
- Wojcik, I.; Szybka, M.; Golanska, E.; Rieske, P.; Blonski, J.Z.; Robak, T.; Bartkowiak, J. Abnormalities of the P53, MDM2, BCL2 and BAX genes in acute leukemias. Neoplasma 2005, 52, 318–324. [Google Scholar]
- Ajayi, S.; Becker, H.; Reinhardt, H.; Engelhardt, M.; Zeiser, R.; Bubnoff, N.V.; Wäsch, R. Ruxolitinib. Recent Results Cancer Res. 2018, 212, 119–132. [Google Scholar]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Yogarajah, M.; Stone, R.M. A concise review of BCL-2 inhibition in acute myeloid leukemia. Expert Rev. Hematol. 2018, 11, 145–154. [Google Scholar]
- Milara, J.; Hernandez, G.; Ballester, B.; Morell, A.; Roger, I.; Montero, P.; Escrivá, J.; Lloris, J.M.; Molina-Molina, M.; Morcillo, E.; et al. The JAK2 pathway is activated in idiopathic pulmonary fibrosis. Respir. Res. 2018, 19, 24. [Google Scholar] [CrossRef]
- Zhang, L.; Gajewski, T.F.; Kline, J. PD-1/PD-L1 interactions inhibit antitumor immune responses in a murine acute myeloid leukemia model. Blood 2009, 114, 1545–1552. [Google Scholar]
- Liu, L.; Chang, Y.-J.; Xu, L.-P.; Zhang, X.-H.; Wang, Y.; Liu, K.-Y.; Huang, X.-J. Reversal of T Cell Exhaustion by the First Donor Lymphocyte Infusion Is Associated with the Persistently Effective Antileukemic Responses in Patients with Relapsed AML after Allo-HSCT. Biol. Blood Marrow Transplant. 2018, 24, 1350–1359. [Google Scholar] [CrossRef]
- Kong, Y.; Zhu, L.; Schell, T.D.; Zhang, J.; Claxton, D.F.; Ehmann, W.C.; Rybka, W.B.; George, M.R.; Zeng, H.; Zheng, H. T-Cell Immunoglobulin and ITIM Domain (TIGIT) Associates with CD8+ T-Cell Exhaustion and Poor Clinical Outcome in AML Patients. Clin. Cancer Res. 2016, 22, 3057–3066. [Google Scholar] [CrossRef]
- Liu, S.; Qin, T.; Liu, Z.; Wang, J.; Jia, Y.; Feng, Y.; Gao, Y.; Li, K. Anlotinib alters tumor immune microenvironment by downregulating PD-L1 expression on vascular endothelial cells. Cell Death Dis. 2020, 11, 309. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Gender | Age | FAB | WBC (109/L) | Blasts (%) | Cytogenetics | Molecular Mutations |
|---|---|---|---|---|---|---|---|
| 1 | M | 39 | M1 | 102.03 | 90.7 | 46,XY | ASXL1,IDH1,TET2 |
| 2 | F | 34 | M5b | 74 | 90.1 | 46,XX | NPM1,FLT3-ITD |
| 3 | M | 51 | M2 | 122 | 81.5 | 46,XY | CEBPA,TET2,ASXL1 |
| 4 | M | 45 | M5 | 190.3 | 79.5 | 46,XY | FLT3-ITD, NPM1 |
| 5 | M | 38 | M2a | 29.3 | 46 | 46,XY | NRAS,PHF6,TET2,TP53 |
| 6 | M | 40 | M2 | 10.75 | 69.3 | 46,XY | CEBPA,TAD1,bZIP |
| 7 | F | 39 | M5b | 48.9 | 59.5 | 46,XX,t (9;11) (p22;q23) | MLL-AF9 |
| 8 | M | 56 | M2b | 44.4 | 21.7 | 45,XY,t(8;21)(q22;q22) | CEBPA,AML1-ETO |
| 9 | M | 16 | M2 | 41.7 | 81 | 46,XY | TET2, CEBPA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Liu, L.; Jiang, Y.; Li, Z.; Feng, L.; Zhuang, X.; Lin, Z.; Chen, Q.; Chen, G.; He, J.; et al. Preclinical Evaluation of the Multiple Tyrosine Kinases Inhibitor Anlotinib in Leukemia Stem Cells. Pharmaceuticals 2022, 15, 1313. https://doi.org/10.3390/ph15111313
Jiang Y, Liu L, Jiang Y, Li Z, Feng L, Zhuang X, Lin Z, Chen Q, Chen G, He J, et al. Preclinical Evaluation of the Multiple Tyrosine Kinases Inhibitor Anlotinib in Leukemia Stem Cells. Pharmaceuticals. 2022; 15(11):1313. https://doi.org/10.3390/ph15111313
Chicago/Turabian StyleJiang, Yuelong, Long Liu, Yirong Jiang, Zhifeng Li, Liying Feng, Xinguo Zhuang, Zhijuan Lin, Qiuling Chen, Guoshu Chen, Jixiang He, and et al. 2022. "Preclinical Evaluation of the Multiple Tyrosine Kinases Inhibitor Anlotinib in Leukemia Stem Cells" Pharmaceuticals 15, no. 11: 1313. https://doi.org/10.3390/ph15111313
APA StyleJiang, Y., Liu, L., Jiang, Y., Li, Z., Feng, L., Zhuang, X., Lin, Z., Chen, Q., Chen, G., He, J., Li, G., Zha, J., & Xu, B. (2022). Preclinical Evaluation of the Multiple Tyrosine Kinases Inhibitor Anlotinib in Leukemia Stem Cells. Pharmaceuticals, 15(11), 1313. https://doi.org/10.3390/ph15111313