Overcoming Hypoxia-Induced Chemoresistance in Cancer Using a Novel Glycoconjugate of Methotrexate

 , , , and
, , , and
Abstract
1. Introduction
2. Results
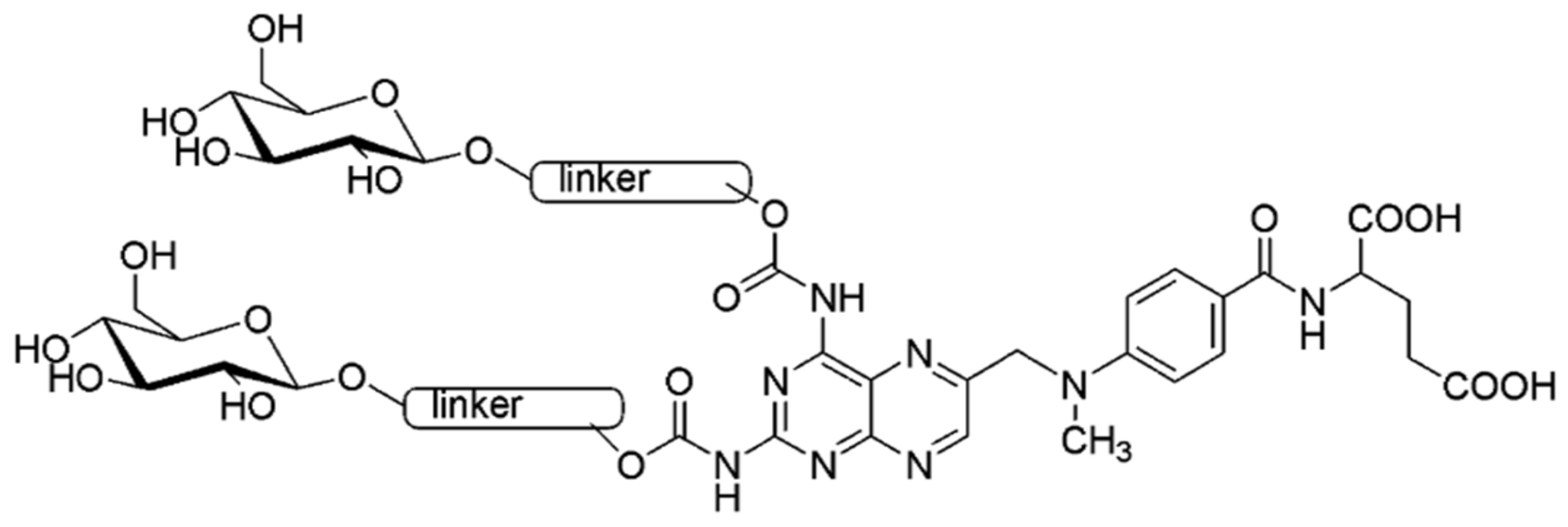
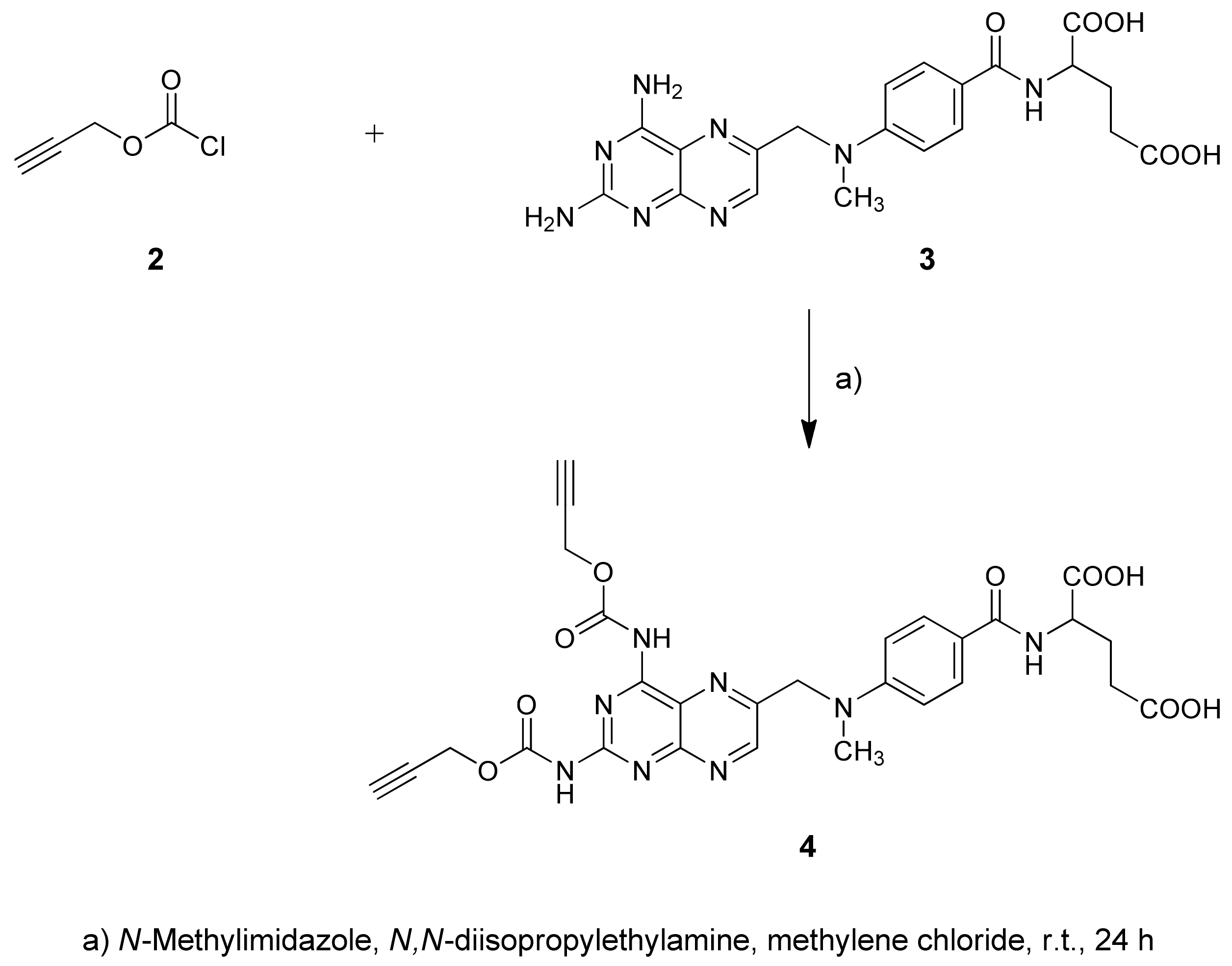
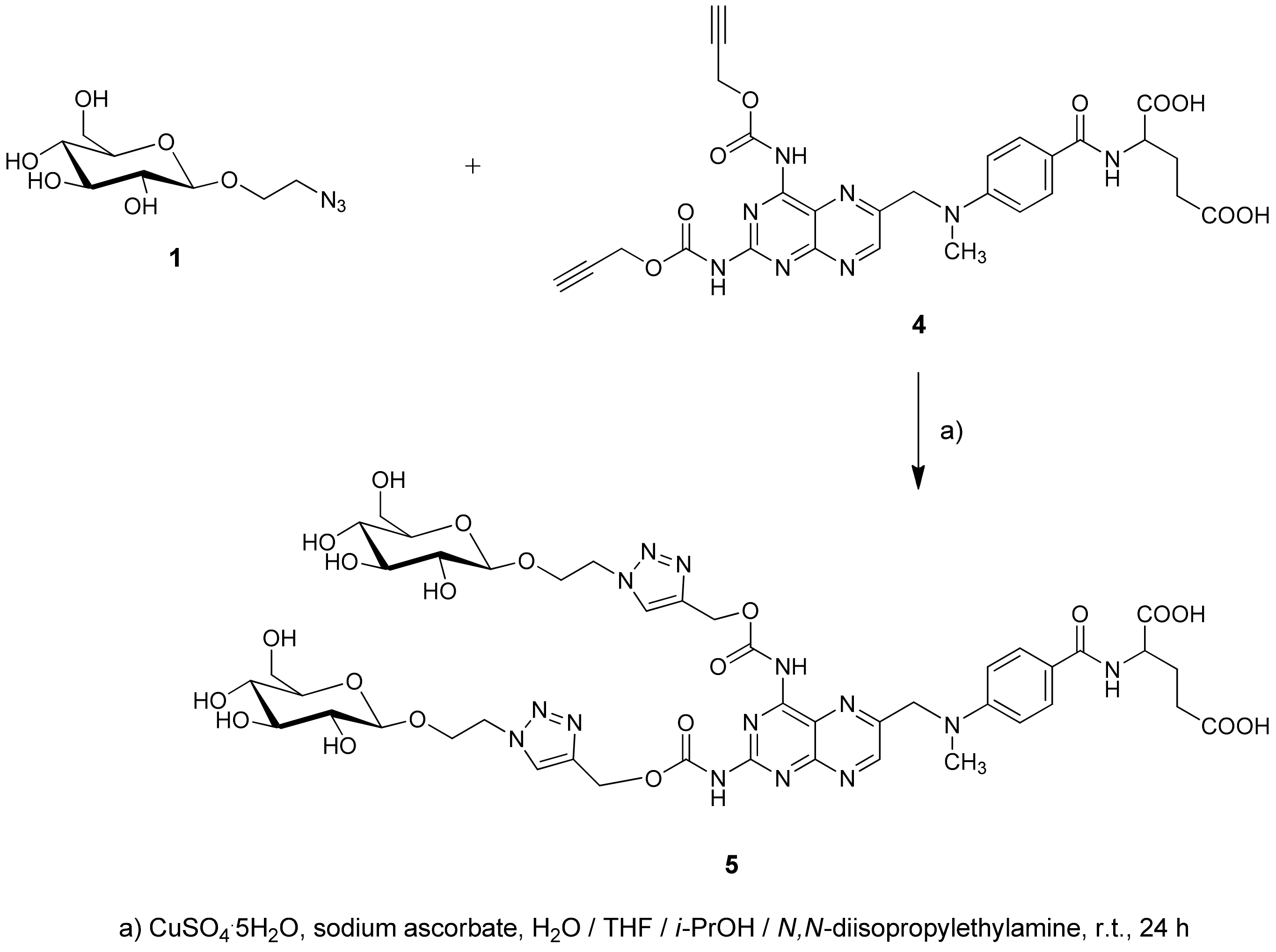
2.1. Chemistry
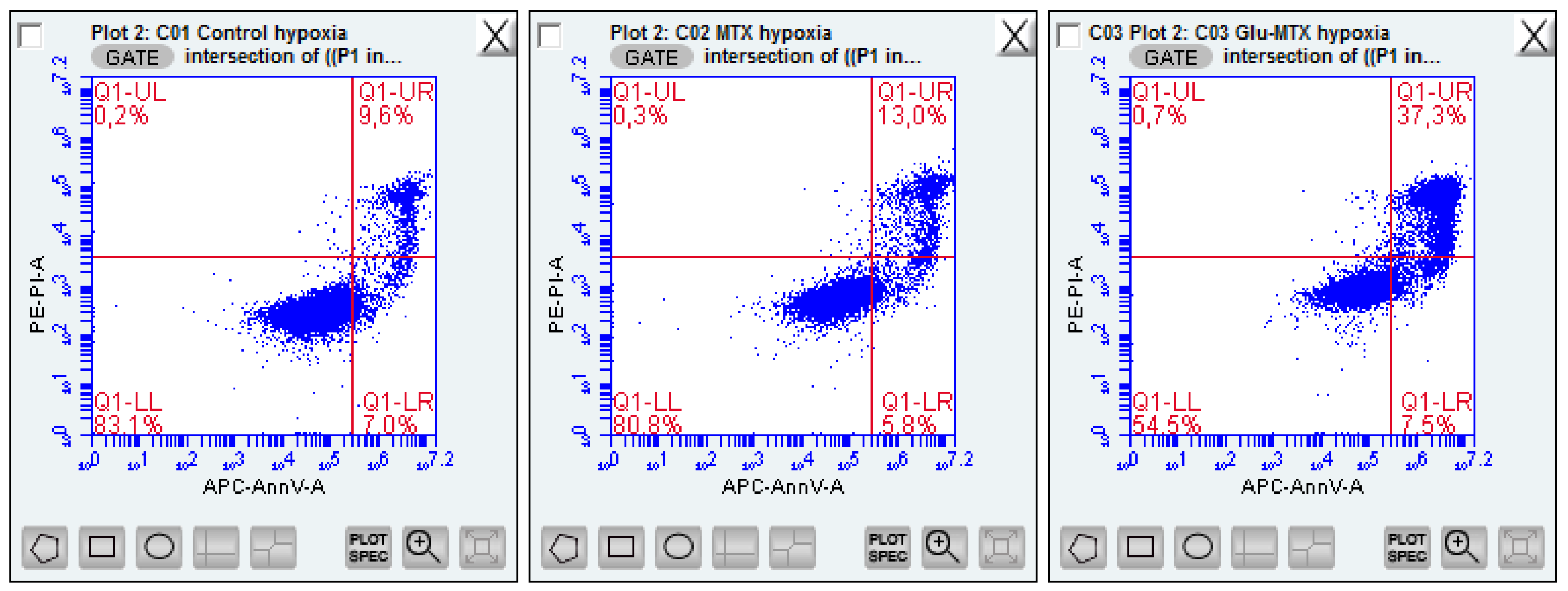
2.2. Biology Experiments
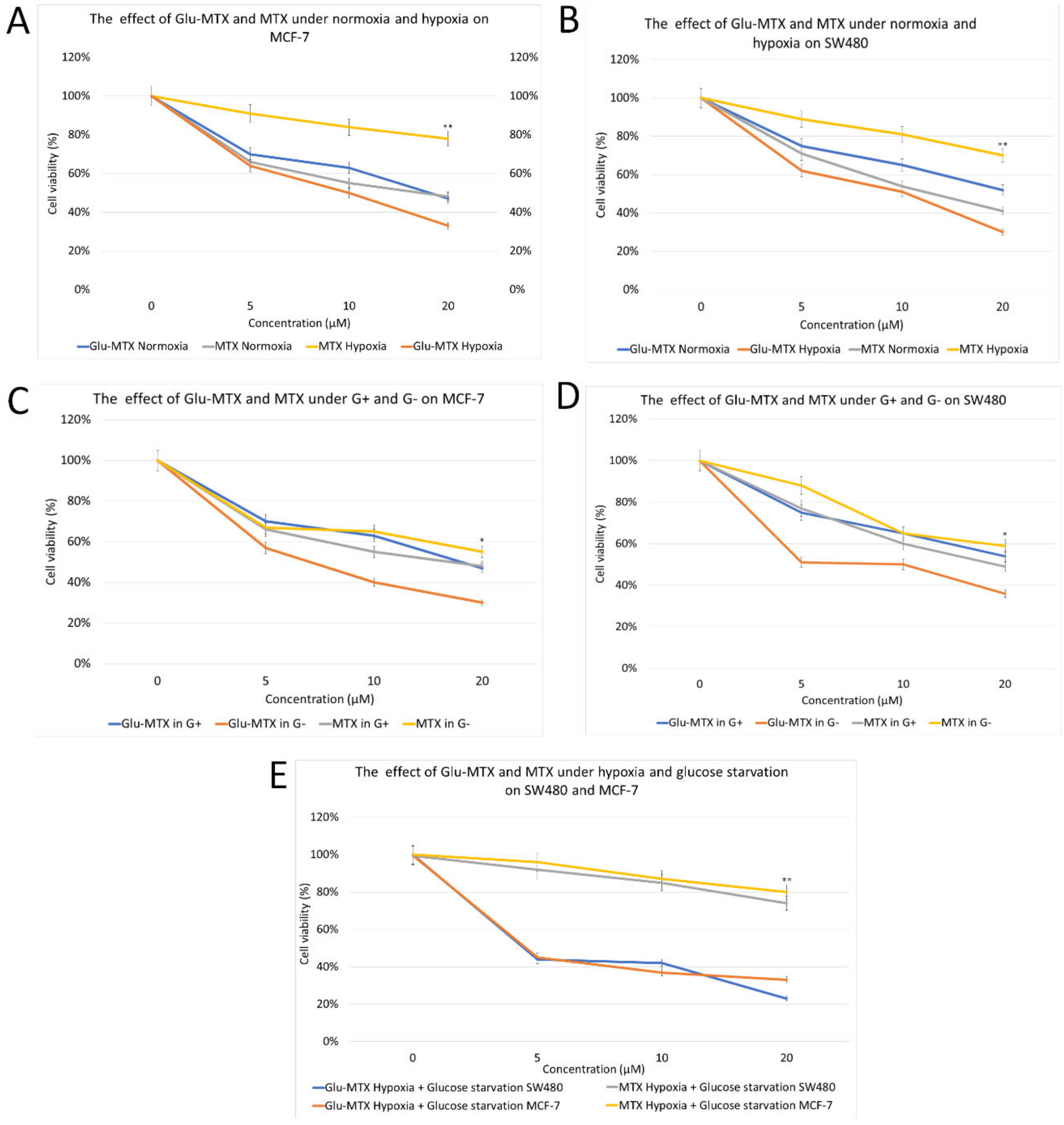
2.2.1. Glu-MTX Compared to MTX Exerted More Potent Cytotoxic Activity on MCF-7 and SW480 Cancer Cell Lines in Hypoxia and Glucose-Deprived Microenvironment
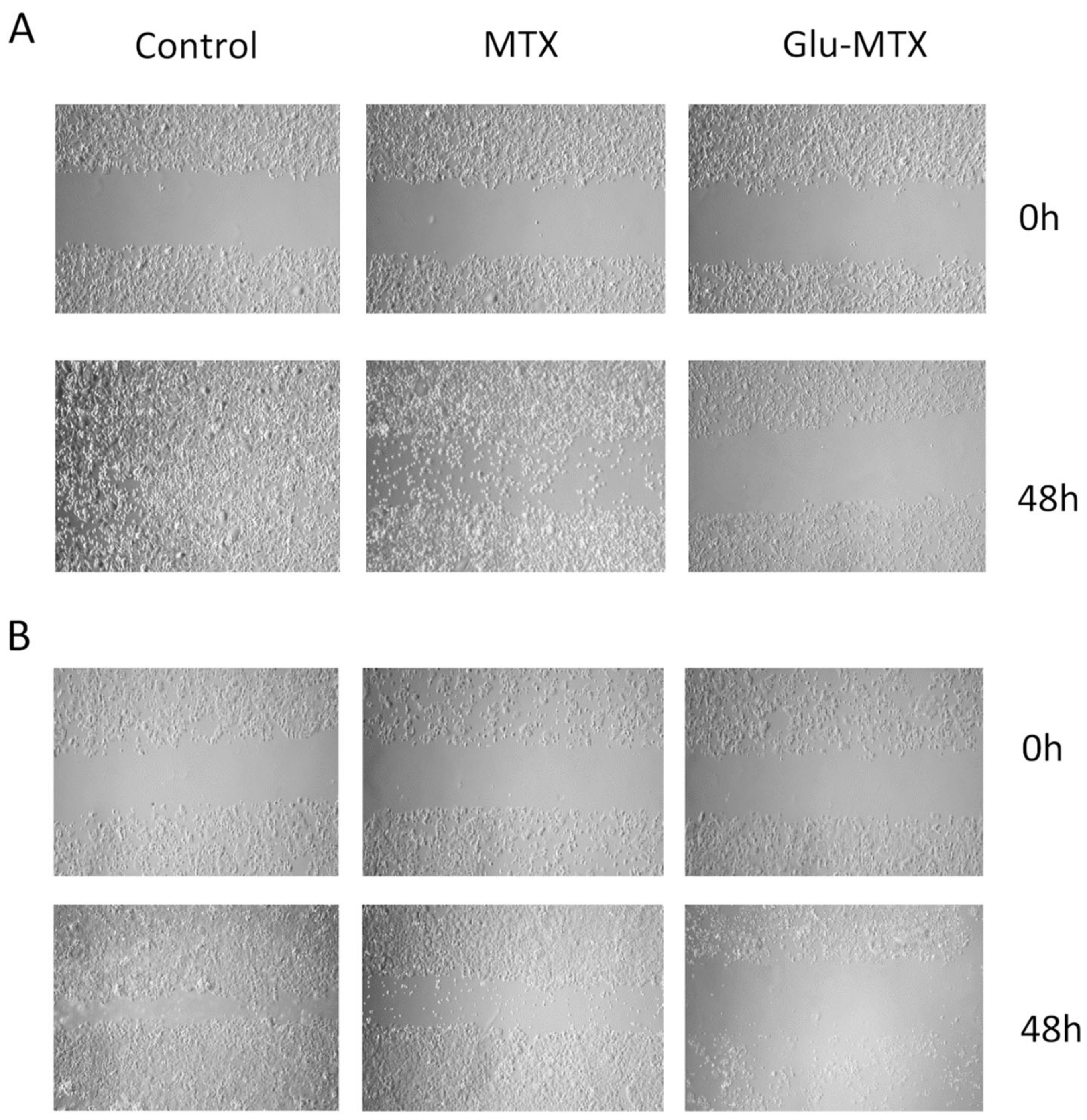
2.2.2. Glu-MTX Inhibits the Wound Healing Process
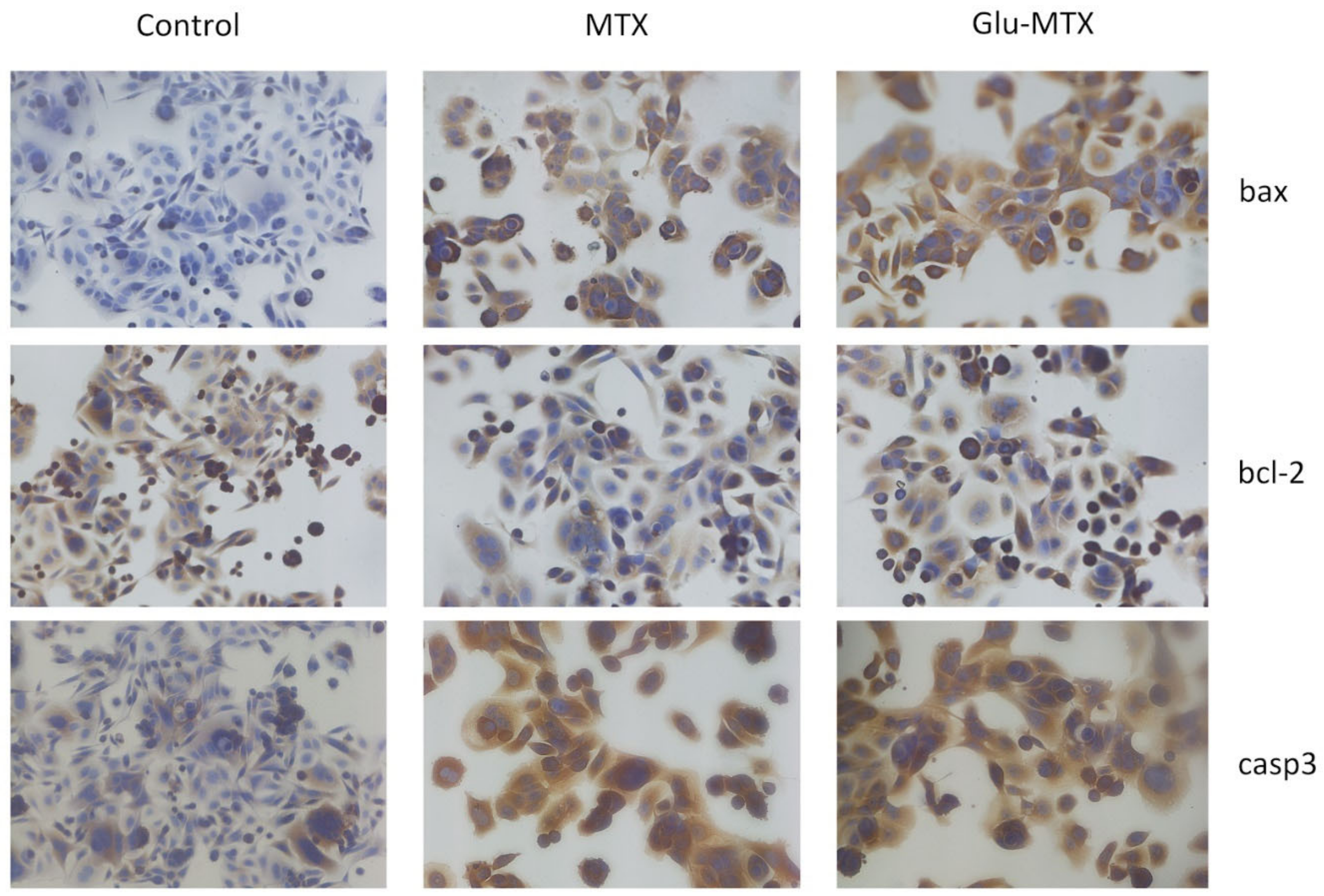
2.2.3. Glu-MTX Induces Apoptosis by Increasing the Expression of Caspase-3 and Bax
3. Discussion
4. Materials and Methods
4.1. Chemistry
4.1.1. Synthesis of Carbamate 4
4.1.2. Synthesis of Glycoconjugate 5
4.1.3. LC/MS Analysis
4.2. Biology
4.2.1. Cell Culture
4.2.2. Experiment Conditions and Cell Viability MTT Assay
4.2.3. Wound-Healing Assay
4.2.4. Flow Cytometry-Apoptosis Assay
4.2.5. ICC Staining for Apoptosis Detection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Roma-Rodrigues, C.; Mendes, R.; Baptista, P.; Fernandes, A. Targeting tumor microenvironment for cancer therapy. Int. J. Mol. Sci. 2019, 20, 840. [Google Scholar] [CrossRef] [PubMed]
- Trédan, O.; Galmarini, C.M.; Patel, K.; Tannock, I.F. Drug resistance and the solid tumor microenvironment. J. Natl. Cancer Inst. 2007, 99, 1441–1454. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.L.; Bissell, M.J. The tumor microenvironment is a dominant force in multidrug resistance. Drug Resist. Updat. 2012, 15, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Nagao, A.; Kobayashi, M.; Koyasu, S.; Chow, C.C.T.; Harada, H. HIF-1-Dependent reprogramming of glucose metabolic pathway of cancer cells and its therapeutic significance. Int. J. Mol. Sci. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, M.; Makuch, S.; Winograd, K.; Wiśniewski, J.; Ziółkowski, P.; Agrawal, S. 6-Shogaol enhances the anticancer effect of 5-fluorouracil, oxaliplatin, and irinotecan via increase of apoptosis and autophagy in colon cancer cells in hypoxic/aglycemic conditions. BMC Complement. Med. Ther. 2020, 20, 141. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Maeda, T.; Suzuki, A.; Baba, Y. Cancer metabolism: New insights into classic characteristics. Jpn. Dent. Sci. Rev. 2018, 54, 8–21. [Google Scholar] [CrossRef]
- Gocheva, G.; Ivanova, A. A look at receptor ligand pairs for active-targeting drug delivery from crystallographic and molecular dynamics perspectives. Mol. Pharm. 2019, 16, 3293–3321. [Google Scholar] [CrossRef]
- Zhu, Y.; Feijen, J.; Zhong, Z. Dual-targeted nanomedicines for enhanced tumor treatment. Nano Today 2018, 18, 65–85. [Google Scholar] [CrossRef]
- Minchinton, A.I.; Tannock, I.F. Drug penetration in solid tumours. Nat. Rev. Cancer 2006, 6, 583–592. [Google Scholar] [CrossRef]
- Srinivasarao, M.; Low, P.S. LiganD-targeted drug delivery. Chem. Rev. 2017, 117, 12133–12164. [Google Scholar] [CrossRef]
- Cantor, J.R.; Sabatini, D.M. Cancer cell metabolism: One hallmark, many faces. Cancer Discov. 2012, 2, 881–898. [Google Scholar] [CrossRef] [PubMed]
- Series, C.; Review, L. F-18 FDG PET/CT for detection of malignant involvement of peripheral nerves. Clin. Nucl. Med. 2011, 36, 96–100. [Google Scholar]
- Heiden, M.G. Vander targeting cancer metabolism: A therapeutic window opens. Nat. Rev. Drug Discov. 2011, 10, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Calvaresi, E.C.; Hergenrother, P.J. Glucose conjugation for the specific targeting and treatment of cancer. Chem. Sci. 2013, 4, 2319. [Google Scholar] [CrossRef]
- Srinivasarao, M.; Galliford, C.V.; Low, P.S. Principles in the design of liganD-targeted cancer therapeutics and imaging agents. Nat. Rev. Drug Discov. 2015, 14, 203–219. [Google Scholar] [CrossRef]
- Abolmaali, S.S.; Tamaddon, A.M.; Dinarvand, R. A review of therapeutic challenges and achievements of methotrexate delivery systems for treatment of cancer and rheumatoid arthritis. Cancer Chemother. Pharmacol. 2013, 71, 1115–1130. [Google Scholar] [CrossRef]
- Kolb, H.C.; Finn, M.G.; Sharpless, K.B. Click chemistry: Diverse chemical function from a few good reactions. Angew. Chem. Int. Ed. 2001, 40, 2004–2021. [Google Scholar] [CrossRef]
- Thirumurugan, P.; Matosiuk, D.; Jozwiak, K. Click chemistry for drug development and diverse chemical-biology applications. Chem. Rev. 2013, 113, 4905–4979. [Google Scholar] [CrossRef]
- Pohl, J.; Bertram, B.; Hilgard, P.; Nowrousian, M.R.; Stüben, J.; Wießler, M. D-19575—A sugar-linked isophosphoramide mustard derivative exploiting transmembrane glucose transport. Cancer Chemother. Pharmacol. 1995, 35, 364–370. [Google Scholar] [CrossRef]
- Granchi, C.; Fortunato, S.; Minutolo, F. Anticancer agents interacting with membrane glucose transporters. Med. Chem. Commun. 2016, 7, 1716–1729. [Google Scholar] [CrossRef]
- Barnett, J.E.G.; Holman, G.D.; Munday, K.A. Structural requirements for binding to the sugar-transport system of the human erythrocyte. Biochem. J. 1973, 131, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.; Makepeace, C. Model of the exofacial substrate-binding site and helical folding of the human Glut1 glucose transporter based on scanning mutagenesis. Biochemistry 2009, 48, 5934–5942. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Bertozzi, C.R. Site-specific antibody-drug conjugates: The nexus of bioorthogonal chemistry, protein engineering, and drug development. Bioconjug. Chem. 2015, 26, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.K.; Mishra, B.B.; Mishra, K.B.; Mishra, N.; Singh, A.S.; Chen, X. Cu-catalyzed click reaction in carbohydrate chemistry. Chem. Rev. 2016, 116, 3086–3240. [Google Scholar] [CrossRef]
- Krawczyk, M.; Pastuch-Gawołek, G.; Pluta, A.; Erfurt, K.; Domiński, A.; Kurcok, P. 8-Hydroxyquinoline glycoconjugates: Modifications in the linker structure and their effect on the cytotoxicity of the obtained compounds. Molecules 2019, 24, 4181. [Google Scholar] [CrossRef]
- Le Roux, A.; Meunier, S.; Le Gall, T.; Denis, J.M.; Bischoff, P.; Wagner, A. Synthesis and radioprotective properties of pulvinic acid derivatives. Chem. Med. Chem. 2011, 6, 561–569. [Google Scholar] [CrossRef]
- Zemplén, G.; Pacsu, E. Über die Verseifung acetylierter Zucker und verwandter Substanzen. Ber. Dtsch. Chem. Ges. 1929, 62, 1613–1614. [Google Scholar] [CrossRef]
- De Figueiredo, R.M.; Suppo, J.S.; Campagne, J.M. Nonclassical routes for amide bond formation. Chem. Rev. 2016, 116, 12029–12122. [Google Scholar] [CrossRef]
- Kim, S.; Lee, J.I.; Kim, Y.C. A simple and mild esterification method for carboxylic acids using mixed carboxylic-carbonic anhydrides. J. Org. Chem. 1985, 50, 561–565. [Google Scholar] [CrossRef]
- Watkins, B.E.; Kiely, J.S.; Rapoport, H. Synthesis of oligodeoxyribonucleotides using N-benzyloxycarbonyl-blocked nucleosides. J. Am. Chem. Soc. 1982, 104, 5702–5708. [Google Scholar] [CrossRef]
- Pandey, M.; Prasad, S.; Tyagi, A.; Deb, L.; Huang, J.; Karelia, D.; Amin, S.; Aggarwal, B. Targeting cell survival proteins for cancer cell death. Pharmaceuticals 2016, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, J.; Zhang, L.; Wei, F.; Lian, Y.; Wu, Y.; Gong, Z.; Zhang, S.; Zhou, J.; Cao, K.; et al. Role of tumor microenvironment in tumorigenesis. J. Cancer 2017, 8, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Doktorova, H.; Hrabeta, J.; Khalil, M.A.; Eckschlager, T. Hypoxia-induced chemoresistance in cancer cells: The role of not only HIF-1. Biomed. Pap. 2015, 159, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, M.; Cao, Y.; Yuan, W.; Pang, T.; Li, B.; Sun, Z.; Chen, L.; Zhao, R.C. Knockdown of hypoxia-inducible factor-1α in breast carcinoma MCF-7 cells results in reduced tumor growth and increased sensitivity to methotrexate. Biochem. Biophys. Res. Commun. 2006, 342, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Sanna, K.; Rofstad, E.K. Hypoxia-induced resistance to doxorubicin and methotrexate in human melanoma cell lines in vitro. Int. J. Cancer 1994, 58, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Petit, C.; Gouel, F.; Dubus, I.; Heuclin, C.; Roget, K.; Vannier, J.P. Hypoxia promotes chemoresistance in acute lymphoblastic leukemia cell lines by modulating death signaling pathways. BMC Cancer 2016, 16, 746. [Google Scholar] [CrossRef]
- Nishimoto, A.; Kugimiya, N.; Hosoyama, T.; Enoki, T.; Li, T.-S.; Hamano, K. HIF-1 activation under glucose deprivation plays a central role in the acquisition of anti-apoptosis in human colon cancer cells. Int. J. Oncol. 2014, 44, 2077–2084. [Google Scholar] [CrossRef]
- Mathews, E.H.; Stander, B.A.; Joubert, A.M.; Liebenberg, L. Tumor cell culture survival following glucose and glutamine deprivation at typical physiological concentrations. Nutrition 2014, 30, 218–227. [Google Scholar] [CrossRef]
- Hu, Y.L.; Yin, Y.; Liu, H.Y.; Feng, Y.Y.; Bian, Z.H.; Zhou, L.Y.; Zhang, J.W.; Fei, B.J.; Wang, Y.G.; Huang, Z.H. Glucose deprivation induces chemoresistance in colorectal cancer cells by increasing ATF4 expression. World J. Gastroenterol. 2016, 22, 6235–6245. [Google Scholar] [CrossRef]
- Fu, J.; Yang, J.; Seeberger, P.H.; Yin, J. Glycoconjugates for glucose transporter-mediated cancer-specific targeting and treatment. Carbohydr. Res. 2020, 498, 108195. [Google Scholar] [CrossRef]
- Deng, D.; Yan, N. GLUT, SGLT, and SWEET: Structural and mechanistic investigations of the glucose transporters. Protein Sci. 2016, 25, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Barron, C.C.; Bilan, P.J.; Tsakiridis, T.; Tsiani, E. Facilitative glucose transporters: Implications for cancer detection, prognosis and treatment. Metabolism 2016, 65, 124–139. [Google Scholar] [CrossRef]
- Parks, S.K.; Cormerais, Y.; Marchiq, I.; Pouyssegur, J. Hypoxia optimises tumour growth by controlling nutrient import and acidic metabolite export. Mol. Aspects Med. 2016, 47, 3–14. [Google Scholar] [CrossRef]
- Yang, Q.R.; Qiao, W.H.; Zhang, S.M.; Qu, J.P.; Liu, D.L. Synthesis and characterization of a new cationic galactolipid with carbamate for gene delivery. Tenside Surf. Det. 2010, 5, 294–299. [Google Scholar] [CrossRef]
- Bhattacharya, B.; Omar, M.F.M.; Soong, R. The Warburg effect and drug resistance. Br. J. Pharmacol. 2016, 173, 970–979. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Procedure | Substrate 1 | Substrate 2 | Reagents | Solvent | Reaction Time [h] | Yield [%] |
|---|---|---|---|---|---|---|
| A | MTX | Propiolic acid | DCC | DMF | 48 | traces |
| DCC/DMAP | DMF | 12 | ||||
| DCC/DMAP | DMF/CH2Cl2 | 18 | ||||
| B | MTX | Propargyl chloroformate | Pyridine | DMF/CH2Cl2 | 24 | 22 |
| Triethylamine | inseparable reaction mixture | |||||
| DMAP | ||||||
| Imidazole | ||||||
| C | MTX | Propargyl chloroformate | N-Methylimidazole | CHCl3 | 24 | - |
| THF | 8 | |||||
| CH3CN | 4 | |||||
| DMF/CH2Cl2 | 21 | |||||
| CH2Cl2 | 32 | |||||
| NMI/Hünigs base | CH2Cl2 | 42 |
| Procedure | Substrate 1 | Substrate 2 | Catalyst | Solvent | Reaction Time [h] | Yield [%] |
|---|---|---|---|---|---|---|
| A | 1 | 4 | NaAsc/CuSO4·5H2O | THF/i-PrOH/H2O | 24 | 11 |
| B | 1 | 4 | NaAsc/CuSO4·5H2O | THF/i-PrOH/H2O NaOH | 24 | 43 |
| C | 1 | 4 | NaAsc/CuSO4·5H2O | THF/i-PrOH/H2O Hünig’s base | 24 | 77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woźniak, M.; Pastuch-Gawołek, G.; Makuch, S.; Wiśniewski, J.; Ziółkowski, P.; Szeja, W.; Krawczyk, M.; Agrawal, S. Overcoming Hypoxia-Induced Chemoresistance in Cancer Using a Novel Glycoconjugate of Methotrexate. Pharmaceuticals 2021, 14, 13. https://doi.org/10.3390/ph14010013
Woźniak M, Pastuch-Gawołek G, Makuch S, Wiśniewski J, Ziółkowski P, Szeja W, Krawczyk M, Agrawal S. Overcoming Hypoxia-Induced Chemoresistance in Cancer Using a Novel Glycoconjugate of Methotrexate. Pharmaceuticals. 2021; 14(1):13. https://doi.org/10.3390/ph14010013
Chicago/Turabian StyleWoźniak, Marta, Gabriela Pastuch-Gawołek, Sebastian Makuch, Jerzy Wiśniewski, Piotr Ziółkowski, Wiesław Szeja, Monika Krawczyk, and Siddarth Agrawal. 2021. "Overcoming Hypoxia-Induced Chemoresistance in Cancer Using a Novel Glycoconjugate of Methotrexate" Pharmaceuticals 14, no. 1: 13. https://doi.org/10.3390/ph14010013
APA StyleWoźniak, M., Pastuch-Gawołek, G., Makuch, S., Wiśniewski, J., Ziółkowski, P., Szeja, W., Krawczyk, M., & Agrawal, S. (2021). Overcoming Hypoxia-Induced Chemoresistance in Cancer Using a Novel Glycoconjugate of Methotrexate. Pharmaceuticals, 14(1), 13. https://doi.org/10.3390/ph14010013