
Chondroitin Sulfate and Fucosylated Chondroitin Sulfate as Stimulators of Hematopoiesis in Cyclophosphamide-Induced Mice


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
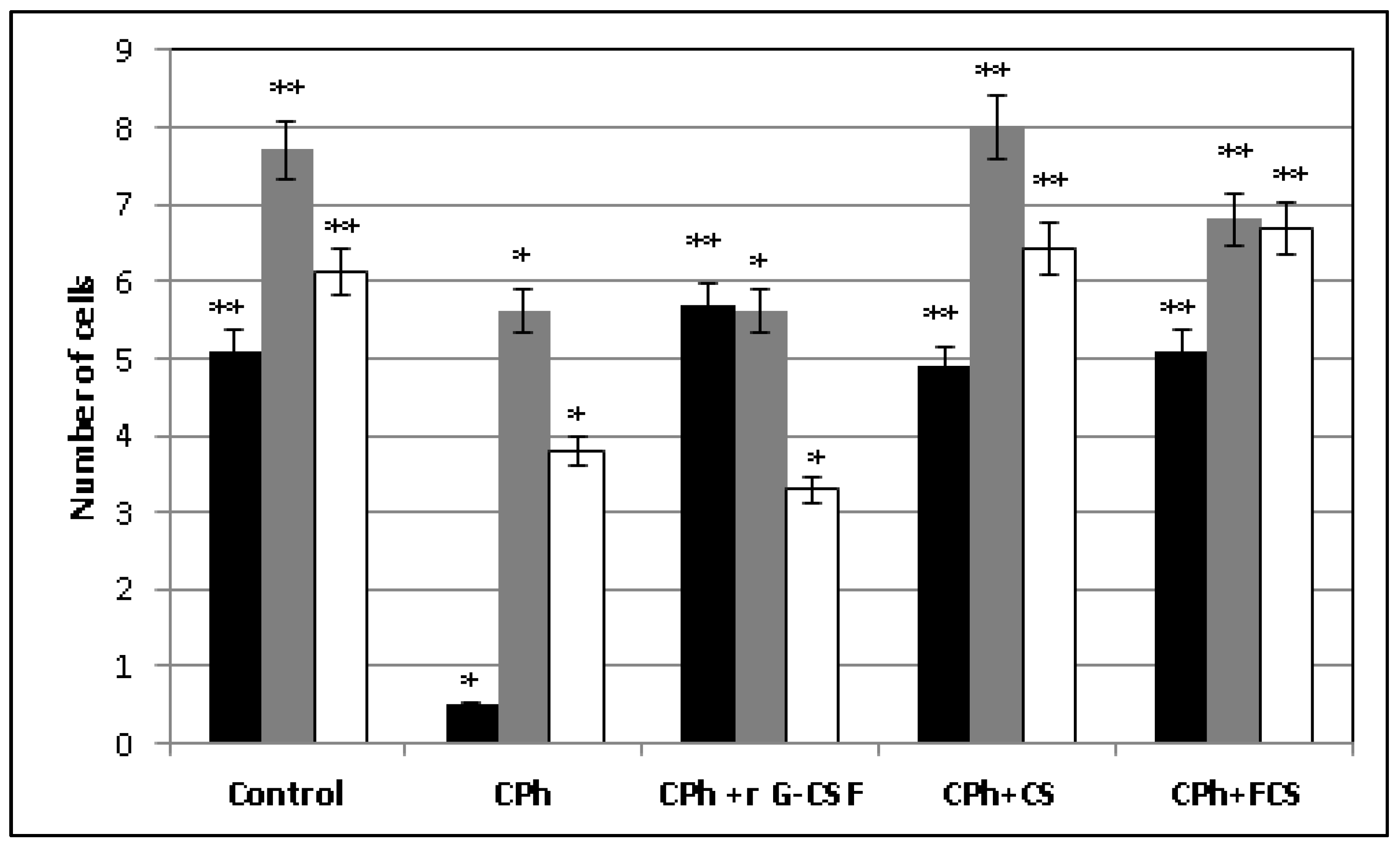
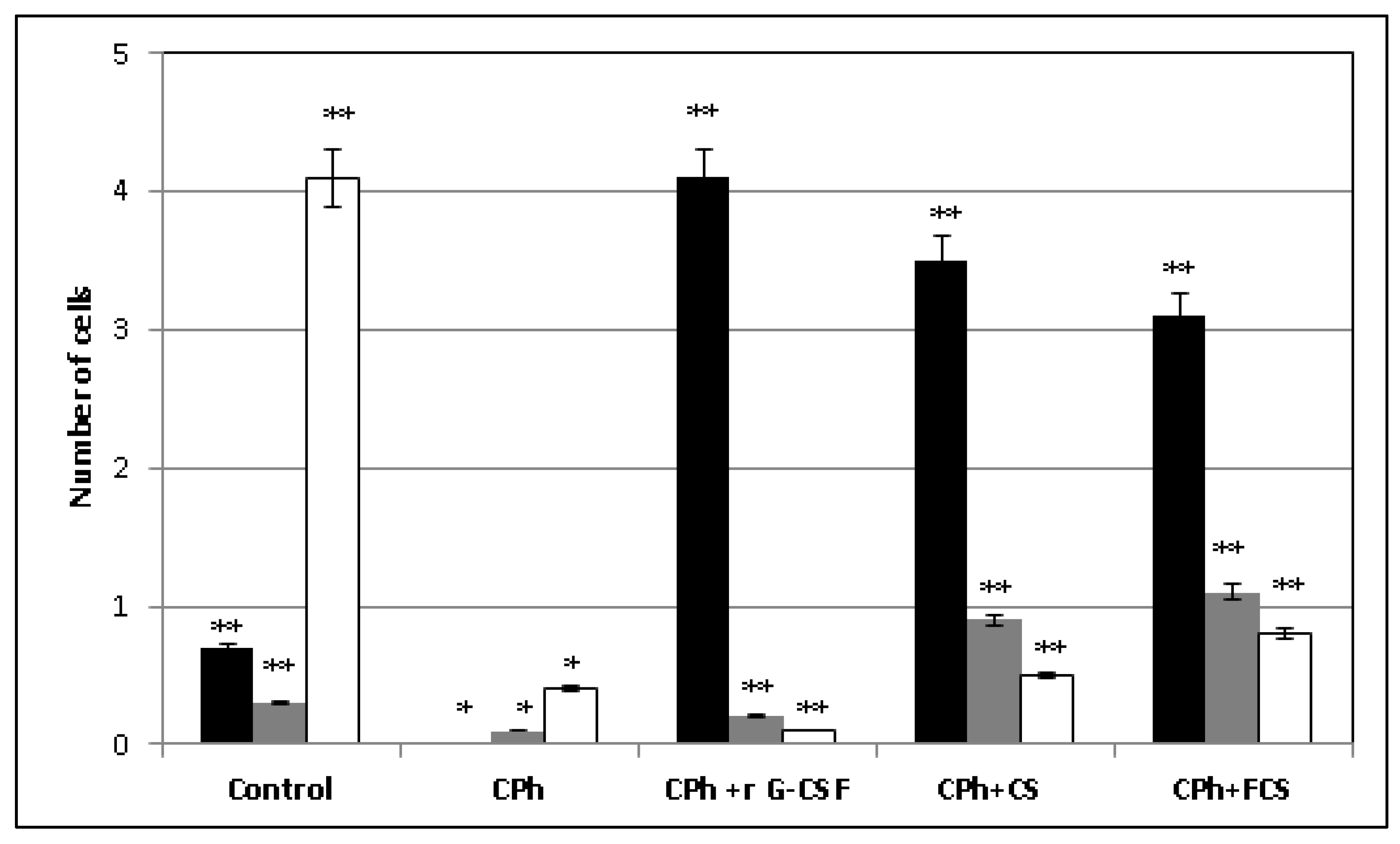
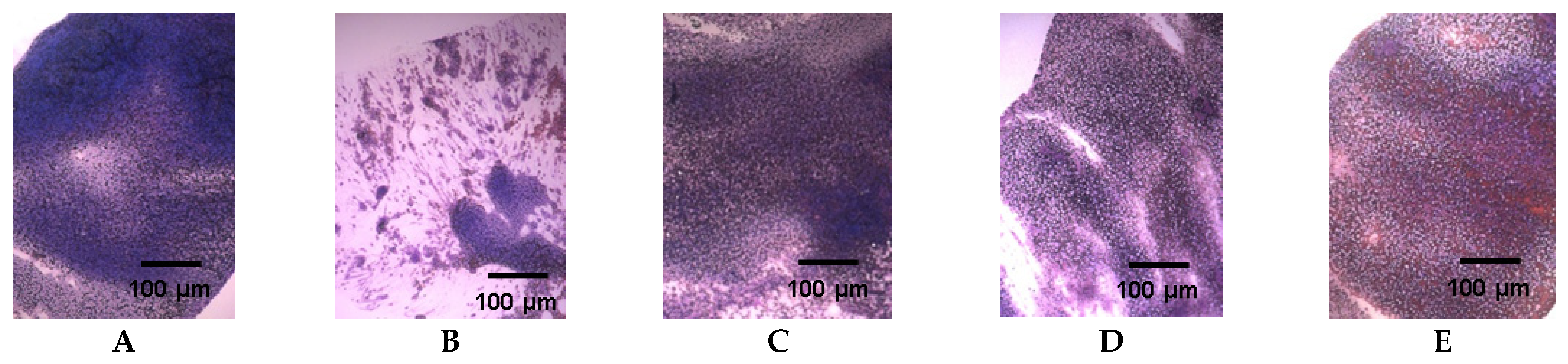
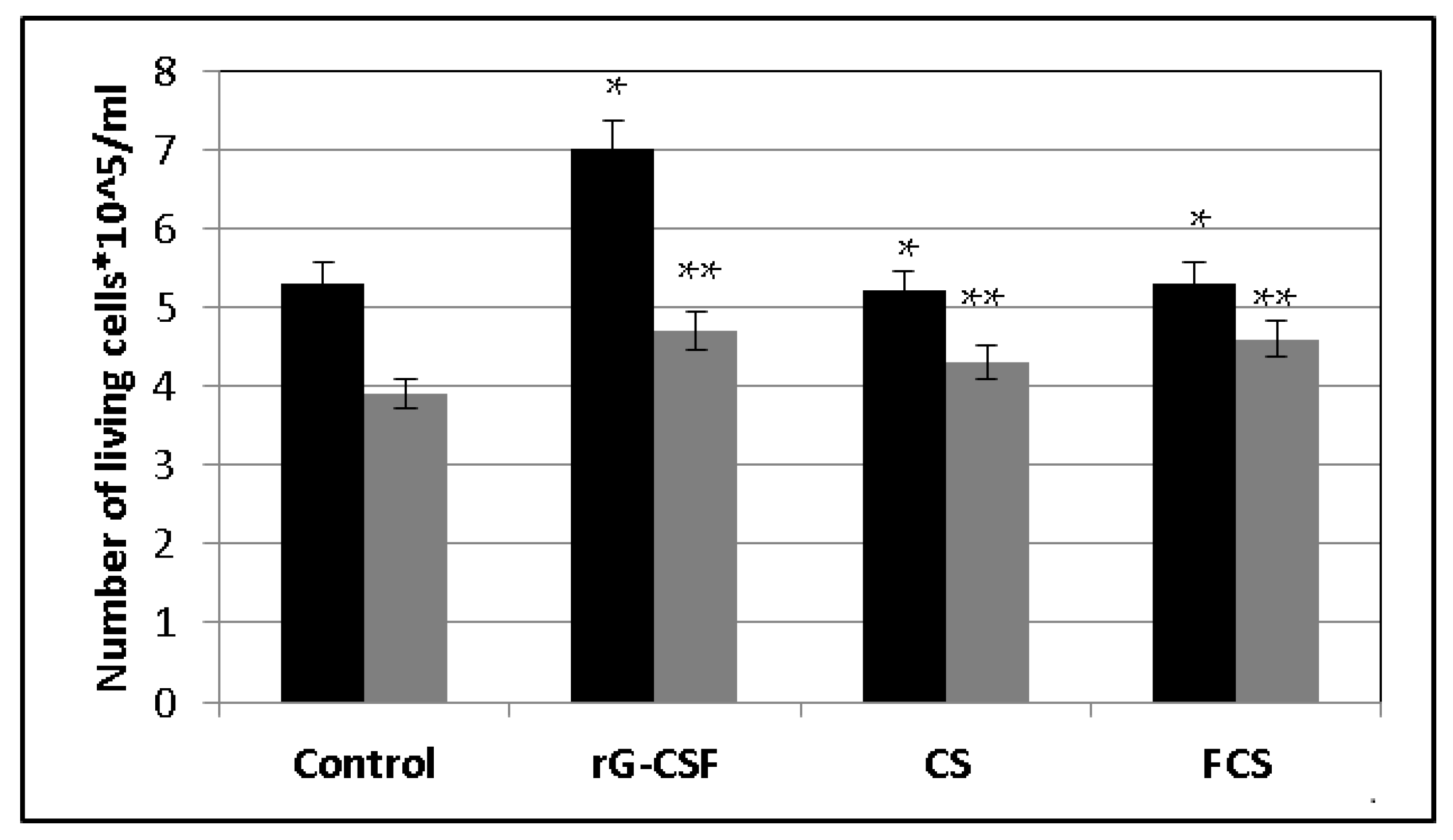
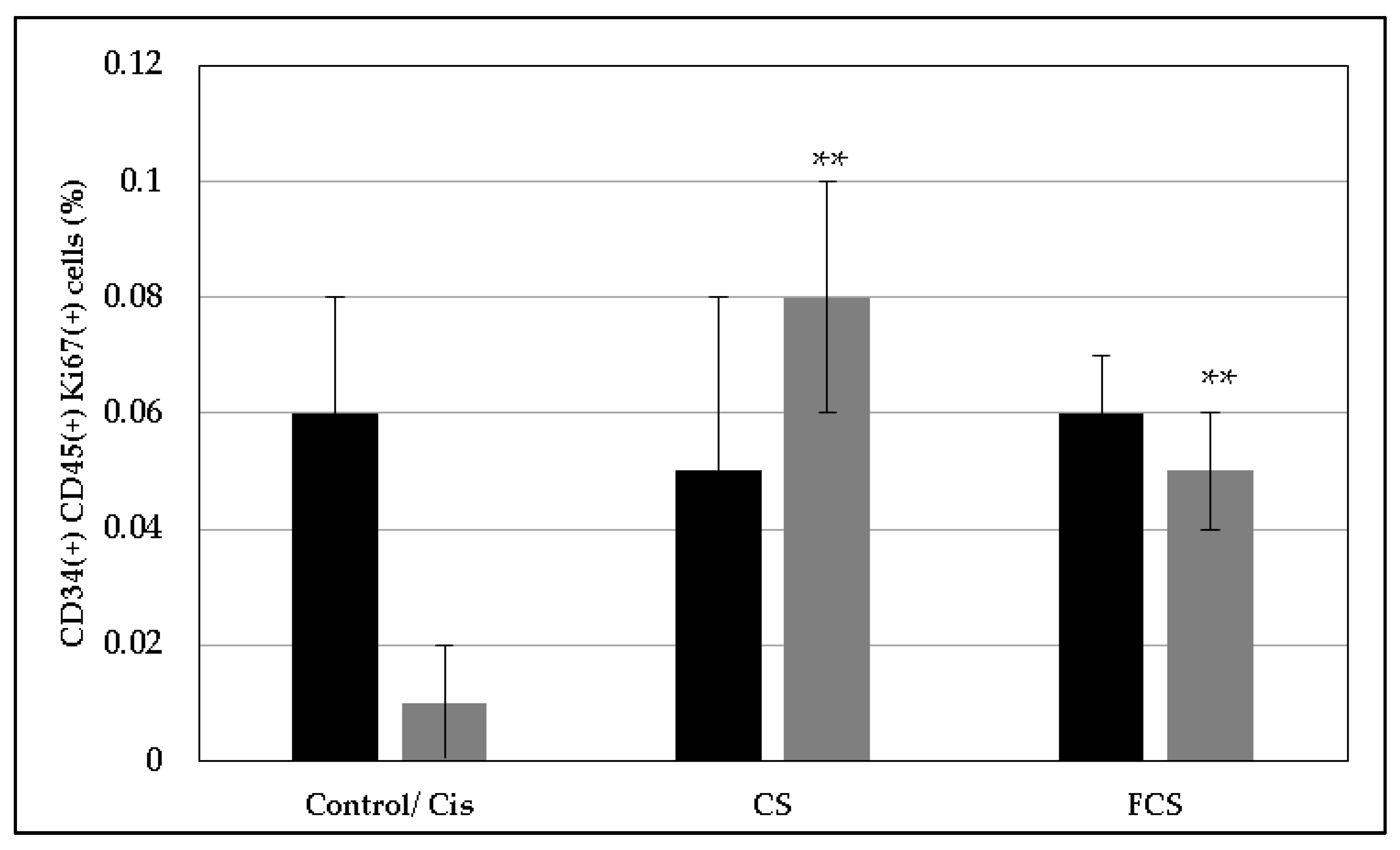
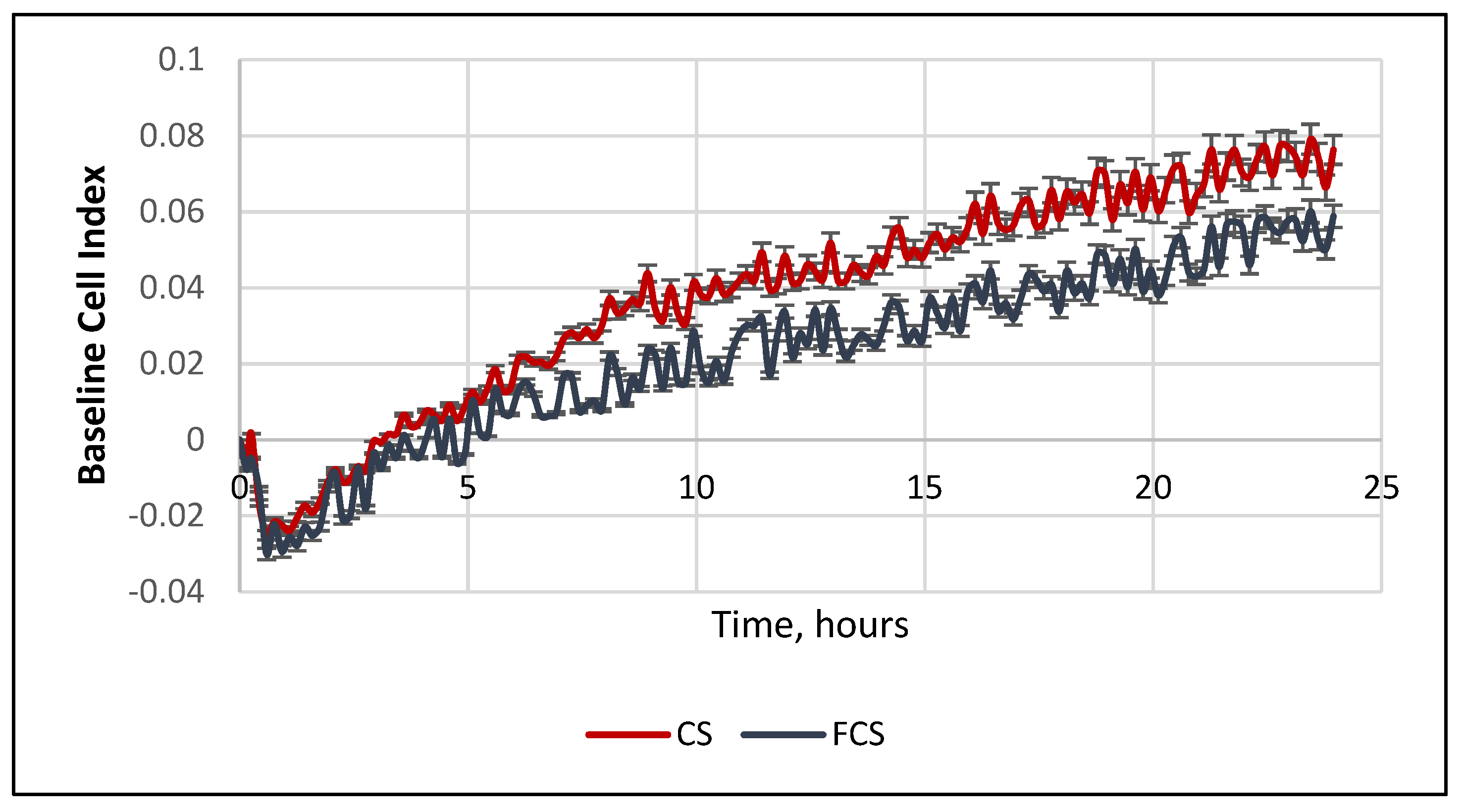
2. Results
3. Discussion
4. Materials and Methods
4.1. General Methods
4.2. Sulfated Polysaccharides
4.3. Animal Model
4.4. Cell Model
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Karagiannis, G.S.; Condeelis, J.S.; Oktay, M.H. Chemotherapy-induced metastasis: Molecular mechanisms, clinical manifestations, therapeutic interventions. Cancer Res. 2019, 79, 4567–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.; Kumar, S.; Tchounwou, P. Cisplatin-Based Chemotherapy of Human Cancers. J. Cancer Sci. Ther. 2019, 11, 97. [Google Scholar]
- Dasari, S.; Tchounwou, P. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, J.H.; Bremer, J.P.; Moosig, F.; Holle, J.U.; Lamprecht, P.; Wieczorek, S.; Haenisch, S.; Cascorbi, I. Cyclophosphamide treatment-induced leukopenia rates in ANCA-associated vasculitis are influenced by variant CYP450 2C9 genotypes. Pharmacogenomics 2016, 17, 367–374. [Google Scholar] [CrossRef]
- Kuter, D.J. Managing thrombocytopenia associated with cancer chemotherapy. Oncology 2015, 29, 282–294. [Google Scholar]
- Yu, J.J.; Hogan, T.; Morley, C.; Crigger, C.; Jiao, S.; Williams, D.J.; Salkini, M.W.; Yang, X.; Liang, X.; Yan, B.; et al. Adverse effects profile of dicycloplatin (DCP) offers chemotherapeutic advantage over cisplatin and carboplatin. Anticancer Res. 2019, 39, 4455–4462. [Google Scholar] [CrossRef]
- Mundry, C.S.; Eberle, K.C.; Singh, P.K.; Hollingsworth, M.A.; Mehla, K. Local and systemic immunosuppression in pancreatic cancer: Targeting the stalwarts in tumor’s arsenal. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188387. [Google Scholar] [CrossRef]
- Floyd, F.; Mirza, I.; Sachs, B.; Perry, M. Hepatotoxicity of chemotherapy. Semin. Oncol. 2006, 33, 50–67. [Google Scholar] [CrossRef] [Green Version]
- Anqi, Y.; Yu, Z.; Mingjun, X.; Xiaoli, K.; Mengmeng, L.; Fangfang, L.; Mei, Z. Use of echocardiography to monitor myocardial damage during anthracycline chemotherapy. Echocardiography 2019, 36, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, D.; Sandri, M.T.; Colombo, A.; Colombo, N.; Boeri, M.; Lamantia, G.; Civelli, M.; Peccatori, F.; Martinelli, G.; Fiorentini, C.; et al. Prognostic value of troponin I in cardiac risk stratification of cancer patients undergoing high-dose chemotherapy. Circulation 2004, 109, 2749–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangi, A.; Lu, S.C. Chemotherapy-associated liver injury in colorectal cancer. Therap. Adv. Gastroenterol. 2020, 13, 1756284820924194. [Google Scholar] [CrossRef]
- Urrutia-Maldonado, E.; Abril-Molina, A.; Alés-Palmer, M.; Gómez-Luque, J.M.; Muñoz de Rueda, P.; Ocete-Hita, E. Chemotherapy-induced liver injury in children. Pediatr 2019, 91, 256–263. [Google Scholar] [CrossRef]
- Passweg, J.R.; Baldomero, H.; Bader, P.; Basak, G.W.; Bonin, C.; Duarte, R.; Dufour, C.; Kröger, N.; Kuball, J.; Lankester, A.; et al. Is the use of unrelated donor transplantation leveling off in Europe? The 2016 European Society for Blood and Marrow Transplant activity survey report. Bone Marrow Transplant. 2018, 53, 1139–1148. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Wang, H.; Zhang, L.; Wei, W. Advances in the treatment of graft-versus-host disease with immunomodulatory cells. Int. Immunopharmacol. 2021, 92, 107349. [Google Scholar] [CrossRef]
- Martin, P.J.; Rizzo, J.D.; Wingard, J.R.; Ballen, K.; Curtin, P.T.; Cutler, C.; Litzow, M.R.; Nieto, Y.; Savani, B.N.; Schriber, J.R.; et al. First- and second-line systemic treatment of acute graft-versus-host disease: Recommendations of the American Society of Blood and Marrow Transplantation. Biol. Blood Marrow Transplant. 2012, 18, 1150–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussetti, A.; Greco, R.; Peccatori, J.; Corradini, P. Post-transplant cyclophosphamide, a promising anti-graft versus host disease prophylaxis: Where do we stand? Expert Rev. Hematol. 2017, 10, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Crobu, D.; Spinetti, G.; Schrepfer, R.; Tonon, G.; Jotti, G.S.; Onali, P.; Dedoni, S.; Orsini, G.; Di Stefano, A. Preclinical and clinical phase I studies of a new recombinant Filgrastim (BK0023) in comparison with Neupogen®. BMC Pharmacol. Toxicol. 2014, 15, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, N.; Ustyuzhanina, N.; Donenko, F.; Bilan, M.; Usov, A.; Nifantiev, N.; Kiselevskiy, M. Fucoidan and fucosylated chondroitin sulfate stimulate hematopoiesis in cyclophosphamide-induced mice. Mar. Drugs 2017, 15, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilan, M.I.; Vinogradova, E.V.; Tsvetkova, E.A.; Grachev, A.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A sulfated glucuronofucan containing both fucofuranose and fucopyranose residues from the brown alga Chordaria flagelliformis. Carbohydr. Res. 2008, 343, 2605–2612. [Google Scholar] [CrossRef] [PubMed]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Borodina, E.Y.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. A highly regular fucosylated chondroitin sulfate from the sea cucumber Massinium magnum: Structure and effects on coagulation. Carbohydr. Polym. 2017, 167, 20–26. [Google Scholar] [CrossRef]
- Anisimova, N.Y.; Ustyuzhanina, N.E.; Bilan, M.I.; Donenko, F.V.; Ushakova, N.A.; Usov, A.I.; Kiselevskiy, M.V.; Nifantiev, N.E. Influence of modified fucoidan and related sulfated oligosaccharides on hematopoiesis in cyclophosphamide-induced mice. Mar. Drugs 2018, 16, 333. [Google Scholar] [CrossRef] [Green Version]
- Ustyuzhanina, N.E.; Bilan, M.I.; Panina, E.G.; Sanamyan, N.P.; Dmitrenok, A.S.; Tsvetkova, E.A.; Ushakova, N.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure and anti-inflammatory activity of a new unusual fucosylated chondroitin sulfate from Cucumaria djakonovi. Mar. Drugs 2018, 16, 389. [Google Scholar] [CrossRef] [Green Version]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Tsvetkova, E.A.; Shashkov, A.S.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. Structural characterization of fucosylated chondroitin sulfates from sea cucumbers Apostichopus japonicus and Actinopyga mauritiana. Carbohydr. Polym. 2016, 153, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Shakiba, E.; Ramezani, M.; Sadeghi, M. Evaluation of serum interleukin-6 levels in hepatocellular carcinoma patients: A systematic review and meta-analysis. Clin. Exp. Hepatol. 2018, 4, 182–190. [Google Scholar] [CrossRef]
- Rose-John, S. IL-6 trans-signaling via the soluble IL-6 receptor: Importance for the pro-inflammatory activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Elsea, C.R.; Kneiss, J.A.; Wood, L.J. Induction of IL-6 by cytotoxic chemotherapy is associated with loss of lean body and fat mass in tumor-free female mice. Biol. Res. Nurs. 2015, 17, 549–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ba, Y.; Shi, Y.; Jiang, W.; Feng, J.; Cheng, Y.; Xiao, L.; Zhang, Q.; Qiu, W.; Xu, B.; Xu, R.; et al. Current management of chemotherapy-induced neutropenia in adults: Key points and new challenges: Committee of Neoplastic Supportive-Care (CONS). Cancer Biol. Med. 2020, 17, 896–909. [Google Scholar] [CrossRef]
- Abdel-Razeq, H.; Hashem, H. Recent update in the pathogenesis and treatment of chemotherapy and cancer induced anemia. Crit. Rev. Oncol. Hematol. 2020, 145, 102837. [Google Scholar] [CrossRef] [PubMed]
- Karagiannidis, I.; Salataj, E.; Said Abu Egal, E.; Beswick, E.J. G-CSF in tumors: Aggressiveness, tumor microenvironment and immune cell regulation. Cytokine 2021, 142, 155479. [Google Scholar] [CrossRef]
- Singbart, G. Adverse events of erythropoietin in long-term and in acute/short-term treatment. Clin. Investig. 1994, 72, S36–S43. [Google Scholar]
- Trivedi, M.; Martinez, S.; Corring-Ham, S.; Medley, K.; Ball, E.D. Optimal use of G-CSF administration after HSCT. Bone Marrow Transplant. 2009, 43, 895–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekker, A.; Bulley, S.; Beyene, J.; Dupuis, L.; Doyle, J.J.; Sung, L. Meta-analysis of randomized controlled trials of prophylactic granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor after autologous and allogeneic stem cell transplantation. J. Clin. Oncol. 2006, 24, 5207–5215. [Google Scholar] [CrossRef]
- Nassar, A.; Elgo-hary, G.; Elhassan, T.; Nurgat, Z.; Mohamed, S.Y.; Aljurf, M.J. Methotrexate for the Treatment of Graft-versus-Host Disease after Allogeneic Hematopoietic Stem Cell Transplantation. J. Transplant. 2014, 980301. [Google Scholar] [CrossRef]
- Wang, G.; Peng, X. A Review of Clinical Applica-tions and Side Effects of Methotrexate in Ophthalmology. J. Ophthalmol. 2020, 2020, 1537689. [Google Scholar] [CrossRef]
- Li, C.; Niu., Q.; Li, S.; Zhang, X.; Liu, C.; Cai, C.; Li, G.; Yu, G. Fucoidan from sea cucumber Holothuria polii: Structural elucidation and stimulation of hematopoietic activity. Int. J. Biol. Macromol. 2020, 154, 1123–1131. [Google Scholar] [CrossRef]
- Kiselevskiy, M.; Shubina, I.; Chikileva, I.; Sitdikova, S.; Samoylenko, I.; Anisimova, N.; Kirgizov, K.; Suleimanova, A.; Gorbunova, T.; Varfolomeev, S. Immune Pathogenesis of COVID-19 Intoxication: Storm or Silence? Pharmaceuticals 2020, 13, 166. [Google Scholar] [CrossRef] [PubMed]
- Tvedt, T.H.; Ersvaer, E.; Tveita, A.A.; Bruserud, O. Interleukin-6 in allogeneic stem cell transplantation: Its possible importance for immunoregulation and as a therapeutic target. Front. Immunol. 2017, 8, 667. [Google Scholar] [CrossRef]
- Tvedt, T.H.; Lie, S.A.; Reikvam, H.; Rye, K.P.; Lindas, R.; Gedde-Dahl, T.; Ahmed, A.B.; Bruserud, O. Pretransplant levels of CRP and interleukin-6 family cytokines; effects on outcome after allogeneic stem cell transplantation. Int. J. Mol. Sci. 2016, 17, 1823. [Google Scholar] [CrossRef] [Green Version]
- Drobyski, W.R.; Pasquini, M.; Kovatovic, K.; Palmer, J.; Rizzo, J.D.; Saad, A.; Saber, W.; Hari, P. Tocilizumab for the treatment of steroid refractory graft-versus-host disease. Biol. Blood Marrow Transplant. 2011, 17, 1862–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesley, J.; Hyman, R.; Kincade, P.W. CD44 and its interaction with extracellular matrix. Adv. Immunol. 1993, 54, 271–335. [Google Scholar]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A multifunctional cell surface adhesion receptor is a regulator of progression and metastasis of cancer cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, G.F.; Ashkar, S.; Glimcher, M.J.; Cantor, H. Receptor-ligand interaction between CD44 and osteopontin (Eta-1). Science 1996, 271, 509–512. [Google Scholar] [CrossRef]
- Kim, Y.; Kumar, S. CD44-mediated adhesion to hyaluronic acid contributes to mechanosensing and invasive motility. Mol. Cancer Res. 2014, 10, 1416–1429. [Google Scholar] [CrossRef] [Green Version]
- Govindaraju, P.; Todd, L.; Shetye, S.; Monslow, J.; Puré, E. CD44-dependent inflammation, fibrogenesis, and collagenolysis regulates extracellular matrix remodeling and tensile strength during cutaneous wound healing. Matrix Biol. 2019, 75–76, 314–330. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.A.; et al. Comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Pomin, V.H. Holothurian fucosylated chondroitin sulfates. Mar. Drugs 2014, 12, 232–254. [Google Scholar] [CrossRef] [Green Version]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S.; Kusaykin, M.I.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. Structure and biological activity of a fucosylated chondroitin sulfate from the sea cucumber. Cucumaria Japonica Glycobiol. 2016, 26, 449–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ustyuzhanina, N.E.; Ushakova, N.A.; Zyuzina, K.A.; Bilan, M.I.; Elizarova, A.L.; Somonova, O.V.; Madzhuga, A.V.; Krylov, V.B.; Preobrazhenskaya, M.E.; Usov, A.I.; et al. Influence of fucoidans on hemostatic system. Mar. Drugs 2013, 11, 2444–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpi, N. Anti-inflammatory activity of chondroitin sulphate: New functions from an old natural macromolecule. Inflammopharmacology 2011, 19, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Ustyuzhanina, N.E.; Bilan, M.I.; Nifantiev, N.E.; Usov, A.I. New insight on the structural diversity of holothurian fucosylated chondroitin sulfates. Pure Appl. Chem. 2019, 91, 1065–1071. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. The structure of a fucosylated chondroitin sulfate from the sea cucumber. Cucumaria Frondosa. Carbohydr. Polym. 2017, 165, 7–12. [Google Scholar] [CrossRef] [PubMed]
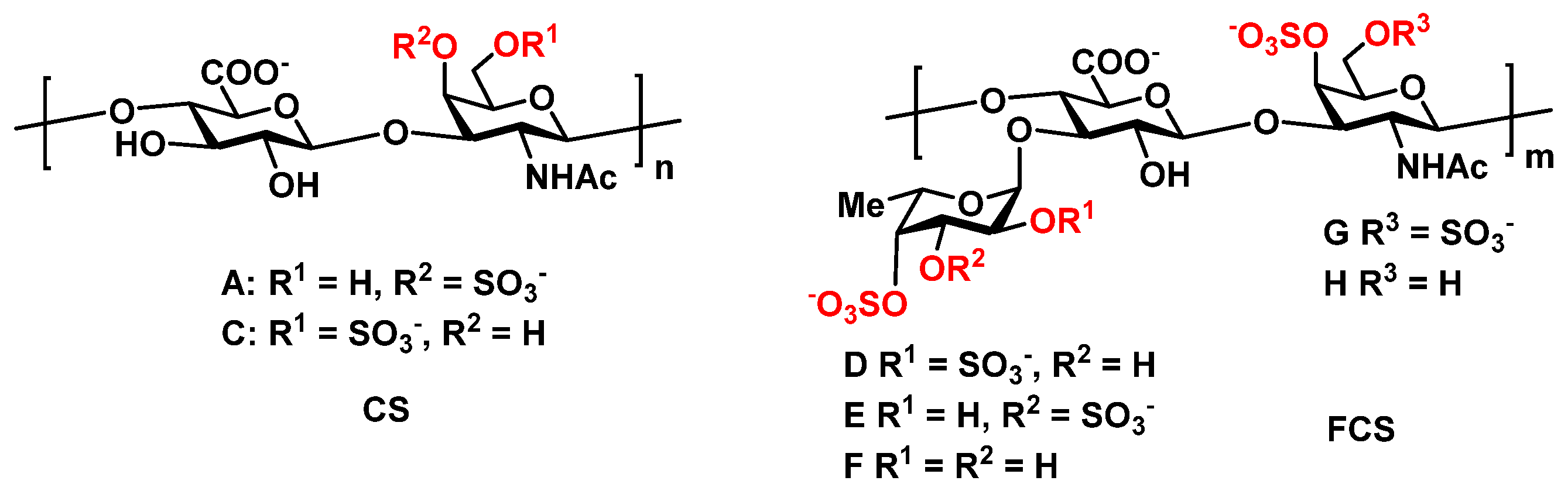



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ustyuzhanina, N.E.; Anisimova, N.Y.; Bilan, M.I.; Donenko, F.V.; Morozevich, G.E.; Yashunskiy, D.V.; Usov, A.I.; Siminyan, N.G.; Kirgisov, K.I.; Varfolomeeva, S.R.; et al. Chondroitin Sulfate and Fucosylated Chondroitin Sulfate as Stimulators of Hematopoiesis in Cyclophosphamide-Induced Mice. Pharmaceuticals 2021, 14, 1074. https://doi.org/10.3390/ph14111074
Ustyuzhanina NE, Anisimova NY, Bilan MI, Donenko FV, Morozevich GE, Yashunskiy DV, Usov AI, Siminyan NG, Kirgisov KI, Varfolomeeva SR, et al. Chondroitin Sulfate and Fucosylated Chondroitin Sulfate as Stimulators of Hematopoiesis in Cyclophosphamide-Induced Mice. Pharmaceuticals. 2021; 14(11):1074. https://doi.org/10.3390/ph14111074
Chicago/Turabian StyleUstyuzhanina, Nadezhda E., Natalia Yu. Anisimova, Maria I. Bilan, Fedor V. Donenko, Galina E. Morozevich, Dmitriy V. Yashunskiy, Anatolii I. Usov, Nara G. Siminyan, Kirill I. Kirgisov, Svetlana R. Varfolomeeva, and et al. 2021. "Chondroitin Sulfate and Fucosylated Chondroitin Sulfate as Stimulators of Hematopoiesis in Cyclophosphamide-Induced Mice" Pharmaceuticals 14, no. 11: 1074. https://doi.org/10.3390/ph14111074
APA StyleUstyuzhanina, N. E., Anisimova, N. Y., Bilan, M. I., Donenko, F. V., Morozevich, G. E., Yashunskiy, D. V., Usov, A. I., Siminyan, N. G., Kirgisov, K. I., Varfolomeeva, S. R., Kiselevskiy, M. V., & Nifantiev, N. E. (2021). Chondroitin Sulfate and Fucosylated Chondroitin Sulfate as Stimulators of Hematopoiesis in Cyclophosphamide-Induced Mice. Pharmaceuticals, 14(11), 1074. https://doi.org/10.3390/ph14111074