
In Silico Studies of Potential Selective Inhibitors of Thymidylate Kinase from Variola virus

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
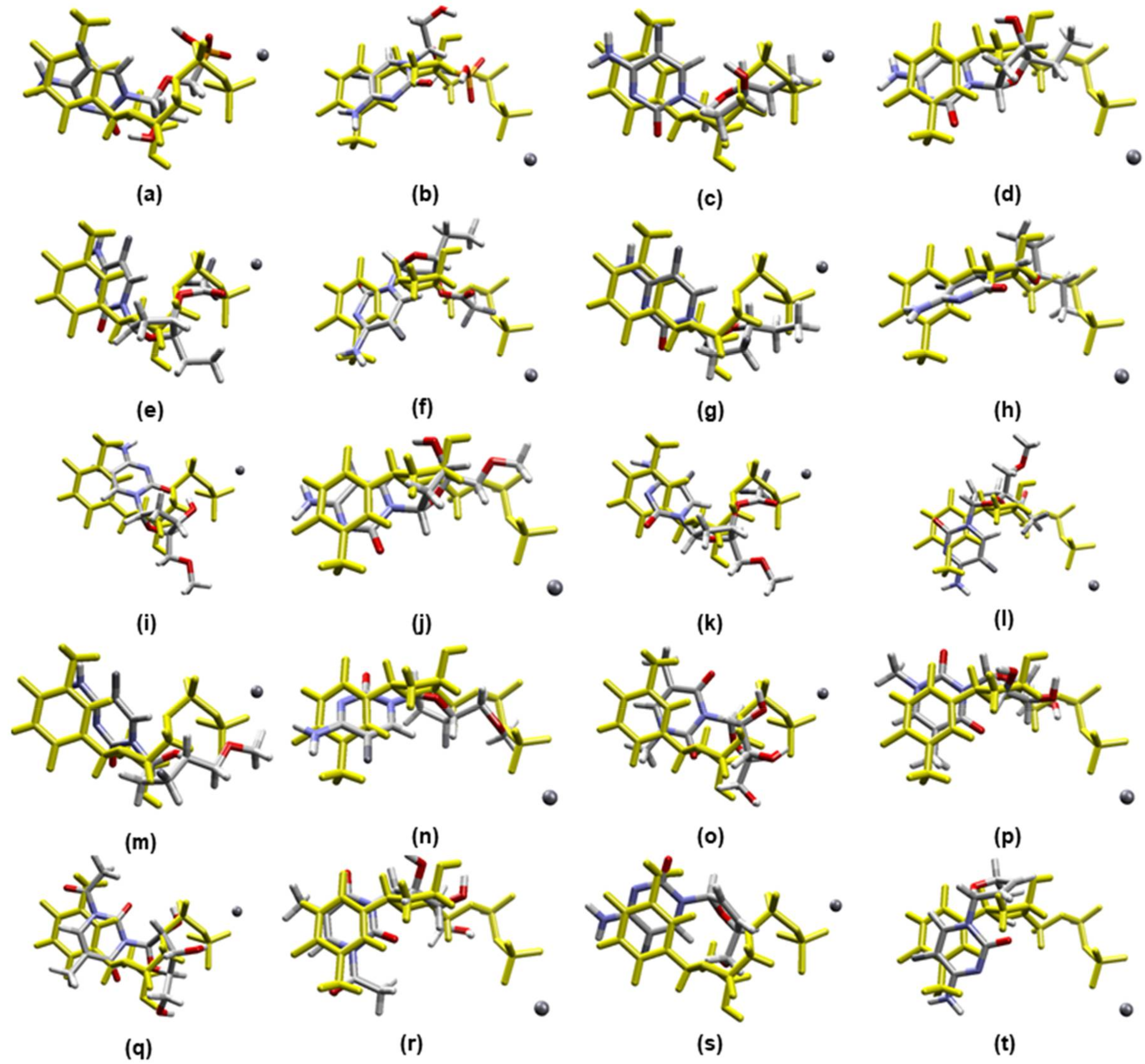
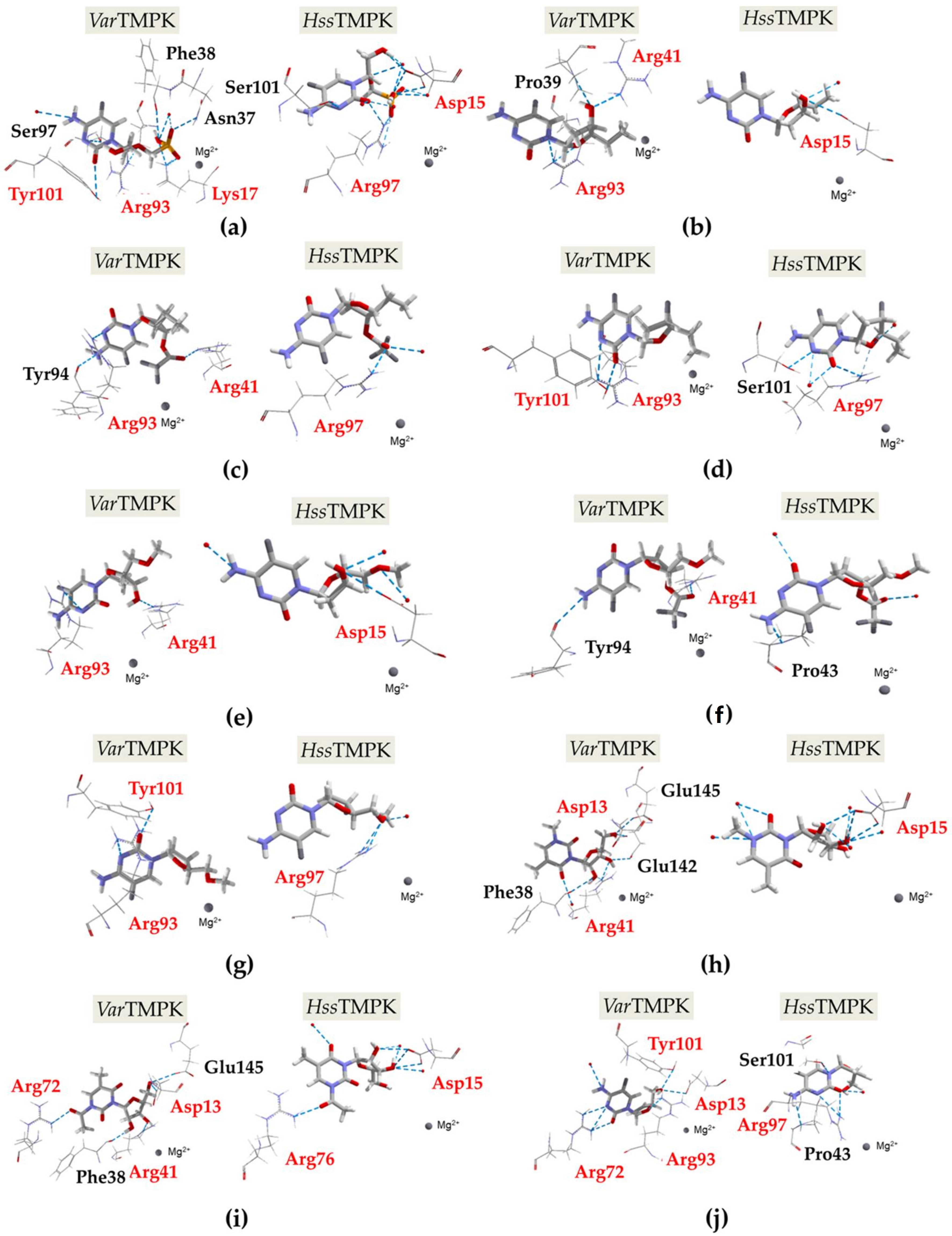
2.1. Docking Calculations
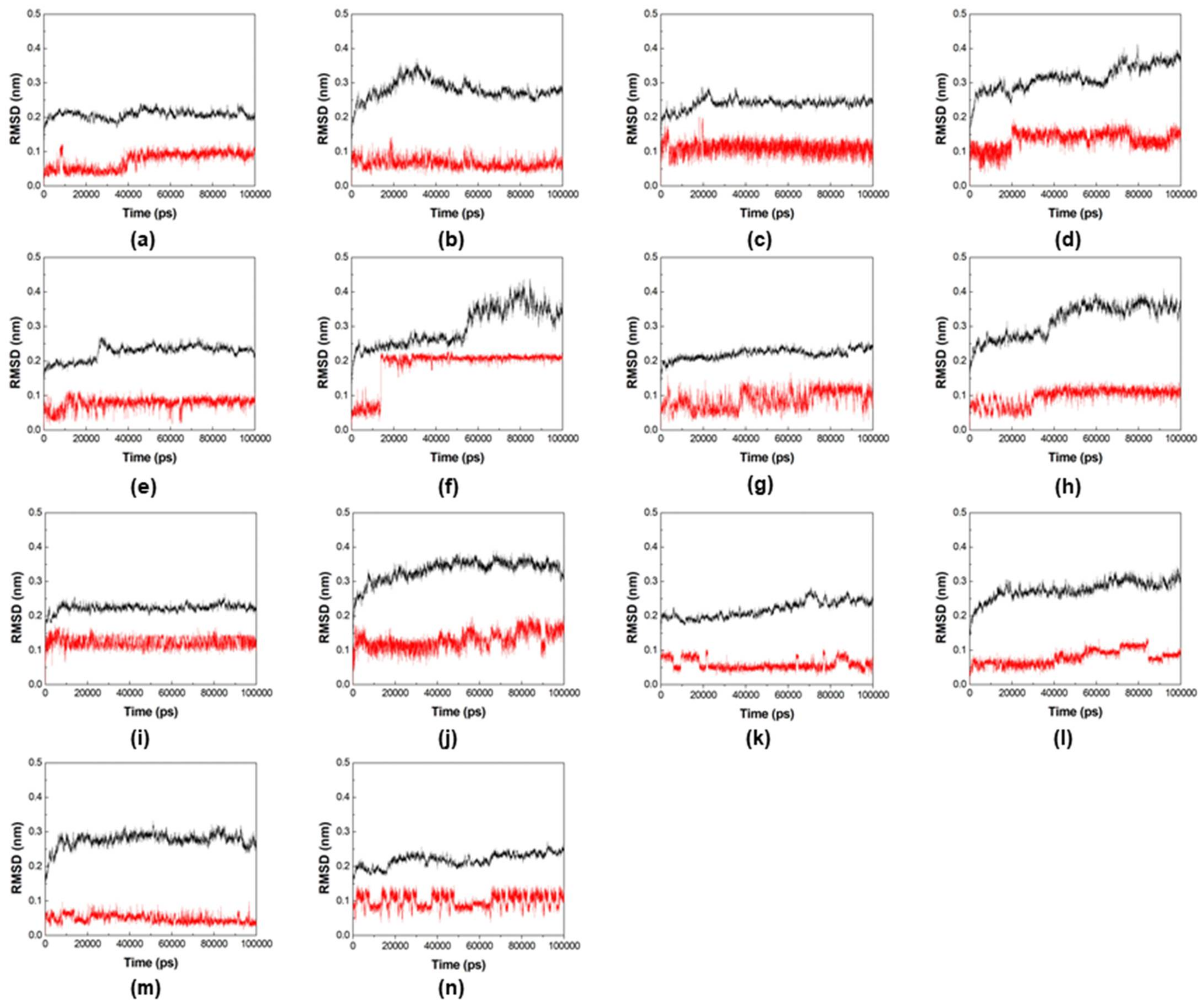
2.2. Molecular Dynamics Simulations
2.3. MM-PBSA Calculations
3. Discussion
4. Materials and Methods
4.1. Compounds Studied
4.2. Applied Protocols
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Russo, A.T.; Berhanu, A.; Bigger, C.B.; Prigge, J.; Silvera, P.M.; Grosenbach, D.W.; Hruby, D. Co-administration of tecovirimat and ACAM2000™ in non-human primates: Effect of tecovirimat treatment on ACAM2000 immunogenicity and efficacy versus lethal monkeypox virus challenge. Vaccine 2020, 38, 644–654. [Google Scholar] [CrossRef]
- Chan-Tack, K.M.; Harrington, P.R.; Choi, S.-Y.; Myers, L.; O’Rear, J.; Seo, S.; McMillan, D.; Ghantous, H.; Birnkrant, D.; Sherwat, A.I. Assessing a drug for an eradicated human disease: US Food and Drug Administration review of tecovirimat for the treatment of smallpox. Lancet Infect. Dis. 2019, 19, e221–e224. [Google Scholar] [CrossRef]
- Bastos, L.D.C.; De Souza, F.R.; Guimarães, A.P.; Sirouspour, M.; Guizado, T.R.C.; Forgione, P.; Ramalho, T.C.; França, T.C.C. Virtual screening, docking, and dynamics of potential new inhibitors of dihydrofolate reductase from Yersinia pestis. J. Biomol. Struct. Dyn. 2016, 34, 2184–2198. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.P.; de Souza, F.R.; Oliveira, A.A.; Gonçalves, A.S.; de Alencastro, R.B.; Ramalho, T.C.; França, T.C.C. Design of inhibitors of thymidylate kinase from Variola virus as new selective drugs against smallpox. Eur. J. Med. Chem. 2015, 91, 72–90. [Google Scholar] [CrossRef] [PubMed]
- Trost, L.C.; Rose, M.L.; Khouri, J.; Keilholz, L.; Long, J.; Godin, S.J.; Foster, S.A. The efficacy and pharmacokinetics of brincidofovir for the treatment of lethal rabbitpox virus infection: A model of smallpox disease. Antivir. Res. 2015, 117, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damon, I.K.; Damaso, C.R.; McFadden, G. Are We There Yet? The Smallpox Research Agenda Using Variola Virus. PLoS Pathog. 2014, 10, e1004108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, A.P.; Ramalho, T.C.; França, T.C.C. Preventing the return of smallpox: Molecular modeling studies on thymidylate kinase fromVariola virus. J. Biomol. Struct. Dyn. 2014, 32, 1601–1612. [Google Scholar] [CrossRef] [PubMed]
- Prichard, M.N.; Kern, E.R. Orthopoxvirus targets for the development of new antiviral agents. Antivir. Res. 2012, 94, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Garcia, D.R.; De Souza, F.R.; Guimarães, A.P.; Ramalho, T.C.; De Aguiar, A.P.; França, T.C.C. Design of inhibitors of thymidylate kinase from Variola virus as new selective drugs against smallpox: Part II. J. Biomol. Struct. Dyn. 2019, 37, 4569–4579. [Google Scholar] [CrossRef]
- Merchlinsky, M.; Albright, A.; Olson, V.; Schiltz, H.; Merkeley, T.; Hughes, C.; Petersen, B.; Challberg, M. The development and approval of tecoviromat (TPOXX®), the first antiviral against smallpox. Antivir. Res. 2019, 168, 168–174. [Google Scholar] [CrossRef]
- Nuth, M.; Guan, H.; Xiao, Y.; Kulp, J.L., III; Parker, M.H.; Strobel, E.D.; Isaacs, S.N.; Scott, R.W.; Reitz, A.B.; Ricciardi, R.P. Mutation and structure guided discovery of an antiviral small molecule that mimics an essential C-Terminal tripeptide of the vaccinia D4 processivity factor. Antivir. Res. 2018, 162, 178–185. [Google Scholar] [CrossRef]
- Chaudhuri, S.; Symons, J.A.; Deval, J. Innovation and trends in the development and approval of antiviral medicines: 1987–2017 and beyond. Antivir. Res. 2018, 155, 76–88. [Google Scholar] [CrossRef]
- Noyce, R.S.; Lederman, S.; Evans, D.H. Construction of an infectious horsepox virus vaccine from chemically synthesized DNA fragments. PLoS ONE 2018, 13, e0188453. [Google Scholar] [CrossRef]
- Chittick, G.; Morrison, M.; Brundage, T.; Nichols, W.G. Short-term clinical safety profile of brincidofovir: A favorable benefit–risk proposition in the treatment of smallpox. Antivir. Res. 2017, 143, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Crump, R.; Korom, M.; Buller, R.M.; Parker, S. Buccal viral DNA as a trigger for brincidofovir therapy in the mousepox model of smallpox. Antivir. Res. 2017, 139, 112–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossi, I.M.; Foster, S.A.; Gainey, M.R.; Krile, R.T.; Dunn, J.A.; Brundage, T.; Khouri, J.M. Efficacy of delayed brincidofovir treatment against a lethal rabbitpox virus challenge in New Zealand White rabbits. Antivir. Res. 2017, 143, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Arita, I. Discovery of forgotten variola specimens at the National Institutes of Health in the USA. Expert Rev. Anti-Infect. Ther. 2014, 12, 1419–1421. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Tatay, L.; Hernández-Andreu, J.M. Biosafety and biosecurity in synthetic biology: A review. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1587–1621. [Google Scholar] [CrossRef]
- Esparza, J.; Schrick, L.; Damaso, C.R.; Nitsche, A. Equination (inoculation of horsepox): An early alternative to vaccination (inoculation of cowpox) and the potential role of horsepox virus in the origin of the smallpox vaccine. Vaccine 2017, 35, 7222–7230. [Google Scholar] [CrossRef]
- Schoepp, R.J.; Morin, M.D.; Martinez, M.J.; Kulesh, D.A.; Hensley, L.; Geisbert, T.W.; Brady, D.R.; Jahrling, P.B. Detection and identification of Variola virus in fixed human tissue after prolonged archival storage. Lab. Investig. 2004, 84, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Berche, P. The threat of smallpox and bioterrorism. Trends Microbiol. 2001, 9, 15–18. [Google Scholar] [CrossRef]
- Chen, D.; Qi, E.Y. Innovative highlights of clinical drug trial design. Transl. Res. 2020, 224, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Mucker, E.M.; Goff, A.J.; Shamblin, J.D.; Grosenbach, D.W.; Damon, I.K.; Mehal, J.M.; Holman, R.C.; Carroll, D.; Gallardo, N.; Olson, V.A.; et al. Efficacy of Tecovirimat (ST-246) in Nonhuman Primates Infected with Variola Virus (Smallpox). Antimicrob. Agents Chemother. 2013, 57, 6246–6253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caillat, C.; Topalis, D.; Agrofoglio, L.A.; Pochet, S.; Balzarini, J.; Deville-Bonne, D.; Meyer, P. Crystal structure of poxvirus thymidylate kinase: An unexpected dimerization has implications for antiviral therapy. Proc. Natl. Acad. Sci. USA 2008, 105, 16900–16905. [Google Scholar] [CrossRef] [Green Version]
- Auvynet, C.; Topalis, D.; Caillat, C.; Munier-Lehmann, H.; Seclaman, E.; Balzarini, J.; Agrofoglio, L.A.; Kaminski, P.A.; Meyer, P.R.; Deville-Bonne, D.; et al. Phosphorylation of dGMP analogs by vaccinia virus TMP kinase and human GMP kinase. Biochem. Biophys. Res. Commun. 2009, 388, 6–11. [Google Scholar] [CrossRef]
- Topalis, D.; Collinet, B.; Gasse, C.; Dugué, L.; Balzarini, J.; Pochet, S.; Deville-Bonne, D. Substrate specificity of vaccinia virus thymidylate kinase. FEBS J. 2005, 272, 6254–6265. [Google Scholar] [CrossRef]
- Cui, Q.; Shin, W.S.; Luo, Y.; Tian, J.; Cui, H.; Yin, D. Thymidylate Kinase: An Old Topic Brings New Perspectives. Curr. Med. Chem. 2013, 20, 1286–1305. [Google Scholar] [CrossRef]
- Irwin, C.R.; Hitt, M.M.; Evans, D.H. Targeting Nucleotide Biosynthesis: A Strategy for Improving the Oncolytic Potential of DNA Viruses. Front. Oncol. 2017, 7, 229. [Google Scholar] [CrossRef] [Green Version]
- El Omari, K.; Solaroli, N.; Karlsson, A.; Balzarini, J.; Stammers, D.K. Structure of vaccinia virus thymidine kinase in complex with dTTP: Insights for drug design. BMC Struct. Biol. 2006, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Solaroli, N.; Johansson, M.; Persoons, L.; Balzarini, J.; Karlsson, A. Substrate specificity of feline and canine herpesvirus thymidine kinase. Antivir. Res. 2008, 79, 128–132. [Google Scholar] [CrossRef]
- Baker, R.O.; Bray, M.; Huggins, J.W. Potential antiviral therapeutics for smallpox, monkeypox and other orthopoxvirus infections. Antivir. Res. 2003, 57, 13–23. [Google Scholar] [CrossRef]
- Bray, M. Pathogenesis and potential antiviral therapy of complications of smallpox vaccination. Antivir. Res. 2003, 58, 101–114. [Google Scholar] [CrossRef]
- Bursulaya, B.D.; Totrov, M.; Abagyan, R.; Brooks, C.L., III. Comparative study of several algorithms for flexible ligand docking. J. Comput. Aided Mol. Des. 2003, 17, 755–763. [Google Scholar] [CrossRef]
- Huang, S.-Y.; Zou, X. Advances and Challenges in Protein-Ligand Docking. Int. J. Mol. Sci. 2010, 11, 3016–3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Greene, D.A.; Xiao, L.; Qi, R.; Luo, R. Recent Developments and Applications of the MMPBSA Method. Front. Mol. Biosci. 2018, 4, 87. [Google Scholar] [CrossRef] [Green Version]
- Genheden, S.; Kuhn, O.; Mikulskis, P.; Hoffmann, D.; Ryde, U. The normal-mode entropy in the MM/GBSA method: Effect of system truncation, buffer region, and dielectric constant. J. Chem. Inf. Model. 2012, 52, 2079–2088. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Kumar, R.; Lynn, A.; Open Source Drug Discovery Consortium. G-mmpbsa -A GROMACS tool for high-throughput MM-PBSA calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef]
- Kar, P.; Lipowsky, R.; Knecht, V. Importance of Polar Solvation and Configurational Entropy for Design of Antiretroviral Drugs Targeting HIV-1 Protease. J. Phys. Chem. B 2013, 117, 5793–5805. [Google Scholar] [CrossRef]
- Evertts, A.G.; Zee, B.M.; Garcia, B.A. Modern approaches for investigating epigenetic signaling pathways. J. Appl. Physiol. 2010, 109, 927–933. [Google Scholar] [CrossRef] [Green Version]
- Norambuena, T.; Melo, F. The Protein-DNA Interface database. BMC Bioinform. 2010, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Molnar, L.F.; Jung, Y.; Kussmann, J.; Ochsenfeld, C.; Brown, S.T.; Gilbert, A.T.; Slipchenko, L.V.; Levchenko, S.V.; O’Neill, D.P.; et al. Advances in methods and algorithms in a modern quantum chemistry program package. Phys. Chem. Chem. Phys. 2006, 8, 3172–3191. [Google Scholar] [CrossRef] [PubMed]
- Luscombe, N.M.; Austin, S.E.; Berman, H.M.; Thornton, J.M. An overview of the structures of protein-DNA complexes. Genome Biol. 2000, 1, REVIEWS001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaram, B.; Sprous, D.; Young, M.; Beveridge, D.L. Free Energy Analysis of the Conformational Preferences of A and B Forms of DNA in Solution. J. Am. Chem. Soc. 1998, 120, 10629–10633. [Google Scholar] [CrossRef]
- Bren, U.; Martínek, V.; Florián, J. Decomposition of the Solvation Free Energies of Deoxyribonucleoside Triphosphates Using the Free Energy Perturbation Method. J. Phys. Chem. B 2006, 110, 12782–12788. [Google Scholar] [CrossRef] [PubMed]
- Bren, M.; Florián, J.; Mavri, J.; Bren, U. Do all pieces make a whole? Thiele cumulants and the free energy decomposition. Theor. Chem. Acc. 2007, 117, 535–540. [Google Scholar] [CrossRef]
- Swain, M. chemicalize.org. J. Chem. Inf. Model. 2012, 52, 613–615. [Google Scholar] [CrossRef]
- Poli, G.; Granchi, C.; Rizzolio, F.; Tuccinardi, T. Application of MM-PBSA Methods in Virtual Screening. Molecules 2020, 25, 1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, C.A. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Rocha, G.B.; Freire, R.O.; Simas, A.M.; Stewart, J.J.P. RM1: A reparameterization of AM1 for H, C, N, O, P, S, F, Cl, Br, and I. J. Comput. Chem. 2006, 27, 1101–1111. [Google Scholar] [CrossRef]
- Gonçalves, A.D.S.; França, T.C.C.; Figueroa-Villar, J.D.; Pascutti, P.G. Conformational Analysis of Toxogonine, TMB-4 and HI-6 using PM6 and RM1 methods. J. Braz. Chem. Soc. 2010, 21, 179–184. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, R.; Christensen, M.H. MolDock: A New Technique for High-Accuracy Molecular Docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef]
- Kaminski, G.A.; Friesner, R.A.; Tirado-Rives, J.; Jorgensen, W.L. Evaluation and Reparametrization of the OPLS-AA Force Field for Proteins via Comparison with Accurate Quantum Chemical Calculations on Peptides†. J. Phys. Chem. B 2001, 105, 6474–6487. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. –GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Byrd, R.H.; Lu, P.; Nocedal, J.; Zhu, C. A Limited Memory Algorithm for Bound Constrained Optimization. SIAM J. Sci. Comput. 1995, 16, 1190–1208. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Da Silva, A.W.S.; Vranken, W.F. ACPYPE-Antechamber python parser interface. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VarTMPK | Asp13, Lys14, Ser15, Lys17, Thr18, Arg41, Leu53, Asn65, Phe68, Cys69, Arg72, Arg93, Tyr101, Ala102 |
| HssTMPK | Asp15, Arg16, Ala17, Lys19, Ser20, Arg45, Leu57, His69, Phe72, Ser73, Arg76, Arg97, Phe105, Thr106 |
| Structure | Acute Toxicity (Algae) | Carcinogenicity (Mouse) | Meets Lipinski’s Rule? |
|---|---|---|---|
| 1 | 1.10 | negative | Yes |
| 2 | 0.30 | positive | Yes |
| 3 | 0.15 | positive | Yes |
| 4 | 0.22 | positive | Yes |
| 5 | 0.51 | positive | Yes |
| 6 | 0.25 | positive | Yes |
| 7 | 0.38 | positive | Yes |
| 8 | 0.31 | positive | Yes |
| 9 | 0.37 | positive | Yes |
| 10 | 0.60 | positive | Yes |
| Cidofovir | 1.19 | Positive | Yes |
| Ribavirin | 0.54 | negative | Yes |
| # | Einteraction (kcal.mol−1) | Ecofactor (kcal.mol−1) | H-Bond Energy (kcal.mol−1) | H-Bond Interactions | ∆Eint. (kcal.mol−1) | ||||
|---|---|---|---|---|---|---|---|---|---|
| VarTMPK | HssTMPK | VarTMPK | HssTMPK | VarTMPK | HssTMPK | VarTMPK | HssTMPK | ||
| 1 | −100.00 | −128.35 | −25.04 | −4.13 | −14.74 | −15.18 | Asn37 (2.91Å) Lys17 (2.08Å; 2.98Å) Phe38 (3.23Å) Arg93 (2.87Å) Tyr101 (3.06Å) Ser97 (3.36Å) H2O (2.67Å; 3.20Å) | Asp15 (3.10Å; 2.97Å; 2.97Å) Arg97 (2.60Å; 2.60Å; 3.05Å) Ser101 (2.93Å) H2O (3.21Å; 2.49Å; 2.52Å; 2.96Å; 2.94Å; 2.87Å; 3.33Å) | +28.35 |
| 2 | −95.43 | −32.58 | −0.24 | −0.19 | −7.93 | −2.50 | Pro39 (3.06Å) Arg93 (2.88Å; 3.10Å) Arg41 (2.76Å) | Asp15 (3.10Å)H2O (3.38Å) | −62.85 |
| 3 | −110.95 | −75.54 | −2.45 | −0.34 | −6.44 | −1.58 | Tyr94 (3.11Å) Arg93 (3.10Å) Arg41 (2.61Å) | Arg97 (2.65Å) H2O (2.84Å) | −35.41 |
| 4 | −89.18 | −50.61 | −0.61 | −0.53 | −4.84 | −2.73 | Arg93 (3.10Å) Tyr101 (3.13Å) | Arg97 (2.6Å; 2.85Å) Ser101 (2.49Å) H2O (2.46Å; 3.43Å; 3.38Å) | −38.57 |
| 5 | −90.77 | −35.37 | −1.47 | −0.21 | −3.86 | −2.58 | Arg41 (3.09Å) Arg93 (3.10Å) | Asp15 (3.10Å) H2O (2.45Å; 3.02Å; 3.09Å) | −55.40 |
| 6 | −112.24 | −84.39 | −2.64 | −0.17 | −3.32 | −2.50 | Tyr94 (3.32Å) Arg41(2.69Å) | Pro43 (2.64Å) H2O (2.93Å; 3.33Å) | −27.85 |
| 7 | −41.11 | −58.87 | −1.33 | −0.47 | −4.22 | −3.70 | Arg93 (3.11Å) Tyr101 (2.63Å) | Arg97 (3.10Å; 3.12Å) H2O (3.27Å) | +17.76 |
| 8 | −106.20 | −94.88 | −0.63 | −0.28 | −12.93 | −7.79 | Phe38 (3.17Å) Arg41 (3.06Å)Glu142 (3.10Å) Asp13 (3.10Å)Glu145 (2.65Å) H2O (3.05Å) | Asp15 (2.72Å; 3.10Å; 3.10Å) H2O (3.28Å; 2.77Å; 3.21Å; 2.94Å; 3.33Å; 3.25Å) | −11.32 |
| 9 | −120.94 | −75.04 | −1.23 | −0.08 | −12.33 | −10.00 | Arg72 (2.70Å) Phe38 (2.90Å) Arg41 (2.98Å) Asp13 (3.10Å) Glu145 (3.10Å) | Arg76 (2.60Å) Asp15 (3.10Å; 3.00Å; 3.00Å) H2O (2.67Å; 3.30Å; 2.73Å; 3.04Å) | −45.90 |
| 10 | −44.85 | −101.15 | −0.80 | −0.64 | −8.63 | −7.40 | Arg72 (2.75Å; 2.88Å; 3.19Å) Arg93 (3.10Å) Tyr101 (3.10Å)Asp13 (3.10Å) H2O (2.84Å) | Pro43 (3.28Å) Arg97 (3.24Å; 2.67Å) Ser101 (3.24Å) H2O (3.36Å) | 56.30 |
| TDP | 228.18 | −195.60 | −50.38 | −18.13 | −3.82 | −13.97 | Asp13, Lys17, Asn37, Arg41, Arg72, Arg93 | Asp15, Arg45, Arg76, Arg97, H2O | |
| Binding Energies kJ.mol−1 | ||
|---|---|---|
| Compound | VarTMPK | HssTMPK |
| 2 | −133.69 ± 6.92 | −36.60 ± 3.71 |
| 3 | −189.90 ± 3.05 | −90.79 ± 3.56 |
| 4 | −59.50 ± 4.37 | −33.50 ± 4.41 |
| 6 | −206.04 ± 12.98 | −100.85 ± 2.24 |
| 9 | −81.72 ± 2.58 | −43.16 ± 4.36 |
| Docking | Software Used in Energy Calculations | Algorithm Used in Energy Calculations | Radius of the Anchoring Area | Enzyme Site Coordinates | No. of Repetitions | No. of Poses | |
|---|---|---|---|---|---|---|---|
| HssTMPK | VarTMPK | ||||||
| Molegro virtual docker (MVD®) | MolDock [52] | 6–10 Å | x = 8.95 y = 22.41 z = 0.69 | x = 13.92 y = 75.19 z = 25.05 | 10 | 30 | |
| Molecular Dynamics | Force Field | Software Used in Coordinate Parameters and Topologies | Software Used in Construction of Cubic Boxes * | Simulation Stage Parameters | Software Used to Analyze MD Results | ||
| 1st | 2nd | ||||||
| OPLS-AA [53] | AnteChamber PYthon Parser InterfacE (AcPype) [57] | GROMACS 5.1.4 [54] | t = 500 ps T = 310 K w/ position restriction | t = 100.000 ps T = 310 K | Visual molecular dynamics (VMD) [56] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, D.R.; Souza, F.R.; Guimarães, A.P.; Valis, M.; Pavelek, Z.; Kuca, K.; Ramalho, T.C.; França, T.C.C. In Silico Studies of Potential Selective Inhibitors of Thymidylate Kinase from Variola virus. Pharmaceuticals 2021, 14, 1027. https://doi.org/10.3390/ph14101027
Garcia DR, Souza FR, Guimarães AP, Valis M, Pavelek Z, Kuca K, Ramalho TC, França TCC. In Silico Studies of Potential Selective Inhibitors of Thymidylate Kinase from Variola virus. Pharmaceuticals. 2021; 14(10):1027. https://doi.org/10.3390/ph14101027
Chicago/Turabian StyleGarcia, Danielle R., Felipe R. Souza, Ana P. Guimarães, Martin Valis, Zbyšek Pavelek, Kamil Kuca, Teodorico C. Ramalho, and Tanos C. C. França. 2021. "In Silico Studies of Potential Selective Inhibitors of Thymidylate Kinase from Variola virus" Pharmaceuticals 14, no. 10: 1027. https://doi.org/10.3390/ph14101027
APA StyleGarcia, D. R., Souza, F. R., Guimarães, A. P., Valis, M., Pavelek, Z., Kuca, K., Ramalho, T. C., & França, T. C. C. (2021). In Silico Studies of Potential Selective Inhibitors of Thymidylate Kinase from Variola virus. Pharmaceuticals, 14(10), 1027. https://doi.org/10.3390/ph14101027