
Echinocandin Drugs Induce Differential Effects in Cytokinesis Progression and Cell Integrity

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Susceptibilities of WT and pbr1-8 Strains to Echinocandins
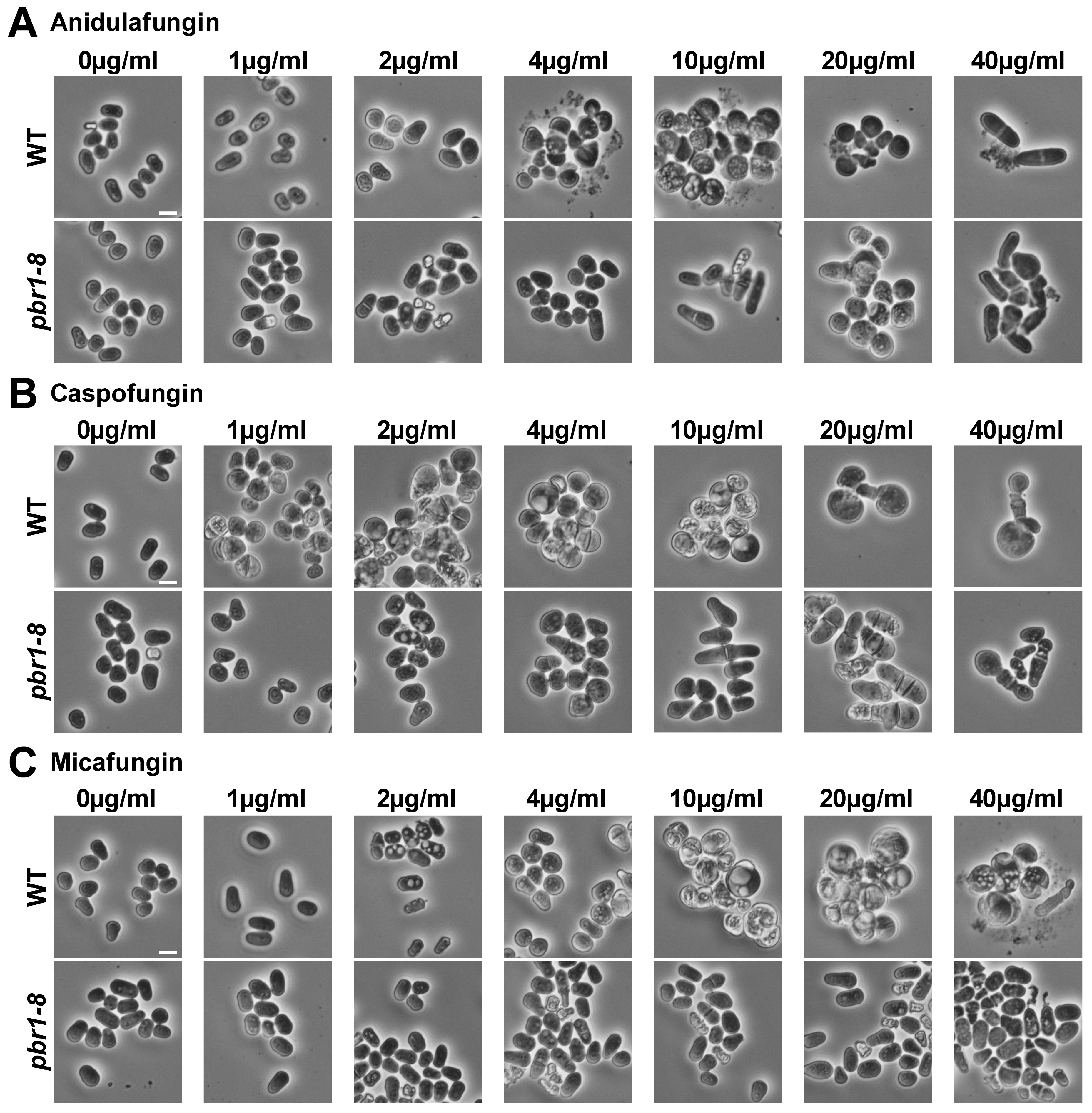
2.2. Differential Effects of the Echinocandins in Cytokinesis Progression
2.2.1. Anidulafungin
2.2.2. Caspofungin
2.2.3. Micafungin
2.3. Differential Effects of the Echinocandins in Cell Integrity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug (Concentration) | Strain | Number of Cells | Cell Death 1 | Cell Lysis 2 | Non-Cell Lysis 3 |
|---|---|---|---|---|---|
| Control (+DMSO) | WT | n = 30 | 0 4 (0) 5 | 0 4 (0) 5 | 0 4 (0) 5 |
| pbr1-8 | n = 33 | 0 (0) | 0 (0) | 0 (0) | |
| Anidulafungin (2 µg/mL) | WT | n = 79 | 89.9 (100) | 65.2 (72.5) | 24.7 (27.5) |
| pbr1-8 | n = 61 | 0 (0) | 0 (0) | 0 (0) | |
| Anidulafungin (20 µg/mL) | WT | n = 60 | 21.7 (100) | 13.3 (61.5) | 8.4 (38.5) |
| pbr1-8 | n = 45 | 8.9 (100) | 2.2 (25) | 6.7 (75) | |
| Caspofungin (2 µg/mL) | WT | n = 96 | 82.3 (100) | 58.3 (70.9) | 24 (29.1) |
| pbr1-8 | n = 85 | 22.4 (100) | 5.9 (26.3) | 16.5 (73.7) | |
| Caspofungin (20 µg/mL) | WT | n = 33 | 27.3 (100) | 12.1 (44.4) | 15.2 (55.6) |
| pbr1-8 | n = 64 | 22.7 (100) | 0 (0) | 22.7 (100) | |
| Micafungin (2 µg/mL) | WT | n = 99 | 80.3 (100) | 13.6 (17) | 69.7 (83) |
| pbr1-8 | n = 36 | 0 (0) | 0 (0) | 0 (0) | |
| Micafungin (20 µg/mL) | WT | n = 61 | 97.5 (100) | 97.5 (100) | 0 (0) |
| pbr1-8 | n = 46 | 0 (0) | 0 (0) | 0 (0) |
| Drug (Concentration) | Strain | Number of Cells | Total Cell Lysis 1 | Lysis at Separation 1 | Lysis at Interphase 1 |
|---|---|---|---|---|---|
| Control (+DMSO) | WT | n = 30 | 0 2 (0) 3 | 0 2 (0) 3 | 0 2 (0) 3 |
| pbr1-8 | n = 33 | 0 (0) | 0 (0) | 0 (0) | |
| Anidulafungin (2 µg/mL) | WT | n = 79 | 65.2 (72.5) | 45.6 (50.7) | 19.6 (21.8) |
| pbr1-8 | n = 61 | 0 (0) | 0 (0) | 0 (0) | |
| Anidulafungin (20 µg/mL) | WT | n = 60 | 13.3 (61.5) | 10.0 (46.1) | 3.3 (15.4) |
| pbr1-8 | n = 45 | 2.2 (25) | 0 (0) | 2.2 (25) | |
| Caspofungin (2 µg/mL) | WT | n = 96 | 58.3 (70.9) | 40.6 (49.4) | 17.7 (21.5) |
| pbr1-8 | n = 85 | 5.9 (26.3) | 2.4 (10.5) | 3.5 (15.8) | |
| Caspofungin (20 µg/mL) | WT | n = 33 | 12.1 (44.4) | 12.1 (44.4) | 0 (0) |
| pbr1-8 | n = 64 | 0 (0) | 0 (0) | 0 (0) | |
| Micafungin (2 µg/mL) | WT | n = 99 | 13.6 (17) | 3.0 (3.8) | 10.6 (13.2) |
| pbr1-8 | n = 36 | 0 (0) | 0 (0) | 0 (0) | |
| Micafungin (20 µg/mL) | WT | n = 61 | 97.5 (100) | 44.3 (45.4) | 53.3 (54.6) |
| pbr1-8 | n = 46 | 0 (0) | 0 (0) | 0 (0) |
3. Materials and Methods
3.1. Strains and Culture Conditions
3.2. Antifungal Drugs and Susceptibility Assays
3.3. Microscopy Techniques and Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases-estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Hoenigl, M.; Meis, J.F.; Cornely, O.A.; Muthu, V.; Gangneux, J.P.; Perfect, J.; Chakrabarti, A.; ECMM; ISHAM. ECMM/ISHAM recommendations for clinical management of COVID-19 associated mucormycosis in low- and middle-income countries. Mycoses 2021, 64, 1028–1037. [Google Scholar] [CrossRef]
- Limper, A.H.; Adenis, A.; Le, T.; Harrison, T.S. Fungal infections in HIV/AIDS. Lancet Infect. Dis. 2017, 17, e334–e343. [Google Scholar] [CrossRef]
- LIFE. Leading International Fungal Education: The Burden of Fungal Disease 2017. Available online: http://www.life-worldwide.org/media-centre/article/the-burden-of-fungal-disease-new-evidence-to-show-the-scale-of-the-problem (accessed on 3 November 2021).
- Perfect, J.R. The antifungal pipeline: A reality check. Nat. Rev. Drug Discov. 2017, 16, 603–616. [Google Scholar] [CrossRef] [Green Version]
- Ostrosky-Zeichner, L.; Casadevall, A.; Galgiani, J.N.; Odds, F.C.; Rex, J.H. An insight into the antifungal pipeline: Selected new molecules and beyond. Nat. Rev. Drug Discov. 2010, 9, 719–727. [Google Scholar] [CrossRef]
- Denning, D.W.; Bromley, M.J. Infectious Disease. How to bolster the antifungal pipeline. Science 2015, 347, 1414–1416. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Gabaldón, T.; Garcia-Rubio, R.; Jenks, J.D.; Hoenigl, M.; Salzer, H.J.F.; Ilkit, M.; Lass-Florl, C.; Perlin, D.S. Drug-resistant fungi: An emerging challenge threatening our limited antifungal armamentarium. Antibiotics 2020, 9, 877. [Google Scholar] [CrossRef]
- Ksiezopolska, E.; Schikora-Tamarit, M.A.; Beyer, R.; Nunez-Rodriguez, J.C.; Schuller, C.; Gabaldón, T. Narrow mutational signatures drive acquisition of multidrug resistance in the fungal pathogen Candida glabrata. Curr. Biol. 2021. [Google Scholar] [CrossRef]
- Cortés, J.C.G.; Curto, M.A.; Carvalho, V.S.D.; Pérez, P.; Ribas, J.C. The fungal cell wall as a target for the development of new antifungal therapies. Biotechnol. Adv. 2019, 37, 107352. [Google Scholar] [CrossRef]
- Hopke, A.; Brown, A.J.P.; Hall, R.A.; Wheeler, R.T. Dynamic Fungal Cell Wall Architecture in Stress Adaptation and Immune Evasion. Trends Microbiol. 2018, 26, 284–295. [Google Scholar] [CrossRef]
- Georgopapadakou, N.H.; Tkacz, J.S. The fungal cell wall as a drug target. Trends Microbiol. 1995, 3, 98–104. [Google Scholar] [CrossRef]
- Curto, M.A.; Butassi, E.; Ribas, J.C.; Svetaz, L.A.; Cortés, J.C.G. Natural products targeting the synthesis of β(1,3)-D-glucan and chitin of the fungal cell wall. Existing drugs and recent findings. Phytomedicine 2021, 88, 153556. [Google Scholar] [CrossRef]
- Cabib, E.; Arroyo, J. How carbohydrates sculpt cells: Chemical control of morphogenesis in the yeast cell wall. Nat. Rev. Microbiol. 2013, 11, 648–655. [Google Scholar] [CrossRef]
- Lesage, G.; Bussey, H. Cell wall assembly in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 317–343. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, V.S.D.; Gómez-Delgado, L.; Curto, M.A.; Moreno, M.B.; Pérez, P.; Ribas, J.C.; Cortés, J.C.G. Analysis and application of a suite of recombinant endo-β(1,3)-D-glucanases for studying fungal cell walls. Microb. Cell Fact. 2021, 20, 126. [Google Scholar] [CrossRef]
- Horiseberger, M.; Rosset, J. Localization of α-galactomannan on the surface of Schizosaccharomyces pombe cells by scanning electron microscopy. Arch. Microbiol. 1977, 112, 123–126. [Google Scholar] [CrossRef]
- Kreger, D.R. Observations on cell walls of yeasts and some other fungi by x-ray diffraction and solubility tests. Biochim. Biophys. Acta 1954, 13, 1–9. [Google Scholar] [CrossRef]
- Cortés, J.C.G.; Ramos, M.; Osumi, M.; Pérez, P.; Ribas, J.C. The Cell Biology of Fission Yeast Septation. Microbiol. Mol. Biol. Rev. 2016, 80, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Humbel, B.M.; Konomi, M.; Takagi, T.; Kamasawa, N.; Ishijima, S.A.; Osumi, M. In situ localization of β-glucans in the cell wall of Schizosaccharomyces pombe. Yeast 2001, 18, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Balasubramanian, M.K. 1,3-β-Glucan synthase: A useful target for antifungal drugs. Curr. Drug Targets-Infect. Disord. 2001, 1, 159–169. [Google Scholar] [CrossRef]
- Latge, J.P. The cell wall: A carbohydrate armour for the fungal cell. Mol. Microbiol. 2007, 66, 279–290. [Google Scholar] [CrossRef]
- Le Goff, X.; Woollard, A.; Simanis, V. Analysis of the cps1 gene provides evidence for a septation checkpoint in Schizosaccharomyces pombe. Mol. Gen. Genet. 1999, 262, 163–172. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Balasubramanian, M.K. A checkpoint that monitors cytokinesis in Schizosaccharomyces pombe. J. Cell Sci. 2000, 113, 1223–1230. [Google Scholar] [CrossRef]
- Ramos, M.; Cortés, J.C.G.; Sato, M.; Rincón, S.A.; Moreno, M.B.; Clemente-Ramos, J.A.; Osumi, M.; Pérez, P.; Ribas, J.C. Two S. pombe septation phases differ in ingression rate, septum structure, and response to F-actin loss. J. Cell Biol. 2019, 218, 4171–4194. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Tang, X.; Wang, H.; Balasubramanian, M. Bgs2p, a 1,3-β-glucan synthase subunit, is essential for maturation of ascospore wall in Schizosaccharomyces pombe. FEBS Lett. 2000, 478, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Roncero, C.; Sánchez, Y. Cell separation and the maintenance of cell integrity during cytokinesis in yeast: The assembly of a septum. Yeast 2010, 27, 521–530. [Google Scholar] [CrossRef]
- Martín, V.; García, B.; Carnero, E.; Durán, A.; Sánchez, Y. Bgs3p, a putative 1,3-β-glucan synthase subunit, is required for cell wall assembly in Schizosaccharomyces pombe. Eukaryot. Cell 2003, 2, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Martins, I.M.; Cortés, J.C.G.; Muñoz, J.; Moreno, M.B.; Ramos, M.; Clemente-Ramos, J.A.; Durán, A.; Ribas, J.C. Differential activities of three families of specific β(1,3)glucan synthase inhibitors in wild-type and resistant strains of fission yeast. J. Biol. Chem. 2011, 286, 3484–3496. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, J.; Cortés, J.C.G.; Sipiczki, M.; Ramos, M.; Clemente-Ramos, J.A.; Moreno, M.B.; Martins, I.M.; Pérez, P.; Ribas, J.C. Extracellular cell wall β(1,3)glucan is required to couple septation to actomyosin ring contraction. J. Cell Biol. 2013, 203, 265–282. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.F. Lysis of yeast cell walls induced by 2-deoxyglucose at their sites of glucan synthesis. J. Bacteriol. 1968, 95, 1169–1172. [Google Scholar] [CrossRef] [Green Version]
- Miyata, M.; Kitamura, J.; Miyata, H. Lysis of growing fissin-yeast cells induced by aculeacin A, a new antifungal antibiotic. Arch. Microbiol. 1980, 127, 11–16. [Google Scholar] [CrossRef]
- Cassone, A.; Mason, R.E.; Kerridge, D. Lysis of growing yeast-form cells of Candida albicans by echinocandin: A cytological study. Sabouraudia 1981, 19, 97–110. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Hiratani, T.; Baba, M.; Osumi, M. Effect of aculeacin A, a wall-active antibiotic, on synthesis of the yeast cell wall. Microbiol. Immunol. 1985, 29, 609–623. [Google Scholar] [CrossRef]
- Vicente, M.F.; Basilio, A.; Cabello, A.; Peláez, F. Microbial natural products as a source of antifungals. Clin. Microbiol. Infect. 2003, 9, 15–32. [Google Scholar] [CrossRef] [Green Version]
- Suwunnakorn, S.; Wakabayashi, H.; Kordalewska, M.; Perlin, D.S.; Rustchenko, E. FKS2 and FKS3 Genes of Opportunistic Human Pathogen Candida albicans Influence Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2018, 62, e02299-17. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vázquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef]
- Perlin, D.S. Echinocandin Resistance in Candida. Clin. Infect. Dis. 2015, 61 (Suppl. 6), S612–S617. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Messer, S.A.; Jones, R.N.; Castanheira, M. Antifungal susceptibilities of Candida, Cryptococcus neoformans and Aspergillus fumigatus from the Asia and Western Pacific region: Data from the SENTRY antifungal surveillance program (2010–2012). J. Antibiot. 2015, 68, 556–561. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.E.; Katiyar, S.K.; Edlind, T.D. New Fks hot spot for acquired echinocandin resistance in Saccharomyces cerevisiae and its contribution to intrinsic resistance of Scedosporium species. Antimicrob. Agents Chemother. 2011, 55, 3774–3781. [Google Scholar] [CrossRef] [Green Version]
- Sawistowska-Schroder, E.T.; Kerridge, D.; Perry, H. Echinocandin inhibition of 1,3-β-D-glucan synthase from Candida albicans. FEBS Lett. 1984, 173, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Douglas, C.M.; Marrinan, J.A.; Li, W.; Kurtz, M.B. A Saccharomyces cerevisiae mutant with echinocandin-resistant 1,3-β−D-glucan synthase. J. Bacteriol. 1994, 176, 5686–5696. [Google Scholar] [CrossRef] [Green Version]
- Taft, C.S.; Stark, T.; Selitrennikoff, C.P. Cilofungin (LY121019) inhibits Candida albicans (1-3)-β-D-glucan synthase activity. Antimicrob. Agents Chemother. 1988, 32, 1901–1903. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S. Resistance to echinocandin-class antifungal drugs. Drug Resist. Updates 2007, 10, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.E.; Edlind, T.D. Topological and mutational analysis of Saccharomyces cerevisiae Fks1. Eukaryot. Cell 2012, 11, 952–960. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Ortigosa, C.; Jiang, J.; Chen, M.; Kuang, X.; Healey, K.R.; Castellano, P.; Boparai, N.; Ludtke, S.J.; Perlin, D.S.; Dai, W. Preliminary structural elucidation of β-(1,3)-glucan synthase from Candida glabrata using cryo-electron tomography. J. Fungi 2021, 7, 120. [Google Scholar] [CrossRef]
- García, R.; Itto-Nakama, K.; Rodríguez-Pena, J.M.; Chen, X.; Sanz, A.B.; de Lorenzo, A.; Pavon-Verges, M.; Kubo, K.; Ohnuki, S.; Nombela, C.; et al. Poacic acid, a β-1,3-glucan-binding antifungal agent, inhibits cell-wall remodeling and activates transcriptional responses regulated by the cell-wall integrity and high-osmolarity glycerol pathways in yeast. FASEB J. 2021, 35, e21778. [Google Scholar] [CrossRef]
- Roncero, C.; Celador, R.; Sánchez, N.; García, P.; Sánchez, Y. The role of the cell integrity pathway in septum assembly in yeast. J. Fungi 2021, 7, 729. [Google Scholar] [CrossRef]
- Cortés, J.C.G.; Ramos, M.; Konomi, M.; Barragán, I.; Moreno, M.B.; Alcaide-Gavilán, M.; Moreno, S.; Osumi, M.; Pérez, P.; Ribas, J.C. Specific detection of fission yeast primary septum reveals septum and cleavage furrow ingression during early anaphase independent of mitosis completion. PLoS Genet. 2018, 14, e1007388. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Splitting of the fission yeast septum. FEMS Yeast Res. 2007, 7, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Makarova, M.; Peter, M.; Balogh, G.; Glatz, A.; MacRae, J.I.; Lopez Mora, N.; Booth, P.; Makeyev, E.; Vigh, L.; Oliferenko, S. Delineating the rules for structural adaptation of membrane-associated proteins to evolutionary changes in membrane lipidome. Curr. Biol. 2020, 30, 367–380.e368. [Google Scholar] [CrossRef] [Green Version]
- Boeck, L.D.; Fukuda, D.S.; Abbott, B.J.; Debono, M. Deacylation of echinocandin B by Actinoplanes utahensis. J. Antibiot. 1989, 42, 382–388. [Google Scholar] [CrossRef]
- Healey, K.R.; Katiyar, S.K.; Raj, S.; Edlind, T.D. CRS-MIS in Candida glabrata: Sphingolipids modulate echinocandin-Fks interaction. Mol. Microbiol. 2012, 86, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Satish, S.; Jiménez-Ortigosa, C.; Zhao, Y.; Lee, M.H.; Dolgov, E.; Kruger, T.; Park, S.; Denning, D.W.; Kniemeyer, O.; Brakhage, A.A.; et al. Stress-induced changes in the lipid microenvironment of β-(1,3)-D-glucan synthase cause clinically important echinocandin resistance in Aspergillus fumigatus. mBio 2019, 10, e00779-19. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Ohnuki, S.; Roncero, C.; Konopka, J.B.; Ohya, Y. Distinct roles of cell wall biogenesis in yeast morphogenesis as revealed by multivariate analysis of high-dimensional morphometric data. Mol. Biol. Cell 2014, 25, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Foltman, M.; Filali-Mouncef, Y.; Crespo, D.; Sánchez-Díaz, A. Cell polarity protein Spa2 coordinates Chs2 incorporation at the division site in budding yeast. PLoS Genet. 2018, 14, e1007299. [Google Scholar] [CrossRef]
- Viana, R.A.; Pinar, M.; Soto, T.; Coll, P.M.; Cansado, J.; Pérez, P. Negative functional interaction between cell integrity MAPK pathway and Rho1 GTPase in fission yeast. Genetics 2013, 195, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, A.; Cunningham, K.W. Vacuolar H+-ATPase (V-ATPase) promotes vacuolar membrane permeabilization and nonapoptotic death in stressed yeast. J. Biol. Chem. 2012, 287, 19029–19039. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Jiang, W.; Liu, Q.; Ryuko, S.; Kuno, T. Genome-wide screening for genes associated with FK506 sensitivity in fission yeast. PLoS ONE 2011, 6, e23422. [Google Scholar] [CrossRef] [Green Version]
- Alfa, C.; Fantes, P.; Hyams, J.; McLeod, M.; Warbrick, E. (Eds.) Experiments with Fission Yeast: A Laboratory Course Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1993; p. 186. [Google Scholar]




| Chemical Structure | Compound (Commercial Name and Proprietary Company) |
|---|---|
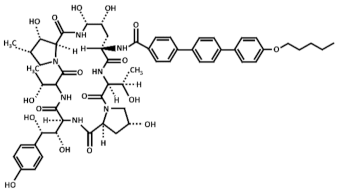 | Anidulafungin (Eraxis® or Ecalta®, Pfizer) |
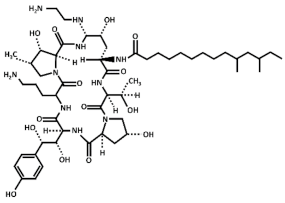 | Caspofungin (Cancidas®, Merck, Sharp and Dhome, MSD) |
 | Micafungin (Mycamine®, Astellas Pharma) |
| Lower Cell Density 2 | Higher Cell Density 3 | |
|---|---|---|
| Strain | Anidulafungin MIC (µg/mL) | |
| WT | 4 | 10 |
| pbr1-8 | 10 | 20 |
| Caspofungin MIC (µg/mL) | ||
| WT | 1 | 10 |
| pbr1-8 | 2 | 40 |
| Micafungin MIC (µg/mL) | ||
| WT | 10 | 20 |
| pbr1-8 | >80 | >80 |
| Drug (Concentration) | Cytokinesis Phenotype | Septation 1 | Cell Separation 2 | ||
|---|---|---|---|---|---|
| WT | pbr1-8 | WT | pbr1-8 | ||
| Control (+DMSO) | Normal cytokinesis | 22.4 ± 2.4 n = 30 | 20.2 ± 1.6 n = 33 | 14.5 ± 2.4 n = 27 | 16.5 ± 3.4 n = 33 |
| Anidulafungin (2 µg/mL) | Cell death 3 | 45.9 ± 10.5 n = 25 | n = 0 | 36.3 ± 6.5 n = 25 | n = 0 |
| Normal cytokinesis | n = 0 | 22.6 ± 2.3 n = 60 | n = 0 | 21.4 ± 4.6 n = 55 | |
| Slow septation Blocked separation 4 | 48.0 ± 0.0 n = 1 | n = 0 | ≥84.0 ± 0.0 n = 1 | n = 0 | |
| Anidulafungin (20 µg/mL) | Blocked cytokinesis 5 | ≥155.1 ± 24.9 n = 23 | ≥160.8 ± 15.4 n = 31 | ≥145.9 ± 36.1 n = 7 | ≥172.0 ± 0.0 n = 1 |
| Caspofungin (2 µg/mL) | Cell death 3 | 31.0 ± 4.3 n = 20 | 32.0 ± 9.0 n = 8 | 32.4 ± 6.2 n = 21 | 26.0 ± 9.2 n = 10 |
| Normal cytokinesis | n = 0 | 24.3 ± 5.2 n = 12 | n = 0 | 22.8 ± 4.9 n = 17 | |
| Blocked separation | 20.0 ± 2.9 n = 14 | 25.1 ± 4.3 n = 11 | ≥80.0 ± 26.2 n = 14 | ≥81.5 ± 36.5 n = 11 | |
| Caspofungin (20 µg/mL) | Extremely slow septation Blocked separation4 | ≥121.6 ± 26.5 n = 24 | n = 0 | ≥ 75.3 ± 28.7 n = 7 | ≥105.1 ± 23.1 n = 29 |
| Slow septation | n = 0 | 51.4 ± 8.4 n = 25 | n = 0 | n = 0 | |
| Micafungin (2 µg/mL) | Cell death 3 | 22.8 ± 1.8 n = 23 | n = 0 | 26.1 ± 6.8 n = 31 | n = 0 |
| Normal cytokinesis | n = 0 | 20.1 ± 1.3 n = 36 | n = 0 | 17.3 ± 2.7 n = 33 | |
| Blocked separation | 20.7 ± 1.8 n = 6 | n = 0 | ≥52.0 ± 21.3 n = 6 | n = 0 | |
| Micafungin (20 µg/mL) | Cell death3 | 27.6 ± 2.0 n = 11 | n = 0 | 33.7 ± 4.3 n = 12 | n = 0 |
| Normal cytokinesis | n = 0 | 19.8 ± 1.2 n = 46 | n = 0 | 25.9 ± 5.0 n = 39 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yagüe, N.; Gómez-Delgado, L.; Curto, M.Á.; Carvalho, V.S.D.; Moreno, M.B.; Pérez, P.; Ribas, J.C.; Cortés, J.C.G. Echinocandin Drugs Induce Differential Effects in Cytokinesis Progression and Cell Integrity. Pharmaceuticals 2021, 14, 1332. https://doi.org/10.3390/ph14121332
Yagüe N, Gómez-Delgado L, Curto MÁ, Carvalho VSD, Moreno MB, Pérez P, Ribas JC, Cortés JCG. Echinocandin Drugs Induce Differential Effects in Cytokinesis Progression and Cell Integrity. Pharmaceuticals. 2021; 14(12):1332. https://doi.org/10.3390/ph14121332
Chicago/Turabian StyleYagüe, Natalia, Laura Gómez-Delgado, M. Ángeles Curto, Vanessa S. D. Carvalho, M. Belén Moreno, Pilar Pérez, Juan Carlos Ribas, and Juan Carlos G. Cortés. 2021. "Echinocandin Drugs Induce Differential Effects in Cytokinesis Progression and Cell Integrity" Pharmaceuticals 14, no. 12: 1332. https://doi.org/10.3390/ph14121332
APA StyleYagüe, N., Gómez-Delgado, L., Curto, M. Á., Carvalho, V. S. D., Moreno, M. B., Pérez, P., Ribas, J. C., & Cortés, J. C. G. (2021). Echinocandin Drugs Induce Differential Effects in Cytokinesis Progression and Cell Integrity. Pharmaceuticals, 14(12), 1332. https://doi.org/10.3390/ph14121332