Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects

 , ,
, ,  ,
,  and
and 
Abstract
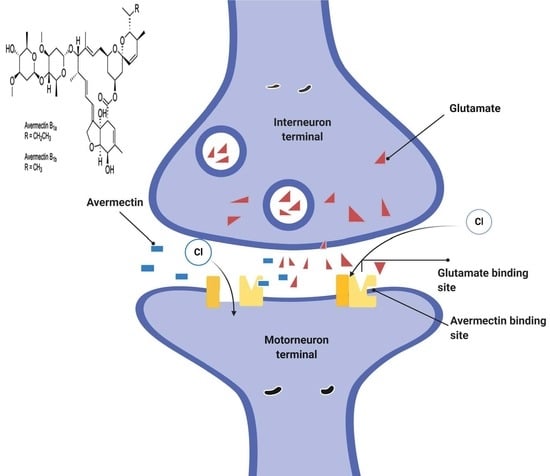
1. Introduction
2. History
3. Structure and Chemical Properties
4. Avermectin Biosynthesis
5. Avermectin Derivatives, Formulations, Therapeutic and Toxic Dosages, and Mechanism of Action
5.1. Avermectin Derivatives and Its Formulations
5.1.1. Ivermectin
5.1.2. Moxidectin
5.1.3. Selamectin
5.1.4. Abamectin (Avermectin B1)
5.1.5. Milbemycin
5.1.6. Doramectin, Eprinomectin, and Nemadectin
5.2. Dosing
5.2.1. Mechanism of Action
5.2.2. Mode of Action of Milbemycins
5.2.3. Mode of Action of Spinosyns
6. Pharmacological Effects of Ivermectin
6.1. Human Uses of Ivermectin
6.2. Efficacy against Plant Parasitic Nematodes
6.3. Antibacterial Action
6.4. Endectocides for Malaria Control
6.5. Anti-Inflammatory Effect of Avermectin
6.6. Anticancer
6.7. Metabolic Effect of IVM
6.8. Anti-Alcohol Therapies
6.9. Antitumor Effect of Avermectins
6.10. Antiviral Effects
6.11. Ivermectin and Coronavirus
6.12. Metabolic Effects of Ivermectin
7. Pharmacokinetics
8. Environmental Effects of the Usage of Avermectins in Livestock
9. Clinical Trials of Avermectins
10. Toxicity and Side Effects
10.1. Neurotoxicology of Ivermectin
10.2. Toxic Effect of Ivermectin
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pitterna, T.; Cassayre, J.; Hüter, O.F.; Jung, P.M.; Maienfisch, P.; Kessabi, F.M.; Quaranta, L.; Tobler, H. New ventures in the chemistry of avermectins. Bioorg. Med. Chem. 2009, 17, 4085–4095. [Google Scholar] [CrossRef]
- Jayakumar, J. Bio-efficacy of Streptomyces avermitilis culture filtrates against root knot nematode, Meloidogyne incognita and reniform nematodes, Rotylenchulus reniformis. Karnataka J. Agric. Sci. 2009, 22, 567–571. [Google Scholar]
- Campbell, W.C. Ivermectin and Abamectin; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Siddique, S.; Syed, Q.; Adnan, A.; Qureshi, F.A. Isolation, characterization and selection of avermectin-producing Streptomyces avermitilis strains from soil samples. Jundishapur J. Microbiol. 2014, 7. [Google Scholar] [CrossRef]
- Prichard, R.; Ménez, C.; Lespine, A. Moxidectin and the avermectins: Consanguinity but not identity. Int. J. Parasitol. 2012, 2, 134–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Panda, S.K.; Luyten, W. Plant-Based Natural Products for the Discovery and Development of Novel Anthelmintics against Nematodes. Biomolecules 2020, 10, 426. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Matsumoto, A.; Seino, A.; Ueno, J.; Iwai, Y.; Omura, S. Streptomyces avermectinius sp. nov., an avermectin-producing strain. Int. J. Syst. Evol. Microbiol. 2002, 52, 2163–2168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yan, Y.-J.; An, J.; Huang, S.-X.; Wang, X.-J.; Xiang, W.-S. Designed biosynthesis of 25-methyl and 25-ethyl ivermectin with enhanced insecticidal activity by domain swap of avermectin polyketide synthase. Microb. Cell Factories 2015, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lumaret, J.-P.; Errouissi, F.; Floate, K.; Rombke, J.; Wardhaugh, K. A review on the toxicity and non-target effects of macrocyclic lactones in terrestrial and aquatic environments. Curr. Pharm. Biotechnol. 2012, 13, 1004–1060. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Beshbishy, A.M.; Tayebwa, D.S.; Adeyemi, O.S.; Yokoyama, N.; Igarashi, I. Evaluation of the inhibitory effect of ivermectin on the growth of Babesia and Theileria parasites in vitro and in vivo. Trop. Med. Health 2019, 47, 42. [Google Scholar] [CrossRef]
- Beshbishy, A.M.; Batiha, G.E.-S.; Yokoyama, N.; Igarashi, I. Ellagic acid microspheres restrict the growth of Babesia and Theileria in vitro and Babesia microti in vivo. Parasites Vectors 2019, 12, 269. [Google Scholar] [CrossRef]
- Pope, L.J. Fate and Effects of Parasiticides in the Pasture Environment; University of York: York, UK, 2009. [Google Scholar]
- Prasse, C.; Löffler, D.; Ternes, T.A. Environmental fate of the anthelmintic ivermectin in an aerobic sediment/water system. Chemosphere 2009, 77, 1321–1325. [Google Scholar] [CrossRef] [PubMed]
- Barrón-Bravo, O.G.; Hernández-Marín, J.A.; Gutiérrez-Chávez, A.J.; Franco-Robles, E.; Molina-Ochoa, J.; Cruz-Vázquez, C.R.; Ángel-Sahagún, C.A. Susceptibility of entomopathogenic nematodes to ivermectin and thiabendazole. Chemosphere 2020, 253, 126658. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, L.E.; Saumell, C.; Sagüés, F.; Sallovitz, J.M.; Lifschitz, A.L. Ivermectin dissipation and movement from feces to soil under field conditions. J. Environ. Sci. Health Part B 2018, 53, 42–48. [Google Scholar] [CrossRef]
- Yoon, Y.; Kim, E.-S.; Hwang, Y.-S.; Choi, C.-Y. Avermectin: Biochemical and molecular basis of its biosynthesis and regulation. Appl. Microbiol. Biotechnol. 2004, 63, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.-A.; Han, S.; Lim, Y.-R.; Kim, V.; Kim, H.; Kim, D. Streptomyces cytochrome P450 enzymes and their roles in the biosynthesis of macrolide therapeutic agents. Biomol. Ther. 2019, 27, 127. [Google Scholar] [CrossRef] [PubMed]
- DeMars, M., II. Structure and Biochemistry of Cytochromes P450 Involved in the Biosynthesis of Macrolide Antibiotics. Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 2017. [Google Scholar]
- Schulman, M.; Acton, S.; Valentino, D.L.; Arison, B.H. Purification and identification of dTDP-oleandrose, the precursor of the oleandrose units of the avermectins. J. Biol. Chem. 1990, 265, 16965–16970. [Google Scholar]
- Plumb, D.C. Plumb’s Veterinary Drug Handbook: Desk; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Pérez, R.; Cabezas, I.; Sutra, J.; Galtier, P.; Alvinerie, M. Faecal excretion profile of moxidectin and ivermectin after oral administration in horses. Vet. J. 2001, 161, 85–92. [Google Scholar] [CrossRef]
- Merola, V.M.; Eubig, P.A. Toxicology of avermectins and milbemycins (macrocylic lactones) and the role of P-glycoprotein in dogs and cats. Vet. Clin. Small Anim. Pract. 2012, 42, 313–333. [Google Scholar] [CrossRef]
- Merola, V.A.; Khan, S.; Gwaltney-Brant, S. Ivermectin toxicosis in dogs: A retrospective study. J. Am. Anim. Hosp. Assoc. 2009, 45, 106–111. [Google Scholar] [CrossRef]
- Hopkins, K.; Marcella, K.; Strecker, A. Ivermectin toxicosis in a dog. J. Am. Vet. Med. Assoc. 1990, 197, 93–94. [Google Scholar]
- Mealey, K.L.; Meurs, K.M. Breed distribution of the ABCB1-1Δ (multidrug sensitivity) polymorphism among dogs undergoing ABCB1 genotyping. J. Am. Vet. Med. Assoc. 2008, 233, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.J.; Vernau, K.M.; Maggs, D.J. Retinopathy associated with ivermectin toxicosis in two dogs. J. Am. Vet. Med. Assoc. 2008, 233, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Bates, N.; Chatterton, J.; Robbins, C.; Wells, K.; Hughes, J.; Stone, M.; Campbell, A. Lipid infusion in the management of poisoning: A report of 6 canine cases. Vet. Rec. Case Rep. 2013, 1. [Google Scholar] [CrossRef]
- Laing, R.; Gillan, V.; Devaney, E. Ivermectin–old drug, new tricks? Trends Parasitol. 2017, 33, 463–472. [Google Scholar] [CrossRef] [PubMed]
- See, A.; McGill, S.; Raisis, A.; Swindells, K. Toxicity in three dogs from accidental oral administration of a topical endectocide containing moxidectin and imidacloprid. Aust. Vet. J. 2009, 87, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Bishop, B.; Bruce, C.; Evans, N.; Goudie, A.; Gration, K.; Gibson, S.; Pacey, M.; Perry, D.; Walshe, N.; Witty, M. Selamectin: A novel broad-spectrum endectocide for dogs and cats. Vet. Parasitol. 2000, 91, 163–176. [Google Scholar] [CrossRef]
- Krautmann, M.; Novotny, M.; De Keulenaer, K.; Godin, C.; Evans, E.; McCall, J.; Wang, C.; Rowan, T.; Jernigan, A. Safety of selamectin in cats. Vet. Parasitol. 2000, 91, 393–403. [Google Scholar] [CrossRef]
- Khalil, M.S. Abamectin and azadirachtin as eco-friendly promising biorational tools in integrated nematodes management programs. J. Plant Pathol. Microbiol. 2013, 4, 2–3. [Google Scholar] [CrossRef]
- Nasr, H.M.; El-Demerdash, F.M.; El-Nagar, W.A. Neuro and renal toxicity induced by chlorpyrifos and abamectin in rats. Environ. Sci. Pollut. Res. 2016, 23, 1852–1859. [Google Scholar] [CrossRef]
- Mingot, J.M.; Kostka, S.; Kraft, R.; Hartmann, E.; Görlich, D. Importin 13: A novel mediator of nuclear import and export. EMBO J. 2001, 20, 3685–3694. [Google Scholar] [CrossRef]
- Lanusse, C.E.; Imperiale, F.A.; Lifschitz, A.L. Macrocyclic Lactones: Endectocide Compounds; Wiley-Blackwell: Hoboken, NJ, USA, 2018. [Google Scholar]
- Kozan, E.; Sevimli, F.K.; Birdane, F.M.; Adanır, R. Efficacy of eprinomectin against Toxacara canis in dogs. Parasitol. Res. 2008, 102, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.; Wang, X.; Xiang, W.; Liang, H.; Gao, J.; Yan, Y. Reversal of P-glycoprotein-mediated multidrug resistance in vitro by doramectin and nemadectin. J. Pharm. Pharmacol. 2010, 62, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Geyer, J.; Klintzsch, S.; Meerkamp, K.; Wöhlke, A.; Distl, O.; Moritz, A.; Petzinger, E. Detection of the nt230 (del4) MDR1 mutation in White Swiss Shepherd dogs: Case reports of doramectin toxicosis, breed predisposition, and microsatellite analysis. J. Vet. Pharmacol. Ther. 2007, 30, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Hotson, I. The avermectins: A new family of antiparasitic agents. J. S. Afr. Vet. Assoc. 1982, 53, 87–90. [Google Scholar] [PubMed]
- Bazzano, M.; Di Salvo, A.; Diaferia, M.; Veronesi, F.; Galarini, R.; Paoletti, F.; Tesei, B.; McLean, A.; Veneziano, V.; Laus, F. Anthelmintic efficacy and pharmacokinetics of ivermectin paste after ooral administration in mules infected by Cyathostomins. Animals 2020, 10, 934. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R.; Greene, B.M. The status of ivermectin in the treatment of human onchocerciasis. Am. J. Trop. Med. Hyg. 1989, 41, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Juarez, M.; Schcolnik-Cabrera, A.; Dueñas-Gonzalez, A. The multitargeted drug ivermectin: From an antiparasitic agent to a repositioned cancer drug. Am. J. Cancer Res. 2018, 8, 317. [Google Scholar]
- Sharmeen, S.; Skrtic, M.; Sukhai, M.A.; Hurren, R.; Gronda, M.; Wang, X.; Fonseca, S.B.; Sun, H.; Wood, T.E.; Ward, R. The antiparasitic agent ivermectin induces chloride-dependent membrane hyperpolarization and cell death in leukemia cells. Blood J. Am. Soc. Hematol. 2010, 116, 3593–3603. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, S.; Sun, Q.; Liu, B. Anthelmintic drug ivermectin inhibits angiogenesis, growth and survival of glioblastoma through inducing mitochondrial dysfunction and oxidative stress. Biochem. Biophys. Res. Commun. 2016, 480, 415–421. [Google Scholar] [CrossRef]
- Dou, Q.; Chen, H.-N.; Wang, K.; Yuan, K.; Lei, Y.; Li, K.; Lan, J.; Chen, Y.; Huang, Z.; Xie, N. Ivermectin induces cytostatic autophagy by blocking the PAK1/Akt axis in breast cancer. Cancer Res. 2016, 76, 4457–4469. [Google Scholar] [CrossRef]
- Yin, J.; Park, G.; Lee, J.E.; Choi, E.Y.; Park, J.Y.; Kim, T.-H.; Park, N.; Jin, X.; Jung, J.-E.; Shin, D. DEAD-box RNA helicase DDX23 modulates glioma malignancy via elevating miR-21 biogenesis. Brain 2015, 138, 2553–2570. [Google Scholar] [CrossRef] [PubMed]
- Sharun, K.; Dhama, K.; Patel, S.K.; Pathak, M.; Tiwari, R.; Singh, B.R.; Sah, R.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J.; Leblebicioglu, H. Ivermectin, a New Candidate Therapeutic Against SARS-CoV-2/COVID-19; BioMed Central: London, UK, 2020. [Google Scholar]
- Patel, A.; Desai, S. Ivermectin in COVID-19 Related Critical Illness; SSRN—Social Science Research Network: 3570270; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Aziz, M.; Diop, I.; Diallo, S.; Lariviere, M.; Porta, M. Efficacy and tolerance of ivermectinin human onchocerciasis. Lancet 1982, 320, 171–173. [Google Scholar] [CrossRef]
- Bloomquist, J.R. Chloride channels as tools for developing selective insecticides. Arch. Insect Biochem. Physiol. Publ. Entomol. Soc. Am. 2003, 54, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Subbanna, A.; Stanley, J.; Rajasekhara, H.; Mishra, K.; Pattanayak, A.; Bhowmick, R. Perspectives of microbial metabolites as pesticides in agricultural pest management. Co-Evol. Second. Metab. 2020, 925–952. [Google Scholar] [CrossRef]
- Srivastava, P.K.; Singh, V.P.; Singh, A.; Singh, S.; Prasad, S.M.; Tripathi, D.K.; Chauhan, D.K. Pesticides in Crop Production; Wiley Online Library: Hoboken, NJ, USA, 2020. [Google Scholar]
- Dent, J.A.; Smith, M.M.; Vassilatis, D.K.; Avery, L. The genetics of ivermectin resistance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2000, 97, 2674–2679. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.G.; Prichard, R.K.; Beech, R.N. A glutamate-gated chloride channel subunit from Haemonchus contortus: Expression in a mammalian cell line, ligand binding, and modulation of anthelmintic binding by glutamate. Biochem. Pharmacol. 2002, 63, 1061–1068. [Google Scholar] [CrossRef]
- Arena, J.P.; Liu, K.K.; Paress, P.S.; Frazier, E.G.; Cully, D.F.; Mrozik, H.; Schaeffer, J.M. The mechanism of action of avermectins in Caenorhabditis elegans: Correlation between activation of glutamate-sensitive chloride current, membrane binding, and biological activity. J. Parasitol. 1995, 81, 286–294. [Google Scholar] [CrossRef]
- Fritz, L.C.; Wang, C.C.; Gorio, A. Avermectin B1a irreversibly blocks postsynaptic potentials at the lobster neuromuscular junction by reducing muscle membrane resistance. Proc. Natl. Acad. Sci. USA 1979, 76, 2062–2066. [Google Scholar] [CrossRef]
- Mrozik, H. Advances in Research and Development of Avermectins; ACS Publications: Washington, DC, USA, 1994. [Google Scholar]
- Fisher, M.H.; Mrozik, H. The chemistry and pharmacology of avermectins. Annu. Rev. Pharmacol. Toxicol. 1992, 32, 537–553. [Google Scholar] [CrossRef]
- Laing, S.T.; Ivens, A.; Butler, V.; Ravikumar, S.P.; Laing, R.; Woods, D.J.; Gilleard, J.S. The transcriptional response of Caenorhabditis elegans to ivermectin exposure identifies novel genes involved in the response to reduced food intake. PLoS ONE 2012, 7, e31367. [Google Scholar] [CrossRef]
- Arena, J.; Liu, K.; Paress, P.; Cully, D. Avermectin-sensitive chloride currents induced by Caenorhabditis elegans RNA in Xenopus oocytes. Mol. Pharmacol. 1991, 40, 368–374. [Google Scholar] [PubMed]
- Gürtler, L.; Bauerfeind, U.; Blümel, D.J.; Burger, R.; Drosten, C.; Gröner, A.; Heiden, M.; Hildebrandt, M.; Jansen, B.; Montag-Lessing, T. Arbonematodes–Nematode Infections Transmissible by Arthropods: Arbeitskreis Blut, Untergruppe «Bewertung Blutassoziierter Krankheitserreger». Transfus. Med. Hemother. 2013, 40, 50. [Google Scholar] [PubMed]
- Cobb, R.; Boeckh, A. Moxidectin: A review of chemistry, pharmacokinetics and use in horses. Parasites Vectors 2009, 2, S5. [Google Scholar] [CrossRef] [PubMed]
- Lifschitz, A.; Virkel, G.; Sallovitz, J.; Imperiale, F.; Pis, A.; Lanusse, C. Loperamide-induced enhancement of moxidectin availability in cattle. J. Vet. Pharmacol. Ther. 2002, 25, 111–120. [Google Scholar] [CrossRef]
- Njue, A.I.; Hayashi, J.; Kinne, L.; Feng, X.P.; Prichard, R.K. Mutations in the extracellular domains of glutamate-gated chloride channel α3 and β subunits from ivermectin-resistant Cooperia oncophora affect agonist sensitivity. J. Neurochem. 2004, 89, 1137–1147. [Google Scholar] [CrossRef]
- Pemberton, D.J.; Franks, C.J.; Walker, R.J.; Holden-Dye, L. Characterization of Glutamate-Gated Chloride Channels in the Pharynx of Wild-Type and Mutant Caenorhabditis elegansDelineates the Role of the Subunit GluCl-α2 in the Function of the Native Receptor. Mol. Pharmacol. 2001, 59, 1037–1043. [Google Scholar] [CrossRef]
- Flório, J.C.; Lebrun, I.; Bernardi, M.M.; de Souza Spinosa, H. Moxidectin interference on motor activity of rats. Braz. Arch. Biol. Technol. 2009, 52, 883–891. [Google Scholar]
- Kirst, H.A. The spinosyn family of insecticides: Realizing the potential of natural products research. J. Antibiot. 2010, 63, 101–111. [Google Scholar] [CrossRef]
- Floate, K.D.; Coghlin, P.C. No Support for Fluctuating Asymmetry as a Biomarker of Chemical Residues in Livestock Dung1. Can. Entomol. 2010, 142, 354–368. [Google Scholar] [CrossRef]
- El-Wakeil, N.; Abdallah, A. Cotton pests and the actual strategies for their management control. Agric. Res. Updates 2012, 7, 161–218. [Google Scholar]
- Orr, N.; Shaffner, A.J.; Richey, K.; Crouse, G.D. Novel mode of action of spinosad: Receptor binding studies demonstrating lack of interaction with known insecticidal target sites. Pestic. Biochem. Physiol. 2009, 95, 1–5. [Google Scholar] [CrossRef]
- Robertson-Plouch, C.; Baker, K.A.; Hozak, R.R.; Zimmermann, A.G.; Parks, S.C.; Herr, C.; Hart, L.M.; Jay, J.; Hutchens, D.E.; Snyder, D.E. Clinical field study of the safety and efficacy of spinosad chewable tablets for controlling fleas on dogs. Vet. Ther. Res. Appl. Vet. Med. 2008, 9, 26–36. [Google Scholar]
- Perry, T.; McKenzie, J.A.; Batterham, P. A Dα6 knockout strain of Drosophila melanogaster confers a high level of resistance to spinosad. Insect Biochem. Mol. Biol. 2007, 37, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Crump, A.; Omura, S. Ivermectin, “Wonder drug” from Japan: The human use perspective. Proc. Jpn. Acad. Ser. B 2011, 87, 13–28. [Google Scholar] [CrossRef]
- dos Santos, A.R.; Falcão, C.A.B.; Muzitano, M.F.; Kaiser, C.R.; Rossi-Bergmann, B.; Férézou, J.-P. Ivermectin-derived leishmanicidal compounds. Bioorg. Med. Chem. 2009, 17, 496–502. [Google Scholar] [CrossRef]
- Geary, T.G. Ivermectin 20 years on: Maturation of a wonder drug. Trends Parasitol. 2005, 21, 530–532. [Google Scholar] [CrossRef]
- Moncayo, A.L.; Vaca, M.; Amorim, L.; Rodriguez, A.; Erazo, S.; Oviedo, G.; Quinzo, I.; Padilla, M.; Chico, M.; Lovato, R. Impact of long-term treatment with ivermectin on the prevalence and intensity of soil-transmitted helminth infections. PLoS Negl. Trop. Dis. 2008, 2, e293. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Tayebwa, D.S.; Beshbishy, A.M.; N’Da, D.D.; Yokoyama, N.; Igarashi, I. Inhibitory effects of novel ciprofloxacin derivatives on the growth of four Babesia species and Theileria equi. Parasitol. Res. 2020, 1–13. [Google Scholar] [CrossRef]
- Bockarie, M.; Alexander, N.; Kazura, J.; Bockarie, F.; Griffin, L.; Alpers, M. Treatment with ivermectin reduces the high prevalence of scabies in a village in Papua New Guinea. Acta Trop. 2000, 75, 127–130. [Google Scholar] [CrossRef]
- Jacovella, J.; Chappuis, J.-P.; Kaoukhov, A.; Graeber, M.; Poncet, M.; Briantais, P.; Salin, L. Treatment of Papulopustular Rosacea with Ivermectin. U.S. Patents 9233118B2, 12 January 2016. [Google Scholar]
- Abegunde, A.T.; Ahuja, R.M.; Okafor, N.J. Doxycycline plus ivermectin versus ivermectin alone for treatment of patients with onchocerciasis. Cochrane Database Syst. Rev. 2016. [Google Scholar] [CrossRef]
- Hoerauf, A.; Mand, S.; Adjei, O.; Fleischer, B.; Büttner, D.W. Depletion of Wolbachia endobacteria in Onchocerca volvulus by doxycycline and microfilaridermia after ivermectin treatment. Lancet 2001, 357, 1415–1416. [Google Scholar] [CrossRef]
- Awadzi, K.; Opoku, N.; Attah, S.; Addy, E.; Duke, B.; Nyame, P.; Kshirsagar, N. The safety and efficacy of amocarzine in African onchocerciasis and the influence of ivermectin on the clinical and parasitological response to treatment. Ann. Trop. Med. Parasitol. 1997, 91, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Ottesen, E.A.; Campbell, W. Ivermectin in human medicine. J. Antimicrob. Chemother. 1994, 34, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Ottesen, E.A.; Vijayasekaran, V.; Kumaraswami, V.; Pillai, S.P.; Sadanandam, A.; Frederick, S.; Prabhakar, R.; Tripathy, S.P. A controlled trial of ivermectin and diethylcarbamazine in lymphatic filariasis. N. Engl. J. Med. 1990, 322, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Lynagh, T.; Lynch, J. Molecular mechanisms of Cys-loop ion channel receptor modulation by ivermectin. Front. Mol. Neurosci. 2012, 5, 60. [Google Scholar] [CrossRef]
- Kane, N.S.; Hirschberg, B.; Qian, S.; Hunt, D.; Thomas, B.; Brochu, R.; Ludmerer, S.W.; Zheng, Y.; Smith, M.; Arena, J.P.; et al. Drug-resistant Drosophila indicate glutamate-gated chloride channels are targets for the antiparasitics nodulisporic acid and ivermectin. Proc. Natl. Acad. Sci. USA 2000, 97, 13949–13954. [Google Scholar] [CrossRef]
- Siddique, S.; Syed, Q.; Adnan, A.; Qureshi, F.A. Production and screening of high yield avermectin B1b mutant of Streptomyces avermitilis 41445 through mutagenesis. Jundishapur J. Microbiol. 2014, 7. [Google Scholar] [CrossRef]
- Khalil, M.S.E.-d.H.; Alam, A.; Barakat, A.S.T. Nematicidal activity of some biopesticide agents and microorganisms against root-knot nematode on tomato plants under greenhouse conditions. J. Plant. Prot. Res. 2012, 52, 47–52. [Google Scholar] [CrossRef]
- Hussein, M.A.; Sabry, A.-K.H. Assessment of some new pesticides as molluscicides against the adult and eggs of chocolate banded snail, Eobania vermiculata. Bull. Nat. Res. Cent. 2019, 43, 75. [Google Scholar] [CrossRef]
- Jayakumar, J.; Rajendran, G.; Ramakrishnan, S. Management of reniform nematode, Rotylenchulus reniformis on okra through Streptomyces avermitilis. Indian J. Nematol. 2005, 35, 59–62. [Google Scholar]
- Blackburn, K.; Alm, S.; Yeh, T. Avermectin B1, isazofos, and fenamiphos for control of Hoplolaimus galeatus and Tylenchorhynchus dubius infesting Poa annua. J. Nematol. 1996, 28, 687. [Google Scholar] [PubMed]
- Das, A.K.; Maji, T.; Kadam, V.; Mukhopadhyay, A. Effect of nursery bed soil drenching with emamectin benzoate 1.9% WP, a novel avermectin derivative, towards eco-friendly management of root-knot nematodes in Chilli. Bioscan 2014, 9, 329–334. [Google Scholar]
- Lim, L.E.; Vilchèze, C.; Ng, C.; Jacobs, W.R.; Ramón-García, S.; Thompson, C.J. Anthelmintic avermectins kill Mycobacterium tuberculosis, including multidrug-resistant clinical strains. Antimicrob. Agents Chemother. 2013, 57, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Scherr, N.; Pluschke, G.; Thompson, C.J.; Ramón-García, S. Selamectin is the avermectin with the best potential for Buruli ulcer treatment. PLoS Negl. Trop. Dis. 2015, 9, e0003996. [Google Scholar] [CrossRef] [PubMed]
- Mshana, R.N.; Tadesse, G.; Abate, G.; Miörner, H. Use of 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide for rapid detection of rifampin-resistantMycobacterium tuberculosis. J. Clin. Microbiol. 1998, 36, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Miró-Canturri, A.; Ayerbe-Algaba, R.; Smani, Y. Drug repurposing for the treatment of bacterial and fungal infections. Front. Microbiol. 2019, 10, 41. [Google Scholar] [CrossRef]
- Butters, M.P.; Kobylinski, K.C.; Deus, K.M.; Da Silva, I.M.; Gray, M.; Sylla, M.; Foy, B.D. Comparative evaluation of systemic drugs for their effects against Anopheles gambiae. Acta Trop. 2012, 121, 34–43. [Google Scholar] [CrossRef][Green Version]
- Chatterjee, D. The mycobacterial cell wall: Structure, biosynthesis and sites of drug action. Curr. Opin. Chem. Biol. 1997, 1, 579–588. [Google Scholar] [CrossRef]
- Wolstenholme, A.; Rogers, A. Glutamate-gated chloride channels and the mode of action of the avermectin/milbemycin anthelmintics. Parasitology 2006, 131, S85–S95. [Google Scholar] [CrossRef]
- Cooper, D.J.; Rajahram, G.S.; William, T.; Jelip, J.; Mohammad, R.; Benedict, J.; Alaza, D.A.; Malacova, E.; Yeo, T.W.; Grigg, M.J. Plasmodium knowlesi malaria in Sabah, Malaysia, 2015–2017: Ongoing increase in incidence despite near-elimination of the human-only Plasmodium species. Clin. Infect. Dis. 2020, 70, 361–367. [Google Scholar] [CrossRef]
- Dhiman, S. Are malaria elimination efforts on right track? An analysis of gains achieved and challenges ahead. Infect. Dis. Poverty 2019, 8, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Foy, B.D.; Alout, H.; Seaman, J.A.; Rao, S.; Magalhaes, T.; Wade, M.; Parikh, S.; Soma, D.D.; Sagna, A.B.; Fournet, F. Efficacy and risk of harms of repeat ivermectin mass drug administrations for control of malaria (RIMDAMAL): A cluster-randomised trial. Lancet 2019, 393, 1517–1526. [Google Scholar] [CrossRef]
- Kobylinski, K.C.; Jittamala, P.; Hanboonkunupakarn, B.; Pukrittayakamee, S.; Pantuwatana, K.; Phasomkusolsil, S.; Davidson, S.A.; Winterberg, M.; Hoglund, R.M.; Mukaka, M. Safety, pharmacokinetics, and mosquito-lethal effects of ivermectin in combination with dihydroartemisinin-piperaquine and primaquine in healthy adult Thai subjects. Clin. Pharmacol. Ther. 2020, 107, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, L.P.; Sandri, T.L.; de Melo, E.J.T.; Fendel, R.; Kremsner, P.G.; Mordmüller, B.; Held, J. Ivermectin impairs the development of sexual and asexual stages of Plasmodium falciparum in vitro. Antimicrob. Agents Chemother. 2019, 63, e00085-19. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Snoeck, S.; Njiru, C.; Inak, E.; Dermauw, W.; Van Leeuwen, T. Geographical distribution and molecular insights into abamectin and milbemectin cross-resistance in European field populations of Tetranychus urticae. Pest. Manag. Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, R.; Mendes, A.M.; Prudêncio, M. Inhibition of Plasmodium sporogonic stages by ivermectin and other avermectins. Parasites Vectors 2019, 12, 549. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, L.A.; Mejias-Carpio, I.E.; Delgado-Noguera, L.A.; Manzanarez-Motezuma, J.P.; Escalona-Rodriguez, M.A.; Sordillo, E.M.; Mogollon-Rodriguez, E.A.; Hernandez-Pereira, C.E.; Marquez-Colmenarez, M.C.; Paniz-Mondolfi, A.E. Ivermectin: Repurposing a multipurpose drug for Venezuela’s humanitarian crisis. Int. J. Antimicrob. Agents 2020, 56, 106037. [Google Scholar] [CrossRef]
- Chaccour, C.J.; Kobylinski, K.C.; Bassat, Q.; Bousema, T.; Drakeley, C.; Alonso, P.; Foy, B.D. Ivermectin to reduce malaria transmission: A research agenda for a promising new tool for elimination. Malar. J. 2013, 12, 153. [Google Scholar] [CrossRef]
- Mekuriaw, W.; Balkew, M.; Messenger, L.A.; Yewhalaw, D.; Woyessa, A.; Massebo, F. The effect of ivermectin® on fertility, fecundity and mortality of Anopheles arabiensis fed on treated men in Ethiopia. Malar. J. 2019, 18, 1–10. [Google Scholar] [CrossRef]
- Dudeck, A.; Dudeck, J.; Scholten, J.; Petzold, A.; Surianarayanan, S.; Köhler, A.; Peschke, K.; Vöhringer, D.; Waskow, C.; Krieg, T. Mast cells are key promoters of contact allergy that mediate the adjuvant effects of haptens. Immunity 2011, 34, 973–984. [Google Scholar] [CrossRef]
- Natsuaki, Y.; Egawa, G.; Nakamizo, S.; Ono, S.; Hanakawa, S.; Okada, T.; Kusuba, N.; Otsuka, A.; Kitoh, A.; Honda, T. Perivascular leukocyte clusters are essential for efficient activation of effector T cells in the skin. Nat. Immunol. 2014, 15, 1064–1069. [Google Scholar] [CrossRef] [PubMed]
- Ventre, E.; Rozières, A.; Lenief, V.; Albert, F.; Rossio, P.; Laoubi, L.; Dombrowicz, D.; Staels, B.; Ulmann, L.; Julia, V. Topical ivermectin improves allergic skin inflammation. Allergy 2017, 72, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- Nörenberg, W.; Sobottka, H.; Hempel, C.; Plötz, T.; Fischer, W.; Schmalzing, G.; Schaefer, M. Positive allosteric modulation by ivermectin of human but not murine P2X7 receptors. Br. J. Pharmacol. 2012, 167, 48–66. [Google Scholar] [CrossRef] [PubMed]
- Wareham, K.; Vial, C.; Wykes, R.; Bradding, P.; Seward, E. Functional evidence for the expression of P2X1, P2X4 and P2X7 receptors in human lung mast cells. Br. J. Pharmacol. 2009, 157, 1215–1224. [Google Scholar] [CrossRef]
- Leyva-Castillo, J.M.; Hener, P.; Jiang, H.; Li, M. TSLP produced by keratinocytes promotes allergen sensitization through skin and thereby triggers atopic march in mice. J. Investig. Dermatol. 2013, 133, 154–163. [Google Scholar] [CrossRef]
- Thakur, P.; Dadsetan, S.; Fomina, A.F. Bidirectional coupling between ryanodine receptors and Ca2+ release-activated Ca2+ (CRAC) channel machinery sustains store-operated Ca2+ entry in human T lymphocytes. J. Biol. Chem. 2012, 287, 37233–37244. [Google Scholar] [CrossRef]
- Ci, X.; Li, H.; Yu, Q.; Zhang, X.; Yu, L.; Chen, N.; Song, Y.; Deng, X. Avermectin exerts anti-inflammatory effect by downregulating the nuclear transcription factor kappa-B and mitogen-activated protein kinase activation pathway. Fundam. Clin. Pharmacol. 2009, 23, 449–455. [Google Scholar] [CrossRef]
- Diao, H.; Cheng, N.; Zhao, Y.; Xu, H.; Dong, H.; Thamm, D.H.; Zhang, D.; Lin, D. Ivermectin inhibits canine mammary tumor growth by regulating cell cycle progression and WNT signaling. BMC Vet. Res. 2019, 15, 276. [Google Scholar] [CrossRef]
- Hashimoto, H.; Messerli, S.M.; Sudo, T.; Maruta, H. Ivermectin inactivates the kinase PAK1 and blocks the PAK1-dependent growth of human ovarian cancer and NF2 tumor cell lines. Drug Discov. Ther. 2009, 3, 243–246. [Google Scholar]
- Yu, Y.; Kanwar, S.S.; Patel, B.B.; Oh, P.-S.; Nautiyal, J.; Sarkar, F.H.; Majumdar, A.P. MicroRNA-21 induces stemness by downregulating transforming growth factor beta receptor 2 (TGFβR2) in colon cancer cells. Carcinogenesis 2012, 33, 68–76. [Google Scholar] [CrossRef]
- Ashraf, S.; Prichard, R. Ivermectin exhibits potent anti-mitotic activity. Vet. Parasitol. 2016, 226, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Feng, X.; Rong, H.; Pan, Z.; Inaba, Y.; Qiu, L.; Zheng, W.; Lin, S.; Wang, R.; Wang, Z. The antiparasitic drug ivermectin is a novel FXR ligand that regulates metabolism. Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Antebi, A.; Culotti, J.G.; Hedgecock, E.M. daf-12 regulates developmental age and the dauer alternative in Caenorhabditis elegans. Development 1998, 125, 1191–1205. [Google Scholar] [PubMed]
- Khoja, S.; Huynh, N.; Warnecke, A.M.; Asatryan, L.; Jakowec, M.W.; Davies, D.L. Preclinical evaluation of avermectins as novel therapeutic agents for alcohol use disorders. Psychopharmacology 2018, 235, 1697–1709. [Google Scholar] [CrossRef]
- Asatryan, L.; Yardley, M.M.; Khoja, S.; Trudell, J.R.; Hyunh, N.; Louie, S.G.; Petasis, N.A.; Alkana, R.L.; Davies, D.L. Avermectins differentially affect ethanol intake and receptor function: Implications for developing new therapeutics for alcohol use disorders. Int. J. Neuropsychopharmacol. 2014, 17, 907–916. [Google Scholar] [CrossRef]
- Yardley, M.M.; Huynh, N.; Rodgers, K.E.; Alkana, R.L.; Davies, D.L. Oral delivery of ivermectin using a fast dissolving oral film: Implications for repurposing ivermectin as a pharmacotherapy for alcohol use disorder. Alcohol 2015, 49, 553–559. [Google Scholar] [CrossRef][Green Version]
- Yardley, M.M.; Wyatt, L.; Khoja, S.; Asatryan, L.; Ramaker, M.J.; Finn, D.A.; Alkana, R.L.; Huynh, N.; Louie, S.G.; Petasis, N.A. Ivermectin reduces alcohol intake and preference in mice. Neuropharmacology 2012, 63, 190–201. [Google Scholar] [CrossRef]
- Franklin, K.M.; Hauser, S.R.; Lasek, A.W.; Bell, R.L.; McBride, W.J. Involvement of purinergic P2X4 receptors in alcohol intake of high-alcohol-drinking (HAD) rats. Alcohol. Clin. Exp. Res. 2015, 39, 2022–2031. [Google Scholar] [CrossRef]
- Yardley, M.M.; Neely, M.; Huynh, N.; Asatryan, L.; Louie, S.G.; Alkana, R.L.; Davies, D.L. Multi-Day Administration of Ivermectin is Effective in Reducing Alcohol Intake in Mice at Doses Shown to be Safe in Humans. Neuroreport 2014, 25, 1018. [Google Scholar] [CrossRef]
- Bortolato, M.; Yardley, M.M.; Khoja, S.; Godar, S.C.; Asatryan, L.; Finn, D.A.; Alkana, R.L.; Louie, S.G.; Davies, D.L. Pharmacological insights into the role of P2X4 receptors in behavioural regulation: Lessons from ivermectin. Int. J. Neuropsychopharmacol. 2013, 16, 1059–1070. [Google Scholar] [CrossRef]
- Guzzo, C.A.; Furtek, C.I.; Porras, A.G.; Chen, C.; Tipping, R.; Clineschmidt, C.M.; Sciberras, D.G.; Hsieh, J.Y.K.; Lasseter, K.C. Safety, tolerability, and pharmacokinetics of escalating high doses of ivermectin in healthy adult subjects. J. Clin. Pharmacol. 2002, 42, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Mendu, S.K.; Birnir, B. GABA is an effective immunomodulatory molecule. Amino Acids 2013, 45, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Khoja, S.; Shah, V.; Garcia, D.; Asatryan, L.; Jakowec, M.W.; Davies, D.L. Role of purinergic P2X4 receptors in regulating striatal dopamine homeostasis and dependent behaviors. J. Neurochem. 2016, 139, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Nuutinen, S.; Kiianmaa, K.; Panula, P. DARPP-32 and Akt regulation in ethanol-preferring AA and ethanol-avoiding ANA rats. Neurosci. Lett. 2011, 503, 31–36. [Google Scholar] [CrossRef]
- Pandey, S.C.; Roy, A.; Zhang, H.; Xu, T. Partial deletion of the cAMP response element-binding protein gene promotes alcohol-drinking behaviors. J. Neurosci. 2004, 24, 5022–5030. [Google Scholar] [CrossRef] [PubMed]
- Lespine, A.; Martin, S.; Dupuy, J.; Roulet, A.; Pineau, T.; Orlowski, S.; Alvinerie, M. Interaction of macrocyclic lactones with P-glycoprotein: Structure–affinity relationship. Eur. J. Pharm. Sci. 2007, 30, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Menez, C.; Sutra, J.-F.; Prichard, R.; Lespine, A. Relative neurotoxicity of ivermectin and moxidectin in Mdr1ab (−/−) mice and effects on mammalian GABA (A) channel activity. PLoS Negl. Trop. Dis. 2012, 6, e1883. [Google Scholar] [CrossRef]
- Janko, C.; Geyer, J. Moxidectin has a lower neurotoxic potential but comparable brain penetration in P-glycoprotein-deficient CF-1 mice compared to ivermectin. J. Vet. Pharmacol. Ther. 2013, 36, 275–284. [Google Scholar] [CrossRef]
- Huynh, N.; Arabian, N.; Naito, A.; Louie, S.; Jakowec, M.W.; Asatryan, L.; Davies, D.L. Preclinical development of moxidectin as a novel therapeutic for alcohol use disorder. Neuropharmacology 2017, 113, 60–70. [Google Scholar] [CrossRef]
- Drinyaev, V.A.; Mosin, V.A.; Kruglyak, E.B.; Novik, T.S.; Sterlina, T.S.; Ermakova, N.V.; Kublik, L.N.; Levitman, M.K.; Shaposhnikova, V.V.; Korystov, Y.N. Antitumor effect of avermectins. Eur. J. Pharmacol. 2004, 501, 19–23. [Google Scholar] [CrossRef]
- Campbell, W.; Fisher, M.; Stapley, E.; Albers-Schonberg, G.; Jacob, T. Ivermectin: A potent new antiparasitic agent. Science 1983, 221, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.J.; Lok, J.B. Macrocyclic lactones in the treatment and control of parasitism in small companion animals. Curr. Pharm. Biotechnol. 2012, 13, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Atif, M.; Estrada-Mondragon, A.; Nguyen, B.; Lynch, J.W.; Keramidas, A. Effects of glutamate and ivermectin on single glutamate-gated chloride channels of the parasitic nematode H. contortus. PLoS Pathog. 2017, 13, e1006663. [Google Scholar]
- Melotti, A.; Mas, C.; Kuciak, M.; Lorente-Trigos, A.; Borges, I.; Ruiz i Altaba, A. The river blindness drug Ivermectin and related macrocyclic lactones inhibit WNT-TCF pathway responses in human cancer. EMBO Mol. Med. 2014, 6, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Yang, Y.; Tian, Y.; Bai, J.; Zhang, X.; Li, X.; Peng, Z.; He, Y.; Chen, L.; Pan, Q. Ascl2 knockdown results in tumor growth arrest by miRNA-302b-related inhibition of colon cancer progenitor cells. PLoS ONE 2012, 7, e32170. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, J.; Liu, Y.; Chen, X.; Yu, T.; Jia, J.; Liu, C. PR55α, a regulatory subunit of PP2A, specifically regulates PP2A-mediated β-catenin dephosphorylation. J. Biol. Chem. 2009, 284, 22649–22656. [Google Scholar] [CrossRef]
- Seeling, J.M.; Miller, J.R.; Gil, R.; Moon, R.T.; White, R.; Virshup, D.M. Regulation of β-catenin signaling by the B56 subunit of protein phosphatase 2A. Science 1999, 283, 2089–2091. [Google Scholar] [CrossRef]
- Varnat, F.; Siegl-Cachedenier, I.; Malerba, M.; Gervaz, P.; Ruiz i Altaba, A. Loss of WNT-TCF addiction and enhancement of HH-GLI1 signalling define the metastatic transition of human colon carcinomas. EMBO Mol. Med. 2010, 2, 440–457. [Google Scholar] [CrossRef]
- Hirsch, D.; Barker, N.; McNeil, N.; Hu, Y.; Camps, J.; McKinnon, K.; Clevers, H.; Ried, T.; Gaiser, T. LGR5 positivity defines stem-like cells in colorectal cancer. Carcinogenesis 2014, 35, 849–858. [Google Scholar] [CrossRef]
- Ziskin, J.L.; Dunlap, D.; Yaylaoglu, M.; Fodor, I.K.; Forrest, W.F.; Patel, R.; Ge, N.; Hutchins, G.G.; Pine, J.K.; Quirke, P.; et al. In situ validation of an intestinal stem cell signature in colorectal cancer. Gut 2013, 62, 1012–1023. [Google Scholar] [CrossRef]
- Felipe de Sousa, E.M.; Colak, S.; Buikhuisen, J.; Koster, J.; Cameron, K.; de Jong, J.H.; Tuynman, J.B.; Prasetyanti, P.R.; Fessler, E.; van den Bergh, S.P. Methylation of cancer-stem-cell-associated Wnt target genes predicts poor prognosis in colorectal cancer patients. Cell Stem Cell 2011, 9, 476–485. [Google Scholar]
- Mastrangelo, E.; Pezzullo, M.; De Burghgraeve, T.; Kaptein, S.; Pastorino, B.; Dallmeier, K.; de Lamballerie, X.; Neyts, J.; Hanson, A.M.; Frick, D.N. Ivermectin is a potent inhibitor of flavivirus replication specifically targeting NS3 helicase activity: New prospects for an old drug. J. Antimicrob. Chemother. 2012, 67, 1884–1894. [Google Scholar] [CrossRef] [PubMed]
- Moseley, G.W.; Filmer, R.P.; DeJesus, M.A.; Jans, D.A. Nucleocytoplasmic distribution of rabies virus P-protein is regulated by phosphorylation adjacent to C-terminal nuclear import and export signals. Biochemistry 2007, 46, 12053–12061. [Google Scholar] [PubMed]
- Pryor, M.J.; Rawlinson, S.M.; Butcher, R.E.; Barton, C.L.; Waterhouse, T.A.; Vasudevan, S.G.; Bardin, P.G.; Wright, P.J.; Jans, D.A.; Davidson, A.D. Nuclear localization of dengue virus nonstructural protein 5 Through Its Importin α/β–recognized nuclear localization sequences is integral to viral infection. Traffic 2007, 8, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Rawlinson, S.M.; Pryor, M.J.; Wright, P.J.; Jans, D.A. CRM1-mediated nuclear export of dengue virus RNA polymerase NS5 modulates interleukin-8 induction and virus production. J. Biol. Chem. 2009, 284, 15589–15597. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, A.J.; Jans, D.A. Regulation of nucleocytoplasmic trafficking of viral proteins: An integral role in pathogenesis? Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 2176–2190. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.M.; Harrich, D.; Jans, D.A. Ivermectin is a specific inhibitor of importin α/β-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef]
- Rizzo, E. Ivermectin, antiviral properties and COVID-19: A possible new mechanism of action. Naunyn Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1153–1156. [Google Scholar] [CrossRef]
- Forwood, J.K.; Jans, D.A. Nuclear import pathway of the telomere elongation supressor TRF1: Inhibition by importin α. Biochemistry 2002, 41, 9333–9340. [Google Scholar]
- Wagstaff, K.M.; Rawlinson, S.M.; Hearps, A.C.; Jans, D.A. An AlphaScreen®-based assay for high-throughput screening for specific inhibitors of nuclear import. J. Biomol. Screen. 2011, 16, 192–200. [Google Scholar] [CrossRef]
- Cimica, V.; Chen, H.-C.; Iyer, J.K.; Reich, N.C. Dynamics of the STAT3 transcription factor: Nuclear import dependent on Ran and importin-β1. PLoS ONE 2011, 6, e20188. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Jans, D.A. Dual nuclear import mechanisms of sex determining factor SRY: Intracellular Ca2+ as a switch. FASEB J. 2011, 25, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Jäkel, S.; Albig, W.; Kutay, U.; Bischoff, F.R.; Schwamborn, K.; Doenecke, D.; Görlich, D. The importin β/importin 7 heterodimer is a functional nuclear import receptor for histone H1. EMBO J. 1999, 18, 2411–2423. [Google Scholar] [CrossRef] [PubMed]
- Mosammaparast, N.; Guo, Y.; Shabanowitz, J.; Hunt, D.F.; Pemberton, L.F. Pathways mediating the nuclear import of histones H3 and H4 in yeast. J. Biol. Chem. 2002, 277, 862–868. [Google Scholar] [CrossRef]
- Azeem, S.; Ashraf, M.; Rasheed, M.A.; Anjum, A.A.; Hameed, R. Evaluation of cytotoxicity and antiviral activity of ivermectin against Newcastle disease virus. Pak. J. Pharm. Sci. 2015, 28, 597–602. [Google Scholar]
- Hearps, A.C.; Jans, D.A. HIV-1 integrase is capable of targeting DNA to the nucleus via an importin α/β-dependent mechanism. Biochem. J. 2006, 398, 475–484. [Google Scholar] [CrossRef]
- Tavassoli, A. Targeting the protein–protein interactions of the HIV lifecycle. Chem. Soc. Rev. 2011, 40, 1337–1346. [Google Scholar] [CrossRef]
- Moreno, Y.; Nabhan, J.F.; Solomon, J.; Mackenzie, C.D.; Geary, T.G. Ivermectin disrupts the function of the excretory-secretory apparatus in microfilariae of Brugia malayi. Proc. Natl. Acad. Sci. USA 2010, 107, 20120–20125. [Google Scholar] [CrossRef]
- Xu, T.-L.; Han, Y.; Liu, W.; Pang, X.-Y.; Zheng, B.; Zhang, Y.; Zhou, X.-N. Antivirus effectiveness of ivermectin on dengue virus type 2 in Aedes albopictus. PLoS Negl. Trop. Dis. 2018, 12, e0006934. [Google Scholar] [CrossRef]
- Smit, M.R.; Ochomo, E.O.; Aljayyoussi, G.; Kwambai, T.K.; Abong’o, B.O.; Chen, T.; Bousema, T.; Slater, H.C.; Waterhouse, D.; Bayoh, N.M. Safety and mosquitocidal efficacy of high-dose ivermectin when co-administered with dihydroartemisinin-piperaquine in Kenyan adults with uncomplicated malaria (IVERMAL): A randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 2018, 18, 615–626. [Google Scholar] [CrossRef]
- Foy, B.D.; Kobylinski, K.C.; da Silva, I.M.; Rasgon, J.L.; Sylla, M. Endectocides for malaria control. Trends Parasitol. 2011, 27, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, E.; Bolognesi, M.; Milani, M. Flaviviral helicase: Insights into the mechanism of action of a motor protein. Biochem. Biophys. Res. Commun. 2012, 417, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Wang, Q.-Y.; Xu, H.Y.; Qing, M.; Kramer, L.; Yuan, Z.; Shi, P.-Y. Inhibition of dengue virus by targeting viral NS4B protein. J. Virol. 2011, 85, 11183–11195. [Google Scholar] [CrossRef] [PubMed]
- Julander, J.G.; Jha, A.K.; Choi, J.-A.; Jung, K.-H.; Smee, D.F.; Morrey, J.D.; Chu, C.K. Efficacy of 2′-C-methylcytidine against yellow fever virus in cell culture and in a hamster model. Antivir. Res. 2010, 86, 261–267. [Google Scholar] [CrossRef]
- Anastasiou, I.A.; Eleftheriadou, I.; Tentolouris, A.; Tsilingiris, D.; Tentolouris, N. In Vitro Data of Current Therapies for SARS-CoV-2. Curr. Med. Chem. 2020, 27, 4542–4548. [Google Scholar] [CrossRef]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antivir. Res. 2020, 178, 104787. [Google Scholar] [CrossRef]
- Lespine, A.; Alvinerie, M.; Sutra, J.-F.; Pors, I.; Chartier, C. Influence of the route of administration on efficacy and tissue distribution of ivermectin in goat. Vet. Parasitol. 2005, 128, 251–260. [Google Scholar] [CrossRef]
- Chaccour, C.; Hammann, F.; Ramón-García, S.; Rabinovich, N.R. Ivermectin and COVID-19: Keeping Rigor in Times of Urgency. Am. J. Trop. Med. Hyg. 2020, 102, 1156. [Google Scholar] [CrossRef]
- Lynch, J.W. Native glycine receptor subtypes and their physiological roles. Neuropharmacology 2009, 56, 303–309. [Google Scholar] [CrossRef]
- Ma, K.; Saha, P.K.; Chan, L.; Moore, D.D. Farnesoid X receptor is essential for normal glucose homeostasis. J. Clin. Investig. 2006, 116, 1102–1109. [Google Scholar] [CrossRef]
- Duran-Sandoval, D.; Cariou, B.; Percevault, F.; Hennuyer, N.; Grefhorst, A.; van Dijk, T.H.; Gonzalez, F.J.; Fruchart, J.-C.; Kuipers, F.; Staels, B. The farnesoid X receptor modulates hepatic carbohydrate metabolism during the fasting-refeeding transition. J. Biol. Chem. 2005, 280, 29971–29979. [Google Scholar] [CrossRef] [PubMed]
- Hubbert, M.L.; Zhang, Y.; Lee, F.Y.; Edwards, P.A. Regulation of hepatic Insig-2 by the farnesoid X receptor. Mol. Endocrinol. 2007, 21, 1359–1369. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Haas, N.; Lindemann, U.; Frank, K.; Sterry, W.; Lademann, J.; Katzung, W. Rapid and preferential sebum secretion of ivermectin: A new factor that may determine drug responsiveness in patients with scabies. Arch. Dermatol. 2002, 138, 1618. [Google Scholar] [CrossRef] [PubMed]
- Canga, A.G.; Prieto, A.M.S.; Liébana, M.J.D.; Martínez, N.F.; Vega, M.S.; Vieitez, J.J.G. The pharmacokinetics and metabolism of ivermectin in domestic animal species. Vet. J. 2009, 179, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Vanapalli, S.; Chen, Y.; Ellingrod, V.; Kitzman, D.; Lee, Y.; Hohl, R.; Fleckenstein, L. Orange juice decreases the oral bioavailability of ivermectin in healthy volunteers. Clin. Pharmacol. Ther. 2003, 73, P94. [Google Scholar] [CrossRef]
- Edwards, G.; Dingsdale, A.; Helsby, N.; Orme, M.E.; Breckenridge, A. The relative systemic availability of ivermectin after administration as capsule, tablet, and oral solution. Eur. J. Clin. Pharmacol. 1988, 35, 681–684. [Google Scholar] [CrossRef]
- Schulz, J.D.; Coulibaly, J.T.; Schindler, C.; Wimmersberger, D.; Keiser, J. Pharmacokinetics of ascending doses of ivermectin in Trichuris trichiura-infected children aged 2–12 years. J. Antimicrob. Chemother. 2019, 74, 1642–1647. [Google Scholar] [CrossRef]
- Schulz, J.D.; Neodo, A.; Coulibaly, J.T.; Keiser, J. Development and validation of a LC-MS/MS method for ivermectin quantification in dried blood spots: Application to a pharmacokinetic study in Trichuris trichiura-infected adults. Anal. Methods 2018, 10, 2901–2909. [Google Scholar] [CrossRef]
- Kudzi, W.; Dodoo, A.N.; Mills, J.J. Genetic polymorphisms in MDR1, CYP3A4 and CYP3A5 genes in a Ghanaian population: A plausible explanation for altered metabolism of ivermectin in humans? BMC Med. Genet. 2010, 11, 1–8. [Google Scholar] [CrossRef]
- Chhaiya, S.B.; Mehta, D.S.; Kataria, B.C. Ivermectin: Pharmacology and therapeutic applications. Int. J. Basic Clin. Pharm. 2012, 1, 132–139. [Google Scholar] [CrossRef]
- Fink, D.W.; Porras, A.G. Pharmacokinetics of ivermectin in animals and humans. In Ivermectin and Abamectin; Springer: Berlin/Heidelberg, Germany, 1989; pp. 113–130. [Google Scholar]
- Canga, A.G.; Prieto, A.M.S.; Liébana, M.J.D.; Martínez, N.F.; Vega, M.S.; Vieitez, J.J.G. The pharmacokinetics and interactions of ivermectin in humans—A mini-review. AAPS J. 2008, 10, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Halley, B.A.; VandenHeuvel, W.J.; Wislocki, P.G. Environmental effects of the usage of avermectins in livestock. Vet. Parasitol 1993, 48, 109–125. [Google Scholar] [CrossRef]
- Yamasmith, E.; Avirutnan, P.; Mairiang, D.; Tanrumluk, S.; Suputtamongkol, Y.; Saleh-arong, F. Efficacy and safety of ivermectin against dengue infection: A phase III, randomized, double-blind, placebo-controlled trial. In Proceedings of the 34th Annual Meeting the Royal College of Physicians of Thailand, Chonburi, Thailand, 28 April 2018. [Google Scholar]
- Smit, M.R.; Ochomo, E.; Aljayyoussi, G.; Kwambai, T.; Abong’o, B.; Bayoh, N.; Gimnig, J.; Samuels, A.; Desai, M.; Phillips-Howard, P.A. Efficacy and safety of high-dose ivermectin for reducing malaria transmission (IVERMAL): Protocol for a double-blind, randomized, placebo-controlled, dose-finding trial in Western Kenya. JMIR Res. Protoc. 2016, 5, e213. [Google Scholar] [CrossRef] [PubMed]
- Chaccour, C.; Lines, J.; Whitty, C.J. Effect of ivermectin on Anopheles gambiae mosquitoes fed on humans: The potential of oral insecticides in malaria control. J. Infect. Dis. 2010, 202, 113–116. [Google Scholar] [CrossRef]
- Foley, D.; Bryan, J.; Lawrence, G. The potential of ivermectin to control the malaria vector Anopheles farauti. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 625–628. [Google Scholar] [CrossRef]
- Ouédraogo, A.L.; Bastiaens, G.J.; Tiono, A.B.; Guelbéogo, W.M.; Kobylinski, K.C.; Ouédraogo, A.; Barry, A.; Bougouma, E.C.; Nebie, I.; Ouattara, M.S. Efficacy and safety of the mosquitocidal drug ivermectin to prevent malaria transmission after treatment: A double-blind, randomized, clinical trial. Clin. Infect. Dis. 2015, 60, 357–365. [Google Scholar] [CrossRef]
- Awadzi, K.; Opoku, N.O.; Attah, S.K.; Lazdins-Helds, J.; Kuesel, A.C. A randomized, single-ascending-dose, ivermectin-controlled, double-blind study of moxidectin in Onchocerca volvulus infection. PLoS Negl. Trop. Dis. 2014, 8, e2953. [Google Scholar] [CrossRef]
- McKellar, Q.A.; Gokbulut, C. Pharmacokinetic features of the antiparasitic macrocyclic lactones. Curr. Pharm. Biotechnol. 2012, 13, 888–911. [Google Scholar] [CrossRef]
- Clark, J.; Scott, J.; Campos, F.; Bloomquist, J. Resistance to avermectins: Extent, mechanisms, and management implications. Annu. Rev. Entomol. 1995, 40, 1–30. [Google Scholar] [CrossRef]
- Viktorov, A.; Yurkiv, V. Effect of ivermectin on function of liver macrophages. Bull. Exp. Biol. Med. 2003, 136, 569–571. [Google Scholar] [CrossRef]
- Yang, C.-C. Acute human toxicity of macrocyclic lactones. Curr. Pharm. Biotechnol. 2012, 13, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.; Breckenridge, C.B.; Wright, J. The Role of P-glycoprotein in Preventing Developmental and Neurotoxicity: Avermectins—A Case Study. In Hayes’ Handbook of Pesticide Toxicology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 2093–2110. [Google Scholar]
- Mealey, K.L.; Bentjen, S.A.; Gay, J.M.; Cantor, G.H. Ivermectin sensitivity in collies is associated with a deletion mutation of the mdr1 gene. Pharm. Genom. 2001, 11, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Umbenhauer, D.R.; Lankas, G.R.; Pippert, T.R.; Wise, L.D.; Cartwright, M.E.; Hall, S.J.; Beare, C.M. Identification of a P-glycoprotein-deficient subpopulation in the CF-1 mouse strain using a restriction fragment length polymorphism. Toxicol. Appl. Pharmacol. 1997, 146, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Sparreboom, A.; Van Asperen, J.; Mayer, U.; Schinkel, A.H.; Smit, J.W.; Meijer, D.K.; Borst, P.; Nooijen, W.J.; Beijnen, J.H.; Van Tellingen, O. Limited oral bioavailability and active epithelial excretion of paclitaxel (Taxol) caused by P-glycoprotein in the intestine. Proc. Natl. Acad. Sci. USA 1997, 94, 2031–2035. [Google Scholar] [CrossRef]
- Kwei, G.; Alvaro, R.; Chen, Q.; Jenkins, H.; Hop, C.; Keohane, C.; Ly, V.; Strauss, J.; Wang, R.; Wang, Z. Disposition of ivermectin and cyclosporin A in CF-1 mice deficient in mdr1a P-glycoprotein. Drug Metab. Dispos. 1999, 27, 581–587. [Google Scholar]
- Rawden, H.C.; Kokwaro, G.O.; Ward, S.A.; Edwards, G. Relative contribution of cytochromes P-450 and flavin-containing monoxygenases to the metabolism of albendazole by human liver microsomes. Br. J. Clin. Pharmacol. 2000, 49, 313–322. [Google Scholar] [CrossRef]
- Rawden, H.C. An Investigation of Potential Chemotherapeutic Drug Combinations with Albendazole for the Treatment of Echinococcus Infection; University of Liverpool: Liverpool, UK, 1999. [Google Scholar]
- Nobmann, S.; Bauer, B.; Fricker, G. Ivermectin excretion by isolated functionally intact brain endothelial capillaries. Br. J. Pharmacol. 2001, 132, 722–728. [Google Scholar] [CrossRef]
- Ibrahim, S.; Peggins, J.; Knapton, A.; Licht, T.; Aszalos, A. Influence of antipsychotic, antiemetic, and Ca2+ channel blocker drugs on the cellular accumulation of the anticancer drug daunorubicin: P-glycoprotein modulation. J. Pharmacol. Exp. Ther. 2000, 295, 1276–1283. [Google Scholar]
- Paasch, U.; Haustein, U.F. Management of endemic outbreaks of scabies with allethrin, permethrin, and ivermectin. Int. J. Dermatol. 2000, 39, 463–470. [Google Scholar] [CrossRef]
- Lankas, G.R.; Cartwright, M.E.; Umbenhauer, D. P-glycoprotein deficiency in a subpopulation of CF-1 mice enhances avermectin-induced neurotoxicity. Toxicol. Appl. Pharm. 1997, 143, 357–365. [Google Scholar] [CrossRef]
- Costa, L.G. Toxic effects of pesticides. In Casarett Doull’s Toxicol: The Basic Science of Poisons; McGraw-Hill Education: New York, NY, USA, 2008; Volume 8, pp. 883–930. [Google Scholar]
- Schinkel, A.H.; Smit, J.J.; van Tellingen, O.; Beijnen, J.H.; Wagenaar, E.; van Deemter, L.; Mol, C.A.; van der Valk, M.A.; Robanus-Maandag, E.C.; te Riele, H.P.; et al. Disruption of the mouse mdr1a P-glycoprotein gene leads to a deficiency in the blood-brain barrier and to increased sensitivity to drugs. Cell 1994, 77, 491–502. [Google Scholar] [CrossRef]
- Bissonnette, S.; Paradis, M.; Daneau, I.; Silversides, D.W. The ABCB1-1Delta mutation is not responsible for subchronic neurotoxicity seen in dogs of non-collie breeds following macrocyclic lactone treatment for generalized demodicosis. Vet. Dermatol. 2009, 20, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Blakley, B.R.; Rousseaux, C.G. Effect of ivermectin on the immune response in mice. Am. J. Vet. Res. 1991, 52, 593–595. [Google Scholar] [PubMed]
- Johnston, N.A.; Trammell, R.A.; Ball-Kell, S.; Verhulst, S.; Toth, L.A. Assessment of immune activation in mice before and after eradication of mite infestation. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2009, 48, 371–377. [Google Scholar]
- Davis, J.A.; Paylor, R.; McDonald, M.P.; Libbey, M.; Ligler, A.; Bryant, K.; Crawley, J.N. Behavioral effects of ivermectin in mice. Comp. Med. 1999, 49, 288–296. [Google Scholar]
- Molinari, G.; Soloneski, S.; Larramendy, M. New ventures in the genotoxic and cytotoxic effects of macrocyclic lactones, abamectin and ivermectin. Cytogenet. Genome Res. 2010, 128, 37–45. [Google Scholar] [CrossRef]
- Lankas, G.R.; Wise, L.D.; Cartwright, M.E.; Pippert, T.; Umbenhauer, D.R. Placental P-glycoprotein deficiency enhances susceptibility to chemically induced birth defects in mice. Reprod. Toxicol. 1998, 12, 457–463. [Google Scholar] [CrossRef]
- Flynn, B.M.; Brown, P.A.; Eckstein, J.M.; Strong, D. Treatment of Syphacia obvelata in mice using ivermectin. Lab. Anim. Sci. 1989, 39, 461–463. [Google Scholar]
- Burdett, E.C.; Heckmann, R.A.; Ochoa, R. Evaluation of five treatment regimens and five diagnostic methods for murine mites (Myocoptes musculinus and Myobia musculi). Contemp. Top. Lab. Anim. Sci. 1997, 36, 73–76. [Google Scholar]
- Conole, J.; Wilkinson, M.J.; McKellar, Q.A. Some observations on the pharmacological properties of ivermectin during treatment of a mite infestation in mice. Contemp. Top. Lab. Anim. Sci. 2003, 42, 42–45. [Google Scholar]
- Klement, P.; Augustine, J.M.; Delaney, K.H.; Klement, G.; Weitz, J.I. An oral ivermectin regimen that eradicates pinworms (Syphacia spp.) in laboratory rats and mice. Lab. Anim. Sci. 1996, 46, 286–290. [Google Scholar] [PubMed]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; Stephen Adeyemi, O.; Nadwa, E.; Rashwan, E.; Yokoyama, N.; Igarashi, I. Safety and efficacy of hydroxyurea and eflornithine against most blood parasites Babesia and Theileria. PLoS ONE 2020, 15, e0228996. [Google Scholar] [CrossRef] [PubMed]
- Lipman, N.S.; Dalton, S.D.; Stuart, A.R.; Arruda, K. Eradication of pinworms (Syphacia obvelata) from a large mouse breeding colony by combination oral anthelmintic therapy. Lab. Anim. Sci. 1994, 44, 517–520. [Google Scholar] [PubMed]
- Toth, L.A.; Oberbeck, C.; Straign, C.M.; Frazier, S.; Rehg, J.E. Toxicity evaluation of prophylactic treatments for mites and pinworms in mice. Contemp. Top. Lab. Anim. Sci. 2000, 39, 18–21. [Google Scholar] [PubMed]
- Silverman, J.; Blatt, H.; Lerro, A. Effect of ivermectin against Myobia-musculi. In Laboratory Animal Science; Amer Assoc Laboratory Animal Science: Cordova, TN, USA, 1983; p. 487. [Google Scholar]
- Boxall, A.B.; Tiede, K.; Bryning, G.; Bevan, R.; Tam, C.; Levy, L. Desk-based study of current knowledge on veterinary medicines in drinking water and estimation of potential levels. Report Ref. DWI 2011, 70, 235. [Google Scholar]
- Ricart Arbona, R.J.; Lipman, N.S.; Riedel, E.R.; Wolf, F.R. Treatment and eradication of murine fur mites: I. Toxicologic evaluation of ivermectin-compounded feed. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2010, 49, 564–570. [Google Scholar]
- Gosting, D.C.; Doyle, M.E. Food Safety 1990: An Annotated Bibliography of the Literature; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Lankas, G.; Minsker, D.; Robertson, R. Effects of ivermectin on reproduction and neonatal toxicity in rats. Food Chem. Toxicol. 1989, 27, 523–529. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Compound | Chemical Structures | Synonyms | IUPAC Name | Molecular Formula | Ref. |
|---|---|---|---|---|---|
| Ivermectin |  | Eqvalan Ivermectin Ivomec Mectizan MK 933 Stromectol | 1R,4S,5′S,6R,6′R,8R,10E,12S,13S,14E,16E,20R,21R,24S)-6′-[(2S)-butan-2-yl]-21,24-dihydroxy-12-[(2R,4S,5S,6S)-5-[(2S,4S,5S,6S)-5-hydroxy-4-methoxy-6-methyloxan-2-yl]oxy-4-methoxy-6-methyloxan-2-yl]oxy-5′,11,13,22-tetramethylspiro[3,7,19-trioxatetracyclo[15.6.1.14,8.020,24]pentacosa-10,14,16,22-tetraene-6,2′-oxane]-2-one | C48H74O14 | [39] |
| Selamectin |  | Selamectin UNII- 220119-17-5 A2669OWX9N UK-124,114 | ((2aE,4E,8E,20Z)-(5′S,6S,6′S,7S,11R,13R,15S,17aR,20aR,20bS)-6′-cyclohexyl-3′,4′,5′,6,6′,7,10,11,14,15,17a,20,20a,20b-tetradecahydro-20b-hydroxy-20-(hydroxyimino)-5′,6,8,19-tetramethyl-17-oxospiro[11,15-methano-2H,13H,17H-furo[4,3,2-pq][2,6]benzodioxacyclooctadecin-13,2′-[2H]pyran]-7-yl 2,6-dideoxy-3-O-methyl-α-L-arabino-hexopyranoside | C43H63NO11 | |
| Moxidectin |  | ProHeart 6 CL301423 Cydectin | 1R,4S,5′S,6R,6′S,8R,10E,12S,13S,14E,16E,20R,21Z,24S)-6′-cyclohexyl-24-hydroxy-21-hydroxyimino-12-[(2R,4S,5S,6S)-5-hydroxy-4-methoxy-6-methyloxan-2-yl]oxy-5′,11,13,22-tetramethylspiro[3,7,19-trioxatetracyclo[15.6.1.14,8.020,24]pentacosa-10,14,16,22-tetraene-6,2′-oxane]-2-one | C37H53NO8 | |
| Doramectin |  | DECTOMAX UK-67,994 | (1′R,2R,3S,4′S,6S,8′R,10′E,12′S,13′S,14′E,16′E,20′R,21′R,24′S)-2-cyclohexyl-21′,24′-dihydroxy-12′-[(2R,4S,5S,6S)-5-[(2S,4S,5S,6S)-5-hydroxy-4-methoxy-6-methyloxan-2-yl]oxy-4-methoxy-6-methyloxan-2-yl]oxy-3,11′,13′,22′-tetramethylspiro[2,3-dihydropyran-6,6′-3,7,19-trioxatetracyclo[15.6.1.14,8.020,24]pentacosa-10,14,16,22-tetraene]-2′-one | C50H74O14 | |
| Eprinomectin |  | Avermectin B1, 4″-(acetylamino)-4″-deoxy-, (4″R)- Eprinex Eprinomectin [USAN:USP:INN] MK 397 ZINC306122586 | (N-[(2S,3R,4S,6S)-6-[(2S,3S,4S,6R)-6-[(1′R,2R,3S,4′S,6S,8′R,10′E,12′S,13′S,14′E,16′E,20′R,21′R,24′S)-21′,24′-dihydroxy-3,11′,13′,22′-tetramethyl-2′-oxo-2-propan-2-ylspiro[2,3-dihydropyran-6,6′-3,7,19-trioxatetracyclo[15.6.1.14,8.020,24]pentacosa-10,14,16,22-tetraene]-12′-yl]oxy-4-methoxy-2-methyloxan-3-yl]oxy-4-methoxy-2-methyloxan-3-yl]acetamido | C49H73NO14 | |
| Nemadectin |  | Nemadectin UNII-1Y8VJ1G3TY 1Y8VJ1G3TY 102130-84-7 Nemadectina | ((1R,4S,4′S,5′S,6R,6′S,8R,10E,13R,14E,16E,20R,21R,24S)-4′,21,24-trihydroxy-5′,11,13,22-tetramethyl-6′-[(E)-4-methylpent-2-en-2-yl]spiro[3,7,19-trioxatetracyclo[15.6.1.14,8.020,24]pentacosa-10,14,16,22-tetraene-6,2′-oxane]-2-one | C36H52O8 | |
| Milbemycin |  | Milbemycin D Antibiotic B 41D Milbemycin B 41D UNII-04S0E2XJQI | ((1R,4S,5′S,6R,6′R,8R,10E,13R,14E,16E,20R,21R,24S)-21,24-dihydroxy-5′,11,13,22-tetramethyl-6′-propan-2-ylspiro[3,7,19-trioxatetracyclo[15.6.1.14,8.020,24]pentacosa-10,14,16,22-tetraene-6,2′-oxane]-2-one | C33H48O7 | |
| Abamectin |  | Abamectinum Agrimek Vertimec Affirm Avomec | ((1′R,2R,3S,4′S,6S,8′R,10′E,12′S,13′S,14′E,16′E,20′R,21′R,24′S)-2-butan-2-yl-21′,24′-dihydroxy-12′-[(2R,4S,5S,6S)-5-[(2S,4S,5S,6S)-5-hydroxy-4-methoxy-6-methyloxan-2-yl]oxy-4-methoxy-6-methyloxan-2-yl]oxy-3,11′,13′,22′-tetramethylspiro[2,3-dihydropyran-6,6′-3,7,19-trioxatetracyclo[15.6.1.14,8.020,24]pentacosa-10,14,16,22-tetraene]-2′-one | C95H142O28 |
| Uses | Ref. |
|---|---|
| Onchocerca volvulus | [73] |
| Strongyloidiasis | [74,75] |
| Scabies | [78] |
| Papulopustular rosacea (PPR) | [79] |
| Pediculosis | [74,76] |
| Gnathostomiasis | |
| Myiasis | |
| Leishmaniasis | |
| Trichuris | |
| Ascaris |
| Activity | Compound | Organism Active Against | Ref. |
|---|---|---|---|
| Nematocidal | Abamectin | Hoplolaimus galeatus and Tylenchorynchus dubius, E. vermiculata | [88,89] |
| Antibacterial | Ivermectin Selamectin Doramectin Moxidectin | Mycobacterium bovis and M. tuberculosis | [93,96,97] |
| Anti-plasmodium | Ivermectin | P. falciparum | [104] |
| Anti-inflammatory | Ivermectin | Skin inflammation | [111] |
| Anticancer | Ivermectin | Lung cancer and colon cancer | [118] |
| Antiviral | Ivermectin | Dengue virus and yellow fever virus | [28] |
| Derivative | Clinical Trial | Ref. |
|---|---|---|
| Ivermectin | Phase III clinical trial in dengue patient in Thailand shows it is safe but of no clinical benefit | [194] |
| Moxidectin | Undergoing phase III clinical trial to treat patient Onchocerca volvulus infection | [199] |
| Selamectin | Not approved for human use | [200] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Saber Batiha, G.; Alqahtani, A.; Ilesanmi, O.B.; Saati, A.A.; El-Mleeh, A.; Hetta, H.F.; Magdy Beshbishy, A. Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects. Pharmaceuticals 2020, 13, 196. https://doi.org/10.3390/ph13080196
El-Saber Batiha G, Alqahtani A, Ilesanmi OB, Saati AA, El-Mleeh A, Hetta HF, Magdy Beshbishy A. Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects. Pharmaceuticals. 2020; 13(8):196. https://doi.org/10.3390/ph13080196
Chicago/Turabian StyleEl-Saber Batiha, Gaber, Ali Alqahtani, Omotayo B. Ilesanmi, Abdullah A. Saati, Amany El-Mleeh, Helal F. Hetta, and Amany Magdy Beshbishy. 2020. "Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects" Pharmaceuticals 13, no. 8: 196. https://doi.org/10.3390/ph13080196
APA StyleEl-Saber Batiha, G., Alqahtani, A., Ilesanmi, O. B., Saati, A. A., El-Mleeh, A., Hetta, H. F., & Magdy Beshbishy, A. (2020). Avermectin Derivatives, Pharmacokinetics, Therapeutic and Toxic Dosages, Mechanism of Action, and Their Biological Effects. Pharmaceuticals, 13(8), 196. https://doi.org/10.3390/ph13080196