
Benzydamine Reverses TMexCD-TOprJ-Mediated High-Level Tigecycline Resistance in Gram-Negative Bacteria


Abstract
:1. Introduction
2. Results
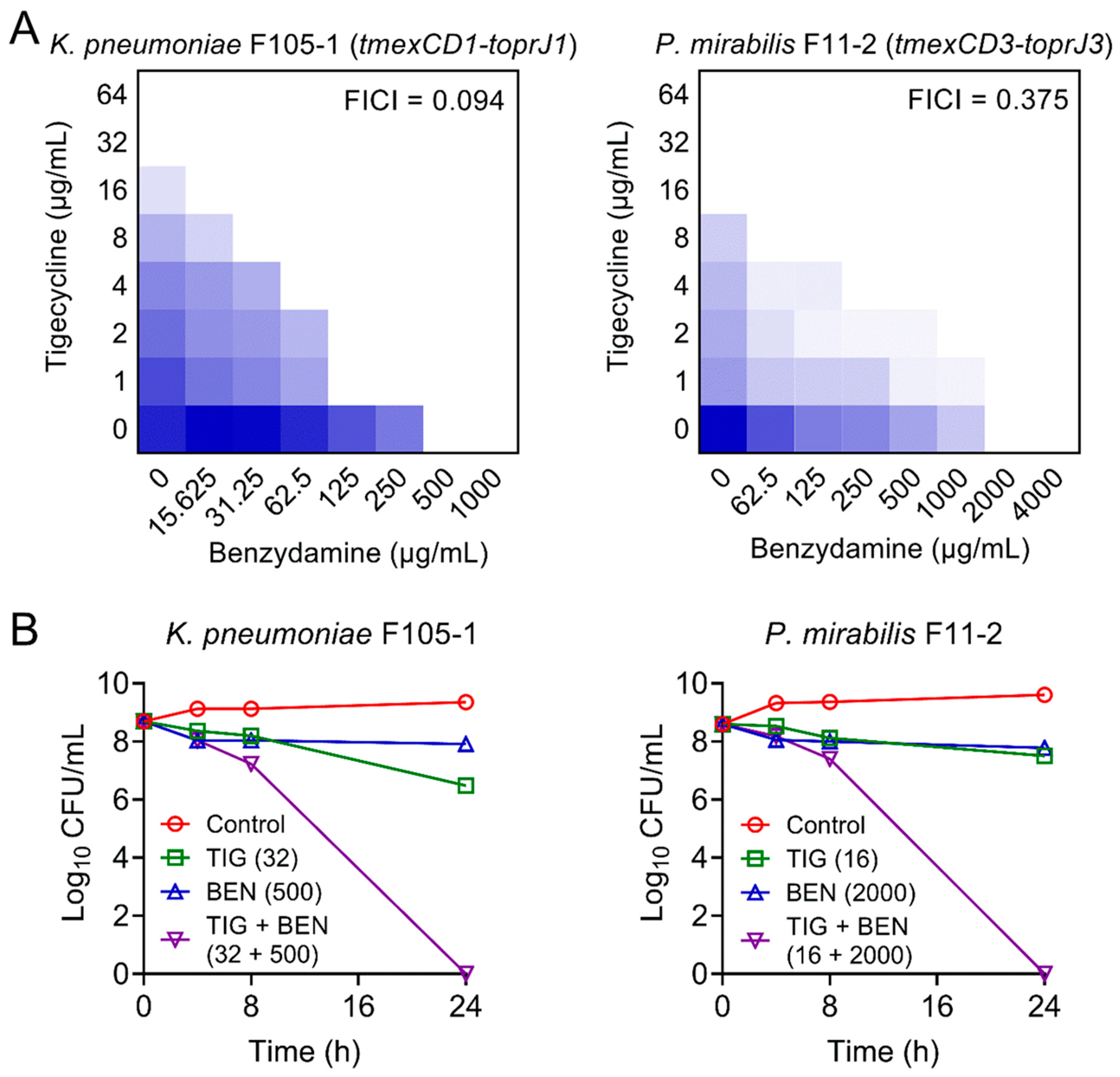
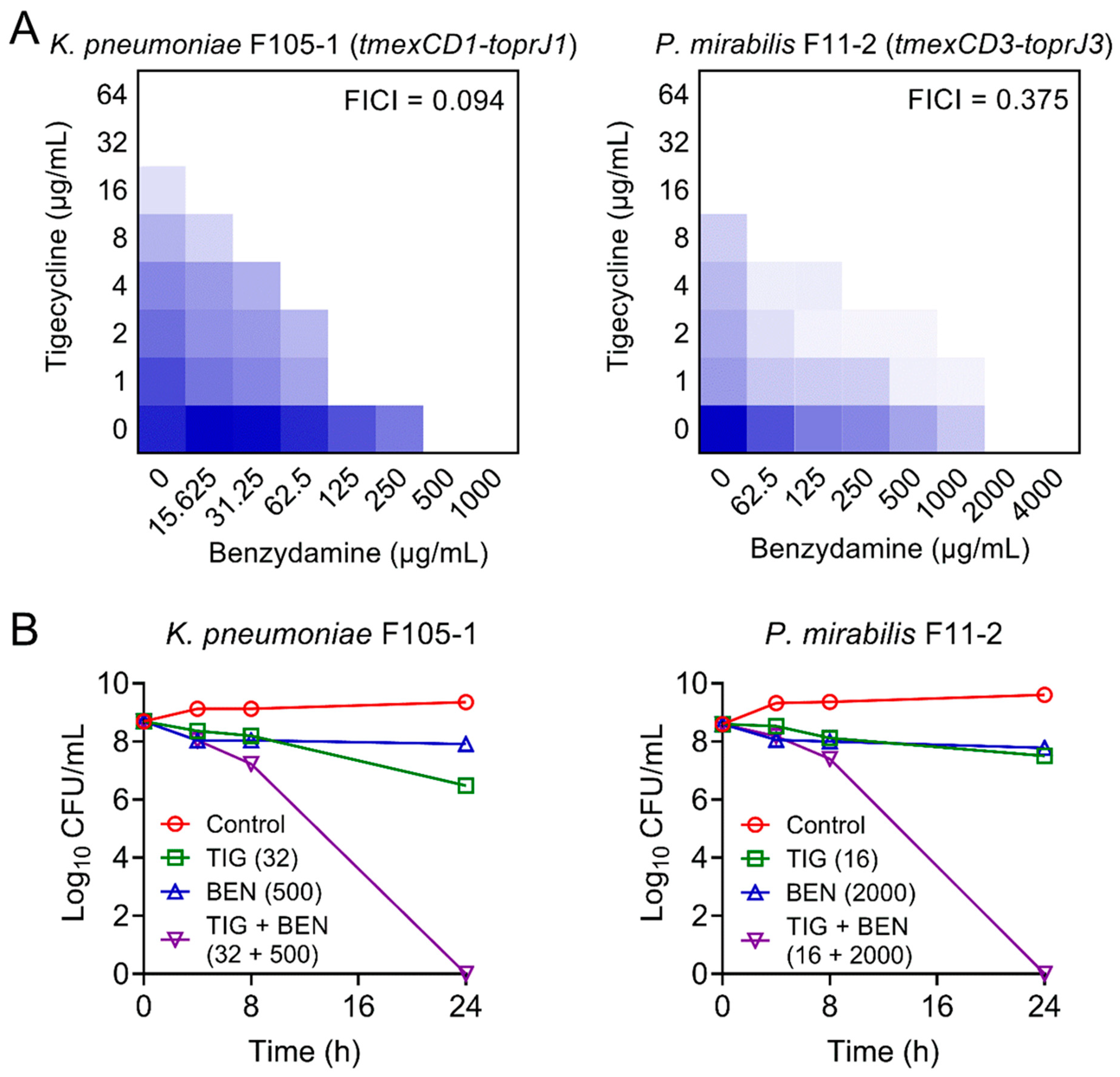
2.1. Synergistic Activity between Benzydamine and Tigecycline against TMexCD-TOprJ-Positive Bacteria
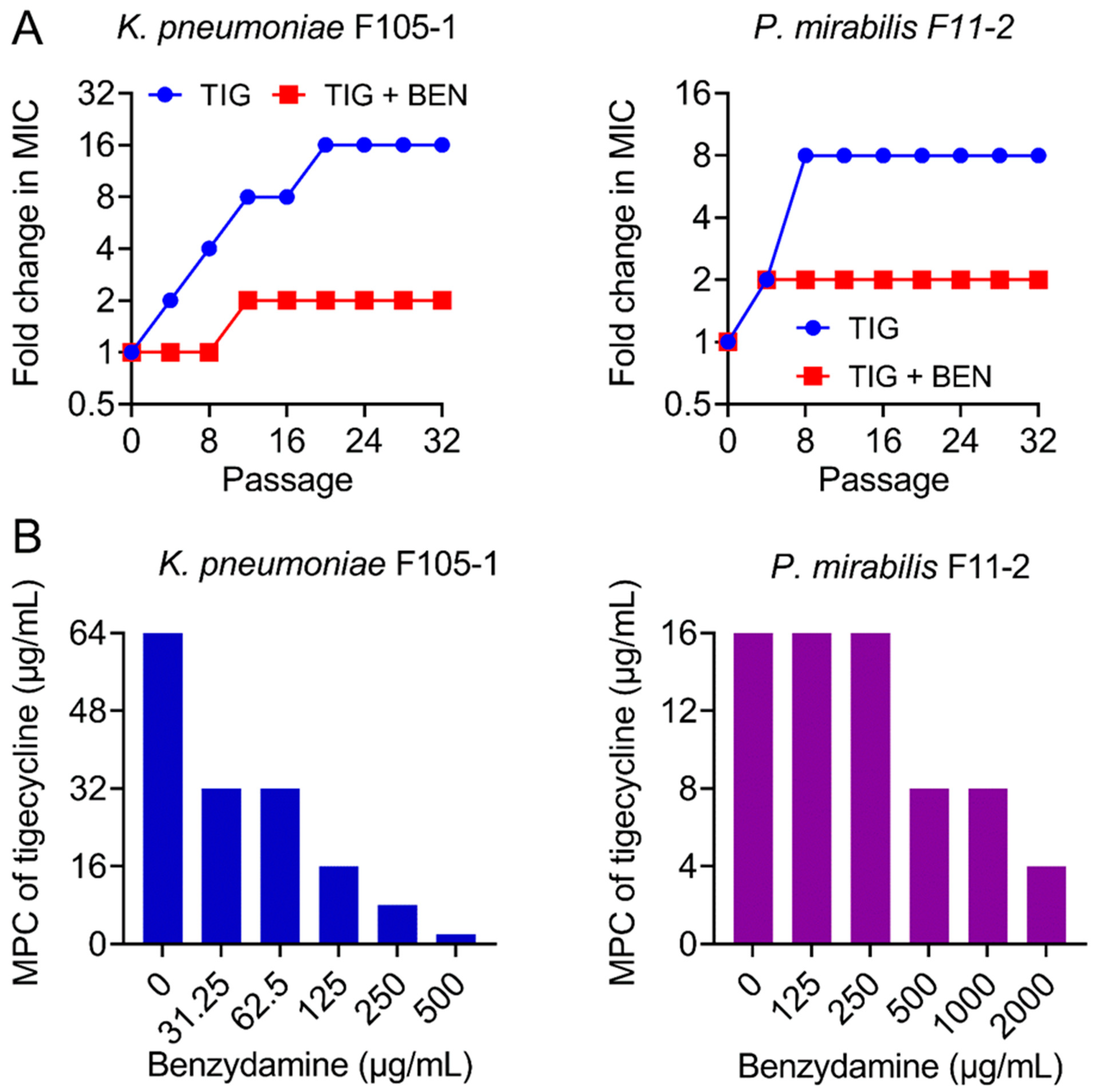
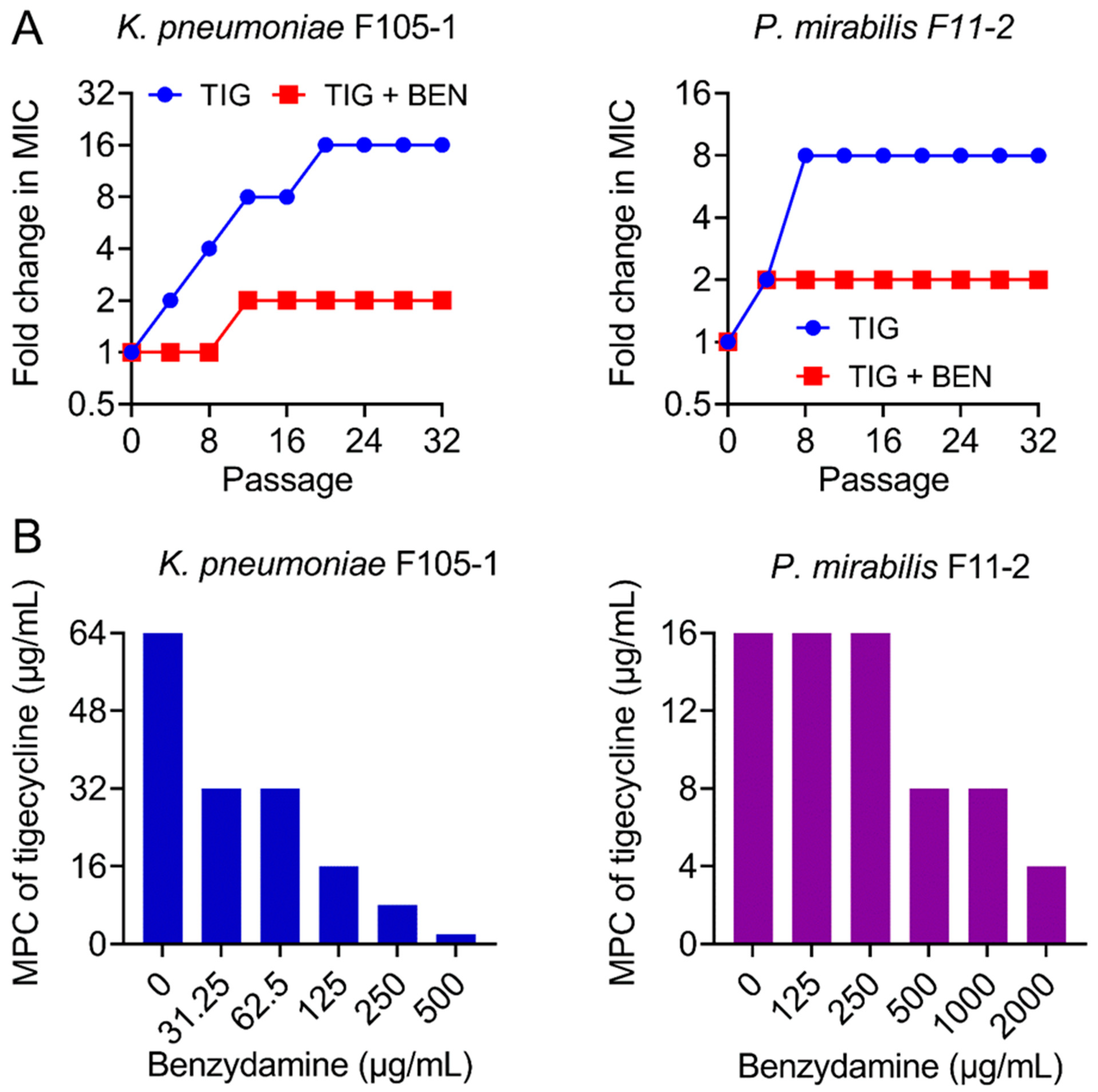
2.2. BEN Prevents the Evolution of Tigecycline Resistance
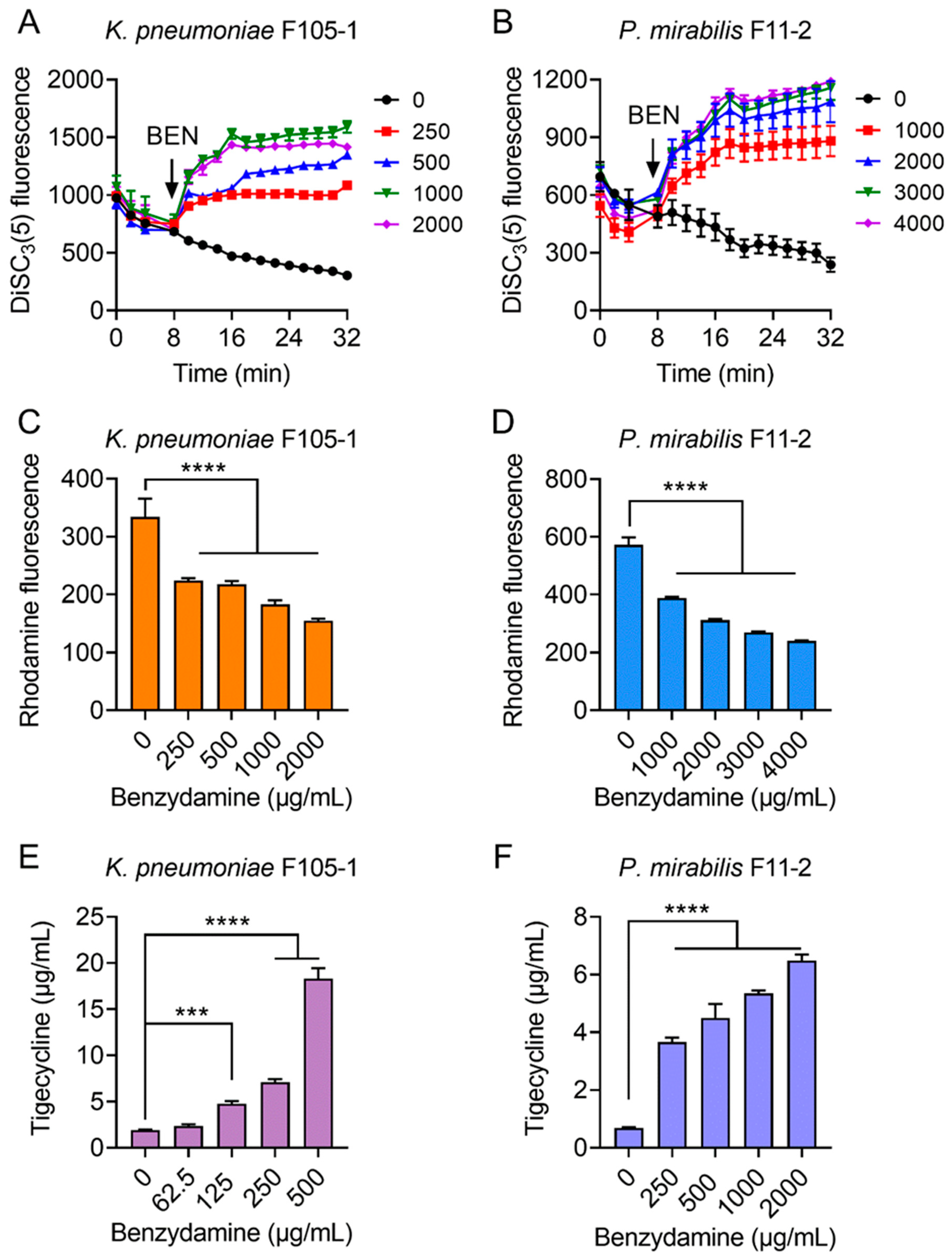
2.3. BEN Deprives the Function of Efflux Pump by Dissipating Proton Motive Force
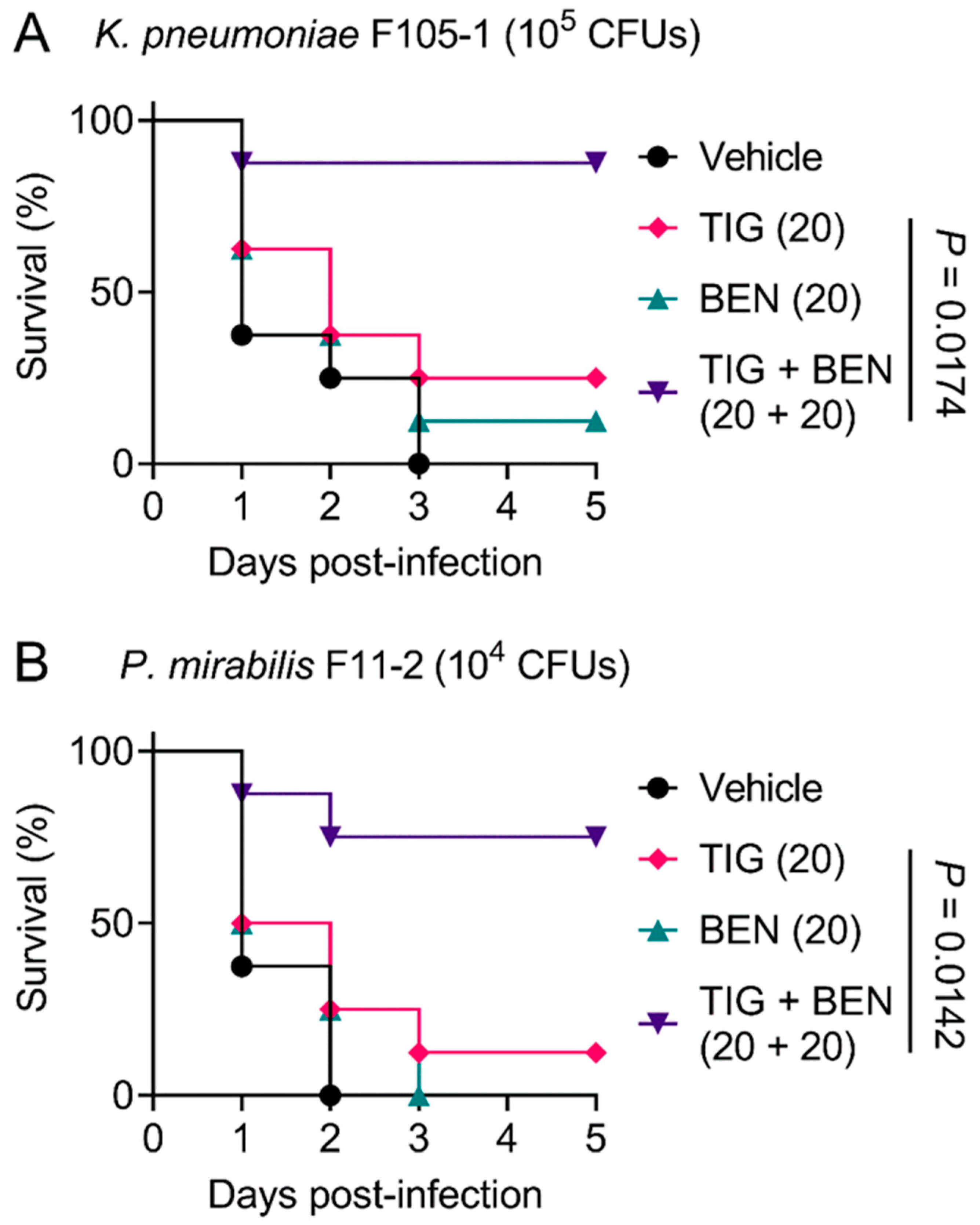
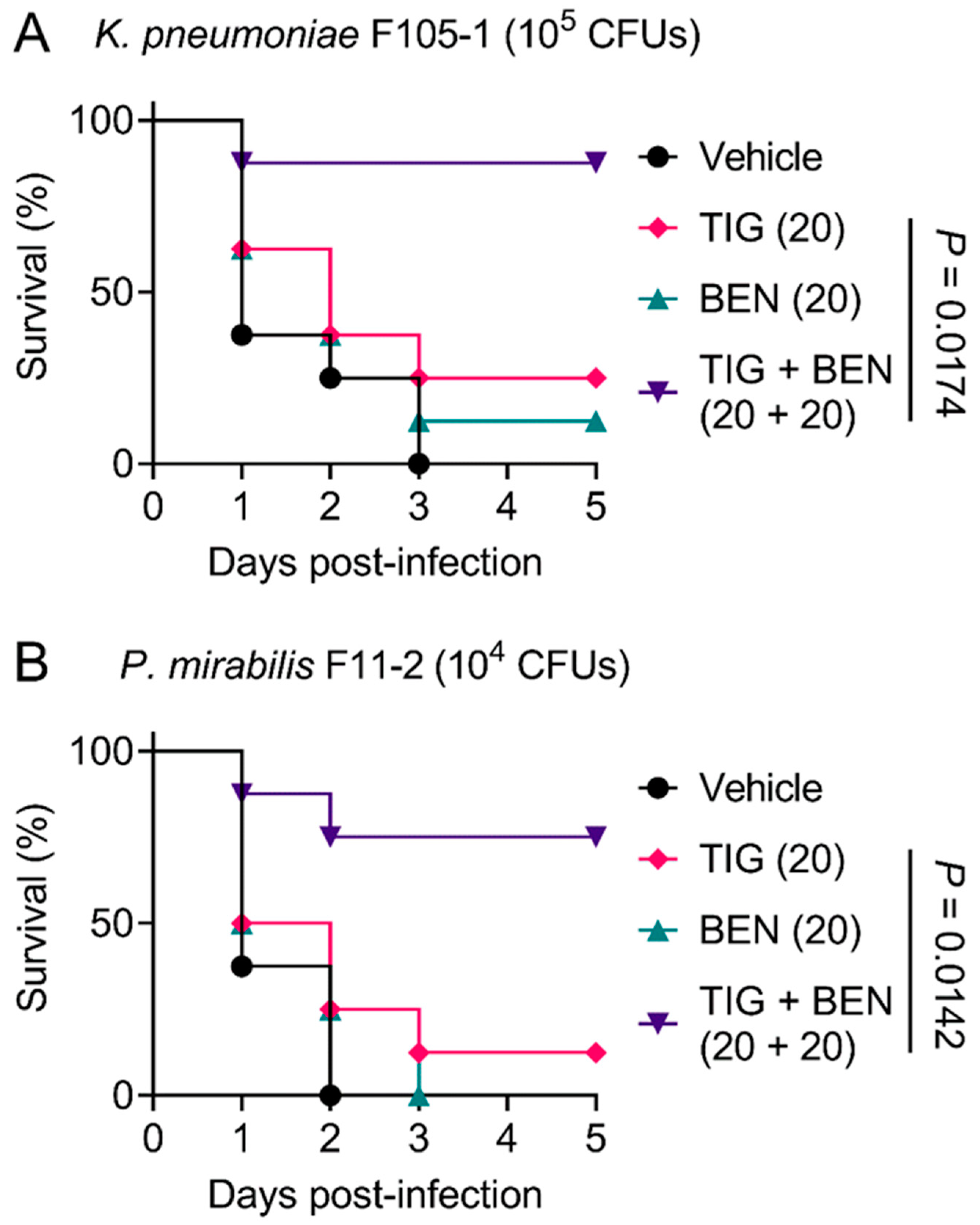
2.4. BEN Restores In Vivo Efficacy of Tigecycline
3. Discussion
4. Materials and Methods
4.1. Minimum Inhibitory Concentration (MIC) Assays
4.2. Checkerboard Assays
4.3. Time-Dependent Killing Studies
4.4. Resistance Development Assessment
4.5. Mutant Prevention Concentration (MPC) Determination
4.6. Proton Motive Force Assay
4.7. Efflux Pump Assay
4.8. Tigecycline Accumulation Analysis
4.9. Galleria mellonella Infection Model
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yelin, I.; Kishony, R. Antibiotic resistance. Cell 2018, 172, 1136.e1. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tong, Z.; Shi, J.; Jia, Y.; Yang, K.; Wang, Z. Correlation between exogenous compounds and the horizontal transfer of plasmid-borne antibiotic resistance genes. Microorganisms 2020, 8, 1211. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Di Pilato, V.; Doi, Y.; Feldgarden, M.; Haft, D.H.; Klimke, W.; Kumar-Singh, S.; Liu, J.-H.; Malhotra-Kumar, S.; Prasad, A.; et al. Proposal for assignment of allele numbers for mobile colistin resistance (mcr) genes. J. Antimicrob. Chemother. 2018, 73, 2625–2630. [Google Scholar] [CrossRef] [PubMed]
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and other enterobacteriaceae: An evolving crisis of global dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef] [Green Version]
- Pankey, G.A. Tigecycline. J. Antimicrob. Chemother. 2005, 56, 470–480. [Google Scholar] [CrossRef]
- Sheu, C.-C.; Chang, Y.-T.; Lin, S.-Y.; Chen, Y.-H.; Hsueh, P.-R. Infections caused by carbapenem-resistant enterobacteriaceae: An update on therapeutic options. Front. Microbiol. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Wang, R.; Liu, D.; Walsh, T.; Zhang, R.; Lv, Y.; Ke, Y.; Ji, Q.; Wei, R.; Liu, Z.; et al. Emergence of plasmid-mediated high-level tigecycline resistance genes in animals and humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef]
- Lv, L.C.; Wan, M.; Wang, C.Z.; Gao, X.; Yang, Q.W.; Partridge, S.R.; Wang, Y.; Zong, Z.Y.; Doi, Y.H.; Shen, J.Z.; et al. Emergence of a plasmid-encoded resistance-nodulation-division efflux pump conferring resistance to multiple drugs, including tigecycline, in Klebsiella pneumoniae. mBio 2020, 11, e02930-19. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Gao, H.; Liu, Y.; Jin, L.; Wang, R.; Wang, X.; Wang, Q.; Yin, Y.; Zhang, Y.; Wang, H. Co-existence of a novel plasmid-mediated efflux pump with colistin resistance gene mcr in one plasmid confers transferable multidrug resistance in Klebsiella pneumoniae. Emerg. Microbes Infect. 2020, 9, 1102–1113. [Google Scholar] [CrossRef]
- Fang, L.; Chen, C.; Cui, C.; Li, X.; Zhang, Y.; Liao, X.; Sun, J.; Liu, Y. Emerging high-level tigecycline resistance: Novel tetracycline destructases spread via the mobile tet(X). BioEssays 2020, 42, e2000014. [Google Scholar] [CrossRef]
- Park, J.; Gasparrini, A.J.; Reck, M.R.; Symister, C.T.; Elliott, J.L.; Vogel, J.P.; Wencewicz, T.A.; Dantas, G.; Tolia, N.H. Plasticity, dynamics, and inhibition of emerging tetracycline resistance enzymes. Nat. Chem. Biol. 2017, 13, 730–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macnair, C.R.; Stokes, J.M.; Carfrae, L.A.; Fiebig-Comyn, A.A.; Coombes, B.K.; Mulvey, M.R.; Brown, E.D. Overcoming mcr-1 mediated colistin resistance with colistin in combination with other antibiotics. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, D.; Wang-Kan, X.; Neuberger, A.; Van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Wang, C.-Z.; Gao, X.; Yang, Q.-W.; Lv, L.-C.; Wan, M.; Yang, J.; Cai, Z.-P.; Liu, J.-H. A novel transferable resistance-nodulation-division pump gene cluster, tmexCD2-toprJ2, confers tigecycline resistance in Raoultella ornithinolytica. Antimicrob. Agents Chemother. 2021, 65, e02229-20. [Google Scholar] [CrossRef]
- Wang, Q.; Peng, K.; Liu, Y.; Xiao, X.; Wang, Z.; Li, R. Characterization of TMexCD3-TOprJ3, an RND-type efflux system conferring resistance to tigecycline in Proteus mirabilis, and its associated integrative conjugative element. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Genet. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Liu, Y.; Li, R.; Xiao, X.; Wang, Z. Antibiotic adjuvants: An alternative approach to overcome multi-drug resistant Gram-negative bacteria. Crit. Rev. Microbiol. 2019, 45, 301–314. [Google Scholar] [CrossRef]
- Liu, Y.; Jia, Y.; Yang, K.; Li, R.; Xiao, X.; Zhu, K.; Wang, Z. Metformin restores tetracyclines susceptibility against multidrug resistant bacteria. Adv. Sci. 2020, 7, 1902227. [Google Scholar] [CrossRef]
- Liu, Y.; Jia, Y.; Yang, K.; Li, R.; Xiao, X.; Wang, Z. Anti-HIV agent azidothymidine decreases Tet(X)-mediated bacterial resistance to tigecycline in Escherichia coli. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuriyama, A.; Aga, M.; Maeda, H. Topical benzydamine hydrochloride for prevention of postoperative sore throat in adults undergoing tracheal intubation for elective surgery: A systematic review and meta-analysis. Anaesthesia 2018, 73, 889–900. [Google Scholar] [CrossRef] [Green Version]
- Son, H.S.; Lee, J.; Lee, H.I.; Kim, N.; Jo, Y.-J.; Lee, G.-R.; Hong, S.-E.; Kwon, M.; Kim, N.Y.; Kim, H.J.; et al. Benzydamine inhibits osteoclast differentiation and bone resorption via down-regulation of interleukin-1 expression. Acta Pharm. Sin. B 2020, 10, 462–474. [Google Scholar] [CrossRef]
- Yang, X.; Ye, L.; Chan, E.W.-C.; Zhang, R.; Chen, S. Characterization of an IncFIB/IncHI1B plasmid encoding efflux pump TMexCD1-TOprJ1 in a clinical tigecycline- and carbapenem-resistant Klebsiella pneumoniae strain. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Wan, M.; Gao, X.; Lv, L.; Cai, Z.; Liu, J.-H. IS 26 mediates the acquisition of tigecycline resistance gene cluster tmexCD1-toprJ1 by IncHI1B-FIB plasmids in Klebsiella pneumoniae and Klebsiella quasipneumoniae from food market sewage. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Liu, Y.; Tong, Z.; Shi, J.; Li, R.; Upton, M.; Wang, Z. Drug repurposing for next-generation combination therapies against multidrug-resistant bacteria. Theranostics 2021, 11, 4910–4928. [Google Scholar] [CrossRef]
- Gianvecchio, C.; Lozano, N.A.; Henderson, C.; Kalhori, P.; Bullivant, A.; Valencia, A.; Su, L.; Bello, G.; Wong, M.; Cook, E.; et al. Variation in mutant prevention concentrations. Front. Microbiol. 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Maier, E.; Benz, R.; Hancock, R. Mechanism of interaction of different classes of cationic antimicrobial peptides with planar bilayers and with the cytoplasmic membrane of Escherichia coli. Biochemistry 1999, 38, 7235–7242. [Google Scholar] [CrossRef] [PubMed]
- Stokes, J.M.; Yang, K.; Swanson, K.; Jin, W.; Cubillos-Ruiz, A.; Donghia, N.M.; Macnair, C.R.; French, S.; Carfrae, L.A.; Bloom-Ackermann, Z.; et al. A deep learning approach to antibiotic discovery. Cell 2020, 180, 688–702. [Google Scholar] [CrossRef] [Green Version]
- Domenech, A.; Brochado, A.R.; Sender, V.; Hentrich, K.; Henriques-Normark, B.; Typas, A.; Veening, J.-W. Proton motive force disruptors block bacterial competence and horizontal gene transfer. Cell Host Microbe 2020, 27, 544–555.e3. [Google Scholar] [CrossRef] [PubMed]
- Farha, M.A.; Verschoor, C.P.; Bowdish, D.; Brown, E.D. Collapsing the proton motive force to identify synergistic combinations against Staphylococcus aureus. Chem. Biol. 2013, 20, 1168–1178. [Google Scholar] [CrossRef] [Green Version]
- Lederman, E.R.; Crum, N.F. Pyogenic liver abscess with a focus on Klebsiella pneumoniae as a primary pathogen: An emerging disease with unique clinical characteristics. Am. J. Gastroenterol. 2005, 100, 322–331. [Google Scholar] [CrossRef]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Pieren, M.; Tigges, M. Adjuvant strategies for potentiation of antibiotics to overcome antimicrobial resistance. Curr. Opin. Pharmacol. 2012, 12, 551–555. [Google Scholar] [CrossRef]
- Domalaon, R.; De Silva, P.M.; Kumar, A.; Zhanel, G.G.; Schweizer, F. The Anthelmintic Drug Niclosamide synergizes with colistin and reverses colistin resistance in Gram-negative bacilli. Antimicrob. Agents Chemother. 2019, 63, e02574-18. [Google Scholar] [CrossRef] [Green Version]
- Maisuria, V.B.; Okshevsky, M.; Déziel, E.; Tufenkji, N. Proanthocyanidin interferes with intrinsic antibiotic resistance mechanisms of Gram-negative bacteria. Adv. Sci. 2019, 6, 1802333. [Google Scholar] [CrossRef]
- Minrovic, B.M.; Jung, D.; Melander, R.J.; Melander, C. New class of adjuvants enables lower dosing of colistin against Acinetobacter baumannii. ACS Infect. Dis. 2018, 4, 1368–1376. [Google Scholar] [CrossRef]
- Hubble, V.B.; Hubbard, B.A.; Minrovic, B.M.; Melander, R.J.; Melander, C. Using small-molecule adjuvants to repurpose azithromycin for use against Pseudomonas aeruginosa. ACS Infect. Dis. 2019, 5, 141–151. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev. 1966, 41, 445–501. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Song, M.; Liu, Y.; Li, T.; Liu, X.; Hao, Z.; Ding, S.; Panichayupakaranant, P.; Zhu, K.; Shen, J. Plant natural flavonoids against multidrug resistant pathogens. Adv. Sci. 2021, 8, 2100749. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, Q.; Wang, R.; Wang, H.; Wong, Y.-T.; Wang, M.; Hao, Q.; Yan, A.; Kao, R.Y.-T.; Ho, P.-L.; et al. Resensitizing carbapenem- and colistin-resistant bacteria to antibiotics using auranofin. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Taber, H.W.; Mueller, J.P.; Miller, P.F.; Arrow, A.S. Bacterial uptake of aminoglycoside antibiotics. Microbiol. Rev. 1987, 51, 439–457. [Google Scholar] [CrossRef]
- Forster, S.; Thumser, A.E.; Hood, S.R.; Plant, N. Characterization of Rhodamine-123 as a tracer dye for use in in vitro drug transport assays. PLoS ONE 2012, 7, e33253. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wang, T.; Wang, X.; Cheng, X.; Dong, H.; Wang, Y.; Zheng, X.; Zhou, L.; Xing, J.; Dong, Y. Quantitative analysis and pharmacokinetics study of tigecycline in human serum using a validated sensitive liquid chromatography with tandem mass spectrometry method. J. Sep. Sci. 2014, 37, 1396–1403. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (μg/mL) a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Tetracyclines | ||||||||||
| MEM | CIP | CL | MIN | OXY | DOX | TET | TIG | TIG b | BEN | |
| K. pneumoniae | ||||||||||
| RGB 7-1 | ≤0.25 | 16 | ≤0.25 | 128 | >256 | 64 | >256 | 32 | 0.125 (256) | 500 |
| RGF 85-1 | ≤0.25 | 8 | ≤0.25 | 64 | >128 | 64 | >128 | 32 | 4 (8) | 500 |
| RGF 105-1 | ≤0.25 | 8 | ≤0.25 | 256 | >256 | 64 | >256 | 32 | 0.125 (256) | 500 |
| RGF 140-1 | ≤0.25 | 8 | ≤0.25 | 128 | >128 | 128 | >128 | 32 | 8 (4) | 500 |
| RGF 172-1 | ≤0.25 | 32 | ≤0.25 | 256 | >256 | 256 | >256 | 32 | 8 (4) | 500 |
| RGT 5-2 | ≤0.25 | 4 | ≤0.25 | 64 | >128 | 64 | >128 | 32 | 0.25 (128) | 500 |
| RGT 31-2 | ≤0.25 | 4 | 0.5 | 128 | >256 | 128 | >256 | 32 | 0.125 (256) | 500 |
| RGT 34-2 | ≤0.25 | 2 | ≤0.25 | 64 | >256 | 256 | >256 | 32 | 4 (8) | 500 |
| RGW 5-1 | ≤0.25 | 16 | ≤0.25 | 64 | >256 | 128 | >256 | 32 | 0.25 (256) | 500 |
| SZP 4-3-1 | 0.5 | 64 | ≤0.25 | >256 | >256 | 256 | >256 | 32 | 16 (2) | 500 |
| TF 18-2 | ≤0.25 | >128 | ≤0.25 | >256 | >128 | 128 | >128 | 32 | 0.5 (64) | 500 |
| TF 44-1 | ≤0.25 | ≤0.25 | ≤0.25 | >256 | >128 | 64 | >128 | 32 | 0.5 (64) | 500 |
| P. mirabilis | ||||||||||
| RGF 11-2 | ≤0.25 | 64 | >128 | >256 | >256 | 256 | 256 | 16 | 4 (4) | 2000 |
| RGF 134-1 | ≤0.25 | 64 | >128 | 128 | >256 | 128 | 128 | 32 | 4 (8) | 2000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, Z.; Xu, T.; Deng, T.; Shi, J.; Wang, Z.; Liu, Y. Benzydamine Reverses TMexCD-TOprJ-Mediated High-Level Tigecycline Resistance in Gram-Negative Bacteria. Pharmaceuticals 2021, 14, 907. https://doi.org/10.3390/ph14090907
Tong Z, Xu T, Deng T, Shi J, Wang Z, Liu Y. Benzydamine Reverses TMexCD-TOprJ-Mediated High-Level Tigecycline Resistance in Gram-Negative Bacteria. Pharmaceuticals. 2021; 14(9):907. https://doi.org/10.3390/ph14090907
Chicago/Turabian StyleTong, Ziwen, Tianqi Xu, Tian Deng, Jingru Shi, Zhiqiang Wang, and Yuan Liu. 2021. "Benzydamine Reverses TMexCD-TOprJ-Mediated High-Level Tigecycline Resistance in Gram-Negative Bacteria" Pharmaceuticals 14, no. 9: 907. https://doi.org/10.3390/ph14090907
APA StyleTong, Z., Xu, T., Deng, T., Shi, J., Wang, Z., & Liu, Y. (2021). Benzydamine Reverses TMexCD-TOprJ-Mediated High-Level Tigecycline Resistance in Gram-Negative Bacteria. Pharmaceuticals, 14(9), 907. https://doi.org/10.3390/ph14090907