
Osteogenic and Chondrogenic Potential of Periosteum-Derived Mesenchymal Stromal Cells: Do They Hold the Key to the Future?

 ,
,  ,
,  ,
, 
 and
and 
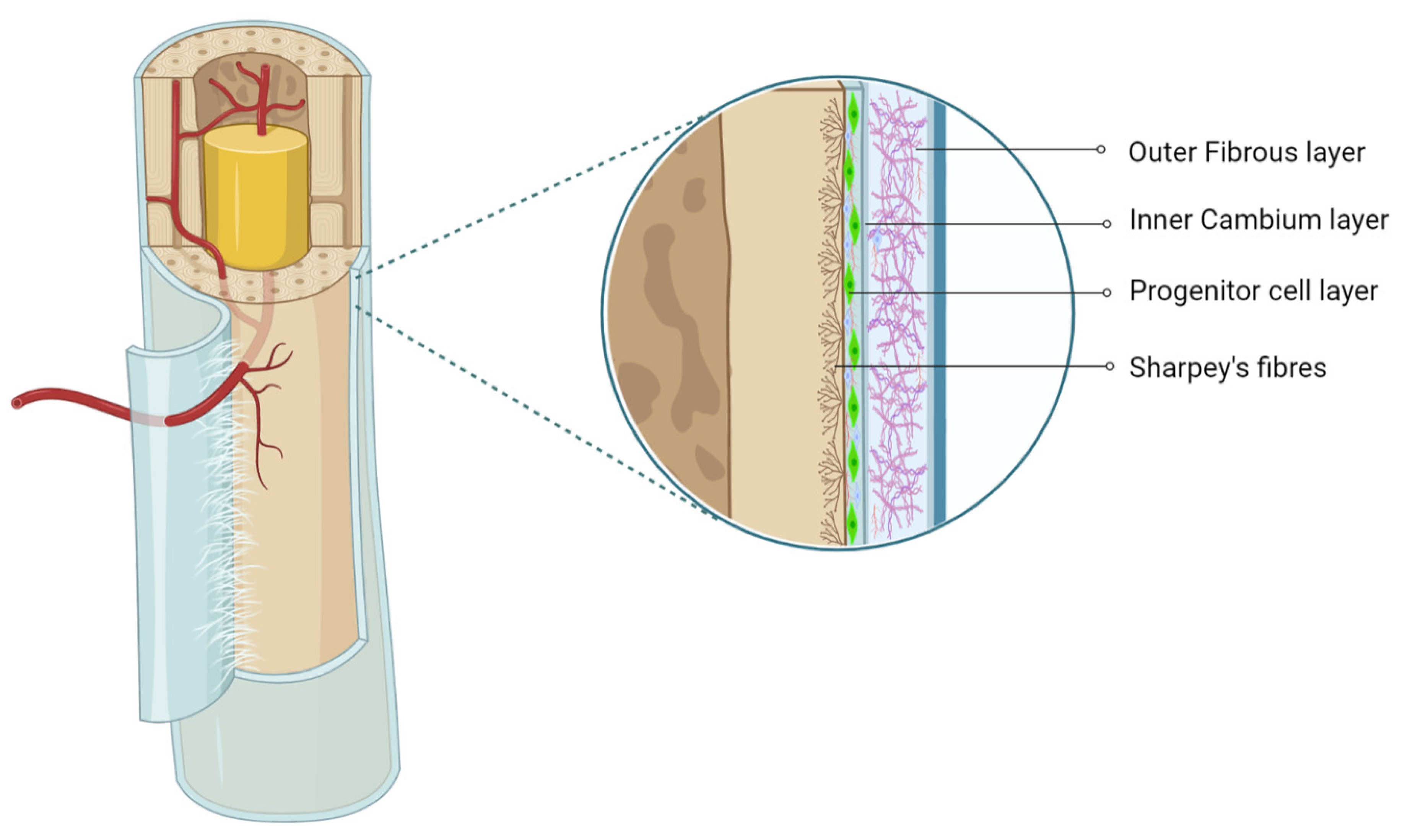{kind=link}
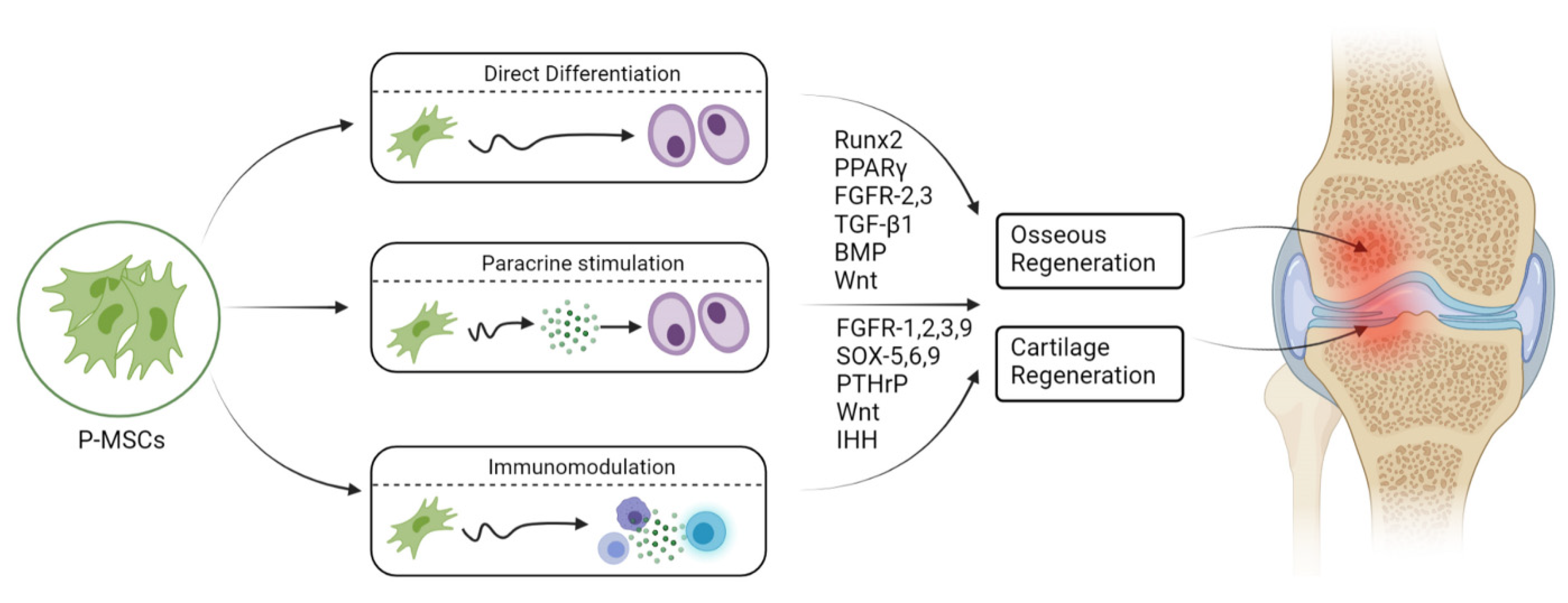{kind=link}
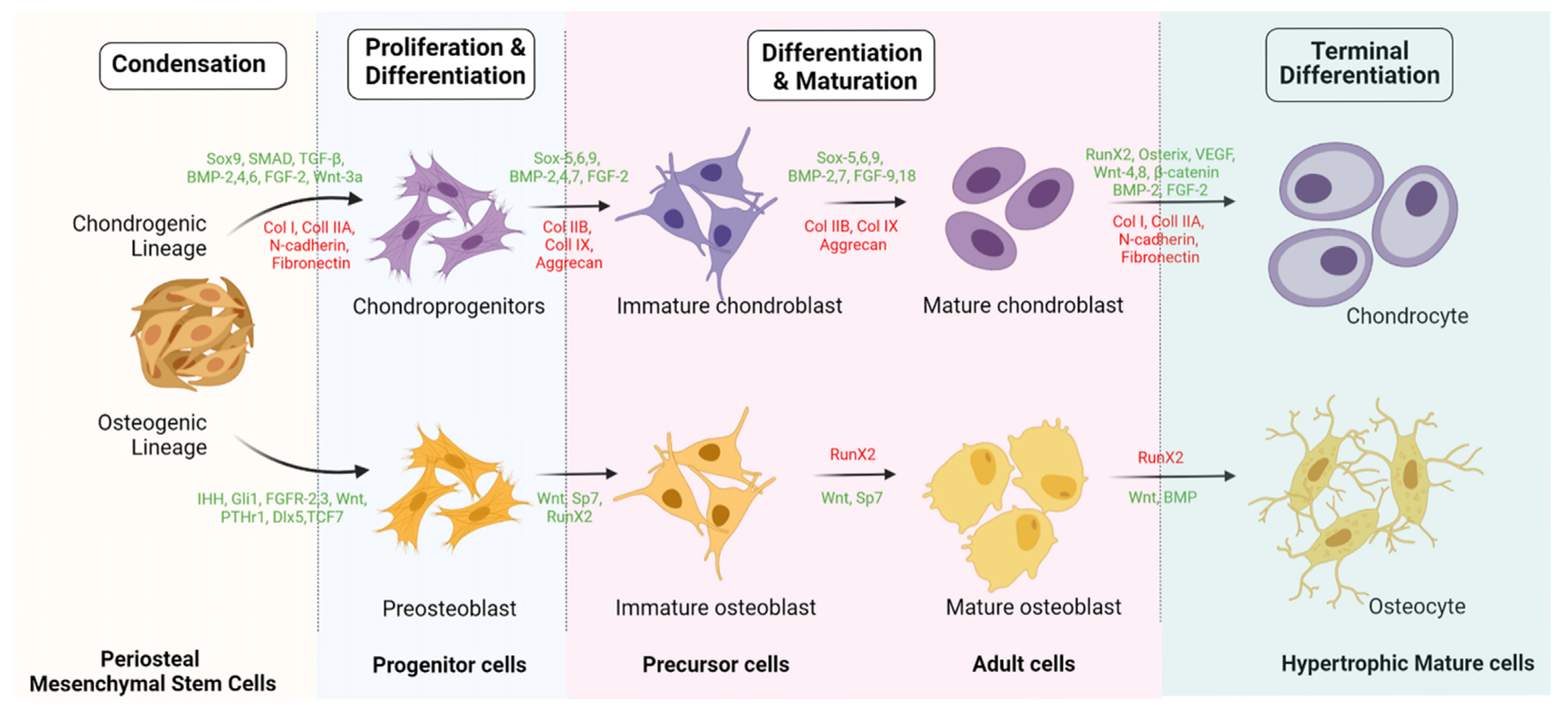{kind=link}
Abstract
1. Introduction
2. Periosteum as MSC Source
3. Characterization and Isolation of P-MSCs
4. Intracellular Signaling in Osteogenic Differentiation
5. Osteogenicity of P-MSCs
6. Engineered Osteogenesis by P-MSCs
7. Intracellular Signaling in Chondrogenic Differentiation
8. Chondrogenicity of P-MSCs
9. Engineered Chondrogenesis of P-MSCs
10. Future Perspectives
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dwek, J.R. The periosteum: What is it, where is it, and what mimics it in its absence? Skelet. Radiol. 2010, 39, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Alencar, C.H.M.F.; Silveira, C.R.S.; Cavalcante, M.M.; Vieira, C.G.M.; Teixeira, M.J.D.; Neto, F.A.; de Abreu, A.; Chhabra, A. Periosteum: An imaging review. Eur. J. Radiol. Open 2020, 7, 100249. [Google Scholar] [CrossRef]
- Egawa, S.; Miura, S.; Yokoyama, H.; Endo, T.; Tamura, K. Growth and differentiation of a long bone in limb development, repair and regeneration. Dev. Growth Differ. 2014, 56, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.F.; Chang, H.; Knothe Tate, M.L. Elucidating multiscale periosteal mechanobiology: A key to unlocking the smart properties and regenerative capacity of the periosteum? Tissue Eng. Part B Rev. 2013, 19, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A. Periosteum: A Highly Underrated Tool in Dentistry. Int. J. Dent. 2011, 2012, e717816. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, C.; Mattioli-Belmonte, M. Periosteum derived stem cells for regenerative medicine proposals: Boosting current knowledge. World J. Stem Cells 2014, 6, 266–277. [Google Scholar] [CrossRef]
- Hutmacher, D.W.; Sittinger, M. Periosteal cells in bone tissue engineering. Tissue Eng. 2003, 9 (Suppl. 1), 45–64. [Google Scholar] [CrossRef]
- Warnke, P.H.; Douglas, T.; Sivananthan, S.; Wiltfang, J.; Springer, I.; Becker, S.T. Tissue engineering of periosteal cell membranes in vitro. Clin. Oral Implants Res. 2009, 20, 761–766. [Google Scholar] [CrossRef]
- Arnsdorf, E.J.; Jones, L.M.; Carter, D.R.; Jacobs, C.R. The periosteum as a cellular source for functional tissue engineering. Tissue Eng. Part A 2009, 15, 2637–2642. [Google Scholar] [CrossRef] [PubMed]
- Colnot, C.; Zhang, X.; Knothe Tate, M.L. Current insights on the regenerative potential of the periosteum: Molecular, cellular, and endogenous engineering approaches. J. Orthop. Res. 2012, 30, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Knothe Tate, M.L. Concise review: The periosteum: Tapping into a reservoir of clinically useful progenitor cells. Stem Cells Transl. Med. 2012, 1, 480–491. [Google Scholar] [CrossRef]
- Moore, E.R.; Zhu, Y.X.; Ryu, H.S.; Jacobs, C.R. Periosteal progenitors contribute to load-induced bone formation in adult mice and require primary cilia to sense mechanical stimulation. Stem Cell Res. Ther. 2018, 9, 190. [Google Scholar] [CrossRef]
- Lin, Z.; Fateh, A.; Salem, D.M.; Intini, G. Periosteum. J. Dent. Res. 2014, 93, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Fitzsimmons, J.S.; Sanyal, A.; Mello, M.A.; Mukherjee, N.; O’Driscoll, S.W. Localization of chondrocyte precursors in periosteum. Osteoarthr. Cartil. 2001, 9, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Conway, S.J.; Izuhara, K.; Kudo, Y.; Litvin, J.; Markwald, R.; Ouyang, G.; Arron, J.R.; Holweg, C.T.J.; Kudo, A. The role of periostin in tissue remodeling across health and disease. Cell. Mol. Life Sci. 2014, 71, 1279–1288. [Google Scholar] [CrossRef]
- Xiao, H.; Wang, L.; Zhang, T.; Chen, C.; Chen, H.; Li, S.; Hu, J.; Lu, H. Periosteum progenitors could stimulate bone regeneration in aged murine bone defect model. J. Cell. Mol. Med. 2020, 24, 12199–12210. [Google Scholar] [CrossRef]
- Gnecchi, M.; Zhang, Z.; Ni, A.; Dzau, V.J. Paracrine mechanisms in adult stem cell signaling and therapy. Circ. Res. 2008, 103, 1204–1219. [Google Scholar] [CrossRef] [PubMed]
- Baraniak, P.R.; McDevitt, T.C. Stem cell paracrine actions and tissue regeneration. Regen. Med. 2010, 5, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Maleki, M.; Ghanbarvand, F.; Reza Behvarz, M.; Ejtemaei, M.; Ghadirkhomi, E. Comparison of Mesenchymal Stem Cell Markers in Multiple Human Adult Stem Cells. Int. J. Stem Cells 2014, 7, 118–126. [Google Scholar] [CrossRef]
- Lv, F.-J.; Tuan, R.S.; Cheung, K.M.C.; Leung, V.Y.L. Concise Review: The Surface Markers and Identity of Human Mesenchymal Stem Cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-S.; Xin, Z.-C.; Dai, J.; Lue, T.F. Commonly used mesenchymal stem cell markers and tracking labels: Limitations and challenges. Histol. Histopathol. 2013, 28, 1109–1116. [Google Scholar] [CrossRef]
- Olivares-Navarrete, R.; Rodil, S.E.; Hyzy, S.L.; Dunn, G.R.; Almaguer-Flores, A.; Schwartz, Z.; Boyan, B.D. Role of Integrin Subunits in Mesenchymal Stem Cell Differentiation and Osteoblast Maturation on Graphitic Carbon-coated Microstructured Surfaces. Biomaterials 2015, 51, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Krebsbach, P.H.; Villa-Diaz, L.G. The Role of Integrin α6 (CD49f) in Stem Cells: More than a Conserved Biomarker. Stem Cells Dev. 2017, 26, 1090–1099. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Roberts, A.I.; Shi, Y. Adhesion molecules: Key players in Mesenchymal stem cell-mediated immunosuppression. Cell Adhes. Migr. 2011, 5, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, F.; Voci, A.; Carminati, E.; Gualandi, F.; Mancardi, G.; Uccelli, A.; Vergani, L. Human mesenchymal stem cells target adhesion molecules and receptors involved in T cell extravasation. Stem Cell Res. Ther. 2015, 6, 245. [Google Scholar] [CrossRef]
- Machado, C.D.V.; Telles, P.D.D.S.; Nascimento, I.L.O. Immunological characteristics of mesenchymal stem cells. Rev. Bras. Hematol. E Hemoter. 2013, 35, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, E.M.; Andreef, M.; Frassoni, F. Mesenchymal Stromal Cells. Curr. Opin. Hematol. 2006, 13, 419–425. [Google Scholar] [CrossRef]
- Frey, S.P.; Jansen, H.; Doht, S.; Filgueira, L.; Zellweger, R. Immunohistochemical and molecular characterization of the human periosteum. Sci. World J. 2013, 2013, 341078. [Google Scholar] [CrossRef]
- Yang, H.; Sun, L.; Cai, W.; Gu, J.; Xu, D.; Deb, A.; Duan, J. DDR2, a discoidin domain receptor, is a marker of periosteal osteoblast and osteoblast progenitors. J. Bone Miner. Metab. 2020, 38, 670–677. [Google Scholar] [CrossRef]
- Deveza, L.; Ortinau, L.; Lei, K.; Park, D. Comparative analysis of gene expression identifies distinct molecular signatures of bone marrow- and periosteal-skeletal stem/progenitor cells. PLoS ONE 2018, 13, e0190909. [Google Scholar] [CrossRef] [PubMed]
- Ortinau, L.C.; Wang, H.; Lei, K.; Deveza, L.; Jeong, Y.; Hara, Y.; Grafe, I.; Rosenfeld, S.B.; Lee, D.; Lee, B.; et al. Identification of Functionally Distinct Mx1+αSMA+ Periosteal Skeletal Stem Cells. Cell Stem Cell 2019, 25, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Deng, R.; Chai, Y.; Chen, H.; Hu, B.; Wang, X.; Zhu, S.; Cao, Y.; Ni, S.; Wan, M.; et al. Macrophage-lineage TRAP+ cells recruit periosteum-derived cells for periosteal osteogenesis and regeneration. J. Clin. Investig. 2019, 129, 2578–2594. [Google Scholar] [CrossRef] [PubMed]
- Groeneveldt, L.C.; Herpelinck, T.; Maréchal, M.; Politis, C.; van IJcken, W.F.J.; Huylebroeck, D.; Geris, L.; Mulugeta, E.; Luyten, F.P. The Bone-Forming Properties of Periosteum-Derived Cells Differ Between Harvest Sites. Front. Cell Dev. Biol. 2020, 8, 554984. [Google Scholar] [CrossRef]
- Olbrich, M.; Rieger, M.; Reinert, S.; Alexander, D. Isolation of Osteoprogenitors from Human Jaw Periosteal Cells: A Comparison of Two Magnetic Separation Methods. PLoS ONE 2012, 7, e47176. [Google Scholar] [CrossRef]
- De Bari, C.; Dell’Accio, F.; Vanlauwe, J.; Eyckmans, J.; Khan, I.M.; Archer, C.W.; Jones, E.A.; McGonagle, D.; Mitsiadis, T.A.; Pitzalis, C.; et al. Mesenchymal multipotency of adult human periosteal cells demonstrated by single-cell lineage analysis. Arthritis Rheum. 2006, 54, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Potier, E.; Ferreira, E.; Andriamanalijaona, R.; Pujol, J.-P.; Oudina, K.; Logeart-Avramoglou, D.; Petite, H. Hypoxia affects mesenchymal stromal cell osteogenic differentiation and angiogenic factor expression. Bone 2007, 40, 1078–1087. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Deng, F.; Wang, L.; Xiang, X.-R.; Zhou, W.-W.; Hu, N.; Xu, L. Hypoxia induces osteogenesis-related activities and expression of core binding factor α1 in mesenchymal stem cells. Tohoku J. Exp. Med. 2011, 224, 7–12. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yu, X.; Wan, Q.; Ye, X.; Cheng, Y.; Pathak, J.L.; Li, Z. Cellular hypoxia promotes osteogenic differentiation of mesenchymal stem cells and bone defect healing via STAT3 signaling. Cell. Mol. Biol. Lett. 2019, 24, 64. [Google Scholar] [CrossRef]
- Cheng, S.-L.; Shao, J.-S.; Charlton-Kachigian, N.; Loewy, A.P.; Towler, D.A. Msx2 Promotes Osteogenesis and Suppresses Adipogenic Differentiation of Multipotent Mesenchymal Progenitors. J. Biol. Chem. 2003, 278, 45969–45977. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shi, K.; Andersen, T.L.; Qiu, W.; Kassem, M. KIAA1199 is a secreted molecule that enhances osteoblastic stem cell migration and recruitment. Cell Death Dis. 2019, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, C.; Miao, X.; Wu, Y.; Yuan, J.; Ding, M.; Li, J.; Shi, Z. Age-Related Insulin-Like Growth Factor Binding Protein-4 Overexpression Inhibits Osteogenic Differentiation of Rat Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2017, 42, 640–650. [Google Scholar] [CrossRef]
- Wang, J.; Li, G.; Hu, L.; Yan, F.; Zhao, B.; Wu, X.; Zhang, C.; Wang, J.; Du, J.; Wang, S. Retinoic Acid Signal Negatively Regulates Osteo/Odontogenic Differentiation of Dental Pulp Stem Cells. Stem Cells Int. 2020, 2020, 5891783. [Google Scholar] [CrossRef]
- Salasznyk, R.M.; Williams, W.A.; Boskey, A.; Batorsky, A.; Plopper, G.E. Adhesion to Vitronectin and Collagen I Promotes Osteogenic Differentiation of Human Mesenchymal Stem Cells. J. Biomed. Biotechnol. 2004, 2004, 24–34. [Google Scholar] [CrossRef]
- Suehiro, F.; Nishimura, M.; Kawamoto, T.; Kanawa, M.; Yoshizawa, Y.; Murata, H.; Kato, Y. Impact of zinc fingers and homeoboxes 3 on the regulation of mesenchymal stem cell osteogenic differentiation. Stem Cells Dev. 2011, 20, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Gong, Y.; Xu, L.; Zhou, M.; Li, J.; Song, J. Bidirectional regulation of osteogenic differentiation by the FOXO subfamily of Forkhead transcription factors in mammalian MSCs. Cell Prolif. 2018, 52, e12540. [Google Scholar] [CrossRef]
- Baglìo, S.R.; Devescovi, V.; Granchi, D.; Baldini, N. MicroRNA expression profiling of human bone marrow mesenchymal stem cells during osteogenic differentiation reveals Osterix regulation by miR-31. Gene 2013, 527, 321–331. [Google Scholar] [CrossRef]
- Li, G.; Han, N.; Yang, H.; Wang, L.; Lin, X.; Diao, S.; Du, J.; Dong, R.; Wang, S.; Fan, Z. Homeobox C10 inhibits the osteogenic differentiation potential of mesenchymal stem cells. Connect. Tissue Res. 2018, 59, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Qu, B.; Liu, O.; Fang, X.; Zhang, H.; Wang, Y.; Quan, H.; Zhang, J.; Zhou, J.; Zuo, J.; Tang, J.; et al. Distal-less homeobox 2 promotes the osteogenic differentiation potential of stem cells from apical papilla. Cell Tissue Res. 2014, 357, 133–143. [Google Scholar] [CrossRef]
- Yang, H.; Fan, J.; Cao, Y.; Gao, R.; Fan, Z. Distal-less homeobox 5 promotes the osteo-/dentinogenic differentiation potential of stem cells from apical papilla by activating histone demethylase KDM4B through a positive feedback mechanism. Exp. Cell Res. 2019, 374, 221–230. [Google Scholar] [CrossRef]
- Gao, R.-T.; Zhan, L.-P.; Meng, C.; Zhang, N.; Chang, S.-M.; Yao, R.; Li, C. Homeobox B7 promotes the osteogenic differentiation potential of mesenchymal stem cells by activating RUNX2 and transcript of BSP. Int. J. Clin. Exp. Med. 2015, 8, 10459–10470. [Google Scholar]
- Diao, S.; Yang, D.-M.; Dong, R.; Wang, L.-P.; Wang, J.-S.; Du, J.; Wang, S.-L.; Fan, Z. Enriched trimethylation of lysine 4 of histone H3 of WDR63 enhanced osteogenic differentiation potentials of stem cells from apical papilla. J. Endod. 2015, 41, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Jia, L.; Li, X.; Guo, R.; Huang, Y.; Zheng, Y.; Li, W. Long Noncoding RNAs: New Players in the Osteogenic Differentiation of Bone Marrow- and Adipose-Derived Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2018, 14, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Wei, W.; Zhao, B.; Guo, X.; Liu, S. Long non-coding RNA HOTAIR inhibits miR-17-5p to regulate osteogenic differentiation and proliferation in non-traumatic osteonecrosis of femoral head. PLoS ONE 2017, 12, e0169097. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, Y.; Zhang, Y.; Ma, L.; Lin, L.; Meng, J.; Jiang, L.; Wang, L.; Zhou, P.; Zhang, Y. LncRNA MEG3 inhibited osteogenic differentiation of bone marrow mesenchymal stem cells from postmenopausal osteoporosis by targeting miR-133a-3p. Biomed. Pharmacother. 2017, 89, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Hong, J.M.; Kang, J.A.; Rhie, J.-W.; Jeong, Y.H.; Cho, D.-W. Regulation of osteogenic differentiation of human adipose-derived stem cells by controlling electromagnetic field conditions. Exp. Mol. Med. 2013, 45, e6. [Google Scholar] [CrossRef] [PubMed]
- Wall, I.; Donos, N.; Carlqvist, K.; Jones, F.; Brett, P. Modified titanium surfaces promote accelerated osteogenic differentiation of mesenchymal stromal cells in vitro. Bone 2009, 45, 17–26. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Nakata, H.; Yamamoto, M.; Miyasaka, M.; Kasugai, S.; Kuroda, S. Osteogenic Potential of Mouse Periosteum-Derived Cells Sorted for CD90 In Vitro and In Vivo. Stem Cells Transl. Med. 2016, 5, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Xie, Y.; Xian, C.J.; Chen, L. Role of FGFs/FGFRs in skeletal development and bone regeneration. J. Cell. Physiol. 2012, 227, 3731–3743. [Google Scholar] [CrossRef]
- Dishowitz, M.I.; Terkhorn, S.P.; Bostic, S.A.; Hankenson, K.D. Notch signaling components are upregulated during both endochondral and intramembranous bone regeneration. J. Orthop. Res. 2012, 30, 296–303. [Google Scholar] [CrossRef] [PubMed]
- James, A.W. Review of Signaling Pathways Governing MSC Osteogenic and Adipogenic Differentiation. Scientifica 2013, 2013, e684736. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Kim, H.J.; Li, Q.L.; Chi, X.Z.; Ueta, C.; Komori, T.; Wozney, J.M.; Kim, E.G.; Choi, J.Y.; Ryoo, H.M.; et al. Runx2 is a common target of transforming growth factor beta1 and bone morphogenetic protein 2, and cooperation between Runx2 and Smad5 induces osteoblast-specific gene expression in the pluripotent mesenchymal precursor cell line C2C12. Mol. Cell. Biol. 2000, 20, 8783–8792. [Google Scholar] [CrossRef]
- Pratap, J.; Wixted, J.J.; Gaur, T.; Zaidi, S.K.; Dobson, J.; Gokul, K.D.; Hussain, S.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Runx2 transcriptional activation of Indian Hedgehog and a downstream bone metastatic pathway in breast cancer cells. Cancer Res. 2008, 68, 7795–7802. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.A.; Bialek, P.; Ahn, J.D.; Starbuck, M.; Patel, M.S.; Clevers, H.; Taketo, M.M.; Long, F.; McMahon, A.P.; Lang, R.A.; et al. Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev. Cell 2005, 8, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Maeda, K.; Ishihara, A.; Uehara, S.; Kobayashi, Y. Regulatory mechanism of osteoclastogenesis by RANKL and Wnt signals. Front. Biosci 2011, 16, 21–30. [Google Scholar] [CrossRef]
- Papapoulos, S.E. Targeting sclerostin as potential treatment of osteoporosis. Ann. Rheum. Dis. 2011, 70 (Suppl. 1), i119–i122. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Liu, X.; Wang, J.; Chen, X.; Zhang, H.; Kim, S.H.; Cui, J.; Li, R.; Zhang, W.; Kong, Y.; et al. Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther. Adv. Musculoskelet. Dis. 2013, 5, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yin, Y.; Li, W.; Zhao, X.; Yu, Y.; Zhu, J.; Qin, Z.; Wang, Q.; Wang, K.; Lu, W.; et al. Over-expression of PDGFR-β promotes PDGF-induced proliferation, migration, and angiogenesis of EPCs through PI3K/Akt signaling pathway. PLoS ONE 2012, 7, e30503. [Google Scholar] [CrossRef]
- Cheng, T.L.; Cantrill, L.C.; Schindeler, A.; Little, D.G. Induction of periosteal bone formation by intraosseous BMP-2 injection in a mouse model of osteogenesis imperfecta. J. Child. Orthop. 2019, 13, 543–550. [Google Scholar] [CrossRef]
- Seeherman, H.J.; Li, X.J.; Smith, E.; Parkington, J.; Li, R.; Wozney, J.M. Intraosseous injection of rhBMP-2/calcium phosphate matrix improves bone structure and strength in the proximal aspect of the femur in chronic ovariectomized nonhuman primates. J. Bone Joint Surg. 2013, 95, 36–47. [Google Scholar] [CrossRef]
- Shaw, A.T.; Maeda, Y.; Gravallese, E.M. IL-17A deficiency promotes periosteal bone formation in a model of inflammatory arthritis. Arthritis Res. Ther. 2016, 18, 104. [Google Scholar] [CrossRef] [PubMed]
- Claudel, M.; Jouzeau, J.-Y.; Cailotto, F. Secreted Frizzled-related proteins (sFRPs) in osteo-articular diseases: Much more than simple antagonists of Wnt signaling? FEBS J. 2019, 286, 4832–4851. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-S.; Lin, C.-L.; Chen, Y.-J.; Wang, C.-J.; Yang, K.D.; Huang, Y.-T.; Sun, Y.-C.; Huang, H.-C. Secreted frizzled-related protein 1 modulates glucocorticoid attenuation of osteogenic activities and bone mass. Endocrinology 2005, 146, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, W.; Osugi, M.; Kawai, T.; Hibi, H. Secreted Frizzled-Related Protein Promotes Bone Regeneration by Human Bone Marrow-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2015, 16, 23250–23258. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Takahashi, T.; Funaki, K.; Yamashita, Y. Periosteal Expansion Osteogenesis Using Highly Purified Beta-Tricalcium Phosphate Blocks: A Pilot Study in Dogs. J. Periodontol. 2008, 79, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- van Gastel, N.; Torrekens, S.; Roberts, S.J.; Moermans, K.; Schrooten, J.; Carmeliet, P.; Luttun, A.; Luyten, F.P.; Carmeliet, G. Engineering vascularized bone: Osteogenic and proangiogenic potential of murine periosteal cells. Stem Cells 2012, 30, 2460–2471. [Google Scholar] [CrossRef]
- Zheng, Y.; Ringe, J.; Liang, Z.; Loch, A.; Chen, L.; Sittinger, M. Osteogenic potential of human periosteum-derived progenitor cells in PLGA scaffold using allogeneic serum. J. Zhejiang Univ. Sci. B 2006, 7, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, H.; Goldberg, V.M.; Caplan, A.I. Culture-expanded human periosteal-derived cells exhibit osteochondral potential in vivo. J. Orthop. Res. 1991, 9, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Xue, M.; Chen, H.; Jiao, J.; Herschman, H.R.; O’Keefe, R.J.; Zhang, X. The Spatiotemporal Role of COX-2 in Osteogenic and Chondrogenic Differentiation of Periosteum-Derived Mesenchymal Progenitors in Fracture Repair. PLoS ONE 2014, 9, e100079. [Google Scholar] [CrossRef][Green Version]
- Collette, N.M.; Yee, C.S.; Hum, N.R.; Murugesh, D.K.; Christiansen, B.A.; Xie, L.; Economides, A.N.; Manilay, J.O.; Robling, A.G.; Loots, G.G. Sostdc1 deficiency accelerates fracture healing by promoting the expansion of periosteal mesenchymal stem cells. Bone 2016, 88, 20–30. [Google Scholar] [CrossRef]
- Kurenkova, A.D.; Medvedeva, E.V.; Newton, P.T.; Chagin, A.S. Niches for Skeletal Stem Cells of Mesenchymal Origin. Front. Cell Dev. Biol. 2020, 8, 592. [Google Scholar] [CrossRef] [PubMed]
- Feigenson, M.; Jonason, J.H.; Shen, J.; Loiselle, A.E.; Awad, H.A.; O’Keefe, R.J. Inhibition of the Prostaglandin EP-1 Receptor in Periosteum Progenitor Cells Enhances Osteoblast Differentiation and Fracture Repair. Ann. Biomed. Eng. 2020, 48, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Bravo, D.; Josephson, A.; Bradaschia-Correa, V.; Wong, M.; Yim, N.; Neibart, S.; Lee, S.; Huo, J.; Coughlin, T.; Mizrahi, M.; et al. Temporary inhibition of the plasminogen activator inhibits periosteal chondrogenesis and promotes periosteal osteogenesis during appendicular bone fracture healing. Bone 2018, 112, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Demol, J.; Eyckmans, J.; Roberts, S.J.; Luyten, F.P.; Van Oosterwyck, H. Does tranexamic acid stabilised fibrin support the osteogenic differentiation of human periosteum derived cells? Eur. Cells Mater. 2011, 21, 272–285. [Google Scholar] [CrossRef]
- Yoon, D.-K.; Park, J.-S.; Rho, G.-J.; Lee, H.-J.; Sung, I.-Y.; Son, J.-H.; Park, B.-W.; Kang, Y.-H.; Byun, S.-H.; Hwang, S.-C.; et al. The involvement of histone methylation in osteoblastic differentiation of human periosteum-derived cells cultured in vitro under hypoxic conditions. Cell Biochem. Funct. 2017, 35, 441–452. [Google Scholar] [CrossRef]
- Park, J.-H.; Park, B.-W.; Kang, Y.-H.; Byun, S.-H.; Hwang, S.-C.; Kim, D.R.; Woo, D.K.; Byun, J.-H. Lin28a enhances in vitro osteoblastic differentiation of human periosteum-derived cells. Cell Biochem. Funct. 2017, 35, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Okamura, H.; Nakashima, Y.; Haneji, T. Histone demethylase Jmjd3 regulates osteoblast differentiation via transcription factors Runx2 and osterix. J. Biol. Chem. 2013, 288, 33530–33541. [Google Scholar] [CrossRef]
- Agger, K.; Cloos, P.A.C.; Christensen, J.; Pasini, D.; Rose, S.; Rappsilber, J.; Issaeva, I.; Canaani, E.; Salcini, A.E.; Helin, K. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature 2007, 449, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Youngstrom, D.W.; Dishowitz, M.I.; Bales, C.B.; Carr, E.; Mutyaba, P.L.; Kozloff, K.M.; Shitaye, H.; Hankenson, K.D.; Loomes, K.M. Jagged1 expression by osteoblast-lineage cells regulates trabecular bone mass and periosteal expansion in mice. Bone 2016, 91, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Xing, Q.; Qian, Z.; Kannan, B.; Tahtinen, M.; Zhao, F. Osteogenic Differentiation Evaluation of an Engineered Extracellular Matrix Based Tissue Sheet for Potential Periosteum Replacement. ACS Appl. Mater. Interfaces 2015, 7, 23239–23247. [Google Scholar] [CrossRef]
- Kang, Y.; Ren, L.; Yang, Y. Engineering vascularized bone grafts by integrating a biomimetic periosteum and β-TCP scaffold. ACS Appl. Mater. Interfaces 2014, 6, 9622–9633. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Li, M.; Chen, K.; Fang, B.; Chen, P.; Tang, Z.; Lin, X.; Fan, S. Periosteal matrix-derived hydrogel promotes bone repair through an early immune regulation coupled with enhanced angio-and osteogenesis. Biomaterials 2020, 227, 119552. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Topol, L.; Lee, H.; Wu, J. Wnt5a and Wnt5b exhibit distinct activities in coordinating chondrocyte proliferation and differentiation. Dev. Camb. Engl. 2003, 130, 1003–1015. [Google Scholar] [CrossRef]
- Ryu, J.-H.; Chun, J.-S. Opposing roles of WNT-5A and WNT-11 in interleukin-1beta regulation of type II collagen expression in articular chondrocytes. J. Biol. Chem. 2006, 281, 22039–22047. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, M.I.; Kapadia, R.M.; Liao, Z.; Naski, M.C. The Wnt-inducible transcription factor Twist1 inhibits chondrogenesis. J. Biol. Chem. 2006, 281, 1381–1388. [Google Scholar] [CrossRef]
- Clancy, R.M.; Gomez, P.F.; Abramson, S.B. Nitric oxide sustains nuclear factor kappaB activation in cytokine-stimulated chondrocytes. Osteoarthr. Cartil. 2004, 12, 552–558. [Google Scholar] [CrossRef]
- Gibson, J.S.; Milner, P.I.; White, R.; Fairfax, T.P.A.; Wilkins, R.J. Oxygen and reactive oxygen species in articular cartilage: Modulators of ionic homeostasis. Pflüg. Arch.-Eur. J. Physiol. 2008, 455, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Shi, Z.; Liu, A.; Lin, T.; Bi, F.; Shi, M.; Yan, S.-G. Effects of cartilage oligomeric matrix protein on bone morphogenetic protein-2-induced differentiation of mesenchymal stem cells. Orthop. Surg. 2014, 6, 280–287. [Google Scholar] [CrossRef]
- Guérit, D.; Brondello, J.-M.; Chuchana, P.; Philipot, D.; Toupet, K.; Bony, C.; Jorgensen, C.; Noël, D. FOXO3A regulation by miRNA-29a Controls chondrogenic differentiation of mesenchymal stem cells and cartilage formation. Stem Cells Dev. 2014, 23, 1195–1205. [Google Scholar] [CrossRef]
- Duan, L.; Liang, Y.; Xu, X.; Xiao, Y.; Wang, D. Recent progress on the role of miR-140 in cartilage matrix remodelling and its implications for osteoarthritis treatment. Arthritis Res. Ther. 2020, 22, 194. [Google Scholar] [CrossRef]
- Yang, B.; Guo, H.; Zhang, Y.; Chen, L.; Ying, D.; Dong, S. MicroRNA-145 regulates chondrogenic differentiation of mesenchymal stem cells by targeting Sox9. PLoS ONE 2011, 6, e21679. [Google Scholar] [CrossRef]
- Xu, J.; Kang, Y.; Liao, W.; Yu, L. MiR-194 Regulates Chondrogenic Differentiation of Human Adipose-Derived Stem Cells by Targeting Sox5. PLoS ONE 2012, 7, e31861. [Google Scholar] [CrossRef]
- Lin, E.A.; Kong, L.; Bai, X.-H.; Luan, Y.; Liu, C.-J. miR-199a, a bone morphogenic protein 2-responsive MicroRNA, regulates chondrogenesis via direct targeting to Smad1. J. Biol. Chem. 2009, 284, 11326–11335. [Google Scholar] [CrossRef] [PubMed]
- Portron, S.; Hivernaud, V.; Merceron, C.; Lesoeur, J.; Masson, M.; Gauthier, O.; Vinatier, C.; Beck, L.; Guicheux, J. Inverse regulation of early and late chondrogenic differentiation by oxygen tension provides cues for stem cell-based cartilage tissue engineering. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 35, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, L.; Arnhold, S.; Brixius, K.; Addicks, K.; Bloch, W. Human mesenchymal stem cells: Influence of oxygen pressure on proliferation and chondrogenic differentiation in fibrin glue in vitro. J. Biomed. Mater. Res. Part A 2010, 93, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Campo, G.M.; Micali, A.; Avenoso, A.; D’Ascola, A.; Scuruchi, M.; Pisani, A.; Bruschetta, A.; Calatroni, A.; Puzzolo, D.; Campo, S. Inhibition of small HA fragment activity and stimulation of A2A adenosine receptor pathway limit apoptosis and reduce cartilage damage in experimental arthritis. Histochem. Cell Biol. 2015, 143. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Yang, H.N.; Woo, D.G.; Jeon, S.Y.; Do, H.-J.; Lim, H.-Y.; Kim, J.-H.; Park, K.-H. Chondrogenesis of human mesenchymal stem cells mediated by the combination of SOX trio SOX5, 6, and 9 genes complexed with PEI-modified PLGA nanoparticles. Biomaterials 2011, 32, 3679–3688. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.Y.; Park, J.S.; Yang, H.N.; Woo, D.G.; Park, K.-H. Co-delivery of SOX9 genes and anti-Cbfa-1 siRNA coated onto PLGA nanoparticles for chondrogenesis of human MSCs. Biomaterials 2012, 33, 4413–4423. [Google Scholar] [CrossRef]
- Choi, Y.-S.; Lim, S.-M.; Shin, H.-C.; Lee, C.-W.; Kim, S.-L.; Kim, D.-I. Chondrogenesis of human periosteum-derived progenitor cells in atelocollagen. Biotechnol. Lett. 2007, 29, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Hall, B.K. Chondrogenic cell differentiation from membrane bone periostea. Anat. Embryol. 1997, 196, 349–362. [Google Scholar] [CrossRef]
- O’Driscoll, S.W.; Saris, D.B.; Ito, Y.; Fitzimmons, J.S. The chondrogenic potential of periosteum decreases with age. J. Orthop. Res. 2001, 19, 95–103. [Google Scholar] [CrossRef]
- Hanada, K.; Solchaga, L.A.; Caplan, A.I.; Hering, T.M.; Goldberg, V.M.; Yoo, J.U.; Johnstone, B. BMP-2 induction and TGF-β1 modulation of rat periosteal cell chondrogenesis. J. Cell. Biochem. 2001, 81, 284–294. [Google Scholar] [CrossRef]
- Reinholz, G.G.; Fitzsimmons, J.S.; Casper, M.E.; Ruesink, T.J.; Chung, H.W.; Schagemann, J.C.; O’Driscoll, S.W. Rejuvenation of periosteal chondrogenesis using local growth factor injection. Osteoarthr. Cartil. 2009, 17, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Im, G.-I.; Quan, Z. The effects of Wnt inhibitors on the chondrogenesis of human mesenchymal stem cells. Tissue Eng. Part A 2010, 16, 2405–2413. [Google Scholar] [CrossRef]
- Gooding, C.R.; Bartlett, W.; Bentley, G.; Skinner, J.A.; Carrington, R.; Flanagan, A. A prospective, randomised study comparing two techniques of autologous chondrocyte implantation for osteochondral defects in the knee: Periosteum covered versus type I/III collagen covered. Knee 2006, 13, 203–210. [Google Scholar] [CrossRef]
- Brittberg, M.; Sjögren-Jansson, E.; Thornemo, M.; Faber, B.; Tarkowski, A.; Peterson, L.; Lindahl, A. Clonal growth of human articular cartilage and the functional role of the periosteum in chondrogenesis. Osteoarthr. Cartil. 2005, 13, 146–153. [Google Scholar] [CrossRef] [PubMed]
- De Bari, C.; Dell’Accio, F.; Luyten, F.P. Human periosteum-derived cells maintain phenotypic stability and chondrogenic potential throughout expansion regardless of donor age. Arthritis Rheum. 2001, 44, 85–95. [Google Scholar] [CrossRef]
- Bian, L.; Zhai, D.Y.; Tous, E.; Rai, R.; Mauck, R.L.; Burdick, J.A. Enhanced MSC chondrogenesis following delivery of TGF-β3 from alginate microspheres within hyaluronic acid hydrogels in vitro and in vivo. Biomaterials 2011, 32, 6425–6434. [Google Scholar] [CrossRef] [PubMed]
- Koga, H.; Muneta, T.; Nagase, T.; Nimura, A.; Ju, Y.-J.; Mochizuki, T.; Sekiya, I. Comparison of mesenchymal tissues-derived stem cells for in vivo chondrogenesis: Suitable conditions for cell therapy of cartilage defects in rabbit. Cell Tissue Res. 2008, 333, 207–215. [Google Scholar] [CrossRef]
- O’Driscoll, S.W.; Recklies, A.D.; Poole, A.R. Chondrogenesis in periosteal explants. An organ culture model for in vitro study. JBJS 1994, 76, 1042–1051. [Google Scholar] [CrossRef]
- Makris, E.A.; Huang, B.J.; Hu, J.C.; Chen-Izu, Y.; Athanasiou, K.A. Digoxin and adenosine triphosphate enhance the functional properties of tissue-engineered cartilage. Tissue Eng. Part A 2015, 21, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Caron, M.M.J.; Janssen, M.P.F.; Peeters, L.; Haudenschild, D.R.; Cremers, A.; Surtel, D.A.M.; van Rhijn, L.W.; Emans, P.J.; Welting, T.J.M. Aggrecan and COMP Improve Periosteal Chondrogenesis by Delaying Chondrocyte Hypertrophic Maturation. Front. Bioeng. Biotechnol. 2020, 8, 1036. [Google Scholar] [CrossRef] [PubMed]
- Kock, L.; van Donkelaar, C.C.; Ito, K. Tissue engineering of functional articular cartilage: The current status. Cell Tissue Res. 2012, 347, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.H.; Rousche, K.T.; Tuan, R.S. Technology Insight: Adult stem cells in cartilage regeneration and tissue engineering. Nat. Clin. Pract. Rheumatol. 2006, 2, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, R.L.; Ni, M.; Meretoja, V.V.; Kasper, F.K.; Mikos, A.G. TGF-β3-induced chondrogenesis in co-cultures of chondrocytes and mesenchymal stem cells on biodegradable scaffolds. Biomaterials 2014, 35, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, R.S.; Won, H.K.; Zeng, L. The role of Nkx3.2 in chondrogenesis. Front. Biol. 2014, 9, 376–381. [Google Scholar] [CrossRef]
- Hsiao, H.-Y.; Cheng, C.-M.; Kao, S.-W.; Liu, J.-W.; Chang, C.-S.; Harhaus, L.; Huang, J.-J. The effect of bone inhibitors on periosteum-guided cartilage regeneration. Sci. Rep. 2020, 10, 8372. [Google Scholar] [CrossRef]
- Choi, Y.S.; Lim, S.M.; Shin, H.C.; Lee, C.W.; Kim, D.-I. Chondrogenic properties of human periosteum-derived progenitor cells (PDPCs) embedded in a thermoreversible gelation polymer (TGP). Biotechnol. Bioprocess Eng. 2006, 11, 550–552. [Google Scholar] [CrossRef]
- Grässel, S.; Rickert, M.; Opolka, A.; Bosserhoff, A.; Angele, P.; Grifka, J.; Anders, S. Coculture between periosteal explants and articular chondrocytes induces expression of TGF-beta1 and collagen I. Rheumatol. Oxf. Engl. 2010, 49, 218–230. [Google Scholar] [CrossRef][Green Version]
- Li, M.; Amizuka, N.; Oda, K.; Tokunaga, K.; Ito, T.; Takeuchi, K.; Takagi, R.; Maeda, T. Histochemical evidence of the initial chondrogenesis and osteogenesis in the periosteum of a rib fractured model: Implications of osteocyte involvement in periosteal chondrogenesis. Microsc. Res. Tech. 2004, 64, 330–342. [Google Scholar] [CrossRef]
- Fukumoto, T.; Sanyal, A.; Fitzsimmons, J.S.; O’Driscoll, S.W. Expression of beta1 integrins during periosteal chondrogenesis. Osteoarthr. Cartil. 2002, 10, 135–144. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Casper, M.E.; Fitzsimmons, J.S.; Stone, J.J.; Meza, A.O.; Huang, Y.; Ruesink, T.J.; O’Driscoll, S.W.; Reinholz, G.G. Tissue engineering of cartilage using poly-ε-caprolactone nanofiber scaffolds seeded in vivo with periosteal cells. Osteoarthr. Cartil. 2010, 18, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Duchamp de Lageneste, O.; Julien, A.; Abou-Khalil, R.; Frangi, G.; Carvalho, C.; Cagnard, N.; Cordier, C.; Conway, S.J.; Colnot, C. Periosteum contains skeletal stem cells with high bone regenerative potential controlled by Periostin. Nat. Commun. 2018, 9, 773. [Google Scholar] [CrossRef] [PubMed]
- Debnath, S.; Yallowitz, A.R.; McCormick, J.; Lalani, S.; Zhang, T.; Xu, R.; Li, N.; Liu, Y.; Yang, Y.S.; Eiseman, M.; et al. Discovery of a periosteal stem cell mediating intramembranous bone formation. Nature 2018, 562, 133–139. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeyaraman, M.; Muthu, S.; Gangadaran, P.; Ranjan, R.; Jeyaraman, N.; Prajwal, G.S.; Mishra, P.C.; Rajendran, R.L.; Ahn, B.-C. Osteogenic and Chondrogenic Potential of Periosteum-Derived Mesenchymal Stromal Cells: Do They Hold the Key to the Future? Pharmaceuticals 2021, 14, 1133. https://doi.org/10.3390/ph14111133
Jeyaraman M, Muthu S, Gangadaran P, Ranjan R, Jeyaraman N, Prajwal GS, Mishra PC, Rajendran RL, Ahn B-C. Osteogenic and Chondrogenic Potential of Periosteum-Derived Mesenchymal Stromal Cells: Do They Hold the Key to the Future? Pharmaceuticals. 2021; 14(11):1133. https://doi.org/10.3390/ph14111133
Chicago/Turabian StyleJeyaraman, Madhan, Sathish Muthu, Prakash Gangadaran, Rajni Ranjan, Naveen Jeyaraman, Gollahalli Shivashankar Prajwal, Prabhu Chandra Mishra, Ramya Lakshmi Rajendran, and Byeong-Cheol Ahn. 2021. "Osteogenic and Chondrogenic Potential of Periosteum-Derived Mesenchymal Stromal Cells: Do They Hold the Key to the Future?" Pharmaceuticals 14, no. 11: 1133. https://doi.org/10.3390/ph14111133
APA StyleJeyaraman, M., Muthu, S., Gangadaran, P., Ranjan, R., Jeyaraman, N., Prajwal, G. S., Mishra, P. C., Rajendran, R. L., & Ahn, B.-C. (2021). Osteogenic and Chondrogenic Potential of Periosteum-Derived Mesenchymal Stromal Cells: Do They Hold the Key to the Future? Pharmaceuticals, 14(11), 1133. https://doi.org/10.3390/ph14111133