
Valorisation of the Inhibitory Potential of Fresh and Dried Fruit Extracts of Prunus spinosa L. towards Carbohydrate Hydrolysing Enzymes, Protein Glycation, Multiple Oxidants and Oxidative Stress-Induced Changes in Human Plasma Constituents
 , ,
, ,  , ,
, ,  and
and
Abstract
1. Introduction
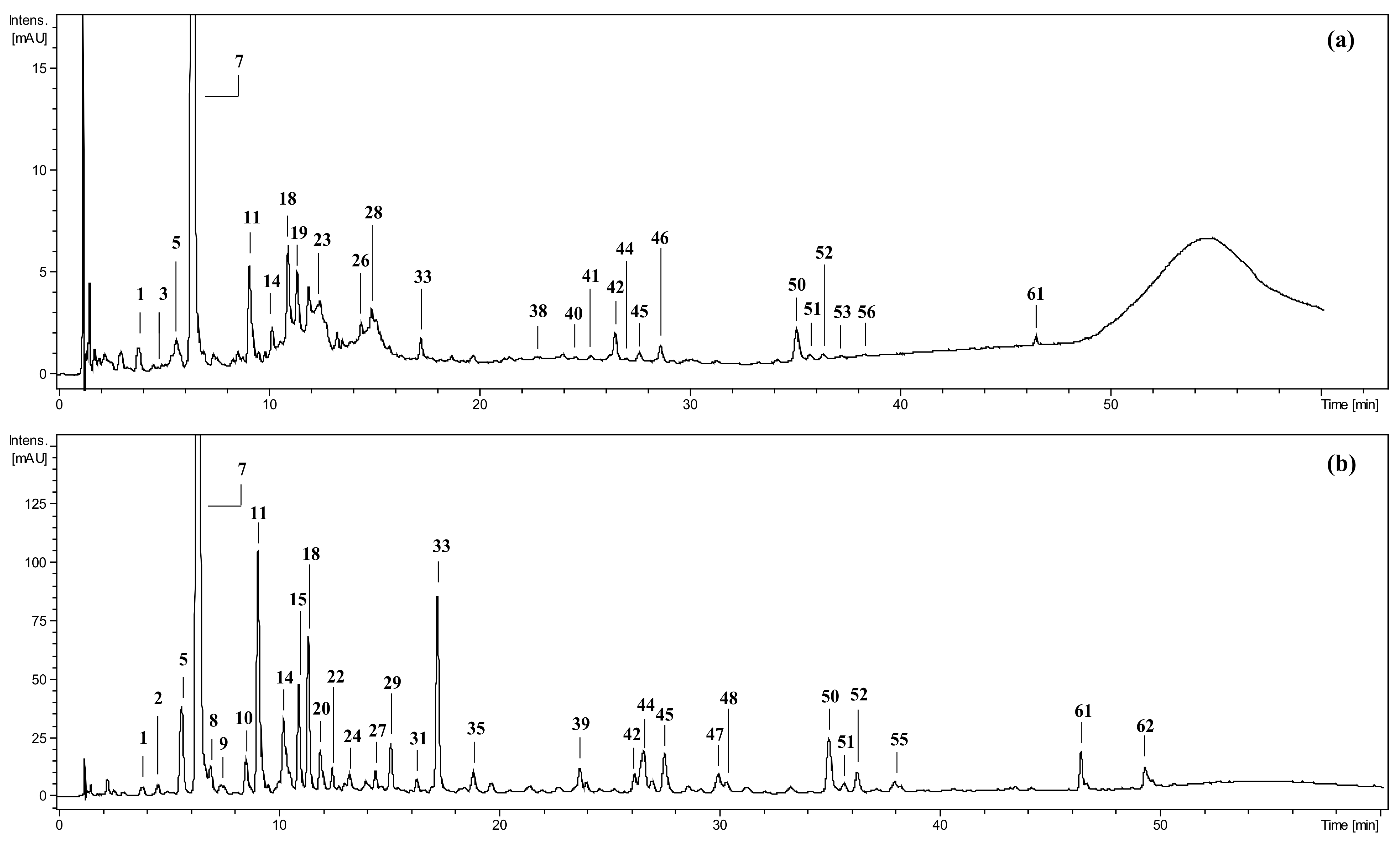
2. Results and Discussion
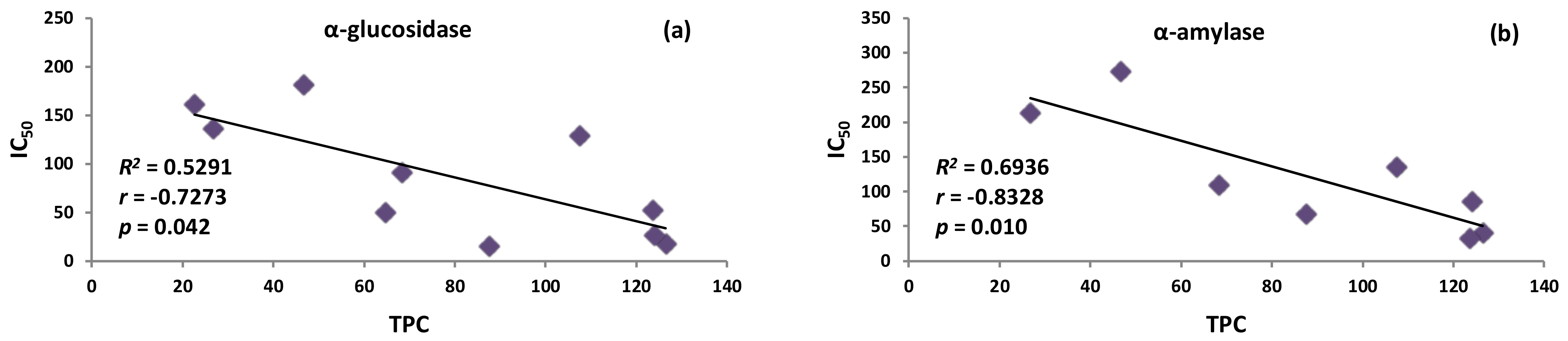
2.1. Inhibition of Digestive Enzymes Related to Diabetes Mellitus (DM)
2.2. Scavenging of Multiple ROS
2.3. Inhibition of the AGEs Generation
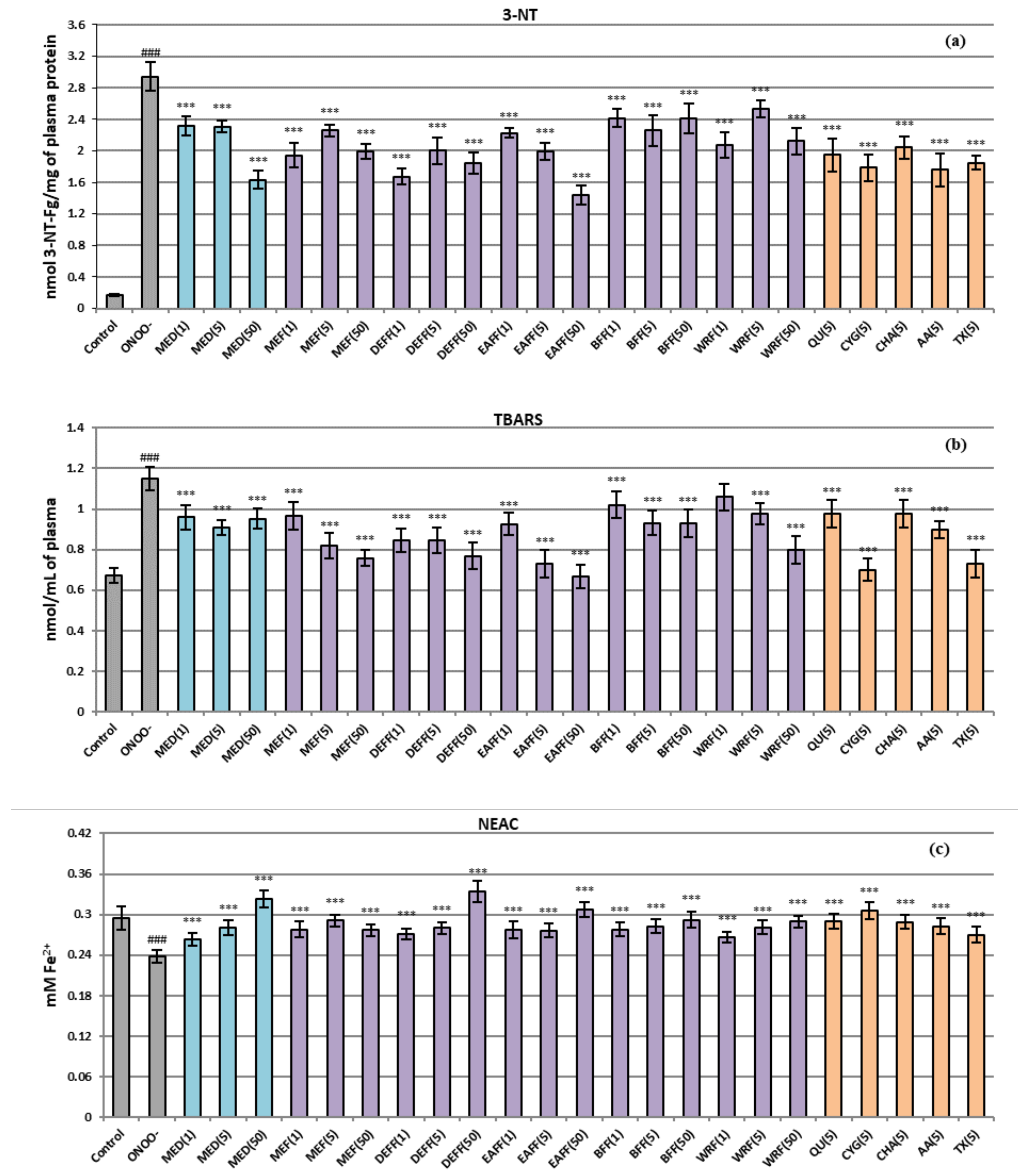
2.4. Antioxidant Protection of Human Plasma Components against Oxidative Stress
2.5. Biological Potential of Maillard Reaction Products (MRPs) in Dried Fruits
3. Materials and Methods
3.1. Study Material, Extraction and Phytochemical Standardisation
3.2. α-Glucosidase and α-Amylase Inhibition Assays
3.3. Multiple Oxidants Scavenging Activity
3.4. Inhibition of the Formation of Advanced Glycation Endproducts (AGEs)
3.5. Protective Effects on Human Plasma Exposed to Oxidative Stress
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3-NT-Fg | nitro-fibrinogen |
| ABTS | 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) |
| AGEs | advanced glycation end products |
| BFD | n-butanol fraction of MED |
| BFF | n-butanol fraction of MEF |
| DEFD | diethyl ether fraction of MED |
| DEFF | diethyl ether fraction of MEF |
| DM | diabetes mellitus |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| dw | dry weight |
| EAFD | ethyl acetate fraction of MED |
| EAFF | ethyl acetate fraction of MEF |
| FRAP | ferric-reducing ability of plasma |
| GAE | gallic acid equivalents |
| H2O2 | hydrogen peroxide |
| HMF | 5-hydroxymetylfurfural |
| HO• | hydroxyl radical |
| HOCl | hypochlorous acid |
| MED | methanol–water (75:25, v/v) extract of dried fruits |
| MEF | methanol–water (75:25, v/v) extract of fresh fruits |
| MRPs | Maillard reaction products |
| NEAC | non-enzymatic antioxidant capacity of human plasma |
| NF-κB | nuclear factor kappa B |
| NO• | nitrogen oxide |
| O2•− | superoxide anion radical |
| ONOO− | peroxynitrite |
| PB2 | procyanidin B2 |
| RAGE | AGEs-specific receptor |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| TAC | total content of anthocyanins |
| TBARS | thiobarbituric acid-reactive substances |
| TFL | total content of flavonoids |
| TPA | total content of phenolic acids |
| TPC | total phenolic content (Folin–Ciocalteu assay) |
| TPH | total phenolic content (sum of individual phenolics by HPLC) |
| TTC | total content of tannin-type proanthocyanidins |
| WRD | water residue of MED after fractionation |
| WRF | water residue of MEF after fractionation |
References
- Ullah, A.; Khan, A.; Khan, I. Diabetes mellitus and oxidative stress—A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar]
- The International Diabetes Federation (IDF). IDF Diabetes Atlas 2021—10th Edition. Available online: https://diabetesatlas.org/atlas/tenth-edition/ (accessed on 22 June 2022).
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Merino, J.; Sun, Q.; Fitó, M.; Salas-Salvadó, J. Dietary polyphenols, mediterranean diet, prediabetes, and type 2 diabetes: A narrative review of the evidence. Oxid. Med. Cell. Longev. 2017, 2017, 6723931. [Google Scholar] [CrossRef] [PubMed]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef] [PubMed]
- Popescu, I.; Caudullo, G. Prunus spinosa in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e01aa69+. Available online: https://w3id.org/mtv/FISE-Comm/v01/e01aa69 (accessed on 22 June 2022).
- Facciola, S. Cornucopia II: A Source Book of Edible Plants, 2nd ed.; Kampong Publications: Vista, CA, USA, 1998; p. 204. [Google Scholar]
- Alarcόn, R.; Pardo-de-Santayana, M.; Priestley, C.; Morales, R.; Heinrich, M. Medicinal and local food plants in the south of Alava (Basque Country, Spain). J. Ethnopharmacol. 2015, 176, 207–224. [Google Scholar] [CrossRef]
- Kültür, S. Medicinal plants used in Kirklareli Province (Turkey). J. Ethnopharmacol. 2007, 111, 341–364. [Google Scholar] [CrossRef]
- Jarić, S.; Mačukanović-Jocić, M.; Djurdjević, L.; Mitrović, M.; Kostić, O.; Karadžić, B.; Pavlović, P. An ethnobotanical survey of traditionally used plants on Suva planina mountain (south-eastern Serbia). J. Ethnopharmacol. 2015, 175, 93–108. [Google Scholar] [CrossRef]
- Magiera, A.; Czerwińska, M.E.; Owczarek, A.; Marchelak, A.; Granica, S.; Olszewska, M.A. Polyphenol-enriched extracts of Prunus spinosa fruits: Anti-inflammatory and antioxidant effects in human immune cells ex vivo in relation to phytochemical profile. Molecules 2022, 27, 1691. [Google Scholar] [CrossRef]
- Magiera, A.; Czerwińska, M.E.; Owczarek, A.; Marchelak, A.; Granica, S.; Olszewska, M.A. Polyphenols and Maillard reaction products in dried Prunus spinosa fruits: Quality aspects and contribution to anti-inflammatory and antioxidant activity in human immune cells ex vivo. Molecules 2022, 27, 3302. [Google Scholar] [CrossRef]
- Sabatini, L.; Fraternale, D.; Di Giacomo, B.; Mari, M.; Albertini, M.C.; Gordillo, B.; Rocchi, M.B.L.; Sisti, D.; Coppari, S.; Semprucci, F.; et al. Chemical composition, antioxidant, antimicrobial and anti-inflammatory activity of Prunus spinosa L. fruit ethanol extract. J. Funct. Foods 2020, 67, 103885. [Google Scholar] [CrossRef]
- Popović, B.M.; Blagojević, B.; Ždero Pavlović, R.; Mićić, N.; Bijelić, S.; Bogdanović, B.; Mišan, A.; Duarte, C.M.M.; Serra, A.T. Comparison between polyphenol profile and bioactive response in blackthorn (Prunus spinosa L.) genotypes from north Serbia-from raw data to PCA analysis. Food Chem. 2020, 302, 125373. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, M.; Olszewska, M.A.; Kolodziejczyk-Czepas, J.; Nowak, P.; Owczarek, A. Sorbus domestica leaf extracts and their activity markers: Antioxidant potential and synergy effects in scavenging assays of multiple oxidants. Molecules 2019, 24, 2289. [Google Scholar] [CrossRef] [PubMed]
- Vlassara, H.; Striker, G.E. Advanced glycation endproducts in diabetes and diabetic complications. Endocrinol. Metab. Clin. N. Am. 2013, 42, 697–719. [Google Scholar] [CrossRef] [PubMed]
- Marín-Peñalver, J.J.; Martín-Timón, I.; Sevillano-Collantes, C.; Del Cañizo-Gómez, F.J. Update on the treatment of type 2 diabetes mellitus. World J. Diabetes 2016, 7, 354–395. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, P.J.; Warren, F.J.; Ellis, P.R. Human α-amylase and starch digestion: An interesting marriage. Starch-Stärke 2011, 63, 395–405. [Google Scholar] [CrossRef]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of polysaccharides in food, digestion, and health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J. Diabetes Metab. Disord. 2013, 12, 43. [Google Scholar] [CrossRef]
- Bento, C.; Gonçalves, A.C.; Silva, B.; Silva, L.R. Assessing the phenolic profile, antioxidant, antidiabetic and protective effects against oxidative damage in human erythrocytes of peaches from Fundão. J. Funct. Foods 2018, 43, 224–233. [Google Scholar] [CrossRef]
- Cui, J.; Gu, X.; Wang, F.; Ouyang, J.; Wang, J. Purification and structural characterization of an α-glucosidase inhibitory polysaccharide from apricot (Armeniaca sibirica L. Lam.) pulp. Carbohydr. Polym. 2015, 121, 309–314. [Google Scholar] [CrossRef]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef]
- Rutkowska, M.; Kołodziejczyk-Czepas, J.; Owczarek, A.; Zakrzewska, A.; Magiera, A.; Olszewska, M.A. Novel insight into biological activity and phytochemical composition of Sorbus aucuparia L. fruits: Fractionated extracts as inhibitors of protein glycation and oxidative/nitrative damage of human plasma components. Food Res. Int. 2021, 147, 110526. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiang, L.; Wang, C.; Tang, C.; He, X. Antidiabetic and antioxidant effects and phytochemicals of mulberry fruit (Morus alba L.) polyphenol enhanced extract. PLoS ONE 2013, 8, e71144. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, Q.; Zhao, M.; Yang, P.; Hu, X.; Ouyang, D. Flavonoid glycosides from seeds of Hippophae rhamnoides subsp. sinensis with α-glucosidase inhibition activity. Fitoterapia 2019, 137, 104248. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Locatelli, M.; Macone, A.; Toniolo, C.; Cesa, S.; Carradori, S.; Eufemi, M.; Mazzanti, G.; Di Giacomo, S. Hypoglycemic, antiglycation, and cytoprotective properties of a phenol-rich extract from waste peel of Punica granatum L. var. Dente di Cavallo DC2. Molecules 2019, 24, 3103. [Google Scholar] [CrossRef]
- Truba, J.; Stanisławska, I.; Walasek, M.; Wieczorkowska, W.; Woliński, K.; Buchholz, T.; Melzig, M.F.; Czerwińska, M.E. Inhibition of digestive enzymes and antioxidant activity of extracts from fruits of Cornus alba, Cornus sanguinea subsp. hungarica and Cornus florida-A comparative study. Plants 2020, 9, 122. [Google Scholar] [CrossRef]
- Amorim, F.L.; de Cerqueira Silva, M.B.; Cirqueira, M.G.; Oliveira, R.S.; Machado, B.A.S.; Gomes, R.G.; de Souza, C.O.; Druzian, J.I.; de Souza Ferreira, E.; Umsza-Guez, M.A. Grape peel (Syrah var.) jam as a polyphenol-enriched functional food ingredient. Food Sci. Nutr. 2019, 7, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Barros, L.; Carvalho, A.M.; Sá Morais, J.; Ferreira, I.C.F.R. Strawberry-tree, blackthorn and rose fruits: Detailed characterisation in nutrients and phytochemicals with antioxidant properties. Food Chem. 2010, 120, 247–254. [Google Scholar] [CrossRef]
- Ruiz-Rodríguez, B.M.; De Ancos, B.; Sánchez-Moreno, C.; Fernández-Ruiz, V.; De Cortes Sánchez-Mata, M.; Cámara, M.; Tardío, J. Wild blackthorn (Prunus spinosa L.) and hawthorn (Crataegus monogyna Jacq.) fruits as valuable sources of antioxidants. Fruits 2014, 69, 61–73. [Google Scholar] [CrossRef]
- Fraternale, D.; Giamperi, L.; Bucchini, A.; Sestili, P.; Paolillo, M.; Riccia, D. Prunus spinosa fresh fruit juice: Antioxidant activity in cell-free and cellular systems. Nat. Prod. Commun. 2009, 4, 1665–1670. [Google Scholar] [PubMed]
- Hajimehdipoor, H.; Shahrestani, R.; Shekarchi, M. Investigating the synergistic antioxidant effects of some flavonoid and phenolic compounds. Res. J. Pharmacogn. 2014, 1, 35–40. [Google Scholar]
- Kicel, A.; Owczarek, A.; Kapusta, P.; Kolodziejczyk-Czepas, J.; Olszewska, M.A. Contribution of individual polyphenols to antioxidant activity of Cotoneaster bullatus and Cotoneaster zabelii leaves-structural relationships, synergy effects and application for quality control. Antioxidants 2020, 9, 69. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef]
- Tan, K.C.; Shiu, S.W.; Wong, Y.; Tam, X. Serum advanced glycation end products (AGEs) are associated with insulin resistance. Diabetes Metab. Res. Rev. 2011, 27, 488–492. [Google Scholar] [CrossRef]
- Ramkisoon, J.S.; Mahomoodally, F.M.; Ahmed, N.; Subratty, H.A. Natural inhibitors of advanced glycation end-products. Nutr. Food Sci. 2012, 42, 397–404. [Google Scholar] [CrossRef]
- Yeh, W.J.; Hsia, S.M.; Lee, W.H.; Wu, C.H. Polyphenols with antiglycation activity and mechanisms of action: A review of recent findings. J. Food Drug. Anal. 2017, 25, 84–92. [Google Scholar] [CrossRef]
- Starowicz, M.; Zieliński, H. Inhibition of advanced glycation end-product formation by high antioxidant-leveled spices commonly used in European cuisine. Antioxidants 2019, 8, 100. [Google Scholar] [CrossRef]
- Ma, H.; Johnson, S.L.; Liu, W.; DaSilva, N.A.; Meschwitz, S.; Dain, J.A.; Seeram, N.P. Evaluation of polyphenol anthocyanin-enriched extracts of blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry for free radical scavenging, reactive carbonyl species trapping, anti-glycation, anti-β-amyloid aggregation, and microglial neuroprotective effects. Int. J. Mol. Sci. 2018, 19, 461. [Google Scholar]
- Kicel, A.; Magiera, A.; Skrzywanek, M.; Malczuk, M.; Olszewska, M.A. Inhibition of α-glucosidase, α-amylase and protein glycation by phenolic extracts of Cotoneaster bullatus, Cotoneaster zabelii and Cotoneaster integerrimus leaves and fruits: Focus on anti-hyperglycemic activity and kinetic parameters. Molecules 2022, 27, 7081. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Szabó, C.; Ischiropoulos, H.; Radi, R. Peroxynitrite: Biochemistry, pathophysiology and development of therapeutics. Nat. Rev. Drug Discov. 2007, 6, 662–680. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Szabo, C. Role of peroxynitrite in the pathogenesis of cardiovascular complications of diabetes. Curr. Opin. Pharmacol. 2006, 6, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Marchelak, A.; Owczarek, A.; Matczak, M.; Pawlak, A.; Kołodziejczyk-Czepas, J.; Nowak, P.; Olszewska, M.A. Bioactivity potential of Prunus spinosa L. flower extracts: Phytochemical profiling, cellular safety, pro-inflammatory enzymes inhibition and protective effects against oxidative stress in vitro. Front. Pharmacol. 2017, 8, 680. [Google Scholar] [CrossRef]
- Ahsan, H. 3-Nitrotyrosine: A biomarker of nitrogen free radical species modified proteins in systemic autoimmunogenic conditions. Hum. Immunol. 2013, 74, 1392–1399. [Google Scholar] [CrossRef]
- Atkin, M.A.; Gasper, A.; Ullegaddi, R.; Powers, H.J. Oxidative susceptibility of unfractionated serum or plasma: Response to antioxidants in vitro and to antioxidant supplementation. Clin. Chem. 2005, 51, 2138–2144. [Google Scholar] [CrossRef][Green Version]
- Shapla, U.M.; Solayman, M.; Alam, N.; Khalil, M.I.; Gan, S.H. 5-Hydroxymethylfurfural (HMF) levels in honey and other food products: Effects on bees and human health. Chem. Cent. J. 2018, 12, 35. [Google Scholar] [CrossRef]
- Kowalski, S.; Lukasiewicz, M.; Duda-Chodak, A.; Zięć, G. 5-Hydroxymethyl-2-Furfural (HMF)—Heat-induced formation, occurrence in food and biotransformation—A review. Pol. J. Food Nutr. Sci. 2013, 63, 207–225. [Google Scholar] [CrossRef]
- Ma, L.F.; Yan, J.J.; Lane, H.Y.; Jin, L.C.; Qiu, F.J.; Wang, Y.J.; Xi, Z.F.; Shan, W.G.; Zhan, Z.J.; Ying, Y.M. Bioassay-guided isolation of lanostane-type triterpenoids as alpha-glucosidase inhibitors from Ganoderma hainanense. Phytochem. Lett. 2019, 29, 154–159. [Google Scholar] [CrossRef]
- Marchelak, A.; Owczarek, A.; Rutkowska, M.; Michel, P.; Kołodziejczyk-Czepas, J.; Nowak, P.; Olszewska, M.A. New insights into antioxidant activity of Prunus spinosa flowers: Extracts, model polyphenols and their phenolic metabolites in plasma towards multiple in vivo-relevant oxidants. Phytochem. Lett. 2019, 30, 288–295. [Google Scholar] [CrossRef]
- Kołodziejczyk-Czepas, J.; Nowak, P.; Wachowicz, B.; Piechocka, J.; Głowacki, R.; Moniuszko-Szajwaj, B.; Stochmal, A. Antioxidant efficacy of Kalanchoe daigremontiana bufadienolide-rich fraction in blood plasma in vitro. Pharm. Biol. 2016, 54, 3182–3188. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| α-Glucosidase IC50 (µg/mL) | α-Amylase IC50 (µg/mL) | AGEs Formation IC50 (µg/mL) | |
|---|---|---|---|
| Analytes: | |||
| MEF | 15.43 ± 0.84 A | 68.32 ± 2.62 C | 42.94 ± 3.18 B |
| MED | 136.31 ± 4.24 D | 214.13 ± 12.20 F | 96.83 ± 3.41 D |
| DEFF | 18.11 ± 0.42 A | 40.89 ± 2.04 B | 73.61 ± 3.09 C |
| DEFD | 26.82 ± 2.75 A | 86.45 ± 3.71 C | 51.51 ± 3.57 B |
| EAFF | 52.39 ± 2.30 B | 33.47 ± 1.57 B | 75.34 ± 4.42 C |
| EAFD | 128.81 ± 6.29 D | 136.17 ± 10.18 E | 79.79 ± 5.39 C |
| BFF | 90.95 ± 13.00 C | 110.12 ± 15.46 D | 45.50 ± 3.03 B |
| BFD | 181.64 ± 9.23 F | 272.83 ± 4.18 G | 95.53 ± 2.67 D |
| WRF | 50.05 ± 2.10 B | *** | 82.93 ± 2.07 C |
| WRD | 161.06 ± 3.52 E | *** | 97.97 ± 5.62 D |
| Quercetin | * | 37.52 ± 2.32 B | - |
| Quercetin 3-O-glucoside (isoquercitrin) | 209.64 ± 5.64 G | - | 3.09 ± 0.17 A |
| Cyanidin 3-O-glucoside | 635.13 ± 34.43 H | 31.28 ± 1.66 B | 10.90 ± 0.85 A |
| Chlorogenic acid | ** | 81.06 ± 3.94 C | 7.07 ± 0.74 A |
| 5-Hydroxymethylfurfural (HMF) | ** | ** | ** |
| Reference standards: | |||
| Acarbose | 177.07 ± 9.10 E,F | 4.90 ± 0.30 A | - |
| Aminoguanidine | - | - | 78.89 ± 4.95 C |
| NO• (µg/mL) | HOCl (µg/mL) | O2•− (µg/mL) | H2O2 (mg/mL) | HO• (mg/mL) | |
|---|---|---|---|---|---|
| Analytes: | |||||
| MEF | 18.28 ± 0.42 H | 40.04 ± 0.51 F | 73.89 ± 3.58 D | 0.17 ± 0.004 E | 0.44 ± 0.02 E,F |
| MED | 43.05 ± 3.08 I | 100.90 ± 0.69 G | 101.81 ± 3.05 E | 0.57 ± 0.007 J | 0.81 ± 0.02 H |
| DEFF | 5.61 ± 0.13 F | 17.34 ± 0.50 B,C | 29.40 ± 0.88 B | 0.13 ± 0.002 C | 0.33 ± 0.01 D |
| DEFD | 3.85 ± 0.22 D | 18.35 ± 0.52 B,C | 32.70 ± 2.74 B | 0.15 ± 0.002 D | 0.32 ± 0.01 D |
| EAFF | 2.15 ± 0.16 C | 31.48 ± 0.64 D | 45.08 ± 2.96 C | 0.14 ± 0.001 C | 0.38 ± 0.01 D,E |
| EAFD | 4.79 ± 0.39 E | 34.44 ± 0.46 D,E | 52.76 ± 2.82 C | 0.21 ± 0.005 F | 0.33 ± 0.01 D |
| BFF | 6.35 ± 0.41 G | 39.78 ± 0.85 F | 73.52 ± 2.68 D | 0.22 ± 0.002 G | 0.47 ± 0.01 F,G |
| BFD | 43.70 ± 2.49 I | 96.48 ± 5.41 G | 106.88 ± 4.39 E | 0.44 ± 0.008 I | 0.96 ± 0.06 I |
| WRF | 51.76 ± 3.89 J | 38.02 ± 0.88 E,F | 103.47 ± 4.06 E | 0.27 ± 0.002 H | 0.52 ± 0.03 G |
| WRD | 40.21 ± 1.99 I | 153.46 ± 0.83 H | 311.63 ± 7.08 G | 0.72 ± 0.003 K | 1.17 ± 0.06 J |
| Quercetin | 0.48 ± 0.03 A | 2.08 ± 0.14 A | 7.89 ± 0.24 A | 0.008 ± 0.001 A | 0.05 ± 0.002 A |
| Cyanidin 3-O-glucoside | 1.40 ± 0.09 B | 5.62 ± 0.11 A | 10.11 ± 0.33 A | 0.012 ± 0.001 A,B | 0.17 ± 0.002 C |
| Chlorogenic acid | 0.69 ± 0.02 A | 13.82 ± 0.53 B | 4.58 ± 0.29 A | 0.018 ± 0.001 A,B | 0.07 ± 0.001 A,B |
| 5-Hydroxymethylfurfural (HMF) | ** | ** | ** | *** | *** |
| Reference standards: | |||||
| Ascorbic acid | 0.59 ± 0.04 A | 6.32 ± 0.07 A | 5.28 ± 0.25 A | 0.014 ± 0.001 A,B | 0.14 ± 0.005 B,C |
| Trolox | 0.59 ± 0.02 A | 21.49 ± 1.21 C | 135.33 ± 5.59 F | 0.020 ± 0.001 B | 0.13 ± 0.003 B,C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magiera, A.; Kołodziejczyk-Czepas, J.; Skrobacz, K.; Czerwińska, M.E.; Rutkowska, M.; Prokop, A.; Michel, P.; Olszewska, M.A. Valorisation of the Inhibitory Potential of Fresh and Dried Fruit Extracts of Prunus spinosa L. towards Carbohydrate Hydrolysing Enzymes, Protein Glycation, Multiple Oxidants and Oxidative Stress-Induced Changes in Human Plasma Constituents. Pharmaceuticals 2022, 15, 1300. https://doi.org/10.3390/ph15101300
Magiera A, Kołodziejczyk-Czepas J, Skrobacz K, Czerwińska ME, Rutkowska M, Prokop A, Michel P, Olszewska MA. Valorisation of the Inhibitory Potential of Fresh and Dried Fruit Extracts of Prunus spinosa L. towards Carbohydrate Hydrolysing Enzymes, Protein Glycation, Multiple Oxidants and Oxidative Stress-Induced Changes in Human Plasma Constituents. Pharmaceuticals. 2022; 15(10):1300. https://doi.org/10.3390/ph15101300
Chicago/Turabian StyleMagiera, Anna, Joanna Kołodziejczyk-Czepas, Karolina Skrobacz, Monika Ewa Czerwińska, Magdalena Rutkowska, Aleksandra Prokop, Piotr Michel, and Monika Anna Olszewska. 2022. "Valorisation of the Inhibitory Potential of Fresh and Dried Fruit Extracts of Prunus spinosa L. towards Carbohydrate Hydrolysing Enzymes, Protein Glycation, Multiple Oxidants and Oxidative Stress-Induced Changes in Human Plasma Constituents" Pharmaceuticals 15, no. 10: 1300. https://doi.org/10.3390/ph15101300
APA StyleMagiera, A., Kołodziejczyk-Czepas, J., Skrobacz, K., Czerwińska, M. E., Rutkowska, M., Prokop, A., Michel, P., & Olszewska, M. A. (2022). Valorisation of the Inhibitory Potential of Fresh and Dried Fruit Extracts of Prunus spinosa L. towards Carbohydrate Hydrolysing Enzymes, Protein Glycation, Multiple Oxidants and Oxidative Stress-Induced Changes in Human Plasma Constituents. Pharmaceuticals, 15(10), 1300. https://doi.org/10.3390/ph15101300