
Peptide Nanoparticle-Mediated Combinatorial Delivery of Cancer-Related siRNAs for Synergistic Anti-Proliferative Activity in Triple Negative Breast Cancer Cells



Abstract
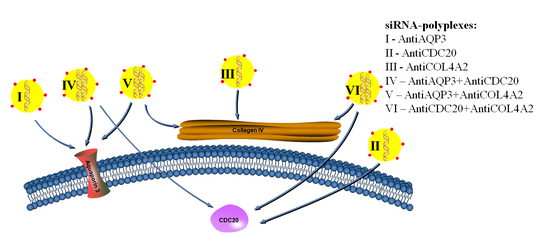
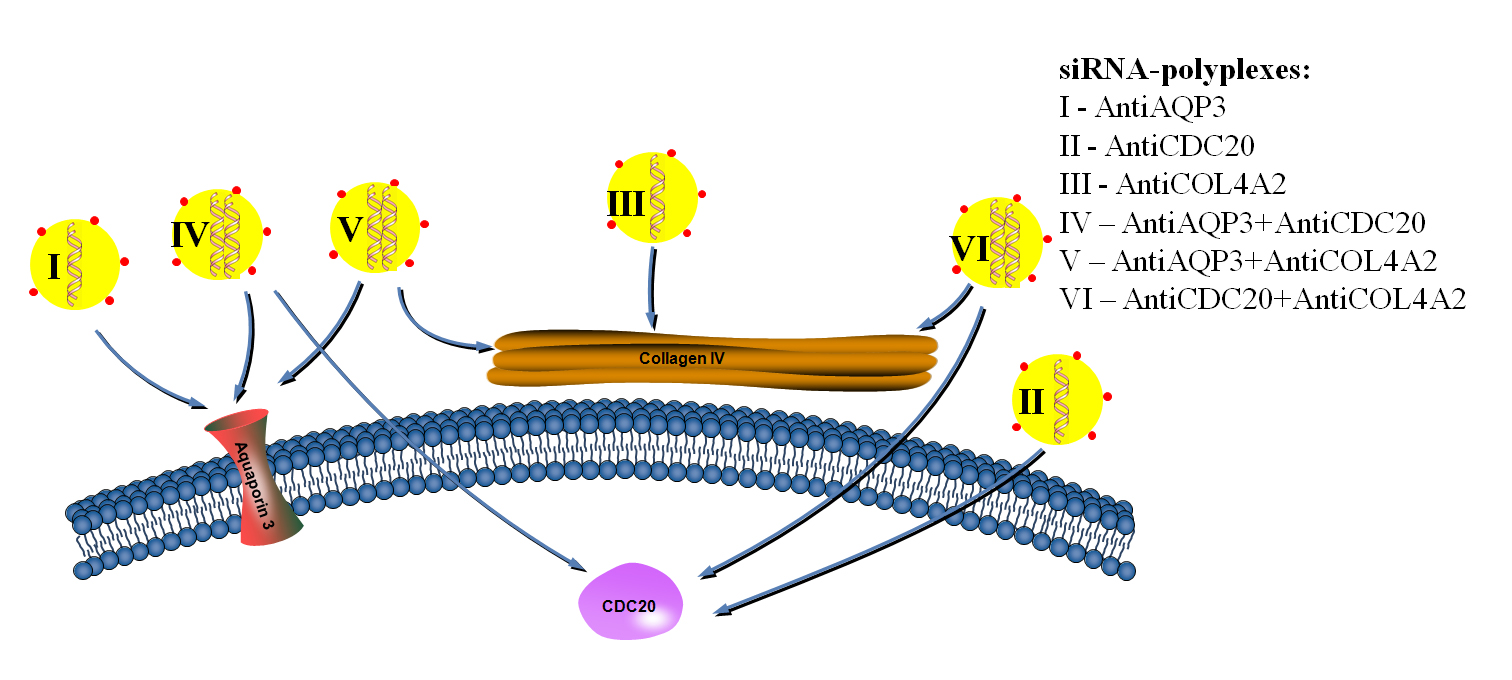
1. Introduction
2. Results and Discussion
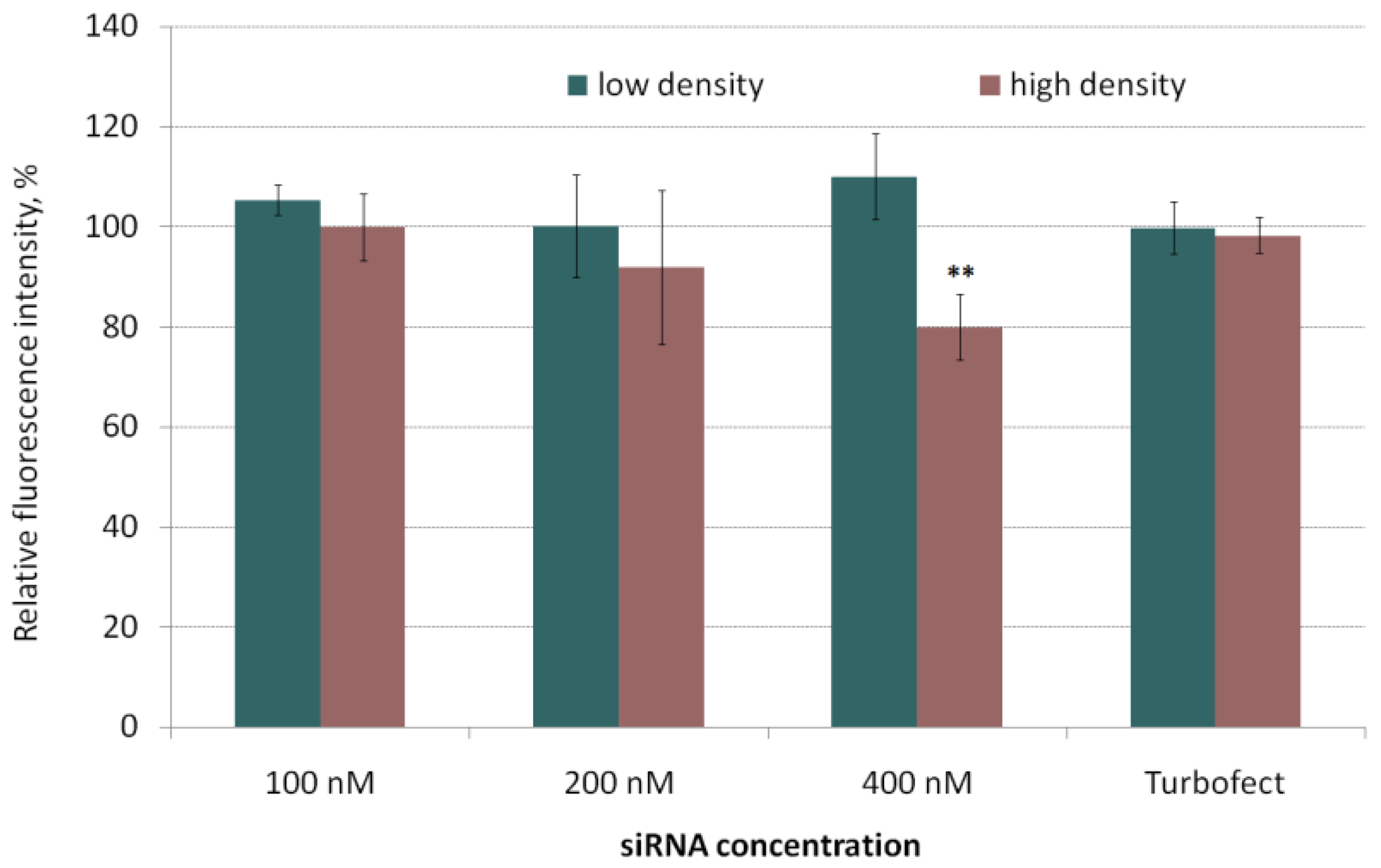
2.1. In Vitro Transfection of MDA-MB-231 Cells
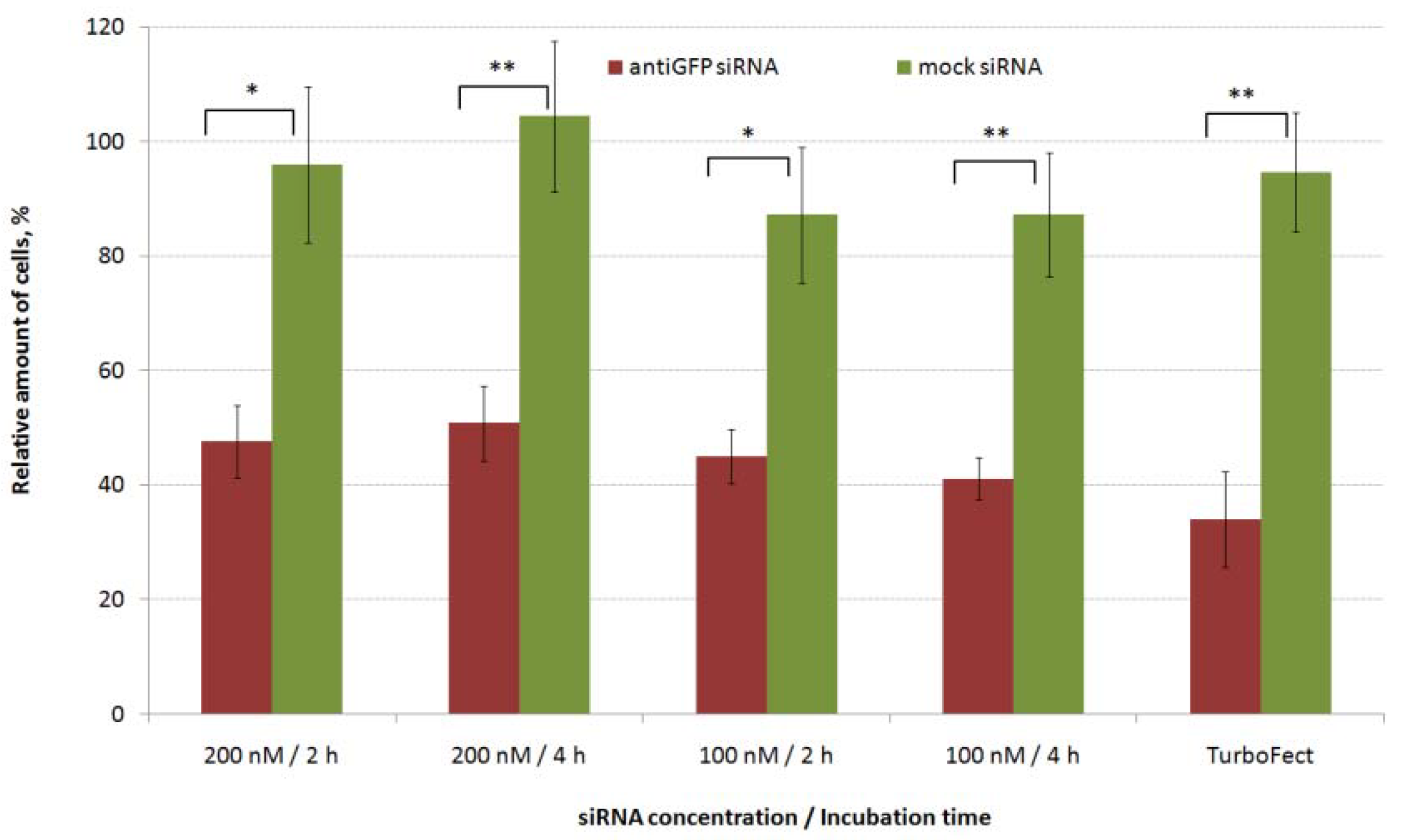
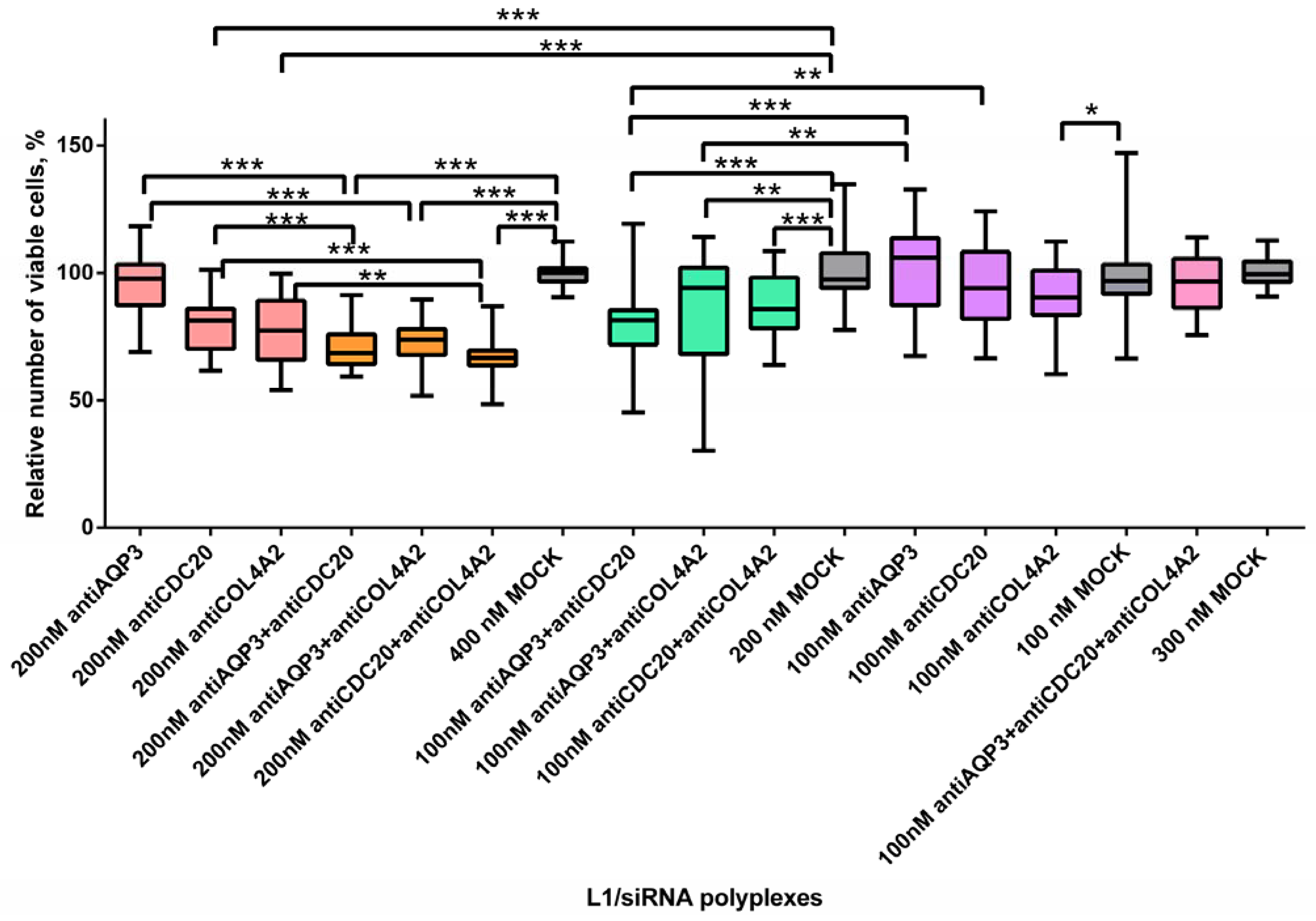
2.2. Evaluation of siRNA-Complexes Cytotoxicity
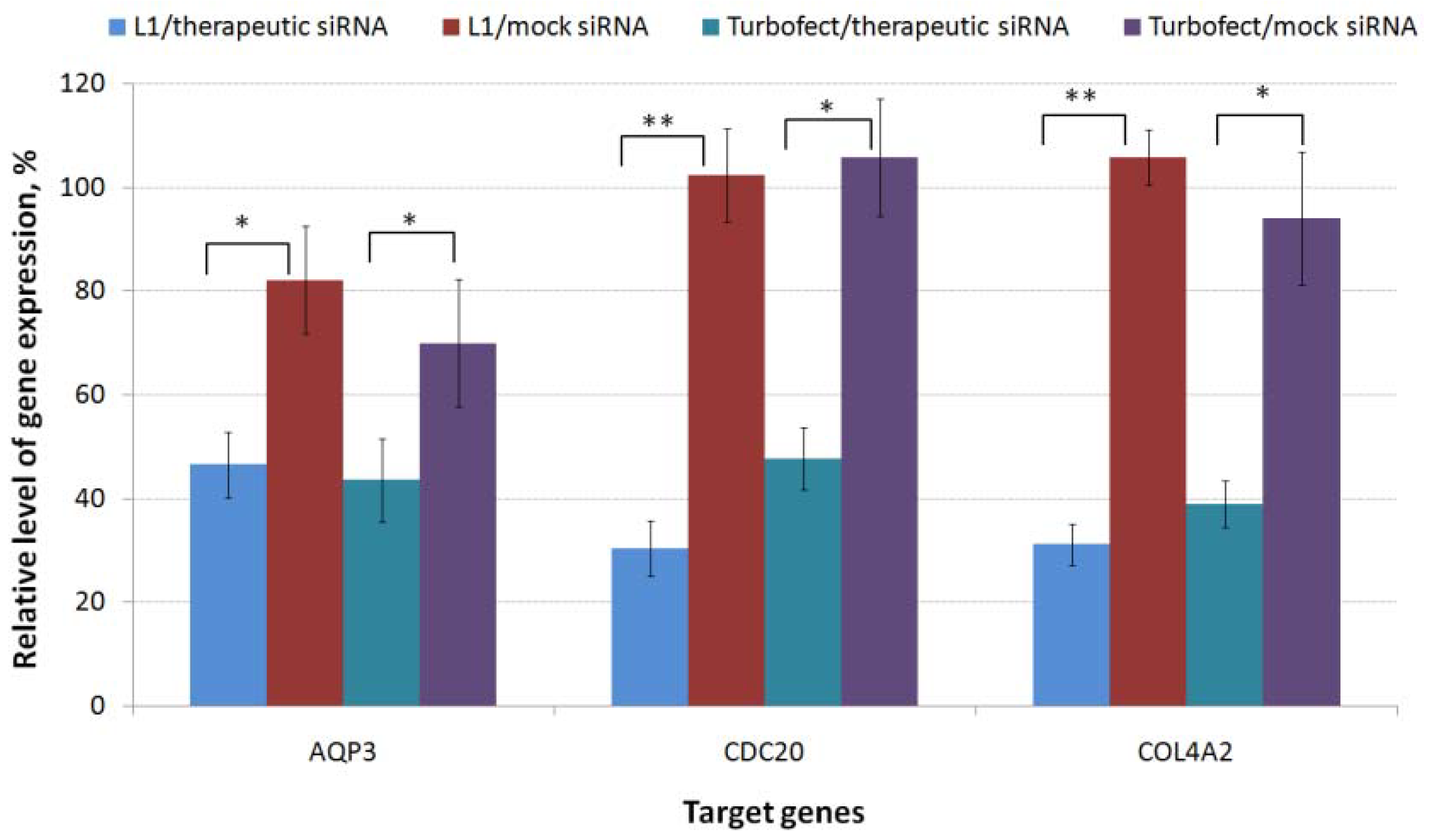
2.3. RNAi-Mediated Knockdown of TNBC-Related Genes
2.4. Inhibition of MDA-MB-231 Cells Proliferation
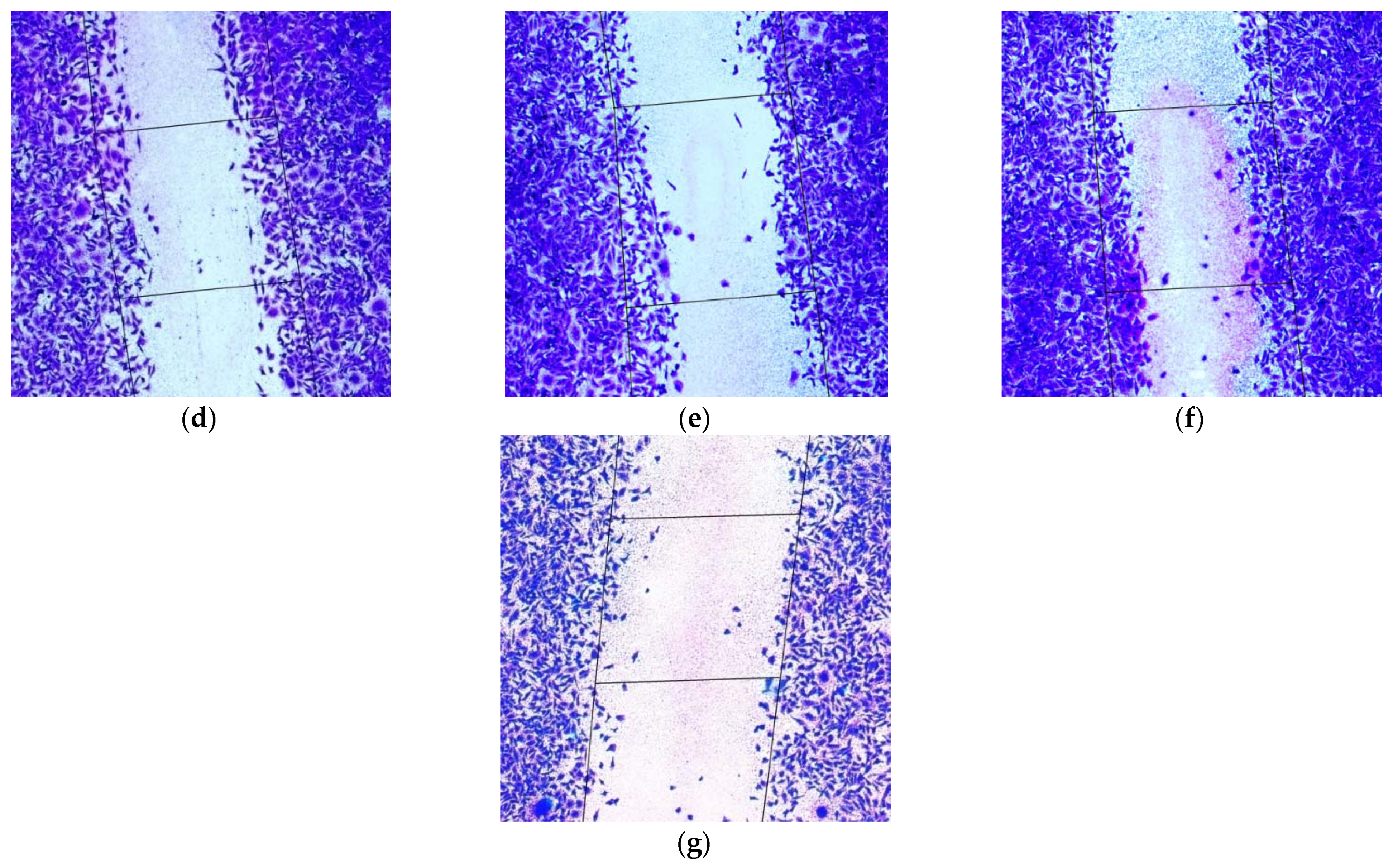
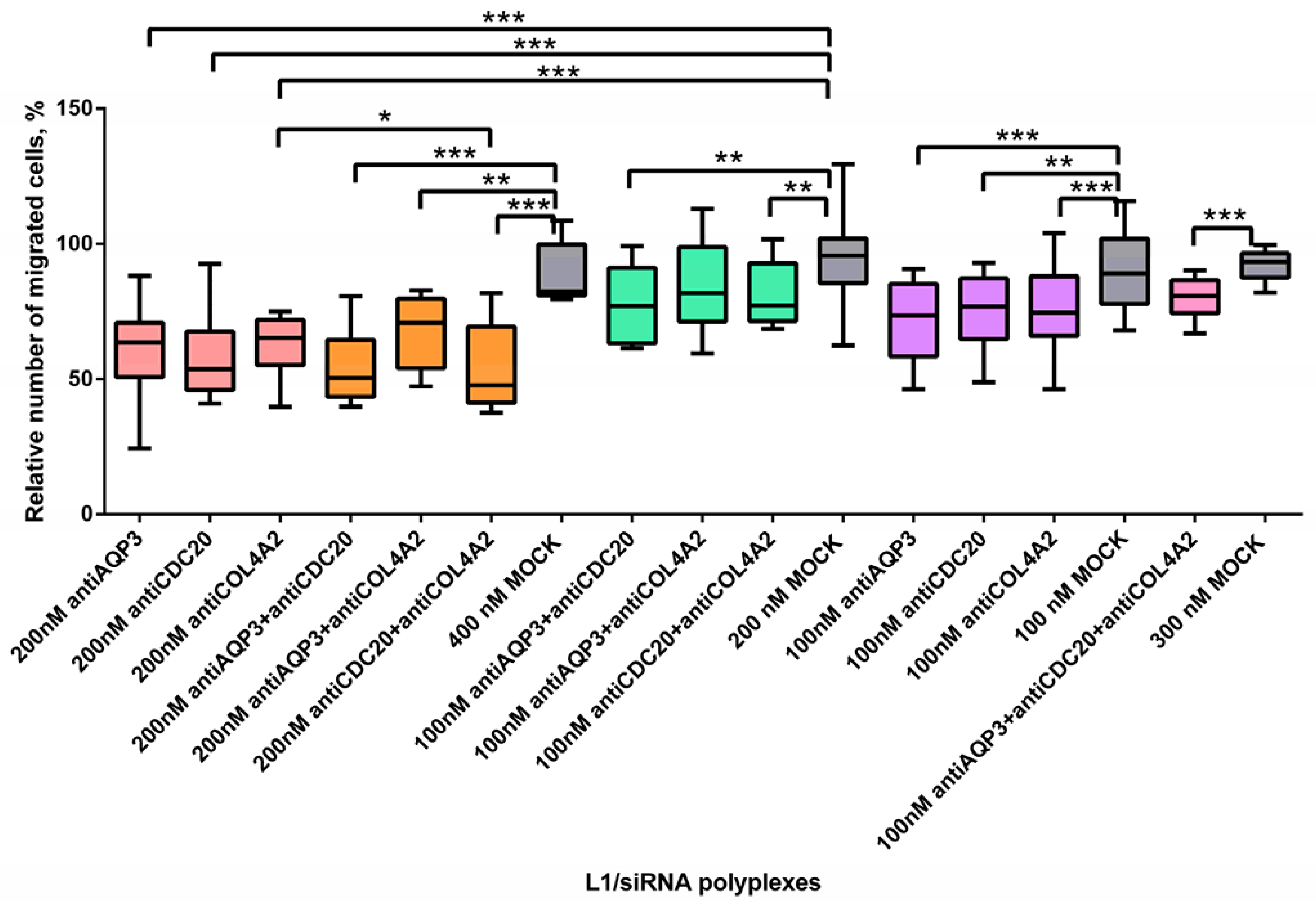
2.5. Inhibition of MDA-MB-231 Cells Migration
3. Materials and Methods
3.1. Cell Lines
3.2. Peptide Synthesis and Design
3.3. siRNA, Preparation of Peptide/siRNA Complexes
3.4. Cytotoxicity Assay
3.5. siRNA Transfer to MDA-MB-231 Cells
3.6. Quantitative RT-PCR
3.7. Proliferation Assay
3.8. Scratch Migration Assay
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patel, S. Breast cancer: Lesser-known facets and hypotheses. Biomed. Pharmacother. 2018, 98, 499–506. [Google Scholar] [CrossRef]
- Miller, K.D.; Fidler-Benaoudia, M.; Keegan, T.H.; Hipp, H.S.; Jemal, A.; Siegel, R.L. Cancer statistics for adolescents and young adults, 2020. CA Cancer J. Clin. 2020, 70, 443–459. [Google Scholar] [CrossRef]
- Ligresti, G.; Libra, M.; Militello, L.; Clementi, S.; Donia, M.; Imbesi, R.; Malaponte, G.; Cappellani, A.; McCubrey, J.A.; Stivala, F. Breast cancer: Molecular basis and therapeutic strategies (Review). Mol. Med. Rep. 2008, 1, 451–458. [Google Scholar] [CrossRef]
- Canine, B.F.; Wang, Y.; Hatefi, A. Biosynthesis and characterization of a novel genetically engineered polymer for targeted gene transfer to cancer cells. J. Control. Release 2009, 138, 188–196. [Google Scholar] [CrossRef]
- Nuciforo, P.; Radosevic-Robin, N.; Ng, T.; Scaltriti, M. Quantification of HER family receptors in breast cancer. Breast Cancer Res. 2015, 17, 53. [Google Scholar] [CrossRef]
- Masoud, V.; Pagès, G. Targeted therapies in breast cancer: New challenges to fight against resistance. World J. Clin. Oncol. 2017, 8, 120–134. [Google Scholar] [CrossRef]
- Rhee, J.; Han, S.-W.; Oh, D.-Y.; Kim, J.H.; Im, S.-A.; Han, W.; Ae Park, I.; Noh, D.-Y.; Bang, Y.-J.; Kim, T.-Y. The clinicopathologic characteristics and prognostic significance of triple-negativity in node-negative breast cancer. BMC Cancer 2008, 8, 307. [Google Scholar] [CrossRef]
- Vici, P.; Pizzuti, L.; Natoli, C.; Gamucci, T.; Di Lauro, L.; Barba, M.; Sergi, D.; Botti, C.; Michelotti, A.; Moscetti, L.; et al. Triple positive breast cancer: A distinct subtype? Cancer Treat. Rev. 2015, 41, 69–76. [Google Scholar] [CrossRef]
- Jiang, X.; Shapiro, D.J. The immune system and inflammation in breast cancer. Mol. Cell. Endocrinol. 2014, 382, 673–682. [Google Scholar] [CrossRef]
- Hon, J.D.C.; Singh, B.; Sahin, A.; Du, G.; Wang, J.; Wang, V.Y.; Deng, F.-M.; Zhang, D.Y.; Monaco, M.E.; Lee, P. Breast cancer molecular subtypes: From TNBC to QNBC. Am. J. Cancer Res. 2016, 6, 1864–1872. [Google Scholar]
- Chadar, R.; Afsana; Kesharwani, P. Nanotechnology-based siRNA delivery strategies for treatment of triple negative breast cancer. Int. J. Pharm. 2021, 605, 120835. [Google Scholar] [CrossRef]
- Amjad, M.W.; Kesharwani, P.; Mohd Amin, M.C.I.; Iyer, A.K. Recent advances in the design, development, and targeting mechanisms of polymeric micelles for delivery of siRNA in cancer therapy. Prog. Polym. Sci. 2017, 64, 154–181. [Google Scholar] [CrossRef]
- Ossovskaya, V.; Wang, Y.; Budoff, A.; Xu, Q.; Lituev, A.; Potapova, O.; Vansant, G.; Monforte, J.; Daraselia, N. Exploring Molecular Pathways of Triple-Negative Breast Cancer. Genes Cancer 2011, 2, 870–879. [Google Scholar] [CrossRef]
- Soni, N.; Soni, N.; Pandey, H.; Maheshwari, R.; Kesharwani, P.; Tekade, R.K. Augmented delivery of gemcitabine in lung cancer cells exploring mannose anchored solid lipid nanoparticles. J. Colloid Interface Sci. 2016, 481, 107–116. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Teicher, B.A.; Fricker, S.P. CXCL12 (SDF-1)/CXCR4 pathway in cancer. Clin. Cancer Res. 2010, 16, 2927–2931. [Google Scholar] [CrossRef]
- Yang, M.; Zeng, C.; Li, P.; Qian, L.; Ding, B.; Huang, L.; Li, G.; Jiang, H.; Gong, N.; Wu, W. Impact of CXCR4 and CXCR7 knockout by CRISPR/Cas9 on the function of triple-negative breast cancer cells. OncoTargets Ther. 2019, 12, 3849–3858. [Google Scholar] [CrossRef]
- Müller, A.; Homey, B.; Soto, H.; Ge, N.; Catron, D.; Buchanan, M.E.; McClanahan, T.; Murphy, E.; Yuan, W.; Wagner, S.N.; et al. Involvement of chemokine receptors in breast cancer metastasis. Nature 2001, 410, 50–56. [Google Scholar] [CrossRef]
- Ma, T.; Verkman, A.S. Aquaporin water channels in gastrointestinal physiology. J. Physiol. 1999, 517, 317–326. [Google Scholar] [CrossRef]
- Satooka, H.; Hara-Chikuma, M. Aquaporin-3 controls breast cancer cell migration by regulating hydrogen peroxide transport and its downstream cell signaling. Mol. Cell. Biol. 2016, 36, 1206–1218. [Google Scholar] [CrossRef]
- Verkman, A.S.; Hara-Chikuma, M.; Papadopoulos, M.C. Aquaporins—New players in cancer biology. J. Mol. Med. 2008, 86, 523–529. [Google Scholar] [CrossRef]
- Arif, M.; Kitchen, P.; Conner, M.T.; Hill, E.J.; Nagel, D.; Bill, R.M.; Dunmore, S.J.; Armesilla, A.L.; Gross, S.; Carmichael, A.R.; et al. Downregulation of aquaporin 3 inhibits cellular proliferation, migration and invasion in the MDA-MB-231 breast cancer cell line. Oncol. Lett. 2018, 16, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Yu, H. Cdc20: A WD40 Activator for a Cell Cycle Degradation Machine. Mol. Cell 2007, 27, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Kimata, Y.; Baxter, J.E.; Fry, A.M.; Yamano, H. A Role for the Fizzy/Cdc20 Family of Proteins in Activation of the APC/C Distinct from Substrate Recruitment. Mol. Cell 2008, 32, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Parmar, M.B.; Aliabadi, H.M.; Mahdipoor, P.; Kucharski, C.; Maranchuk, R.; Hugh, J.C.; Uludag, H. Targeting Cell Cycle Proteins in Breast Cancer Cells with siRNA by Using Lipid-Substituted Polyethylenimines. Front. Bioeng. Biotechnol. 2015, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Lowe, V.J.; Lee, S. Inhibition of Cdc20 suppresses the metastasis in triple negative breast cancer (TNBC). Breast Cancer 2021, 28, 1073–1086. [Google Scholar] [CrossRef]
- Parmar, M.B.; Remant Bahadur, K.C.; Löbenberg, R.; Uludag, H. Additive Polyplexes to Undertake siRNA Therapy against CDC20 and Survivin in Breast Cancer Cells. Biomacromolecules 2018, 19, 4193–4206. [Google Scholar] [CrossRef]
- Jeanne, M.; Jorgensen, J.; Gould, D.B. Molecular and genetic analyses of collagen type IV mutant mouse models of spontaneous intracerebral hemorrhage identify mechanisms for stroke prevention. Circulation 2015, 131, 1555–1565. [Google Scholar] [CrossRef]
- Zhu, J.; Pan, C.; Jiang, J.; Deng, M.; Gao, H.; Men, B.; McClelland, M.; Mercola, D.; Zhong, W.-D.; Jia, Z. Six stroma-based RNA markers diagnostic for prostate cancer in European-Americans validated at the RNA and protein levels in patients in China. Oncotarget 2015, 6, 16757–16765. [Google Scholar] [CrossRef][Green Version]
- Brown, C.W.; Brodsky, A.S.; Freiman, R.N. Notch3 Overexpression Promotes Anoikis Resistance in Epithelial Ovarian Cancer via Upregulation of COL4A2. Mol. Cancer Res. 2015, 13, 78–85. [Google Scholar] [CrossRef]
- Maekawa, R.; Sato, S.; Yamagata, Y.; Asada, H.; Tamura, I.; Lee, L.; Okada, M.; Tamura, H.; Takaki, E.; Nakai, A.; et al. Genome-Wide DNA Methylation Analysis Reveals a Potential Mechanism for the Pathogenesis and Development of Uterine Leiomyomas. PLoS ONE 2013, 8, e66632. [Google Scholar] [CrossRef]
- Wang, C.; Gao, C.; Zhuang, J.-L.; Ding, C.; Wang, Y. A combined approach identifies three mRNAs that are down-regulated by microRNA-29b and promote invasion ability in the breast cancer cell line MCF-7. J. Cancer Res. Clin. Oncol. 2012, 138, 2127–2136. [Google Scholar] [CrossRef]
- He, J.; Yang, J.; Chen, W.; Wu, H.; Yuan, Z.; Wang, K.; Li, G.; Sun, J.; Yu, L. Molecular features of triple negative breast cancer: Microarray evidence and further integrated analysis. PLoS ONE 2015, 10, e0129842. [Google Scholar] [CrossRef]
- He, J.; Hong, G.; Yang, J.; Zheng, D.; Li, F.; Chen, W.; Luo, X.; Mao, Y.; OuYang, Y.; Pan, Y.; et al. siRNA-Mediated suppression of collagen type iv alpha 2 (COL4A2) mRNA inhibits triple-negative breast cancer cell proliferation and migration. Oncotarget 2017, 8, 2585–2593. [Google Scholar] [CrossRef]
- David, S.; Pitard, B.; Benoît, J.-P.; Passirani, C. Non-viral nanosystems for systemic siRNA delivery. Pharmacol. Res. 2010, 62, 100–114. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Egorova, A.; Kiselev, A.; Hakli, M.; Ruponen, M.; Baranov, V.; Urtti, A. Chemokine-derived peptides as carriers for gene delivery to CXCR4 expressing cells. J. Gene Med. 2009, 11, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, A.; Egorova, A.; Laukkanen, A.; Baranov, V.; Urtti, A. Characterization of reducible peptide oligomers as carriers for gene delivery. Int. J. Pharm. 2013, 441, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Egorova, A.; Bogacheva, M.; Shubina, A.; Baranov, V.; Kiselev, A. Development of a receptor-targeted gene delivery system using CXCR4 ligand-conjugated cross-linking peptides. J. Gene Med. 2014, 16, 336–351. [Google Scholar] [CrossRef]
- Egorova, A.; Shubina, A.; Sokolov, D.; Selkov, S.; Baranov, V.; Kiselev, A. CXCR4-targeted modular peptide carriers for efficient anti-VEGF siRNA delivery. Int. J. Pharm. 2016, 515, 431–440. [Google Scholar] [CrossRef]
- Egorova, A.; Petrosyan, M.; Maretina, M.; Balashova, N.; Polyanskih, L.; Baranov, V.; Kiselev, A. Anti-angiogenic treatment of endometriosis via anti-VEGFA siRNA delivery by means of peptide-based carrier in a rat subcutaneous model. Gene Ther. 2018, 25, 548–555. [Google Scholar] [CrossRef]
- Egorova, A.A.; Shtykalova, S.V.; Maretina, M.A.; Sokolov, D.I.; Selkov, S.A.; Baranov, V.S.; Kiselev, A.V. Synergistic Anti-Angiogenic Effects Using Peptide-Based Combinatorial Delivery of siRNAs Targeting VEGFA, VEGFR1, and Endoglin Genes. Pharmaceutics 2019, 11, 261. [Google Scholar] [CrossRef]
- Wang, J.; Mi, P.; Lin, G.; Wáng, Y.X.J.; Liu, G.; Chen, X. Imaging-guided delivery of RNAi for anticancer treatment. Adv. Drug Deliv. Rev. 2016, 104, 44–60. [Google Scholar] [CrossRef]
- Peer, D.; Lieberman, J. Special delivery: Targeted therapy with small RNAs. Gene Ther. 2011, 18, 1127–1133. [Google Scholar] [CrossRef]
- Liang, S.; Peng, X.; Li, X.; Yang, P.; Xie, L.; Li, Y.; Du, C.; Zhang, G. Silencing of CXCR4 sensitizes triple-negative breast cancer cells to cisplatin. Oncotarget 2015, 6, 1020–1030. [Google Scholar] [CrossRef]
- Ren, Y.; Jiang, X.; Pan, D.; Mao, H.-Q. Charge density and molecular weight of polyphosphoramidate gene carrier are key parameters influencing its DNA compaction ability and transfection efficiency. Biomacromolecules 2010, 11, 3432–3439. [Google Scholar] [CrossRef]
- Nikcevic, G.; Kovacevic-Grujicic, N.; Stevanovic, M. Improved transfection efficiency of cultured human cells. Cell Biol. Int. 2003, 27, 735–737. [Google Scholar] [CrossRef]
- Mayer, I.A.; Abramson, V.G.; Lehmann, B.D.; Pietenpol, J.A. New strategies for triple-negative breast cancer-deciphering the heterogeneity. Clin. Cancer Res. 2014, 20, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.; Custodio, S.; Maestro De Las Casas, M.-L.; García-Sáenz, J.-A.; De La Torre, J.-C.; Bellón-Cano, J.-M.; López-Tarruella, S.; Vidaurreta-Lazaro, M.; De La Orden, V.; Jerez, Y.; et al. Circulating tumor cells following first chemotherapy cycle: An early and strong predictor of outcome in patients with metastatic breast cancer. Oncologist 2013, 18, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhu, Z.; Sun, M.; Wang, J.; Guo, R.; Shen, L.; Wu, W. Critical role of aquaporin-3 in the human epidermal growth factor-induced migration and proliferation in the human gastric adenocarcinoma cells. Cancer Biol. Ther. 2010, 9, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Ma, Y.-F.; Yu, C.-H.; Li, Y.-J.; Tang, J.; Li, J.-B.; Zhao, Y.-N.; Liu, Y. Aquaporin 3 knockdown suppresses tumour growth and angiogenesis in experimental non-small cell lung cancer. Exp. Physiol. 2014, 99, 974–984. [Google Scholar] [CrossRef]
- Tang, J.; Lu, M.; Cui, Q.; Zhang, D.; Kong, D.; Liao, X.; Ren, J.; Gong, Y.; Wu, G. Overexpression of ASPM, CDC20, and TTK Confer a Poorer Prognosis in Breast Cancer Identified by Gene Co-expression Network Analysis. Front. Oncol. 2019, 9, 310. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, Y.; Williams, J.; Hang, Y.; Richter, L.; Becker, M.; Amador, C.; Oupický, D.; Hyde, R.K. Use of polymeric CXCR4 inhibitors as siRNA delivery vehicles for the treatment of acute myeloid leukemia. Cancer Gene Ther. 2020, 27, 45–55. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Y.; Li, J.; Hang, Y.; Oupický, D. Promise of chemokine network-targeted nanoparticles in combination nucleic acid therapies of metastatic cancer. WIREs Nanomed. Nanobiotechnol. 2019, 11, e1528. [Google Scholar] [CrossRef]
- Castanotto, D.; Sakurai, K.; Lingeman, R.; Li, H.; Shively, L.; Aagaard, L.; Soifer, H.; Gatignol, A.; Riggs, A.; Rossi, J.J. Combinatorial delivery of small interfering RNAs reduces RNAi efficacy by selective incorporation into RISC. Nucleic Acids Res. 2007, 35, 5154–5164. [Google Scholar] [CrossRef]
- Su, S.; Tian, Y.; Li, Y.; Ding, Y.; Ji, T.; Wu, M.; Wu, Y.; Nie, G. “Triple-Punch” Strategy for Triple Negative Breast Cancer Therapy with Minimized Drug Dosage and Improved Antitumor Efficacy. ACS Nano 2015, 9, 1367–1378. [Google Scholar] [CrossRef]
- Entschladen, F.; Drell, T.L., IV; Lang, K.; Joseph, J.; Zaenker, K.S. Tumour-cell migration, invasion, and metastasis: Navigation by neurotransmitters. Lancet Oncol. 2004, 5, 254–258. [Google Scholar] [CrossRef]
- Cao, X.-C.; Zhang, W.-R.; Cao, W.-F.; Liu, B.-W.; Zhang, F.; Zhao, H.-M.; Meng, R.; Zhang, L.; Niu, R.-F.; Hao, X.-S.; et al. Aquaporin3 Is Required for FGF-2-Induced Migration of Human Breast Cancers. PLoS ONE 2013, 8, e56735. [Google Scholar] [CrossRef]
- Cheng, S.; Castillo, V.; Sliva, D. CDC20 associated with cancer metastasis and novel mushroom-derived CDC20 inhibitors with antimetastatic activity. Int. J. Oncol. 2019, 54, 2250–2256. [Google Scholar] [CrossRef]
- Bakhoum, S.F.; Cantley, L.C. The Multifaceted Role of Chromosomal Instability in Cancer and Its Microenvironment. Cell 2018, 174, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Paul, D.; Ghorai, S.; Dinesh, U.S.; Shetty, P.; Chattopadhyay, S.; Santra, M.K. Cdc20 directs proteasome-mediated degradation of the tumor suppressor SMAR1 in higher grades of cancer through the anaphase promoting complex. Cell Death Dis. 2017, 8, e2882. [Google Scholar] [CrossRef] [PubMed]
- Poage, G.M.; Hartman, Z.C.; Brown, P.H. Revealing targeted therapeutic opportunities in triple-negative breast cancers: A new strategy. Cell Cycle 2013, 12, 2705–2706. [Google Scholar] [CrossRef]
- Slita, A.; Egorova, A.; Casals, E.; Kiselev, A.; Rosenholm, J.M. Characterization of modified mesoporous silica nanoparticles as vectors for siRNA delivery. Asian J. Pharm. Sci. 2018, 13, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhang, Y.; Chen, H.; Guo, Z. MicroRNA-488 inhibits proliferation, invasion and EMT in osteosarcoma cell lines by targeting aquaporin 3. Int. J. Oncol. 2018, 53, 1493–1504. [Google Scholar] [CrossRef]
- Hemati, M.; Haghiralsadat, F.; Jafary, F.; Moosavizadeh, S.; Moradi, A. Targeting cell cycle protein in gastric cancer with CDC20siRNA and anticancer drugs (Doxorubicin and quercetin) co-loaded cationic PEGylated nanoniosomes. Int. J. Nanomed. 2019, 14, 6575–6585. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Yang, H.; Wu, J.; Wang, A.; Chen, X.; Hu, S.; Zhang, Y.; Bai, D.; Jin, Z. COL4A2 in the tissue-specific extracellular matrix plays important role on osteogenic differentiation of periodontal ligament stem cells. Theranostics 2019, 9, 4265–4286. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, L.; Yeh, J.; Wu, X.; Cao, Z.; Wang, Y.A.; Zhang, M.; Yang, L.; Mao, H. Reducing non-specific binding and uptake of nanoparticles and improving cell targeting with an antifouling PEO-b-PγMPS copolymer coating. Biomaterials 2010, 31, 5397–5407. [Google Scholar] [CrossRef]
- Wu, J.; Qu, L.; Meng, L.; Zeng, Y.; Shou, C.; Xu, H.; Jiang, B.; Ren, T. N-α-Acetyltransferase 10 protein inhibits apoptosis through RelA/p65-regulated MCL1 expression. Carcinogenesis 2012, 33, 1193–1202. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Analysis | Type of siRNA | |||||
|---|---|---|---|---|---|---|
| AntiAQP3 | AntiCDC20 | AntiCOL4A2 | AntiAQP3 + AntiCDC20 | AntiAQP3 + AntiCOL4A2 | AntiCDC20 + AntiCOL4A2 | |
| Proliferation | - | ↓ | ↓ | ↓↓ | ↓↓ | ↓↓↓ |
| Migration | ↓ | ↓ | ↓ | ↓ | ↓ | ↓↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, A.; Pyankov, I.; Maretina, M.; Baranov, V.; Kiselev, A. Peptide Nanoparticle-Mediated Combinatorial Delivery of Cancer-Related siRNAs for Synergistic Anti-Proliferative Activity in Triple Negative Breast Cancer Cells. Pharmaceuticals 2021, 14, 957. https://doi.org/10.3390/ph14100957
Egorova A, Pyankov I, Maretina M, Baranov V, Kiselev A. Peptide Nanoparticle-Mediated Combinatorial Delivery of Cancer-Related siRNAs for Synergistic Anti-Proliferative Activity in Triple Negative Breast Cancer Cells. Pharmaceuticals. 2021; 14(10):957. https://doi.org/10.3390/ph14100957
Chicago/Turabian StyleEgorova, Anna, Ivan Pyankov, Marianna Maretina, Vladislav Baranov, and Anton Kiselev. 2021. "Peptide Nanoparticle-Mediated Combinatorial Delivery of Cancer-Related siRNAs for Synergistic Anti-Proliferative Activity in Triple Negative Breast Cancer Cells" Pharmaceuticals 14, no. 10: 957. https://doi.org/10.3390/ph14100957
APA StyleEgorova, A., Pyankov, I., Maretina, M., Baranov, V., & Kiselev, A. (2021). Peptide Nanoparticle-Mediated Combinatorial Delivery of Cancer-Related siRNAs for Synergistic Anti-Proliferative Activity in Triple Negative Breast Cancer Cells. Pharmaceuticals, 14(10), 957. https://doi.org/10.3390/ph14100957