
Phytomedicines Targeting Cancer Stem Cells: Therapeutic Opportunities and Prospects for Pharmaceutical Development

 , , , , ,
, , , , ,  ,
,  ,
,  ,
, 
 and
and 
Abstract
1. Introduction
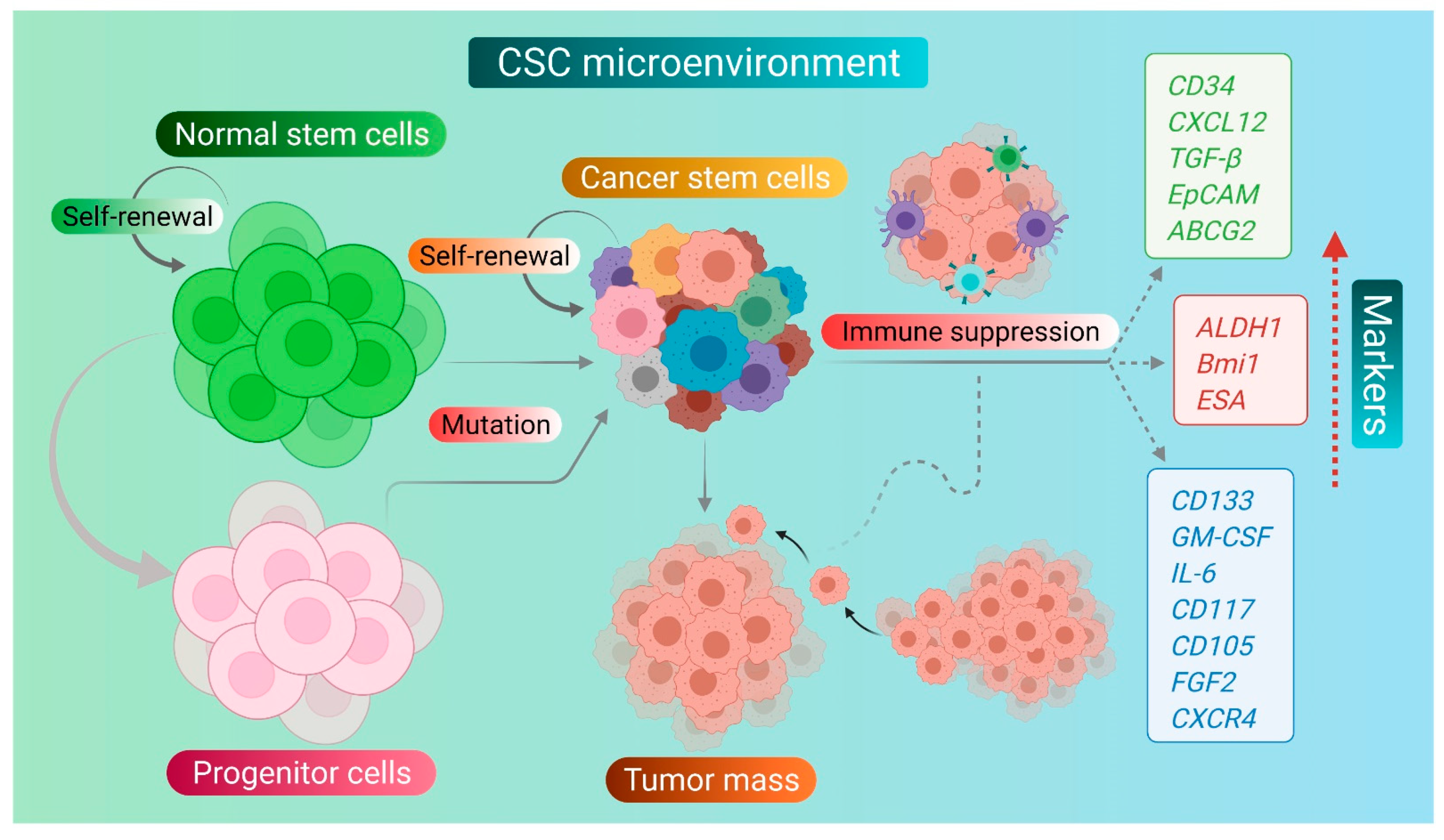
2. Cancer Stem Cells and Their Markers
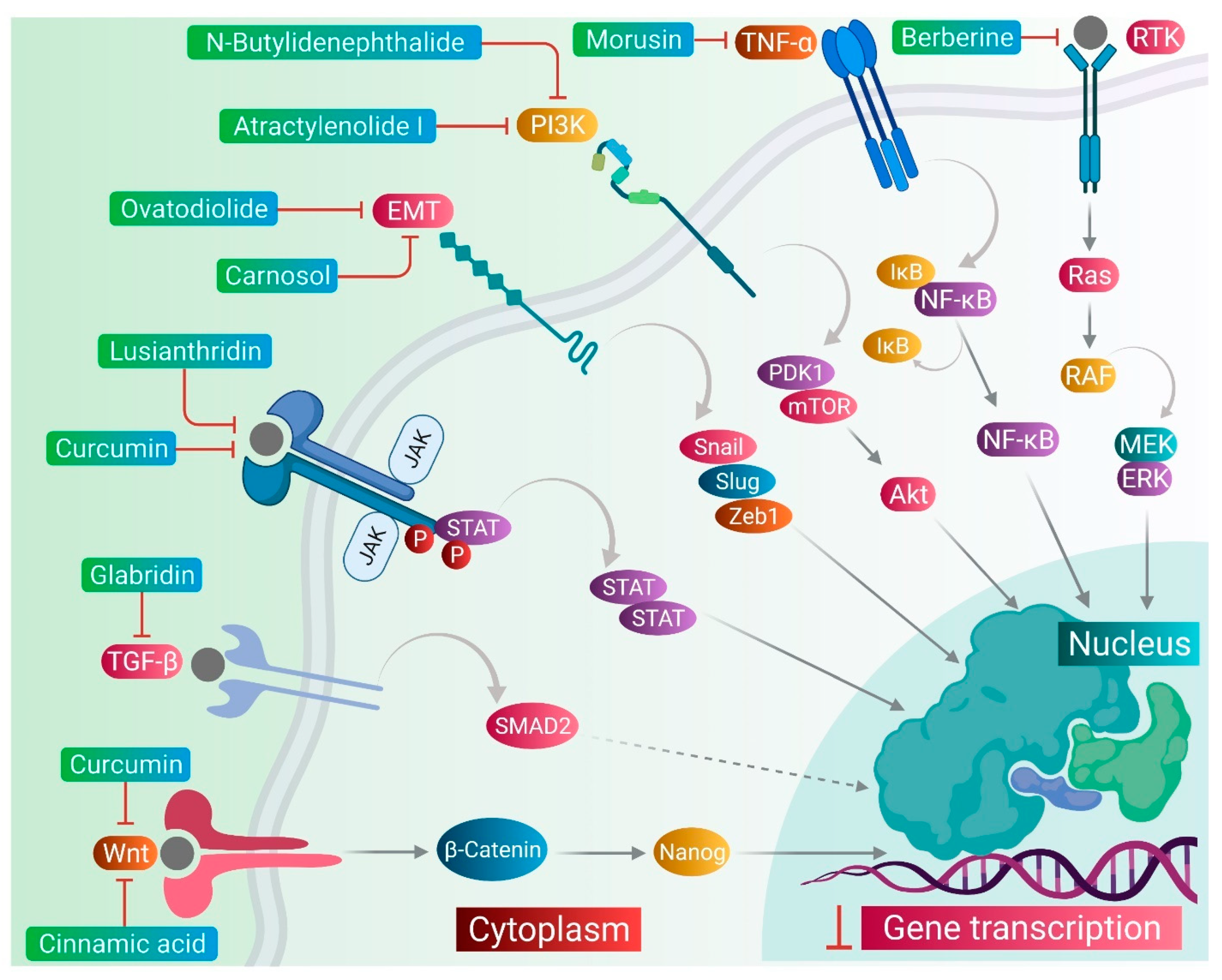
2.1. JAK/STAT Pathway
2.2. PI3K-Akt Pathway
2.3. NF-κB Pathway
2.4. Hedgehog Pathway
2.5. Wnt/β-Catenin Pathway
2.6. Notch Pathway
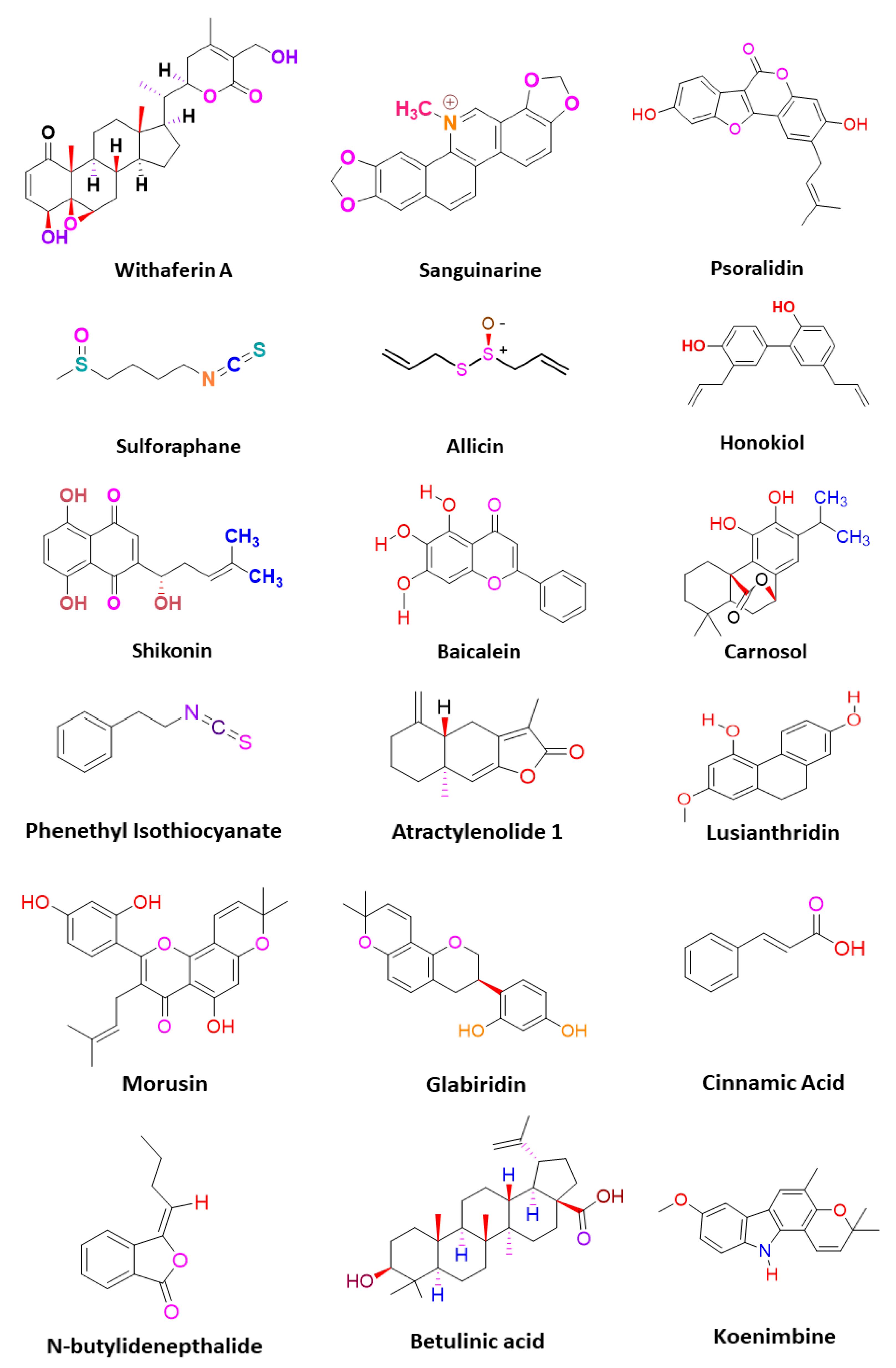
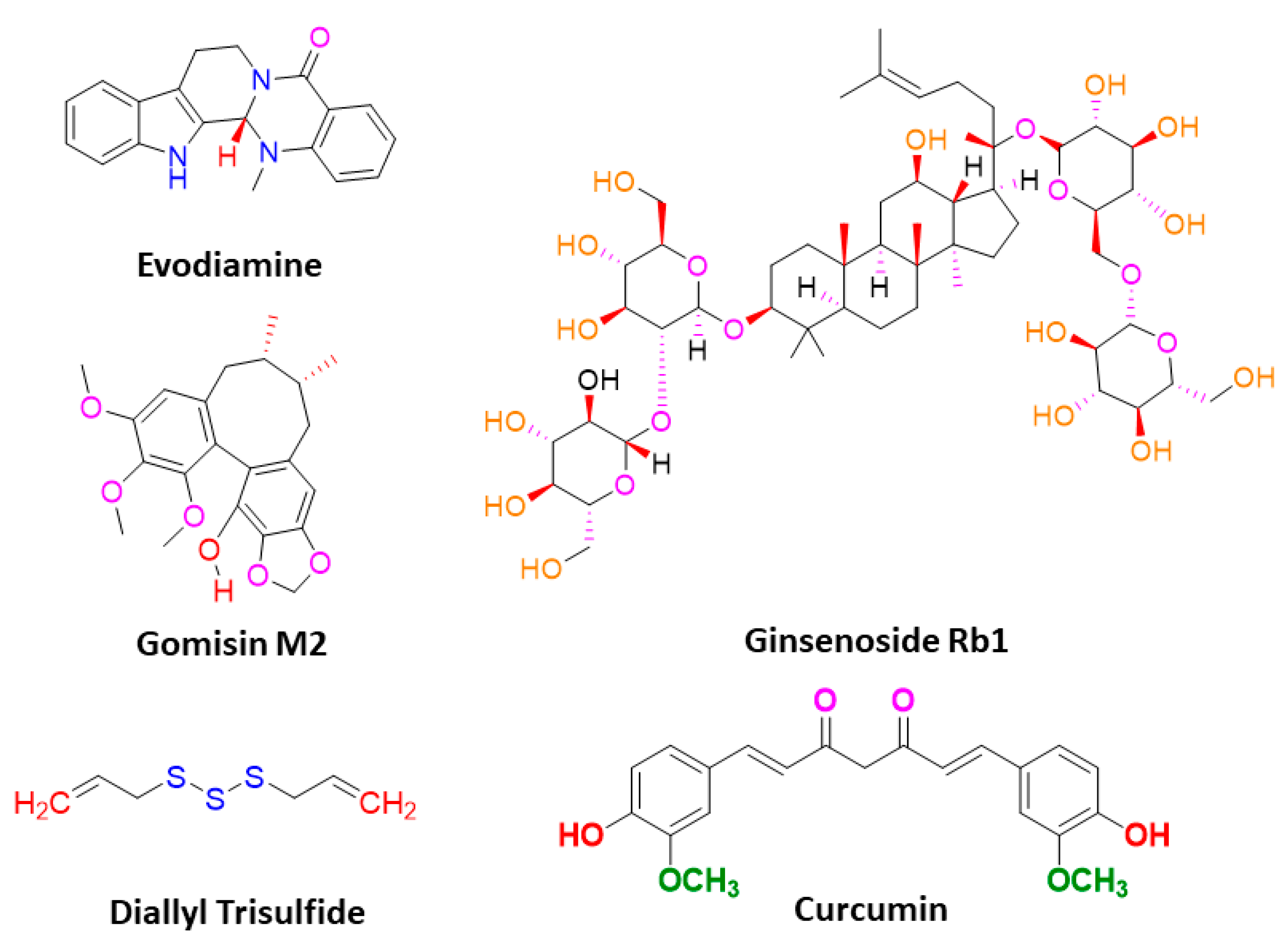
3. Phytomedicines Targeting Key Regulators of Anti-Cancer Drug Resistance in CSCs
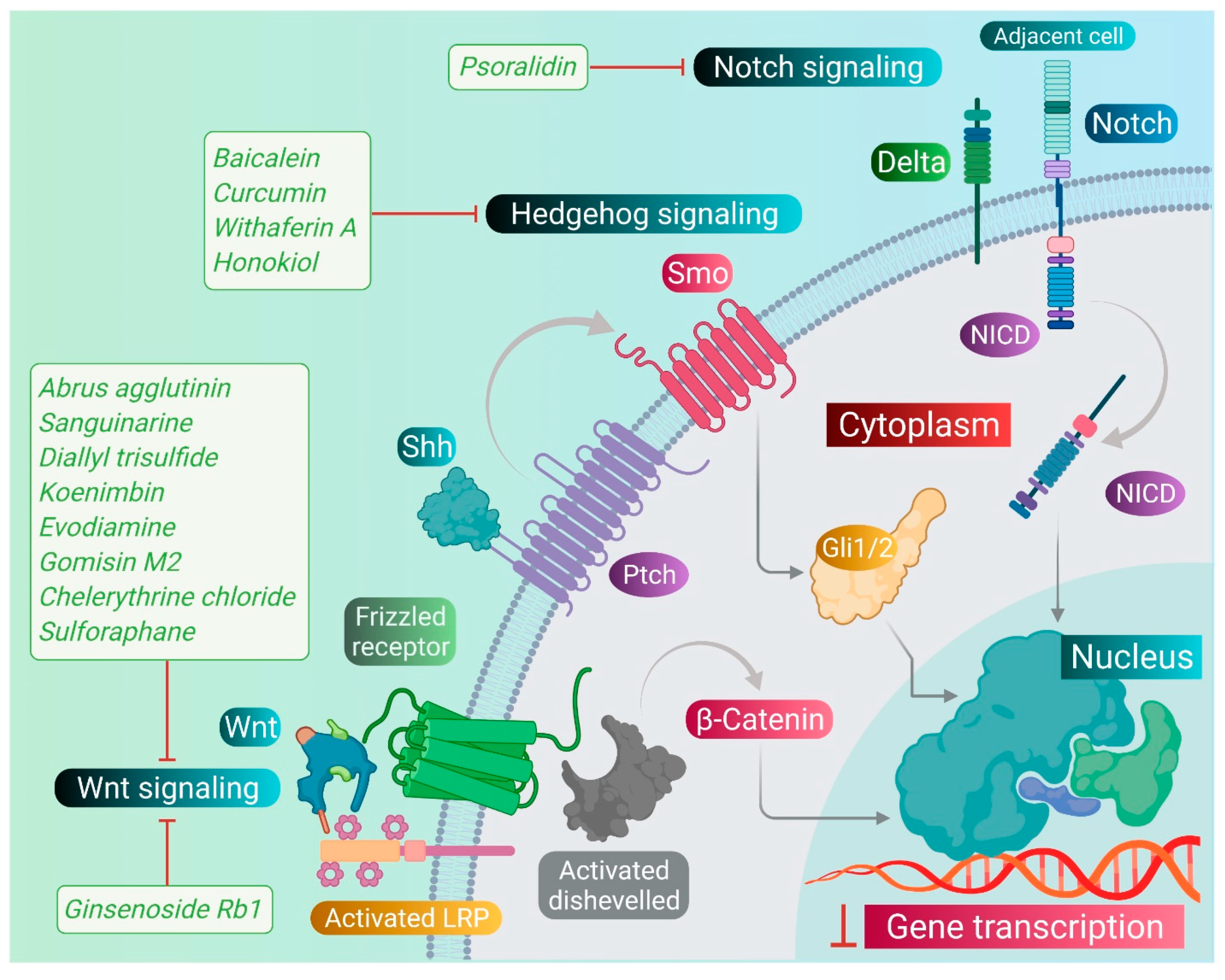
4. Phytomedicines Targeting Wnt/β-Catenin, Notch, and Hedgehog Signaling in CSCs
4.1. Phytomedicines Targeting Wnt/β-Catenin Signaling Pathway
4.2. Phytomedicines Targeting Notch Signaling Pathway
4.3. Phytomedicines Targeting Hedgehog Signaling Pathway
5. Clinical Studies on Phytomedicines
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Torquato, H.F.; Goettert, M.I.; Justo, G.Z.; Paredes-Gamero, E.J. Anti-Cancer Phytometabolites Targeting Cancer Stem Cells. Curr. Genom. 2017, 18, 156–174. [Google Scholar] [CrossRef]
- Chang, J.C. Cancer stem cells: Role in tumor growth, recurrence, metastasis, and treatment resistance. Medicine 2016, 95, S20–S25. [Google Scholar] [CrossRef]
- Liu, A.; Yu, X.; Liu, S. Pluripotency transcription factors and cancer stem cells: Small genes make a big difference. Chin. J. Cancer 2013, 32, 483–487. [Google Scholar] [CrossRef]
- Kusoglu, A.; Biray, A.C. Cancer stem cells: A brief review of the current status. Gene 2019, 681, 80–85. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Dudas, J.; Skvortsov, S.; Ganswindt, U.; Riechelmann, H.; Skvortsova, I.-I. Therapy resistance mediated by cancer stem cells. Semin. Cancer Biol. 2018, 53, 156–167. [Google Scholar] [CrossRef]
- Cianciosi, D.; Varela-Lopez, A.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Afrin, S.; Reboredo-Rodriguez, P.; Zhang, J.; Quiles, J.L.; Nabavi, S.F.; Battino, M.; et al. Tar-geting molecular pathways in cancer stem cells by natural bioactive compounds. Pharm. Res. 2018, 135, 150–165. [Google Scholar] [CrossRef]
- Oh, J.; Hlatky, L.; Jeong, Y.-S.; Kim, D. Therapeutic Effectiveness of Anticancer Phytochemicals on Cancer Stem Cells. Toxins 2016, 8, 199. [Google Scholar] [CrossRef]
- Yadav, A.K.; Desai, N.S. Cancer Stem Cells: Acquisition, Characteristics, Therapeutic Implications, Targeting Strategies and Future Prospects. Stem Cell Rev. Rep. 2019, 15, 331–355. [Google Scholar] [CrossRef]
- Bonam, S.R.; Wu, Y.S.; Tunki, L.; Chellian, R.; Halmuthur, M.S.K.; Muller, S.; Pandy, V. What Has Come out from Phytomedicines and Herbal Edibles for the Treatment of Cancer? ChemMedChem 2018, 13, 1854–1872. [Google Scholar] [CrossRef]
- Hermawan, A.; Putri, H. Current report of natural product development against breast cancer stem cells. Int. J. Biochem. Cell Biol. 2018, 104, 114–132. [Google Scholar] [CrossRef]
- Barbato, L.; Bocchetti, M.; Di Biase, A.; Regad, T. Cancer Stem Cells and Targeting Strategies. Cells 2019, 8, 926. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Gupta, P.K.; Dharanivasan, G.; Misra, R.; Gupta, S.; Verma, R.S. Nanomedicine in Cancer Stem Cell Therapy; Springer Science and Business Media LLC: Singapore, 2020; pp. 67–105. [Google Scholar]
- Makena, M.R.; Ranjan, A.; Thirumala, V.; Reddy, A.P. Cancer stem cells: Road to therapeutic resistance and strategies to overcome resistance. Biochim. Et Biophys. Acta (Bba) Mol. Basis Dis. 2020, 1866, 165339. [Google Scholar] [CrossRef]
- Leong, K.H.; Mahdzir, M.A.; Din, M.F.M.; Awang, K.; Tanaka, Y.; Kulkeaw, K.; Ishitani, T.; Sugiyama, D. Induction of intrinsic apoptosis in leu-kaemia stem cells and in vivo zebrafish model by betulonic acid isolated from Walsura pinnata Hassk (Meliaceae). Phytomedicine 2017, 26, 11–21. [Google Scholar] [CrossRef]
- Kaur, G.; Sharma, P.; Dogra, N.; Singh, S. Eradicating Cancer Stem Cells: Concepts, Issues, and Challenges. Curr. Treat. Options Oncol. 2018, 19, 20. [Google Scholar] [CrossRef]
- Li, S.-H.; Fu, J.; Watkins, D.N.; Srivastava, R.K.; Shankar, S. Sulforaphane regulates self-renewal of pancreatic cancer stem cells through the modulation of Sonic hedgehog–GLI pathway. Mol. Cell. Biochem. 2012, 373, 217–227. [Google Scholar] [CrossRef]
- Soltanian, S.; Riahirad, H.; Pabarja, A.; Jafari, E.; Khandani, B.K. Effect of Cinnamic acid and FOLFOX in diminishing side population and downregulating cancer stem cell markers in colon cancer cell line HT-29. Daru J. Pharm. Sci. 2018, 26, 19–29. [Google Scholar] [CrossRef]
- Yen, C.-H.; Lai, C.-C.; Shia, T.-H.; Chen, M.; Yu, H.-C.; Liu, Y.-P.; Chang, F.-R. Gynura divaricata attenuates tumor growth and tumor relapse after cisplatin therapy in HCC xenograft model through suppression of cancer stem cell growth and Wnt/β-catenin signalling. J. Ethnopharmacol. 2018, 213, 366–375. [Google Scholar] [CrossRef]
- Kamalidehghan, B.; Ahmadipour, F.; Ibrahim Noordin, M.; Mohan, S.; Arya, A.; Paydar, M.; Looi, C.Y. Koenimbin, a natural dietary compound of Murraya koenigii (L) Spreng: Inhibition of MCF7 breast cancer cells and targeting of derived MCF7 breast cancer stem cells (CD44+/CD24-/low): An in vitro study. Drug Des. Dev. 2015, 9, 1193–1208. [Google Scholar] [CrossRef][Green Version]
- Bhummaphan, N.; Petpiroon, N.; Prakhongcheep, O.; Sritularak, B.; Chanvorachote, P. Lusianthridin targeting of lung cancer stem cells via Src-STAT3 suppression. Phytomedicine 2019, 62, 152932. [Google Scholar] [CrossRef]
- Liu, S.-C.; Huang, C.-M.; Bamodu, O.A.; Lin, C.-S.; Liu, B.-L.; Tzeng, Y.-M.; Tsai, J.-T.; Lee, W.-H.; Chen, T.-M. Ovatodiolide suppresses nasopharyngeal cancer by targeting stem cell-like population, inducing apoptosis, inhibiting EMT and dysregulating JAK/STAT signaling pathway. Phytomedicine 2019, 56, 269–278. [Google Scholar] [CrossRef]
- Lin, C.S.; Bamodu, O.A.; Kuo, K.T.; Huang, C.M.; Liu, S.C.; Wang, C.H.; Tzeng, Y.M.; Chao, T.-Y.; Yeh, C.-T. Investigation of ovatodiolide, a macrocyclic diterpenoid, as a potential inhibitor of oral cancer stem-like cells properties via the inhibition of the JAK2/STAT3/JARID1B signal circuit. Phytomedicine 2018, 46, 93–103. [Google Scholar] [CrossRef]
- Jobani, B.M.; Najafzadeh, N.; Mazani, M.; Arzanlou, M.; Vardin, M.M. Molecular mechanism and cytotoxicity of allicin and all-trans retinoic acid against CD44+ versus CD117+ melanoma cells. Phytomedicine 2018, 48, 161–169. [Google Scholar] [CrossRef]
- Su, Y.-K.; Bamodu, O.A.; Tzeng, Y.-M.; Hsiao, M.; Yeh, C.-T.; Lin, C.-M. Ovatodiolide inhibits the oncogenicity and cancer stem cell-like phenotype of glioblastoma cells, as well as potentiate the anticancer effect of temozolomide. Phytomedicine 2019, 61, 152840. [Google Scholar] [CrossRef]
- Nisticò, P.; Bissell, M.J.; Radisky, D.C. Epithelial-Mesenchymal Transition: General Principles and Pathological Relevance with Special Emphasis on the Role of Matrix Metalloproteinases. Cold Spring Harb. Perspect. Biol. 2012, 4, a011908. [Google Scholar] [CrossRef]
- Salehi, B.; Varoni, E.M.; Sharifi-Rad, M.; Rajabi, S.; Zucca, P.; Iriti, M.; Rad, J.D. Epithelial-mesenchymal transition as a target for botanicals in cancer metastasis. Phytomedicine 2019, 55, 125–136. [Google Scholar] [CrossRef]
- Antony, J.; Thiery, J.P.; Huang, R.Y.-J. Epithelial-to-mesenchymal transition: Lessons from development, insights into cancer and the potential of EMT-subtype based therapeutic intervention. Phys. Biol. 2019, 16, 041004. [Google Scholar] [CrossRef]
- Tsai, J.H.; Yang, J. Epithelial-mesenchymal plasticity in carcinoma metastasis. Genes Dev. 2013, 27, 2192–2206. [Google Scholar] [CrossRef]
- Ayob, A.Z.; Ramasamy, T.S. Cancer stem cells as key drivers of tumour progression. J. Biomed. Sci. 2018, 25, 20. [Google Scholar] [CrossRef]
- Naveen, C.R.; Gaikwad, S.; Agrawal-Rajput, R. Berberine induces neuronal differentiation through inhibition of cancer stemness and epithelial-mesenchymal transition in neuroblastoma cells. Phytomedicine 2016, 23, 736–744. [Google Scholar] [CrossRef]
- Matsui, W.H. Cancer stem cell signaling pathways. Medicine 2016, 95, S8–S19. [Google Scholar] [CrossRef]
- Stine, R.R.; Matunis, E.L. JAK-STAT Signaling in Stem Cells. In Transcriptional and Translational Regulation of Stem Cells; Hime, G., Abud, H., Eds.; Springer Nature: Dordrecht, The Netherlands, 2012; pp. 247–267. [Google Scholar]
- Hemmings, B.A.; Restuccia, D.F. PI3K-PKB/Akt Pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011189. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]
- Kumar, G.; Farooqui, M.; Rao, C.V. Role of Dietary Cancer-Preventive Phytochemicals in Pancreatic Cancer Stem Cells. Curr. Pharm. Rep. 2018, 4, 326–335. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Karamboulas, C.; Ailles, L. Developmental signaling pathways in cancer stem cells of solid tumors. Biochim. Et Biophys. Acta (Bba) Gen. Subj. 2013, 1830, 2481–2495. [Google Scholar] [CrossRef]
- Wang, B.; Dai, Z.; Yang, X.-W.; Liu, Y.-P.; Khan, A.; Yang, Z.-F.; Huang, W.-Y.; Wang, X.-H.; Zhao, X.-D.; Luo, X.-D. Novel nor-monoterpenoid indole alkaloids inhibiting glioma stem cells from fruits of Alstonia scholaris. Phytomedicine 2018, 48, 170–178. [Google Scholar] [CrossRef]
- Yun, J.H.; Kim, K.A.; Yoo, G.; Kim, S.Y.; Shin, J.M.; Kim, J.H.; Jung, S.H.; Kim, J.; Nho, C.W. Phenethyl isothiocyanate suppresses cancer stem cell properties in vitro and in a xenograft model. Phytomedicine 2017, 30, 42–49. [Google Scholar] [CrossRef]
- Upadhyaya, B.; Liu, Y.; Dey, M. Phenethyl Isothiocyanate Exposure Promotes Oxidative Stress and Suppresses Sp1 Tran-scription Factor in Cancer Stem Cells. Int. J. Mol. Sci. 2019, 20, 1027. [Google Scholar] [CrossRef]
- Wang, K.; Huang, W.; Sang, X.; Wu, X.; Shan, Q.; Tang, D.; Xu, X.; Cao, G. Atractylenolide I inhibits colorectal cancer cell proliferation by affecting metabolism and stemness via AKT/mTOR signaling. Phytomedicine 2020, 68, 153191. [Google Scholar] [CrossRef]
- Cao, X.; Zou, H.; Cao, J.; Cui, Y.; Sun, S.; Ren, K.; Song, Z.; Li, D.; Quan, M. A candidate Chinese medicine preparation-Fructus Viticis Total Fla-vonoids inhibits stem-like characteristics of lung cancer stem-like cells. Bmc Complementary Altern. Med. 2016, 16, 364. [Google Scholar] [CrossRef]
- Seo, E.-J.; Wiench, B.; Hamm, R.; Paulsen, M.; Zu, Y.; Fu, Y.; Efferth, T. Cytotoxicity of natural products and derivatives toward MCF-7 cell monolayers and cancer stem-like mammospheres. Phytomedicine 2015, 22, 438–443. [Google Scholar] [CrossRef]
- El-Merahbi, R.; Liu, Y.N.; Eid, A.; Daoud, G.; Hosry, L.; Monzer, A.; Mouhieddine, T.H.; Hamade, A.; Najjar, F.; Abou-Kheir, W. Berberis libanotica Ehrenb Extract Shows An-ti-Neoplastic Effects on Prostate Cancer Stem/Progenitor Cells. PLoS ONE 2014, 9, e112453. [Google Scholar] [CrossRef]
- Hu, C.; Li, M.; Guo, T.; Wang, S.; Huang, W.; Yang, K.; Liao, Z.; Wang, J.; Zhang, F.; Wang, H. Anti-metastasis activity of curcumin against breast cancer via the inhibition of stem cell-like properties and EMT. Phytomedicine 2019, 58, 152740. [Google Scholar] [CrossRef]
- Zhou, Q.; Ye, M.; Lu, Y.; Zhang, H.; Chen, Q.; Huang, S.; Su, S. Curcumin Improves the Tumoricidal Effect of Mitomycin C by Suppressing ABCG2 Expression in Stem Cell-Like Breast Cancer Cells. PLoS ONE 2015, 10, e0136694. [Google Scholar] [CrossRef]
- Khan, A.Q.; Ahmed, E.I.; Elareer, N.; Fathima, H.; Prabhu, K.S.; Siveen, K.S.; Kulinski, M.; Azizi, F.; Dermime, S.; Ahmad, A.; et al. Curcumin-Mediated Apoptotic Cell Death in Papillary Thyroid Cancer and Cancer Stem-Like Cells through Targeting of the JAK/STAT3 Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 438. [Google Scholar] [CrossRef]
- Wang, Y.; Ying, X.; Xu, H.; Yan, H.; Li, X.; Tang, H. The functional curcumin liposomes induce apoptosis in C6 glioblastoma cells and C6 glioblastoma stem cells in vitro and in animals. Int. J. Nanomed. 2017, 12, 1369–1384. [Google Scholar] [CrossRef]
- Elkady, A.I. Targeting prostate cancer cell proliferation, stemness and metastatic potential using Costus speciosus derived phytochemicals. Am. J. Transl. Res. 2019, 11, 2550–2569. [Google Scholar]
- Yousefnia, S.; Naseri, D.; Forootan, F.S.; Tabatabaeian, M.; Moattar, F.; Ghafghazi, T.; Esfahani, M.H.N.; Ghaedi, K. Suppressive role of Viola odorata extract on malignant characters of mammosphere-derived breast cancer stem cells. Clin. Transl. Oncol. 2020, 22, 1619–1634. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Q.; Chen, K.; Sima, Z.; Liu, J.; Yu, Q.; Liu, J. 2-Ethoxystypandrone, a novel small-molecule STAT3 signaling inhibitor from Polygonum cuspidatum, inhibits cell growth and induces apoptosis of HCC cells and HCC Cancer stem cells. Bmc Complement. Altern. Med. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Jiang, F.; Li, Y.; Mu, J.; Hu, C.; Zhou, M.; Wang, X.; Si, L.; Ning, S.; Li, Z. Glabridin inhibits cancer stem cell-like properties of human breast cancer cells: An epigenetic regulation of miR-148a/SMAd2 signaling: GLA INHIBITS CSC-LIKE PROPERTIES VIA mIR-148a. Mol. Carcinog. 2016, 55, 929–940. [Google Scholar] [CrossRef]
- Wang, L.; Guo, H.; Yang, L.; Dong, L.; Lin, C.; Zhang, J.; Lin, P.; Wang, X. Morusin inhibits human cervical cancer stem cell growth and migration through attenuation of NF-κB activity and apoptosis induction. Mol. Cell. Biochem. 2013, 379, 7–18. [Google Scholar] [CrossRef]
- Liu, J.; Wang, P.; Xue, Y.-X.; Li, Z.; Qu, C.-B.; Liu, Y.-H. Enhanced antitumor effect of shikonin by inhibiting Endoplasmic Reticulum Stress via JNK/c-Jun pathway in human glioblastoma stem cells. Biochem. Biophys. Res. Commun. 2015, 466, 103–110. [Google Scholar] [CrossRef]
- Giacomelli, C.; Daniele, S.; Natali, L.; Iofrida, C.; Flamini, G.; Braca, A.; Trincavelli, M.L.; Martini, C. Carnosol controls the human glioblastoma stemness features through the epithelial-mesenchymal transition modulation and the induction of cancer stem cell apoptosis. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef]
- Wei, L.; Chen, P.; Chen, Y.; Shen, A.; Chen, H.; Lin, W.; Hong, Z.; Sferra, T.J.; Peng, J. Pien Tze Huang suppresses the stem-like side population in colorectal cancer cells. Mol. Med. Rep. 2013, 9, 261–266. [Google Scholar] [CrossRef]
- Gupta, P.K.; Pappuru, S.; Gupta, S.; Patra, B.; Chakraborty, D.; Verma, R.S. Self-assembled dual-drug loaded core-shell na-noparticles based on metal-free fully alternating polyester for cancer theranostics. Mater. Sci. Eng. C 2019, 101, 448–463. [Google Scholar] [CrossRef]
- Gupta, P.K.; Tripathi, S.K.; Pappuru, S.; Chabattula, S.C.; Govarthanan, K.; Gupta, S.; Biswal, B.K.; Chakraborty, D.; Verma, R.S. Metal-free semi-aromatic polyester as nanodrug carrier: A novel tumor targeting drug delivery vehicle for potential clinical application. Mater. Sci. Eng. C 2020, 107, 110285. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, X.-T.; Chen, Y.; Chen, J.-Q.; Zhu, J.-Y.; Meng, Y.; Wang, X.-Q.; Li, Y.; Geng, S.-S.; Xie, C.-F.; et al. Wnt/β-catenin signaling mediates the suppressive effects of diallyl trisulfide on colorectal cancer stem cells. Cancer Chemother. Pharm. 2018, 81, 969–977. [Google Scholar] [CrossRef]
- Deng, S.; Wong, C.K.C.; Lai, H.-C.; Wong, A.S.T. Ginsenoside-Rb1 targets chemotherapy-resistant ovarian cancer stem cells via simultaneous inhibition of Wnt/β-catenin signaling and epithelial-to-mesenchymal transition. Oncotarget 2016, 8, 25897–25914. [Google Scholar] [CrossRef]
- Sinha, N.; Panda, P.K.; Naik, P.P.; Maiti, T.K.; Bhutia, S.K. Abrus agglutinin targets cancer stem-like cells by eliminating self-renewal capacity accompanied with apoptosis in oral squamous cell carcinoma. Tumor Biol. 2017, 39, 1010428317701634. [Google Scholar] [CrossRef]
- Chen, L.; Chan, L.S.; Lung, H.L.; Yip, T.T.C.; Ngan, R.K.C.; Wong, J.W.C.; Lo, K.W.; Ng, W.T.; Lee, A.W.M.; Tsao, G.S.W.; et al. Crucifera sulforaphane (SFN) inhibits the growth of nasopharyngeal carcinoma through DNA methyltransferase 1 (DNMT1)/Wnt inhibitory factor 1 (WIF1) axis. Phytomedicine 2019, 63, 153058. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, S.; Chen, Y.; Li, X.; Jiang, Y.; Yang, X.; Li, Y.; Wang, X.; Meng, Y.; Zhu, M.; et al. miR-19 targeting of GSK3β mediates sulforaphane suppression of lung cancer stem cells. J. Nutr. Biochem. 2017, 44, 80–91. [Google Scholar] [CrossRef]
- Heng, W.S.; Cheah, S.-C. Chelerythrine Chloride Downregulates β-Catenin and Inhibits Stem Cell Properties of Non-Small Cell Lung Carcinoma. Molecules 2020, 25, 224. [Google Scholar] [CrossRef]
- Yang, J.; Fang, Z.; Wu, J.; Yin, X.; Fang, Y.; Zhao, F.; Zhu, S.; Li, Y. Construction and application of a lung cancer stem cell model: An-titumor drug screening and molecular mechanism of the inhibitory effects of sanguinarine. Tumor Biol. 2016, 37, 13871–13883. [Google Scholar] [CrossRef]
- Yang, Y.; Hao, E.; Pan, X.; Tan, D.; Du, Z.; Xie, J.; Hou, X.; Deng, J.; Wei, K. Gomisin M2 from Baizuan suppresses breast cancer stem cell proliferation in a zebrafish xenograft model. Aging 2019, 11, 8347–8361. [Google Scholar] [CrossRef]
- Kim, H.; Yu, Y.; Choi, S.; Lee, H.; Yu, J.; Lee, J.-H.; Kim, W.-Y. Evodiamine Eliminates Colon Cancer Stem Cells via Suppressing Notch and Wnt Signaling. Molecules 2019, 24, 4520. [Google Scholar] [CrossRef]
- Wen, Z.; Feng, S.; Wei, L.; Wang, Z.; Hong, D.; Wang, Q. Evodiamine, a novel inhibitor of the Wnt pathway, inhibits the self-renewal of gastric cancer stem cells. Int. J. Mol. Med. 2015, 36, 1657–1663. [Google Scholar] [CrossRef]
- Yanli, X.; Feiye, Z.; Shan, X.; Luming, L. Anti-tumor effect of the extract from Qingyihuaji formula on pancreatic cancer by down-regulating Notch-4 and Jagged-1. J. Tradit. Chin. Med. 2015, 35, 77–83. [Google Scholar] [CrossRef]
- Yan, B.; Liu, L.; Zhao, Y.; Xiu, L.-J.; Sun, D.-Z.; Liu, X.; Lu, Y.; Shi, J.; Zhang, Y.-C.; Li, Y.-J.; et al. Xiaotan Sanjie decoction attenuates tumor angiogenesis by manipulating Notch-1-regulated proliferation of gastric cancer stem-like cells. World J. Gastroenterol. 2014, 20, 13105–13118. [Google Scholar] [CrossRef]
- Youqin, C.; Wei, L.; Shen, A.; Chen, Y.; Lin, J.; Chu, J.; Cai, Q.; Pan, J.; Peng, J. Pien Tze Huang inhibits the proliferation, and induces the apoptosis and differentiation of colorectal cancer stem cells via suppression of the Notch1 pathway. Oncol. Rep. 2015, 35, 511–517. [Google Scholar] [CrossRef]
- Suman, S.; Das, T.P.; Damodaran, C. Silencing NOTCH signaling causes growth arrest in both breast cancer stem cells and breast cancer cells. Br. J. Cancer 2013, 109, 2587–2596. [Google Scholar] [CrossRef]
- Yoneyama, T.; Arai, M.A.; Sadhu, S.K.; Ahmed, F.; Ishibashi, M. Hedgehog inhibitors from Withania somnifera. Bioorganic Med. Chem. Lett. 2015, 25, 3541–3544. [Google Scholar] [CrossRef]
- Wang, D.; Kong, X.; Li, Y.; Qian, W.; Ma, J.; Wang, D.; Yu, D.; Zhong, C. Curcumin inhibits bladder cancer stem cells by suppressing Sonic Hedgehog pathway. Biochem. Biophys. Res. Commun. 2017, 493, 521–527. [Google Scholar] [CrossRef]
- Huynh, D.L.; Koh, H.; Chandimali, N.; Zhang, J.J.; Kim, N.; Kang, T.Y.; Ghosh, M.; Gera, M.; Park, Y.-H.; Kwon, T.; et al. BRM270 Inhibits the Proliferation of CD44 Positive Pan-creatic Ductal Adenocarcinoma Cells via Downregulation of Sonic Hedgehog Signaling. Evid. Based Complementary Altern. Med. 2019, 2019, 1–8. [Google Scholar] [CrossRef]
- Song, L.; Chen, X.; Wang, P.; Gao, S.; Qu, C.; Liu, L. Effects of baicalein on pancreatic cancer stem cells via modulation of sonic Hedgehog pathway. Acta Biochim. Et Biophys. Sin. 2018, 50, 586–596. [Google Scholar] [CrossRef]
- Yao, C.-J.; Han, T.-Y.; Shih, P.-H.; Yi, T.-Y.; Lai, I.-C.; Chang, K.-H.; Lai, T.-Y.; Chang, C.-L.; Lai, G.-M. Elimination of Cancer Stem–Like Side Population in Human Glioblastoma Cells Accompanied with Stemness Gene Suppression by Korean Herbal Recipe Mscintegr. Cancer. 2014, 13, 541–554. [Google Scholar] [CrossRef]
- Kamalidehghan, B.; Ghafouri-Fard, S.; Motevaseli, E.; Ahmadipour, F. Inhibition of human prostate cancer (PC-3) cells and targeting of PC-3-derived prostate cancer stem cells with koenimbin, a natural dietary compound from Murraya koenigii (L) Spreng. Drug Des. Dev. 2018, ume 12, 1119–1133. [Google Scholar] [CrossRef]
- Wang, J.; Lai, Z.; Chen, X.; Li, D.; Zhang, Y.; Ma, W.; Chu, Y.-t.; Shi, F.-q.; Yang, L.; Hou, L. Effect of Compound Zhebei Granule combined with chemotherapy on surface markers of leukemia stem cell in patients with acute myeloid leukemia. Chin. J. Integr. Med. 2016, Z22, 438–444. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | CSC Markers | References |
|---|---|---|
| Ovarian cancer | CD133+/CD44+/CD117+/ALDH1+/ABCG2+ | [2] |
| Stomach cancer | CD44+/CD133+ | [8] |
| Breast cancer | CD44+/ESA+/CD24−/ALDH1+/ABCG2+/EpCAM+/CXCR4 | [11] |
| Leukemia | CD34+CD38- | [16] |
| Brain tumor | CD133+/CD90+/ALDH1+ | [17] |
| Renal cancer | CD105+ | [17] |
| Pancreatic cancer | CD44+/CD24+/ESA+/CD133+/Bmi1/ALDH1+/ABCG2+/CXCR4 | [18] |
| Colon cancer | CD133+/ALDH1+/CD44+/EpCAM | [19] |
| Liver cancer | CD133+/CD90+/CD44+/ABCG2+/EpCAM+/CD13+ | [20] |
| Prostate cancer | CD44+/CD133+/ALDH1+/Bmi1 | [21] |
| Lung cancer | CD133+/CD117+/ALDH1+/ABCG2+/EpCAM | [22] |
| Nasopharyngeal cancer | CD44+/CD133+/ALDH1+/ABCG2+/Bmi1 | [23] |
| Oral cancer | CD44+/ALDH1+/CD117+/Bmi1 | [24] |
| Melanoma | ABCB5+/ALDH1+/CD133+/CD44+/CD117+ | [25] |
| Glioblastoma | CD133+/CD44+/Bmi1 | [26] |
| Plant Source | Extract | Bioactive Compound | Mode of Action | In Vivo Dose | Cells/Model | References |
|---|---|---|---|---|---|---|
| Alstonia scholaris | Fruit extract | Scholarisine Q(1) and R(2) |
| ------ | Glioma stem cells | [40] |
| Anisomeles indica | ------ | Ovatodiolide |
| ------ | Glioblastoma stem-like cells | [26] |
| ------ | CSC population in nasopharyngeal carcinoma | [23] | |||
| 3.6 mg/kg | Oral CSCs and xenograft tumor mice | [24] | |||
| Cruciferous vegetables | ------ | Phenethyl isothiocyanate |
| 20 mg/kg | Colon CSCs and xenograft tumor mice | [41] |
| 10 µM | Cervical CSCs and Xenograft NOD-SCID tumor mice | [42] | |||
| Atractylodes macrocephala Koidz | Rhizome extract | Atractylenolide I |
| 25 mg/kg and 75 mg/kg | Stemness of colon cancer cells and xenograft tumor mice | [43] |
| Fructus viticis | ------ | Flavonoids |
| ------ | Lung CSCs | [44] |
| Pigeon pea | ------ | Cajaninstilbene acid derivatives |
| ------ | Breast cancer stem-like cells | [45] |
| Berberis libanotica Ehrenb | Root extract | ------ |
| ------ | Prostate CSCs | [46] |
| Berberis, Arcangelisia Hydrastis | ------ | Berberine |
| ------ | Stemness in neuroblastoma cells | [32] |
| Dendrobium venustum | Stem extract | Lusianthridin |
| ------ | Lung CSCs | [22] |
| Curcuma longa | ------ | Curcumin |
| ------ | Breast CSCs | [47] |
| ------ | Curcumin |
| ------ | Breast CSCs | [48] | |
| ------ | Curcumin |
| ------ | Papillary thyroid CSCs | [49] | |
| ------ | Curcumin |
| 5 mg/kg | Glioblastoma stem cells and xenograft tumor mice | [50] | |
| Walsura pinnata Hassk | Bark extract | Betulonic acid |
| 18, 36, or 54 μM | Leukemia stem cells and xenotransplanted zebrafish | [16] |
| Costus speciosus | Rhizome extract | ------ |
| ------ | Stemness of prostate cancer cells | [51] |
| Viola odorata | Hydro-alcoholic extract of aerial part | ------ |
| ------ | Breast CSCs | [52] |
| Polygonum cuspidatum | Root extract | 2-Ethoxystypandrone |
| ------ | Hepatocellular CSCs | [53] |
| Cinnamomum cassia | ------ | Cinnamic acid |
| ------ | Colon CSCs | [19] |
| Glycyrrhiza glabra | ------ | Glabridin |
| 20 mg/kg | Breast cancer stem-like cells and xenograft tumor mice | [54] |
| Morus australis | ------ | Morusin |
| ------ | Cervical CSCs | [55] |
| Lithospermum erythrorhizon | ------ | Shikonin |
| 2 mg/kg | Glioblastoma stem cells and xenograft tumor mice | [56] |
| Rosmarinus officinalis | ------ | Carnosol |
| ------ | Glioblastoma CSCs | [57] |
| PienTze Huang | ------ | ------ |
| ------ | Colorectal CSCs | [58] |
| Allium sativum | ------ | Allicin (diallyl thiosulfinate) |
| ------ | Melanoma stem-like cells | [25] |
| Plant Source | Extract | Bioactive Compound | Mode of Action | In Vivo Dose | Cell Line/Model | References |
|---|---|---|---|---|---|---|
| Abrus precatorius | Seed extract | Abrus agglutinin |
| 50 μg/kg | Oral squamous carcinoma stem-like cells and xenograft tumor mice | [63] |
| Celandine (TCM) | ------ | Sanguinarine |
| 0.5 mg/20 g | Lung CSCs and xenograft tumor mice | [67] |
| Gynura divaricata | Aqueous extract of aerial part | ------ |
| 300 mg/kg | Hepatocellular CSCs and xenograft tumor mice | [20] |
| Panax quinquefolius | ------ | Ginsenoside-Rb1 |
| 50 mg/kg | Ovarian CSCs and xenograft tumor mice | [62] |
| Allium sativum | ------ | Diallyl-trisulfide |
| ------ | Colorectal CSCs | [61] |
| Murraya koenigii(L) Spreng | Leaf extract | Koenimbin |
| ------ | Breast CSCs | [21] |
| ------ | Prostate CSCs | [80] | |||
| Chelidonium majus/Macleaya cordata | ------ | Chelerythrine chloride |
| ------ | Non-small cell lung carcinoma stem-like cells | [66] |
| Evodiae rutaecarpa | ------ | Evodiamine |
| ------ | Gastric CSCs | [70] |
| Evodiae fructus | ------ | Evodiamine |
| ------ | Colon CSCs | [69] |
| Baizuan (TCM) | ------ | Gomisin M2 |
| 10 μM | Breast CSCs and zebrafish xenograft | [68] |
| Cruciferous vegetables | ------ | Sulforaphane |
| 60 mg/kg | Nasopharyngeal CSCs and xenograft tumor mice | [64] |
| ------ | Lung CSCs | [65] | |||
| ------ | Pancreatic CSCs | [18] | |||
| Curcuma longa | ------ | Curcumin |
| ------ | Bladder CSCs | [50] |
| Withania somnifera | Leaf extract | Withaferin A |
| ------ | Pancreatic, prostate, and breast CSCs | [75] |
| BRM270 | Alcohol extract | ------ |
| 5 mg/kg | Pancreatic ductal adenocarcinoma stem cells and xenograft tumor mice | [77] |
| Qingyihuaji (TCM) | ------ | Baicalein |
| 20 or 60 mg/kg | Pancreatic CSCs and xenograft tumor mice | [78] |
| Qingyihuaji (TCM) | Aqueous extract | ------ |
| 36 g/kg | Pancreatic cancer stem-like cells and xenograft tumor mice | [71] |
| Xiaotansanjie (TCM) | ------ | ------ |
| 1.46, 2.92, and 5.84 g/mL | Gastric CSCs and xenograft tumor mice | [72] |
| Psoralea corylifolia | ------ | Psoralidin |
| ------ | Breast CSCs | [74] |
| PienTze Huang (TCM) | ------ | ------ |
| ------ | Colorectal CSCs | [73] |
| MSC500 | ------ | ------ |
| ------ | Glioblastoma stem cells | [79] |
| Clinical Trial No. | Sponsors and Collaborators | Title of the Study | Clinical Status | Year of Study (Start Date–Completion Date) |
|---|---|---|---|---|
| ||||
| NCT00982319 | Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins | Study to Evaluate the Effect of Sulforaphane in Broccoli Sprout Extract on Breast Tissue | Phase 2 | 2009–2013 |
| NCT03665922 | University of Pittsburgh | Biomarkers of Sulforaphane/Broccoli Sprout Extract in Prostate Cancer | Recruiting (Phase not applicable) | 2019–2024 |
| ||||
| NCT01740323 | Andrew H Miller and National Cancer Institute | Phase II Study of Curcumin vs. Placebo for Chemotherapy-Treated Breast Cancer Patients Undergoing Radiotherapy | Phase 2 | 2015–2018 |
| NCT03980509 | Medical University of South Carolina | A “Window Trial” on Curcumin for Invasive Breast Cancer Primary Tumors | Phase 1 | 2020–2021 |
| NCT03072992 | National Center of Oncology, Armenia, and BRIU GmbH | “Curcumin” in Combination with Chemotherapy in Advanced Breast Cancer | Phase 2 | 2017–2019 |
| ||||
| NCT01790204 | Georgetown University | A Study of the Effects of PEITC on Oral Cells with Mutant p53 | Phase 2 | 2012–2014 |
| NCT00691132 | University of Minnesota and National Cancer Institute | Phenethyl Isothiocyanate in Preventing Lung Cancer in Smokers | Phase 2 | 2009–2013 |
| ||||
| NCT00079170 | National Cancer Institute | Docetaxel Plus Garlic in Treating Patients with Locally Advanced or Metastatic Breast Cancer | Pilot study (Phase not applicable) | 2004–2007 |
| ||||
| NCT02226185 | Shanghai Jiao Tong University School of Medicine | Study of Berberine Hydrochloride in Prevention of Colorectal Adenomas Recurrence | Phase 3 | 2014–2018 |
| ||||
| NCT00176631 | Rutgers, The State University of New Jersey, and National Cancer Institute | Licorice Root Extract and Docetaxel in Treating Patients with Metastatic Prostate Cancer That Did Not Respond to Hormone Therapy | Phase 2 | 2007–2008 |
| ||||
| NCT03234595 | Everfront Biotech Co., Ltd. | A Phase I/IIa Study of Cerebraca Wafer Plus Adjuvant Temozolomide (TMZ) in Patients with Recurrent High-Grade Glioma | Phase 2 | 2017–2021 |
| ||||
| NCT02714608 | Tasly Pharmaceuticals, Inc. | A Study of Ginsenoside H Dripping Pills for Advanced Non-Small Cell Lung Cancer (NSCLC) | Phase 2 | 2016–2018 |
| ||||
| NCT00689195 | Tata Memorial Hospital and Pharmanza Herbals Pvt Limited (PHPL) | Pilot Study of Curcumin Formulation and Ashwagandha Extract in Advanced Osteosarcoma (OSCAT) | Phase 2 | 2008–2013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, P.K.; Saraff, M.; Gahtori, R.; Negi, N.; Tripathi, S.K.; Kumar, J.; Kumar, S.; Aldhayan, S.H.; Dhanasekaran, S.; Abomughaid, M.M.; et al. Phytomedicines Targeting Cancer Stem Cells: Therapeutic Opportunities and Prospects for Pharmaceutical Development. Pharmaceuticals 2021, 14, 676. https://doi.org/10.3390/ph14070676
Gupta PK, Saraff M, Gahtori R, Negi N, Tripathi SK, Kumar J, Kumar S, Aldhayan SH, Dhanasekaran S, Abomughaid MM, et al. Phytomedicines Targeting Cancer Stem Cells: Therapeutic Opportunities and Prospects for Pharmaceutical Development. Pharmaceuticals. 2021; 14(7):676. https://doi.org/10.3390/ph14070676
Chicago/Turabian StyleGupta, Piyush Kumar, Mrunmayee Saraff, Rekha Gahtori, Nidhi Negi, Surya Kant Tripathi, Jatin Kumar, Sanjay Kumar, Saad Hamad Aldhayan, Sugapriya Dhanasekaran, Mosleh Mohammad Abomughaid, and et al. 2021. "Phytomedicines Targeting Cancer Stem Cells: Therapeutic Opportunities and Prospects for Pharmaceutical Development" Pharmaceuticals 14, no. 7: 676. https://doi.org/10.3390/ph14070676
APA StyleGupta, P. K., Saraff, M., Gahtori, R., Negi, N., Tripathi, S. K., Kumar, J., Kumar, S., Aldhayan, S. H., Dhanasekaran, S., Abomughaid, M. M., Dua, K., Gundamaraju, R., Ojha, S., Ruokolainen, J., Jha, N. K., & Kesari, K. K. (2021). Phytomedicines Targeting Cancer Stem Cells: Therapeutic Opportunities and Prospects for Pharmaceutical Development. Pharmaceuticals, 14(7), 676. https://doi.org/10.3390/ph14070676