
Ginkgo biloba Extract Stimulates Adipogenesis in 3T3-L1 Preadipocytes

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
2.1. GbE Treatment Enhanced Cell Viability and Stimulated Cell Proliferation in 3T3-L1 Cells
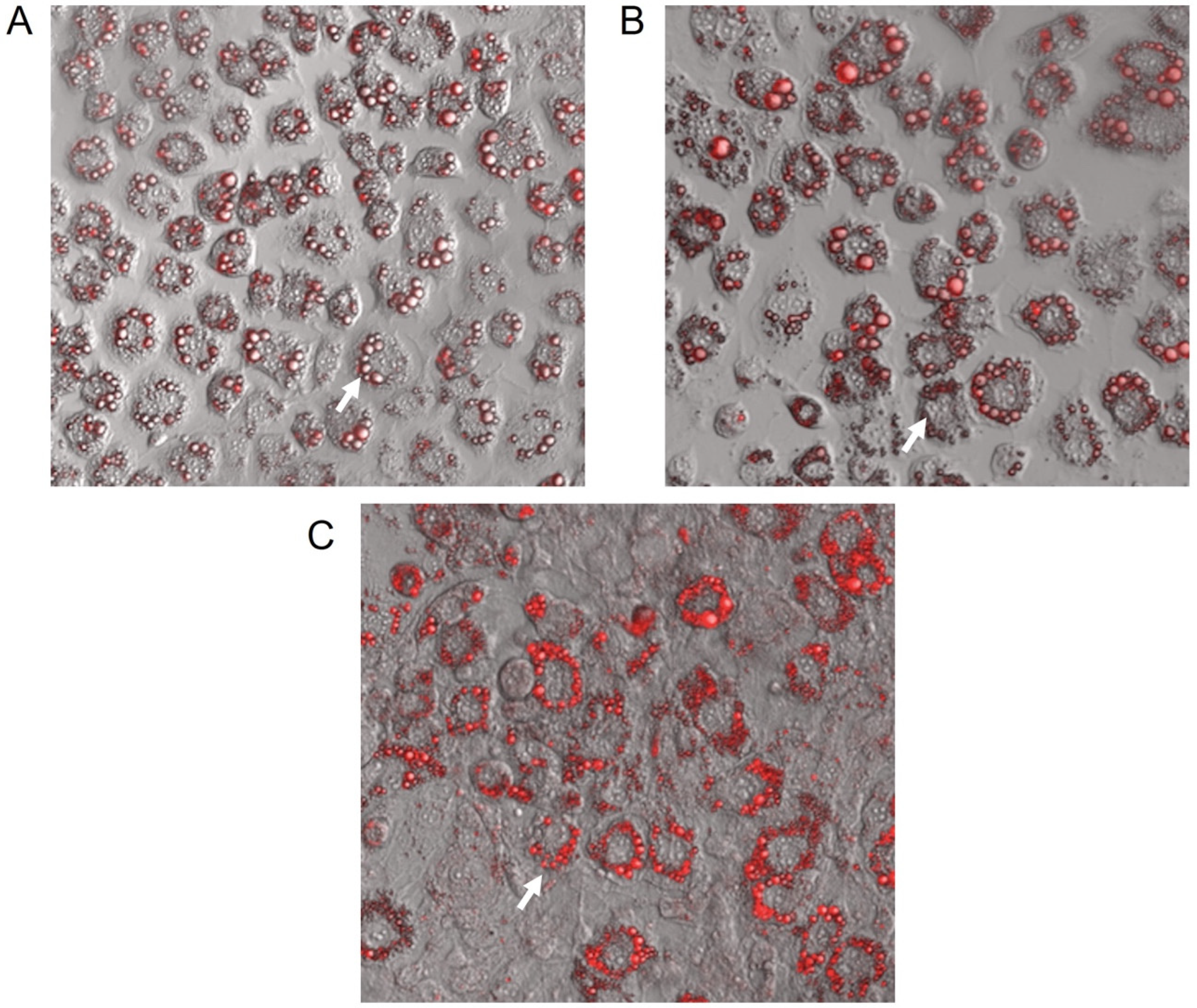
2.2. GbE Increased the Lipid Content in Mature 3T3-L1 Adipocytes
2.3. GbE Increased Gene Expression of the Key Markers of Adipogenesis in 3T3-L1 Cells
2.4. GbE Treatment Accelerated Protein Expression of Specific Adipocytes Proteins in 3T3-L1 Cells during the Differentiation Process
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Ginkgo Biloba Extract Treatment
4.3. Oil Red O Staining
4.4. PCR Array Gene Expression Analysis
4.5. Western Blotting
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sethi, J.K.; Vidal-Puig, A.J. Thematic review series: Adipocyte biology. Adipose tissue function and plasticity orchestrate nutritional adaptation. J. Lipid Res. 2007, 48, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Hammarstedt, A.; Gogg, S.; Hedjazifar, S.; Nerstedt, A.; Smith, U. Impaired Adipogenesis and Dysfunctional Adipose Tissue in Human Hypertrophic Obesity. Physiol. Rev. 2018, 98, 1911–1941. [Google Scholar] [CrossRef] [PubMed]
- Frühbeck, G. Overview of adipose tissue and its role in obesity and metabolic disorders. Methods Mol. Biol. 2008, 456, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Curat, C.A.; Miranville, A.; Sengenès, C.; Diehl, M.; Tonus, C.; Busse, R.; Bouloumié, A. From blood monocytes to adipose tissue-resident macrophages: Induction of diapedesis by human mature adipocytes. Diabetes 2004, 53, 1285–1292. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Heinonen, S.; Jokinen, R.; Rissanen, A.; Pietiläinen, K.H. White adipose tissue mitochondrial metabolism in health and in obesity. Obes. Rev. 2020, 21, e12958. [Google Scholar] [CrossRef]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef]
- McLaughlin, T.; Sherman, A.; Tsao, P.; Gonzalez, O.; Yee, G.; Lamendola, C.; Reaven, G.M.; Cushman, S.W. Enhanced proportion of small adipose cells in insulin-resistant vs insulin-sensitive obese individuals implicates impaired adipogenesis. Diabetologia 2007, 50, 1707–1715. [Google Scholar] [CrossRef]
- Lundgren, M.; Svensson, M.; Lindmark, S.; Renström, F.; Ruge, T.; Eriksson, J.W. Fat cell enlargement is an independent marker of insulin resistance and ‘hyperleptinaemia’. Diabetologia 2007, 50, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Halberg, N.; Khan, T.; Trujillo, M.E.; Wernstedt-Asterholm, I.; Attie, A.D.; Sherwani, S.; Wang, Z.V.; Landskroner-Eiger, S.; Dineen, S.; Magalang, U.J.; et al. Hypoxia-inducible factor 1alpha induces fibrosis and insulin resistance in white adipose tissue. Mol. Cell. Biol. 2009, 29, 4467–4483. [Google Scholar] [CrossRef] [PubMed]
- Mahadevan, S.; Park, Y. Multifaceted therapeutic benefits of Ginkgo biloba L.: Chemistry, efficacy, safety, and uses. J. Food Sci. 2008, 73, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Belwal, T.; Giri, L.; Bahukhandi, A.; Tariq, M.; Kewlani, P.; Bhatt, I.; Rawal, R.S. Ginkgo biloba. In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Silva, A.S., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 241–250. [Google Scholar] [CrossRef]
- Hirata, B.K.S.; Banin, R.M.; Dornellas, A.P.S.; de Andrade, I.S.; Zemdegs, J.C.S.; Caperuto, L.C.; Oyama, L.M.; Ribeiro, E.B.; Telles, M.M. Ginkgo biloba extract improves insulin signaling and attenuates inflammation in retroperitoneal adipose tissue depot of obese rats. Mediat. Inflamm. 2015, 2015, 419106. [Google Scholar] [CrossRef] [PubMed]
- Hirata, B.K.; Pedroso, A.; Machado, M.M.F.; Neto, N.I.; Perestrelo, B.O.; De Sá, R.D.; Alonso-Vale, M.I.C.; Nogueira, F.; Oyama, L.M.; Ribeiro, E.B.; et al. Ginkgo biloba extract modulates the retroperitoneal fat depot proteome and reduces oxidative stress in diet-induced obese rats. Front. Pharmacol. 2019, 10, 686. [Google Scholar] [CrossRef]
- Hirata, B.K.S.; Cruz, M.M.; De Sá, R.D.C.C.; Farias, T.S.M.; Machado, M.M.F.; Bueno, A.A.; Alonso-Vale, M.I.; Telles, M.M. Potential anti-obesogenic effects of Ginkgo biloba observed in epididymal white adipose tissue of obese rats. Front. Endocrinol. 2019, 10, 284. [Google Scholar] [CrossRef]
- Meyer, L.K.; Ciaraldi, T.P.; Henry, R.R.; Wittgrove, A.C.; Phillips, S.A. Adipose tissue depot and cell size dependency of adiponectin synthesis and secretion in human obesity. Adipocyte 2013, 2, 217–226. [Google Scholar] [CrossRef]
- Van Heemst, D. Insulin, IGF-1 and longevity. Aging Dis. 2010, 1, 147–157. [Google Scholar]
- Fu, Y.; Luo, N.; Klein, R.L.; Garvey, W.T. Adiponectin promotes adipocyte differentiation, insulin sensitivity, and lipid accumulation. J. Lipid Res. 2005, 46, 1369–1379. [Google Scholar] [CrossRef]
- Gustafson, B.; Hammarstedt, A.; Hedjazifar, S.; Smith, U. Restricted adipogenesis in hypertrophic obesity: The role of WISP2, WNT, and BMP4. Diabetes 2013, 62, 2997–3004. [Google Scholar] [CrossRef]
- Prospective Studies Collaboration; Whitlock, G.; Lewington, S.; Sherliker, P.; Clarke, R.; Emberson, J.; Halsey, J.; Qizilbash, N.; Collins, R.; Peto, R. Body-mass index and cause-specific mortality in 900,000 adults: Collaborative analyses of 57 prospective studies. Lancet 2009, 37, 1083–1096. [Google Scholar] [CrossRef]
- Trompezinski, S.; Bonneville, M.; Pernet, I.; Denis, A.; Schmitt, D.; Viac, J. Ginkgo biloba extract reduces VEGF and CXCL-8/IL-8 levels in keratinocytes with cumulative effect with epigallocatechin-3-gallate. Arch. Dermatol. Res. 2010, 302, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Cefali, L.C.; Ataide, J.A.; Fernandes, A.R.; Sanchez-Lopez, E.; Sousa, I.M.D.O.; Figueiredo, M.C.; Ruiz, A.L.T.G.; Foglio, M.A.; Mazzola, P.G.; Souto, E.B. Evaluation of in vitro solar protection factor (SPF), antioxidant activity, and cell viability of mixed vegetable extracts from Dirmophandra mollis Benth, Ginkgo biloba L., Ruta graveolens L., and Vitis vinifera L. Plants 2019, 8, 453. [Google Scholar] [CrossRef] [PubMed]
- Wronska, A.; Kmiec, Z. Structural and biochemical characteristics of various white adipose tissue depots. Acta Physiol. 2012, 205, 194–208. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Cheng, J.; Suzuki, M.; Shinohara, Y.; Fujita, A.; Fujimoto, T. Biogenesis of cytoplasmic lipid droplets: From the lipid ester globule in the membrane to the visible structure. Biochim. Biophys. Acta 2009, 1791, 399–407. [Google Scholar] [CrossRef]
- Khan, T.; Muise, E.S.; Iyengar, P.; Wang, Z.; Chandalia, M.; Abate, N.; Zhang, B.B.; Bonaldo, P.; Chua, S.; Scherer, P.E. Metabolic dysregulation and adipose tissue fibrosis: Role of collagen VI. Mol. Cell. Biol. 2009, 29, 1575–1591. [Google Scholar] [CrossRef]
- Laurencikiene, J.; Skurk, T.; Kulyté, A.; Hedén, P.; Åström, G.; Sjölin, E.; Rydén, M.; Hauner, H.; Arner, P. Regulation of lipolysis in small and large fat cells of the same subject. J. Clin. Endocrinol. Metab. 2011, 96, E2045–E2049. [Google Scholar] [CrossRef]
- Bambace, C.; Telesca, M.; Zoico, E.; Sepe, A.; Olioso, D.; Rossi, A.; Corzato, F.; Di Francesco, V.; Mazzucco, A.; Santini, F.; et al. Adiponectin gene expression and adipocyte diameter: A comparison between epicardial and subcutaneous adipose tissue in men. Cardiovasc. Pathol. 2011, 20, e153–e156. [Google Scholar] [CrossRef]
- Trayhurn, P. Endocrine and signalling role of adipose tissue: New perspectives on fat. Acta Physiol. Scand. 2005, 184, 285–293. [Google Scholar] [CrossRef]
- Cao, Z.; Umek, R.M.; McKnight, S.L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991, 5, 1538–1552. [Google Scholar] [CrossRef]
- Audano, M.; Pedretti, S.; Caruso, D.; Crestani, M.; De Fabiani, E.; Mitro, N. Regulatory mechanisms of the early phase of white adipocyte differentiation: An overview. Cell. Mol. Life Sci. 2022, 79, 139. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Jakab, J.; Miškić, B.; Mikšić, Š.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a Potential Anti-Obesity Target: A Review of Pharmacological Treatment and Natural Products. Diabetes Metab. Syndr. Obes. 2021, 14, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Dixon, T.M.; Daniel, K.W.; Farmer, S.R.; Collins, S. CCAAT/enhancer-binding protein alpha is required for transcription of the beta 3-adrenergic receptor gene during adipogenesis. J. Biol. Chem. 2001, 276, 722–728. [Google Scholar] [CrossRef]
- Richard, J.E.; López-Ferreras, L.; Chanclón, B.; Eerola, K.; Micallef, P.; Skibicka, K.P.; Wernstedt Asterholm, I. CNS β3-adrenergic receptor activation regulates feeding behavior, white fat browning, and body weight. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E344–E358. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Lu, D.; Li, J.; Liu, M.; He, Y.; Williams, B.; Li, J.; Yang, T. Ginkgolic acid, a sumoylation inhibitor, promotes adipocyte commitment but suppresses adipocyte terminal differentiation of mouse bone marrow stromal cells. Sci. Rep. 2018, 8, 2545. [Google Scholar] [CrossRef]
- Liou, C.J.; Lai, X.Y.; Chen, Y.L.; Wang, C.L.; Wei, C.H.; Huang, W.C. Ginkgolide C Suppresses Adipogenesis in 3T3-L1 Adipocytes via the AMPK Signaling Pathway. Evid.-Based Complement. Altern. Med. 2015, 2015, 298635. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Kim, S.; Park, J.; Ha, T. The anti-obesity effect of quercetin is mediated by the AMPK and MAPK signaling pathways. Biochem. Biophys. Res. Commun. 2008, 373, 545–549. [Google Scholar] [CrossRef]
- Bu, S.; Yuan, C.Y.; Xue, Q.; Chen, Y.; Cao, F. Bilobalide Suppresses Adipogenesis in 3T3-L1 Adipocytes via the AMPK Signaling Pathway. Molecules 2019, 24, 3503. [Google Scholar] [CrossRef]
- Cho, Y.-L.; Park, J.-G.; Kang, H.J.; Kim, W.; Cho, M.J.; Jang, J.-H.; Kwon, M.-G.; Kim, S.; Lee, S.-H.; Lee, J.; et al. Ginkgetin, a biflavone from Ginkgo biloba leaves, prevents adipogenesis through STAT5-mediated PPARγ and C/EBPα regulation. Pharmacol. Res. 2019, 139, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Ntambi, J.M.; Young-Cheul, K. Adipocyte Differentiation and Gene Expression. J. Nutr. 2000, 130, 3122S–3126S. [Google Scholar] [CrossRef]
- Cruz, M.M.; Lopes, A.B.; Crisma, A.R.; de Sá, R.C.C.; Kuwabara, W.M.T.; Curi, R.; de Andrade, P.B.M.; Alonso-Vale, M.I.C. Palmitoleic acid (16, 1n7) increases oxygen consumption, fatty acid oxidation and ATP content in white adipocytes. Lipids Health Dis. 2018, 17, 55. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.M.F.; Banin, R.M.; Thomaz, F.M.; de Andrade, I.S.; Boldarine, V.T.; de Souza Figueiredo, J.; Hirata, B.K.S.; Oyama, L.M.; Lago, J.H.G.; Ribeiro, E.B.; et al. Ginkgo biloba Extract (GbE) restores serotonin and leptin receptor levels and plays an antioxidative role in the hippocampus of ovariectomized rats. Mol. Neurobiol. 2021, 58, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Fold Regulation | p-Value | Pathway Related |
|---|---|---|---|
| Up-Regulated | |||
| Adipoq | 8.78 | 0.002505 | Adipokines |
| Acaca | 27.36 | 0.047027 | Lipases and lipogenic enzymes |
| Gpd1 | 26.25 | 0.001161 | Lipases and lipogenic enzymes |
| Lipe | 5.01 | 0.006739 | Lipases and lipogenic enzymes |
| Scd1 | 113.17 | 0.021186 | Lipases and lipogenic enzymes |
| Pck1 | 743.06 | 0.015354 | Lipases and lipogenic enzymes |
| Fasn | 698.12 | 0.016781 | Lipases and lipogenic enzymes |
| Cebpa | 19.84 | 0.00061 | Pro-adipogenesis |
| Pparg | 2002.15 | 0.026924 | Pro-adipogenesis |
| Srebf1 | 12.84 | 0.027642 | Pro-adipogenesis |
| Fabp4 | 7.31 | 0.047331 | Pro-adipogenesis |
| Plin1 | 5.49 | 0.000060 | Pro-adipogenesis |
| Fgf10 | 7.52 | 0.013401 | Pro-adipogenesis |
| Slc2a4 | 125.57 | 0.015493 | Pro-adipogenesis |
| Adrb2 | 7.28 | 0.035326 | Anti-adipogenesis |
| Ncor2 | 113.96 | 0.021250 | Anti-adipogenesis |
| Sirt1 | 281.57 | 0.020575 | Anti-adipogenesis |
| Klf2 | 7.99 | 0.033226 | Anti-adipogenesis |
| Cpt1b | 34.31 | 0.024551 | Pro-browning, fatty acid oxidation, and thermogenesis |
| Elovl3 | 43.33 | 0.017401 | Pro-browning, fatty acid oxidation, and thermogenesis |
| Ppara | 7.58 | 0.031916 | Pro-browning, fatty acid oxidation, and thermogenesis |
| Tfam | 601.46 | 0.019365 | Pro-browning, fatty acid oxidation, and thermogenesis |
| Nr1h3 | 3.92 | 0.000593 | Anti-browning |
| Rb1 | 25.76 | 0.021335 | Anti-browning |
| Cxcl10 | 20.78 | 0.018731 | Cytokines, growth factors, and signal transduction |
| Il4 | 14.05 | 0.025102 | Cytokines, growth factors, and signal transduction |
| Il6 | 8.4 | 0.037268 | Cytokines, growth factors, and signal transduction |
| Il13 | 90.34 | 0.015970 | Cytokines, growth factors, and signal transduction |
| Tgfb1 | 416.55 | 0.015650 | Cytokines, growth factors, and signal transduction |
| Irs2 | 3.18 | 0.029002 | Cytokines, growth factors, and signal transduction |
| Ptpn1 | 2.22 | 0.012208 | Cytokines, growth factors, and signal transduction |
| Mapk8 | 29.8 | 0.018482 | Cytokines, growth factors, and signal transduction |
| Nfkb1 | 1043.59 | 0.016021 | Cytokines, growth factors, and signal transduction |
| Irf4 | 6.65 | 0.034403 | Cytokines, growth factors, and signal transduction |
| Retnla | 2153.3 | 0.021842 | Cytokines, growth factors, and signal transduction |
| Down-regulated | |||
| Cdkn1b | −2.0 | 0.007710 | Anti-adipogenesis |
| Ddit3 | −5.53 | 0.019482 | Anti-adipogenesis |
| Cd68 | −39.47 | 0.000351 | Cytokines, growth factors, and signal transduction |
| Gapdh | −3.76 | 0.018267 | Endogenous |
| Pathways | Genes |
|---|---|
| Adipokines | Adipoq (Acrp30), Cfd (Adipisin), Lep, Retn |
| Lipases and lipogenic enzymes | Acaca (Acc1), Gpd, Lipe (HSL), Scd1, Lpl, Pnpla2 (Atgl), Lipin 1, Pck1, Fasn |
| Pro-adipogenesis | Cebpa, Cebpb, Cebpd, Pparg, Srebf1, Fabp4(aP2), Pilin1, Fgf2 (bFGF), Fgf10, Jun, Lmna, Sfrp1, Slc2a4 (Glut4), Klf15, Klf4 |
| Anti-adipogenesis | Adrb2, Cdkn1a, Cdkn1b, Ddit3, Dlk1(Pref1), Foxo1, Ncor2, Shh, Sirt1, Wnt1, Wnt3a, Gata2, Klf |
| Pro-browning, fatty acid oxidation, and thermogenesis | Bmp7, Cidea, Cpt1b, Creb1, Dio2, Elovl3, Foxc2, Mapk14 (p38alpha), Nrf1, Ppara, Ppard, Ppargc1a, Ppargc1b, Prdm16, Sirt3, Src, Tbx1, Tfam, Ucp1, Wnt5a |
| Anti-browning | Ncoa2, Nr1h3, Rb1, Wnt10b |
| Adipokines receptors | Lepr, Adipor2, Adrb1 |
| Cytokines, growth factors, and signal transduction | Ccl2 (MCP1), Cxcl10, Ifng, Il1b, Il4, Il6, Il10, Il12b, Il13, Tgfb1, Tnf, Insr, Irs1, Irs2, Akt2, Ptpn1 (PTP1B), Ikbkb (IKKbeta), Mapk8 (JNK1), Nfkb1, Pik3r1 (p85alpha), Irf4, Retnla, Cd68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomaz, F.M.; de Jesus Simão, J.; da Silva, V.S.; Machado, M.M.F.; Oyama, L.M.; Ribeiro, E.B.; Alonso Vale, M.I.C.; Telles, M.M. Ginkgo biloba Extract Stimulates Adipogenesis in 3T3-L1 Preadipocytes. Pharmaceuticals 2022, 15, 1294. https://doi.org/10.3390/ph15101294
Thomaz FM, de Jesus Simão J, da Silva VS, Machado MMF, Oyama LM, Ribeiro EB, Alonso Vale MIC, Telles MM. Ginkgo biloba Extract Stimulates Adipogenesis in 3T3-L1 Preadipocytes. Pharmaceuticals. 2022; 15(10):1294. https://doi.org/10.3390/ph15101294
Chicago/Turabian StyleThomaz, Fernanda Malanconi, Jussara de Jesus Simão, Viviane Simões da Silva, Meira Maria Forcelini Machado, Lila Missae Oyama, Eliane Beraldi Ribeiro, Maria Isabel Cardoso Alonso Vale, and Monica Marques Telles. 2022. "Ginkgo biloba Extract Stimulates Adipogenesis in 3T3-L1 Preadipocytes" Pharmaceuticals 15, no. 10: 1294. https://doi.org/10.3390/ph15101294
APA StyleThomaz, F. M., de Jesus Simão, J., da Silva, V. S., Machado, M. M. F., Oyama, L. M., Ribeiro, E. B., Alonso Vale, M. I. C., & Telles, M. M. (2022). Ginkgo biloba Extract Stimulates Adipogenesis in 3T3-L1 Preadipocytes. Pharmaceuticals, 15(10), 1294. https://doi.org/10.3390/ph15101294