
Biphasic Production of Anti-ApoB100 Autoantibodies in Obese Humans and Mice


Abstract
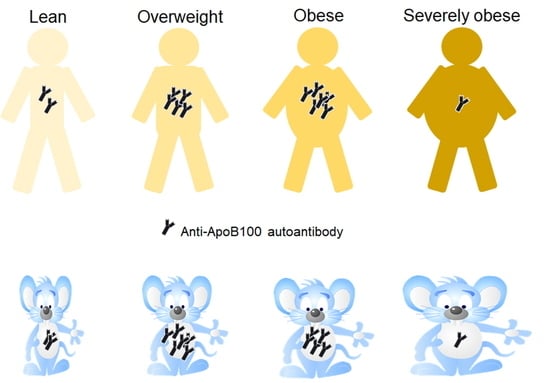
1. Introduction
2. Results
2.1. Metabolic Context
2.2. Obese Mice and Humans Produce ApoB100/p210/pB1-Specific Autoantibodies
2.3. The Autoantibody Response in Obese Mice Exhibits a Biphasic Pattern
2.4. The Autoantibody Response in Obese Humans Also Exhibits a Biphasic Pattern
3. Discussion
4. Materials and Methods
4.1. Animals, Diets, and Serum Harvest
4.2. Human Subjects and Serum Harvest
4.3. Peptide Preparation
4.4. Serum Antibody Analysis
4.5. Purification of Autoantibodies against ApoB100
4.6. Dot-Blots
4.7. Identification of Isotype
4.8. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saltiel, A.R. New therapeutic approaches for the treatment of obesity. Sci. Transl. Med. 2016, 8, 323rv322. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, G.; Apovian, C. Future Pharmacotherapy for Obesity: New Anti-Obesity Drugs on the Horizon. Curr. Obes. Rep. 2018, 7, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.; Muscogiuri, G.; Savastano, S.; Barrea, L.; Guida, B.; Taglialatela, M.; Colao, A. Gender-related issues in the pharmacology of new anti-obesity drugs. Obes. Rev. 2019, 20, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S. Peptides and Peptidomimetics as Potential Antiobesity Agents: Overview of Current Status. Front. Nutr. 2019, 6, 11. [Google Scholar] [CrossRef]
- Hersoug, L.G.; Linneberg, A. The link between the epidemics of obesity and allergic diseases: Does obesity induce decreased immune tolerance? Allergy 2007, 62, 1205–1213. [Google Scholar] [CrossRef]
- Versini, M.; Jeandel, P.Y.; Rosenthal, E.; Shoenfeld, Y. Obesity in autoimmune diseases: Not a passive bystander. Autoimmun. Rev. 2014, 13, 981–1000. [Google Scholar] [CrossRef]
- Arai, S.; Maehara, N.; Iwamura, Y.; Honda, S.; Nakashima, K.; Kai, T.; Ogishi, M.; Morita, K.; Kurokawa, J.; Mori, M.; et al. Obesity-associated autoantibody production requires AIM to retain the immunoglobulin M immune complex on follicular dendritic cells. Cell Rep. 2013, 3, 1187–1198. [Google Scholar] [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Thaller, S.; Blomberg, B.B. Secretion of autoimmune antibodies in the human subcutaneous adipose tissue. PLoS ONE 2018, 13, e0197472. [Google Scholar] [CrossRef]
- Elkon, K.; Casali, P. Nature and functions of autoantibodies. Nat. Clin. Pract. Rheumatol. 2008, 4, 491–498. [Google Scholar] [CrossRef]
- Leslie, D.; Lipsky, P.; Notkins, A.L. Autoantibodies as predictors of disease. J. Clin. Investig. 2001, 108, 1417–1422. [Google Scholar] [CrossRef]
- Wildbaum, G.; Nahir, M.A.; Karin, N. Beneficial autoimmunity to proinflammatory mediators restrains the consequences of self-destructive immunity. Immunity 2003, 19, 679–688. [Google Scholar] [CrossRef]
- Zohar, Y.; Wildbaum, G.; Karin, N. Beneficial autoimmunity participates in the regulation of rheumatoid arthritis. Front. Biosci. 2006, 11, 368–379. [Google Scholar] [CrossRef]
- Meyer, S.; Woodward, M.; Hertel, C.; Vlaicu, P.; Haque, Y.; Karner, J.; Macagno, A.; Onuoha, S.C.; Fishman, D.; Peterson, H.; et al. AIRE-Deficient Patients Harbor Unique High-Affinity Disease-Ameliorating Autoantibodies. Cell 2016, 166, 582–595. [Google Scholar] [CrossRef]
- Karin, N. Autoantibodies to Chemokines and Cytokines Participate in the Regulation of Cancer and Autoimmunity. Front. Immunol. 2018, 9, 623. [Google Scholar] [CrossRef]
- Wolf, D.; Ley, K. Immunity and Inflammation in Atherosclerosis. Circ. Res. 2019, 124, 315–327. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Browne, S.; Holland, S.M.; Iadarola, M.J.; Alevizos, I. Clinical features of Sjogren’s syndrome patients with autoantibodies against interferons. Clin. Transl. Med. 2019, 8, 1. [Google Scholar] [CrossRef]
- Hertel, C.; Fishman, D.; Lorenc, A.; Ranki, A.; Krohn, K.; Peterson, P.; Kisand, K.; Hayday, A. Response to comment on ‘AIRE-deficient patients harbor unique high-affinity disease-ameliorating autoantibodies’. eLife 2019, 8. [Google Scholar] [CrossRef]
- Sjogren, P.; Fredrikson, G.N.; Samnegard, A.; Ericsson, C.G.; Ohrvik, J.; Fisher, R.M.; Nilsson, J.; Hamsten, A. High plasma concentrations of autoantibodies against native peptide 210 of apoB-100 are related to less coronary atherosclerosis and lower risk of myocardial infarction. Eur. Heart J. 2008, 29, 2218–2226. [Google Scholar] [CrossRef]
- Bjorkbacka, H.; Alm, R.; Persson, M.; Hedblad, B.; Nilsson, J.; Fredrikson, G.N. Low Levels of Apolipoprotein B-100 Autoantibodies Are Associated with Increased Risk of Coronary Events. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 765–771. [Google Scholar] [CrossRef]
- Fredrikson, G.N.; Soderberg, I.; Lindholm, M.; Dimayuga, P.; Chyu, K.Y.; Shah, P.K.; Nilsson, J. Inhibition of atherosclerosis in apoE-null mice by immunization with apoB-100 peptide sequences. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 879–884. [Google Scholar] [CrossRef]
- Amirfakhryan, H. Vaccination against atherosclerosis: An overview. Hell. J. Cardiol. 2020, 61, 78–91. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, H.J.; Choi, J.S.; Han, J.; Kim, J.Y.; Na, H.K.; Joung, H.J.; Kim, Y.S.; Binas, B. An apolipoprotein B100 mimotope prevents obesity in mice. Clin. Sci. 2016, 130, 105–116. [Google Scholar] [CrossRef]
- Liu, J.; Han, L.; Zhu, L.; Yu, Y. Free fatty acids, not triglycerides, are associated with non-alcoholic liver injury progression in high fat diet induced obese rats. Lipids Health Dis. 2016, 15, 27. [Google Scholar] [CrossRef]
- Fernandez, I.; Pena, A.; Del Teso, N.; Perez, V.; Rodriguez-Cuesta, J. Clinical biochemistry parameters in C57BL/6J mice after blood collection from the submandibular vein and retroorbital plexus. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 202–206. [Google Scholar]
- List, E.O.; Berryman, D.E.; Wright-Piekarski, J.; Jara, A.; Funk, K.; Kopchick, J.J. The effects of weight cycling on lifespan in male C57BL/6J mice. Int. J. Obes. 2013, 37, 1088–1094. [Google Scholar] [CrossRef]
- Fredrikson, G.N.; Bjorkbacka, H.; Soderberg, I.; Ljungcrantz, I.; Nilsson, J. Treatment with apo B peptide vaccines inhibits atherosclerosis in human apo B-100 transgenic mice without inducing an increase in peptide-specific antibodies. J. Intern. Med. 2008, 264, 563–570. [Google Scholar] [CrossRef]
- Klingenberg, R.; Lebens, M.; Hermansson, A.; Fredrikson, G.N.; Strodthoff, D.; Rudling, M.; Ketelhuth, D.F.; Gerdes, N.; Holmgren, J.; Nilsson, J.; et al. Intranasal immunization with an apolipoprotein B-100 fusion protein induces antigen-specific regulatory T cells and reduces atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 946–952. [Google Scholar] [CrossRef]
- Morrow, J.D. Is oxidant stress a connection between obesity and atherosclerosis? Arterioscler. Thromb. Vasc. Biol. 2003, 23, 368–370. [Google Scholar] [CrossRef]
- King, V.L.; Hatch, N.W.; Chan, H.W.; de Beer, M.C.; de Beer, F.C.; Tannock, L.R. A murine model of obesity with accelerated atherosclerosis. Obesity 2010, 18, 35–41. [Google Scholar] [CrossRef]
- Fredrikson, G.N.; Hedblad, B.; Berglund, G.; Alm, R.; Ares, M.; Cercek, B.; Chyu, K.Y.; Shah, P.K.; Nilsson, J. Identification of immune responses against aldehyde-modified peptide sequences in apoB associated with cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 872–878. [Google Scholar] [CrossRef]
- Fredrikson, G.N.; Schiopu, A.; Berglund, G.; Alm, R.; Shah, P.K.; Nilsson, J. Autoantibody against the amino acid sequence 661–680 in apo B-100 is associated with decreased carotid stenosis and cardiovascular events. Atherosclerosis 2007, 194, e188–e192. [Google Scholar] [CrossRef] [PubMed]
- McLeod, O.; Silveira, A.; Fredrikson, G.N.; Gertow, K.; Baldassarre, D.; Veglia, F.; Sennblad, B.; Strawbridge, R.J.; Larsson, M.; Leander, K.; et al. Plasma autoantibodies against apolipoprotein B-100 peptide 210 in subclinical atherosclerosis. Atherosclerosis 2014, 232, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.K.; Choe, M.K.; Kim, H.J.; Kim, Y.S.; Binas, B.; Kim, H.J. An ApoB100-mimetic vaccine prevents obesity and liver steatosis in ApoE-/- mice. Pharmacol. Rep. 2017, 69, 1140–1144. [Google Scholar] [CrossRef] [PubMed]
- Ketelhuth, D.F.; Rios, F.J.; Wang, Y.; Liu, H.; Johansson, M.E.; Fredrikson, G.N.; Hedin, U.; Gidlund, M.; Nilsson, J.; Hansson, G.K.; et al. Identification of a danger-associated peptide from apolipoprotein B100 (ApoBDS-1) that triggers innate proatherogenic responses. Circulation 2011, 124, 2433–2443. [Google Scholar] [CrossRef]
- Kim, N.H.; Ahn, J.; Choi, Y.M.; Son, H.J.; Choi, W.H.; Cho, H.J.; Yu, J.H.; Seo, J.A.; Jang, Y.J.; Jung, C.H.; et al. Differential circulating and visceral fat microRNA expression of non-obese and obese subjects. Clin. Nutr. 2020, 39, 910–916. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | BMI (kg/m2) | |||
|---|---|---|---|---|
| <23.0 (n = 33) | 23.0 to <25.0 (n = 41) | 25.0 to <27.0 (n = 30) | ≥27.0 (n = 44) | |
| Age (years) | 54.4 ± 1.6 | 54.4 ± 1.5 | 54.3 ± 1.6 | 52.0 ± 1.4 |
| Hight (cm) | 162.2 ± 1.8 | 165.5 ± 1.2 | 163.2 ± 1.6 | 164.4 ± 1.5 |
| Weight (kg) | 58.0 ± 1.3 | 65.2 ± 1.0 | 69.7 ± 1.4 | 78.6 ± 1.5 |
| BMI (kg/m2) | 21.8 ± 0.1 | 23.8 ± 0.1 | 26.0 ± 0.1 | 29.2 ± 0.3 |
| Diabetes (%) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Hypertension (%) | 9 (27.3) | 7 (16.7) | 3 (10.0) | 13 (39.4) |
| Dyslipidemia (%) | 2 (6.1) | 5 (11.9) | 2 (6.7) | 6 (18.2) |
| Smoking (%) | 7 (21.2) | 8 (19.0) | 4 (13.3) | 8 (24.2) |
| Alcohol (%) | 12 (36.4) | 21 (50.0) | 14 (46.7) | 22 (66.7) |
| Exercise (%) | 14 (42.4) | 22 (52.4) | 15 (50.0) | 19 (57.6) |
| WC (cm) | 78.7 ± 1.1 | 83.8 ± 0.7 | 86.2 ± 0.6 | 92.3 ± 0.9 |
| SBP (mm/Hg) | 116.3 ± 2.3 | 120.8 ± 2.2 | 120.0 ± 2.3 | 121.7 ± 1.7 |
| FPG (mg/dL) | 97.1 ± 2.3 | 95.0 ± 1.9 | 92.4 ± 2.0 | 97.6 ± 1.6 |
| HbA1c (%) | 5.5 ± 0.1 | 5.4 ± 0.1 | 5.4 ± 0.1 | 5.5 ± 0.1 |
| Total cholesterol (mg/dL) | 201.9 ± 5.9 | 203.0 ± 5.9 | 194.8 ± 5.2 | 200.3 ± 5.5 |
| Triglyceride (mg/dL) | 141.6 ± 23.2 | 172.1 ± 14.5 | 142.5 ± 14.7 | 173.5 ± 15.2 |
| LDL-C (mg/dL) | 125.4 ± 5.0 | 117.0 ± 6.1 | 123.8 ± 4.9 | 122.3 ± 5.2 |
| HDL-C (mg/dL) | 48.5 ± 2.6 | 45.8 ± 1.8 | 47.6 ± 2.1 | 47.1 ± 1.8 |
| hsCRP (mg/L) | 0.5 ± 0.2 | 0.5 ± 0.1 | 2.5 ± 1.6 | 0.6 ± 0.1 |
| AST (IU/L) | 24.1 ± 1.2 | 24.2 ± 0.9 | 24.8 ± 1.2 | 26.9 ± 1.3 |
| ALT (IU/L) | 19.6 ± 2.2 | 21.4 ± 1.3 | 22.3 ± 1.5 | 32.6 ± 2.8 |
| WBC (×103 cells/ul) | 5.5 ± 0.2 | 6.2 ± 0.2 | 5.8 ± 0.2 | 6.2 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choe, M.K.; Kim, H.-J.; Kim, N.H.; Binas, B.; Kim, H.J. Biphasic Production of Anti-ApoB100 Autoantibodies in Obese Humans and Mice. Pharmaceuticals 2021, 14, 330. https://doi.org/10.3390/ph14040330
Choe MK, Kim H-J, Kim NH, Binas B, Kim HJ. Biphasic Production of Anti-ApoB100 Autoantibodies in Obese Humans and Mice. Pharmaceuticals. 2021; 14(4):330. https://doi.org/10.3390/ph14040330
Chicago/Turabian StyleChoe, Moon Kyung, Hyung-Ji Kim, Nan Hee Kim, Bert Binas, and Hyo Joon Kim. 2021. "Biphasic Production of Anti-ApoB100 Autoantibodies in Obese Humans and Mice" Pharmaceuticals 14, no. 4: 330. https://doi.org/10.3390/ph14040330
APA StyleChoe, M. K., Kim, H.-J., Kim, N. H., Binas, B., & Kim, H. J. (2021). Biphasic Production of Anti-ApoB100 Autoantibodies in Obese Humans and Mice. Pharmaceuticals, 14(4), 330. https://doi.org/10.3390/ph14040330