Is the Way to Fight Cancer Paved with Gold? Metal-Based Carbene Complexes with Multiple and Fascinating Biological Features

 ,
, 



 ,
,  and
and 
Abstract
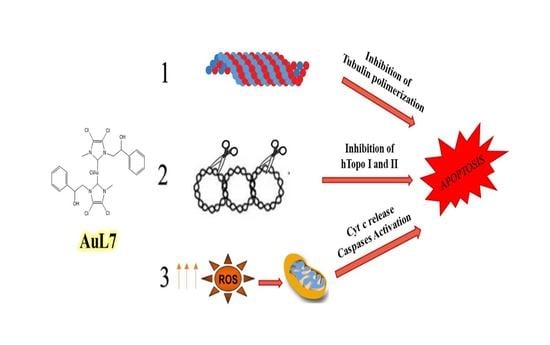
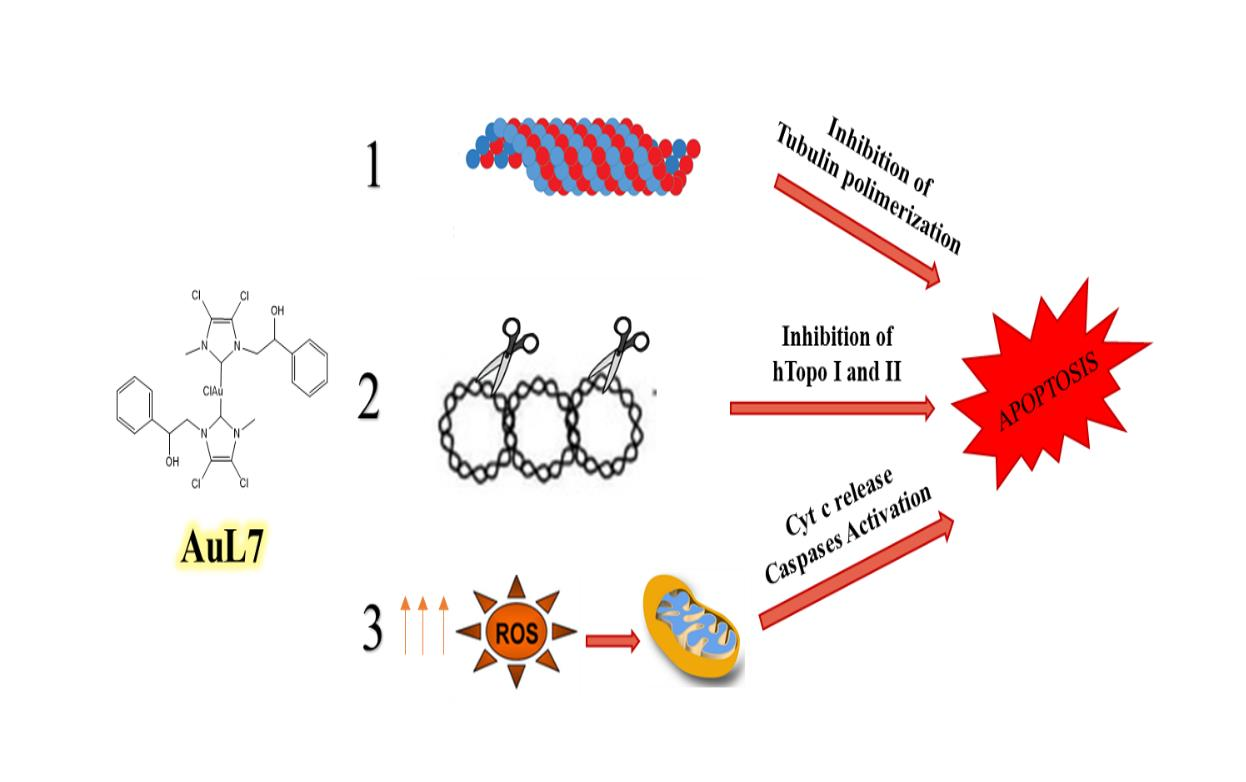
1. Introduction
2. Results
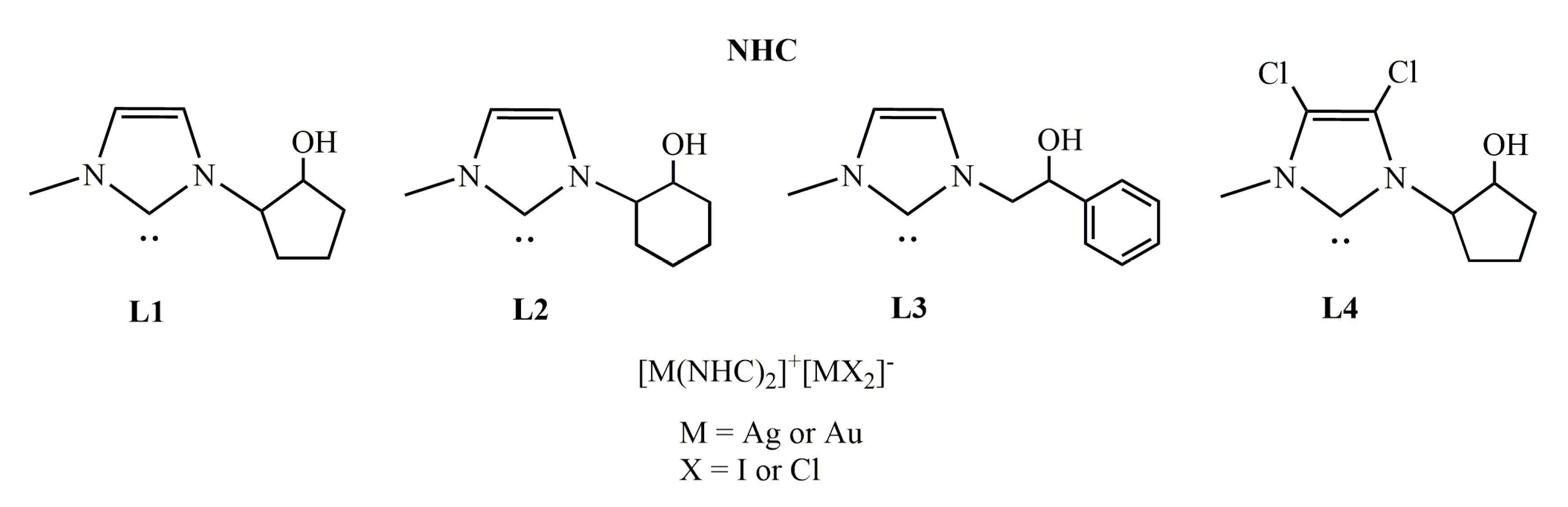
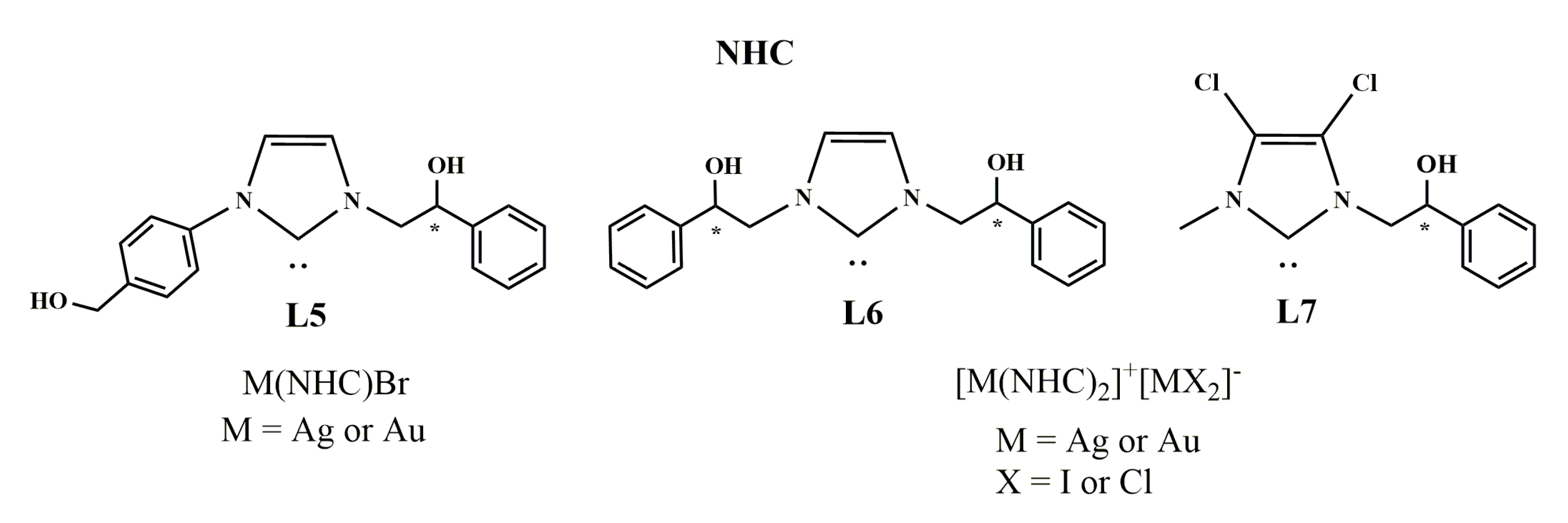
2.1. Chemistry
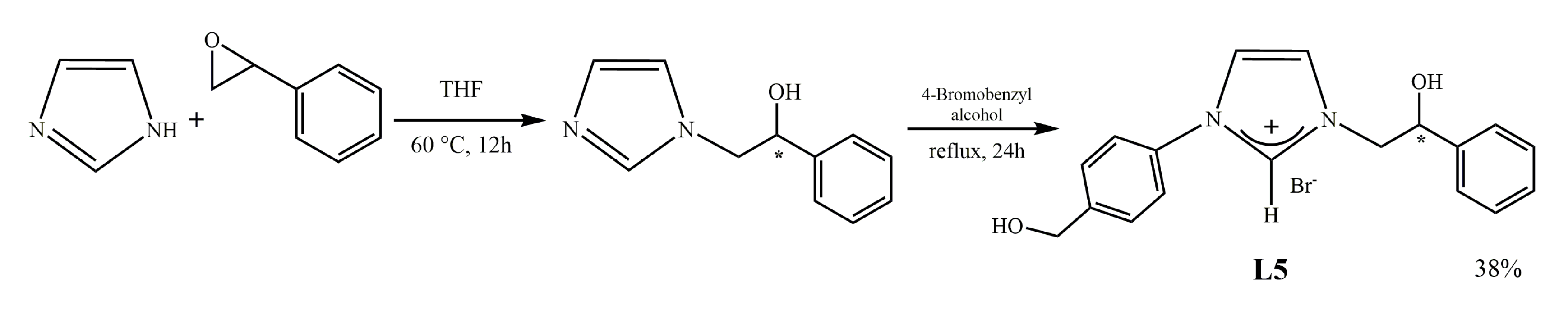
2.1.1. Synthesis of N-[4-(hydroxymethyl)phenyl]-N’-[(2-hydroxy-2-phenyl)ethyl]-imidazolium bromide (L5)
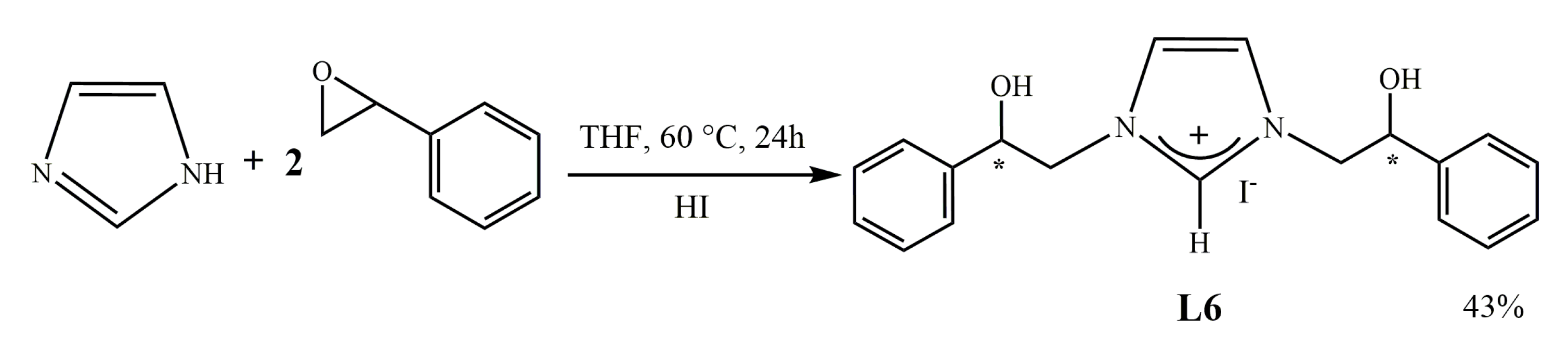
2.1.2. Synthesis of [N,N’-[(2-hydroxy-2-phenyl)ethyl]-imidazolium iodide (L6)
2.1.3. Synthesis of N-[4-(hydroxymethyl)phenyl]-N’-[(2-hydroxy-2-phenyl)ethyl]-imidazole-2-ylidene]-silver(I)bromide] (AgL5)
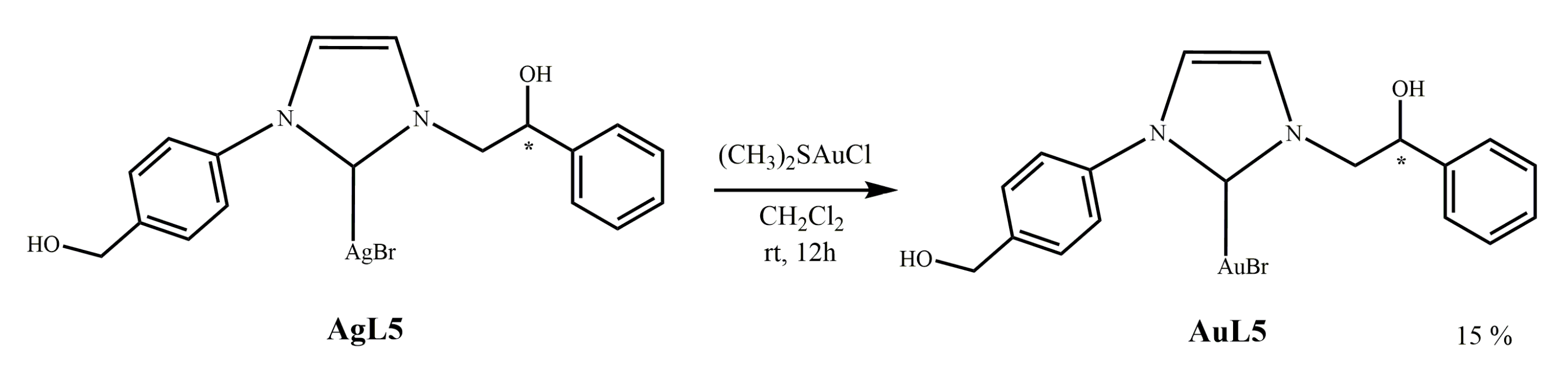
2.1.4. Synthesis of N-[4-(hydroxymethyl)phenyl]-N’-(2-hydroxy-2-phenyl)ethyl]-imidazole- 2-ylidene]-gold(I)bromide](AuL5)
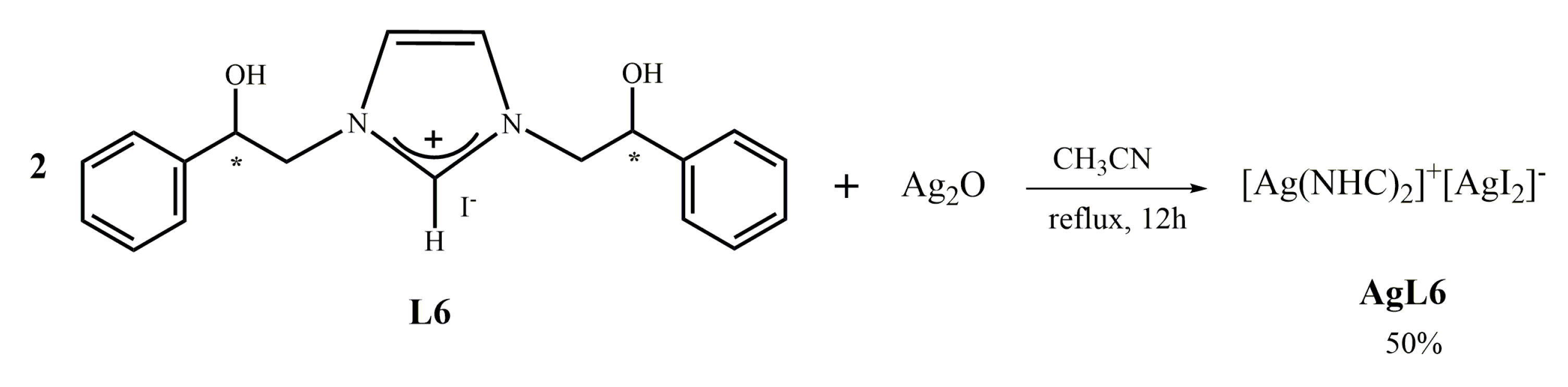
2.1.5. Synthesis of bis-[N,N’-(2-hydroxy-2-phenyl)ethyl]-imidazole-2-ylidene]silver(I)]+[di-iodide-silver]− (AgL6)
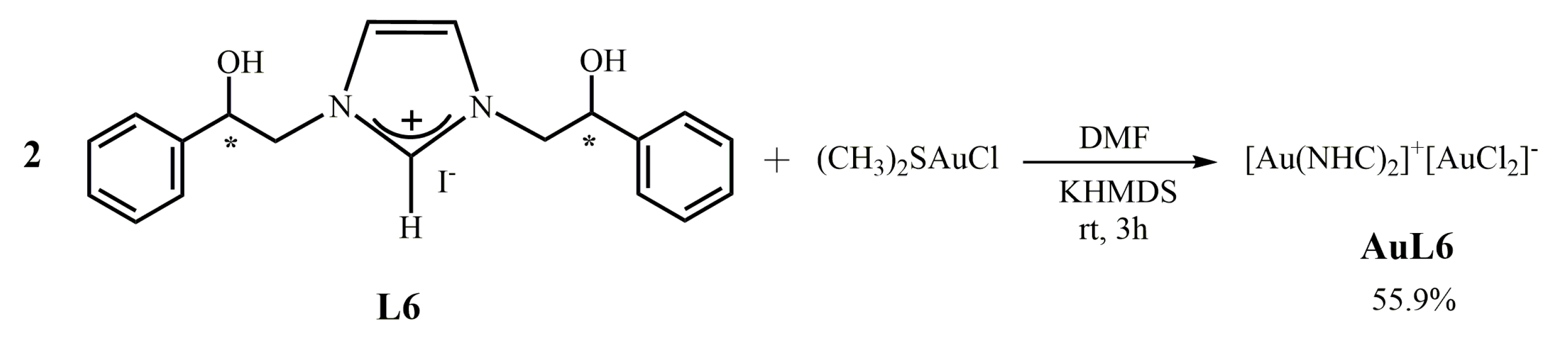
2.1.6. Synthesis of bis-[N,N’-(2-hydroxy-2-phenyl)ethyl]-imidazole-2-ylidene]gold(I)]+[dichloro-gold]− (AuL6)
2.2. Biology
2.2.1. Anti-Tumor Activity
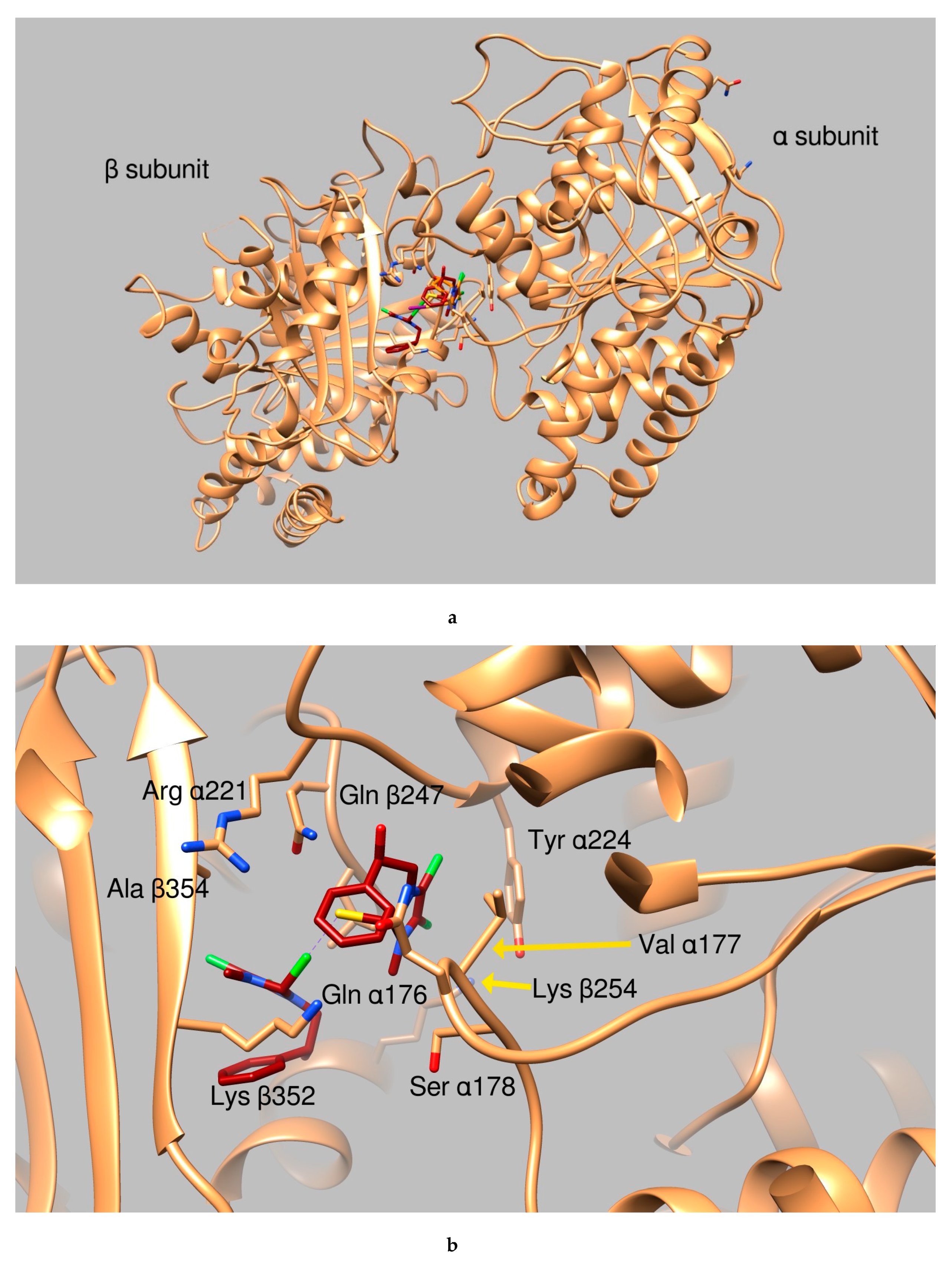
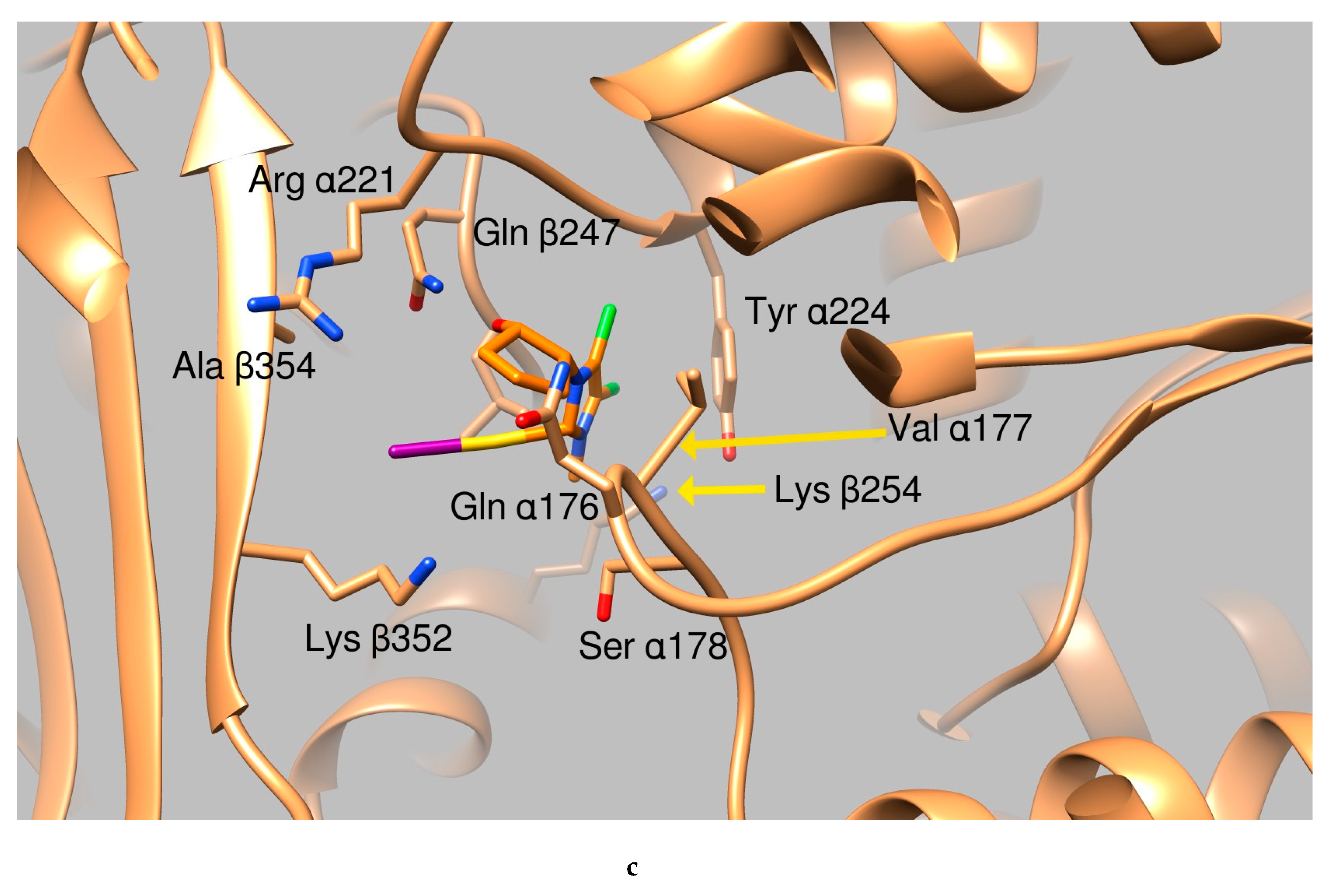
2.2.2. Docking Studies
- green background: good clusterization of the simulation results and ligand binding modes that occur in a protein area which is either involved in DNA binding or in protein-protein interactions (i.e., tubulin oligomer formation, topoisomerase dimerization etc.)
- orange background: no (or small) clusterization of the results indicating low probability of ligand binding
- yellow background: good clusterization of the simulation results but the ligand binding mode occurs in an area not involved in interactions with DNA or in a zone not involved in oligomeric formation.
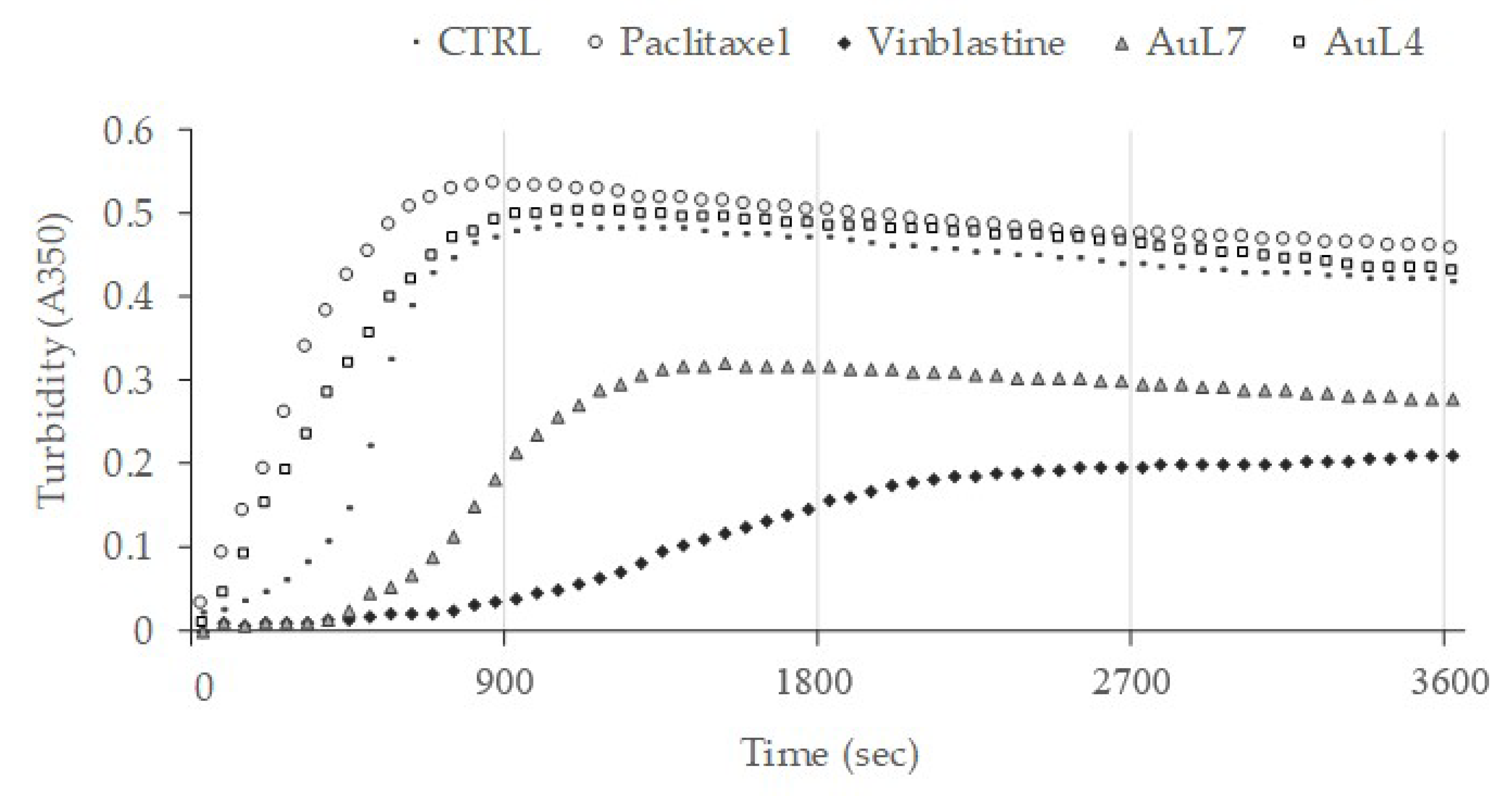
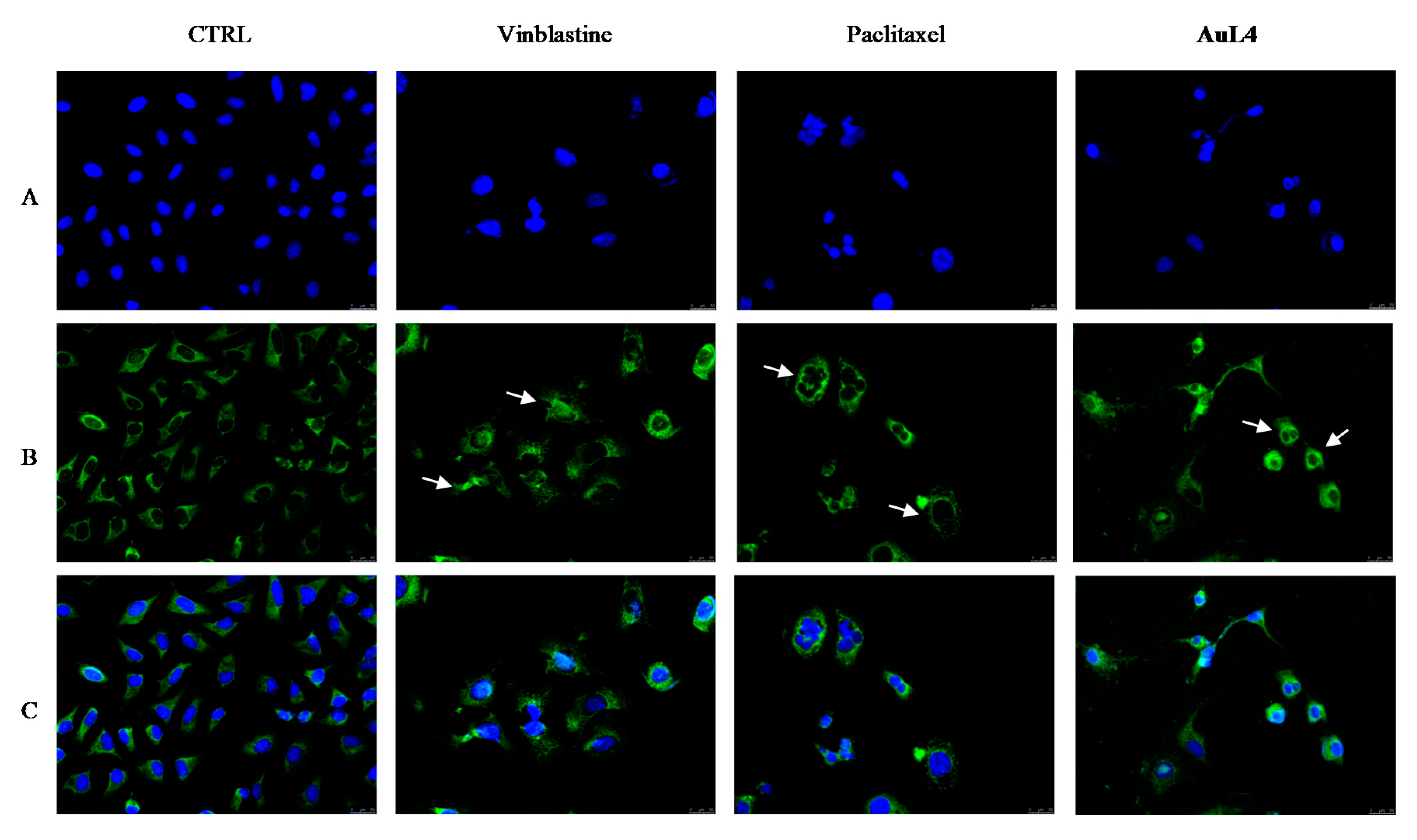
2.2.3. AuL7 Induces Cell Cytoskeleton Destabilization
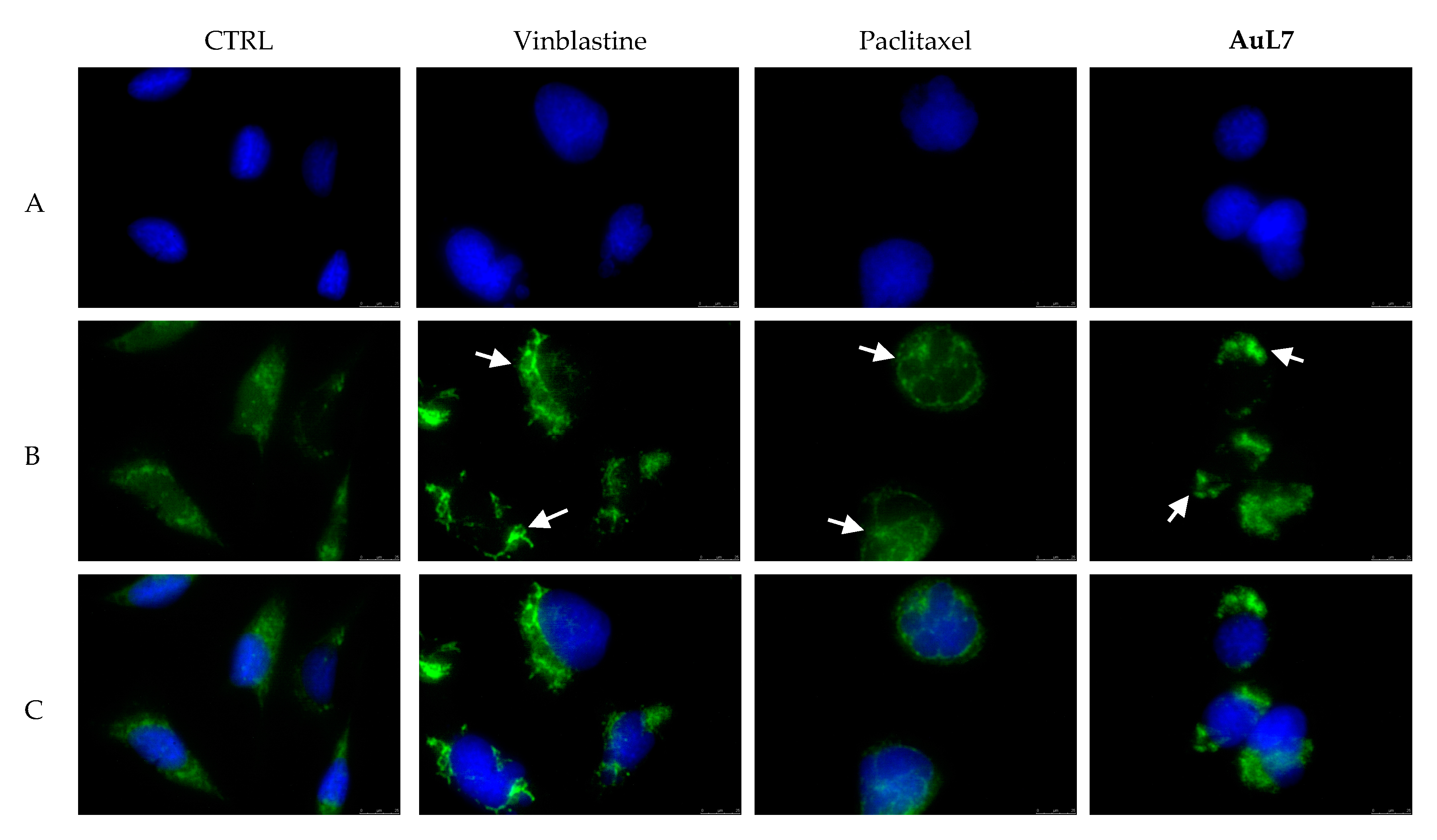
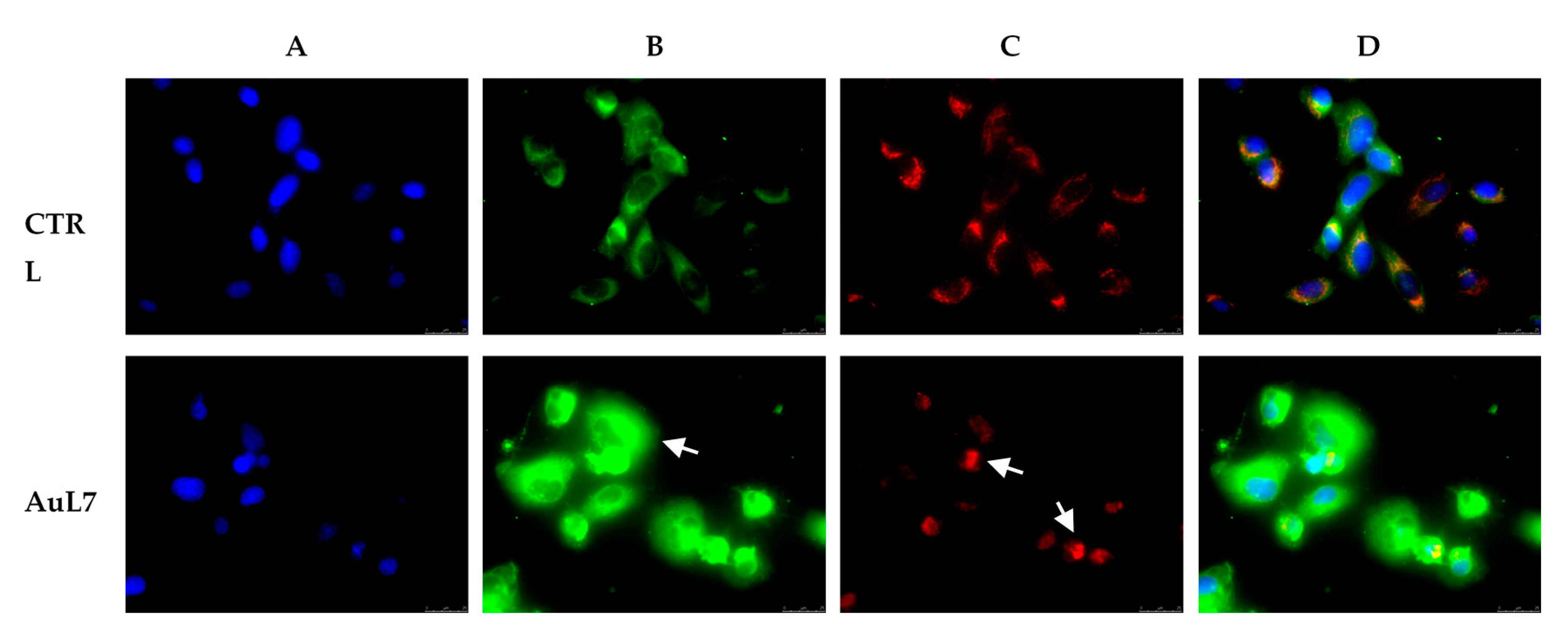
2.2.4. Immunostaining Studies
2.2.5. Human Topoisomerases Inhibition
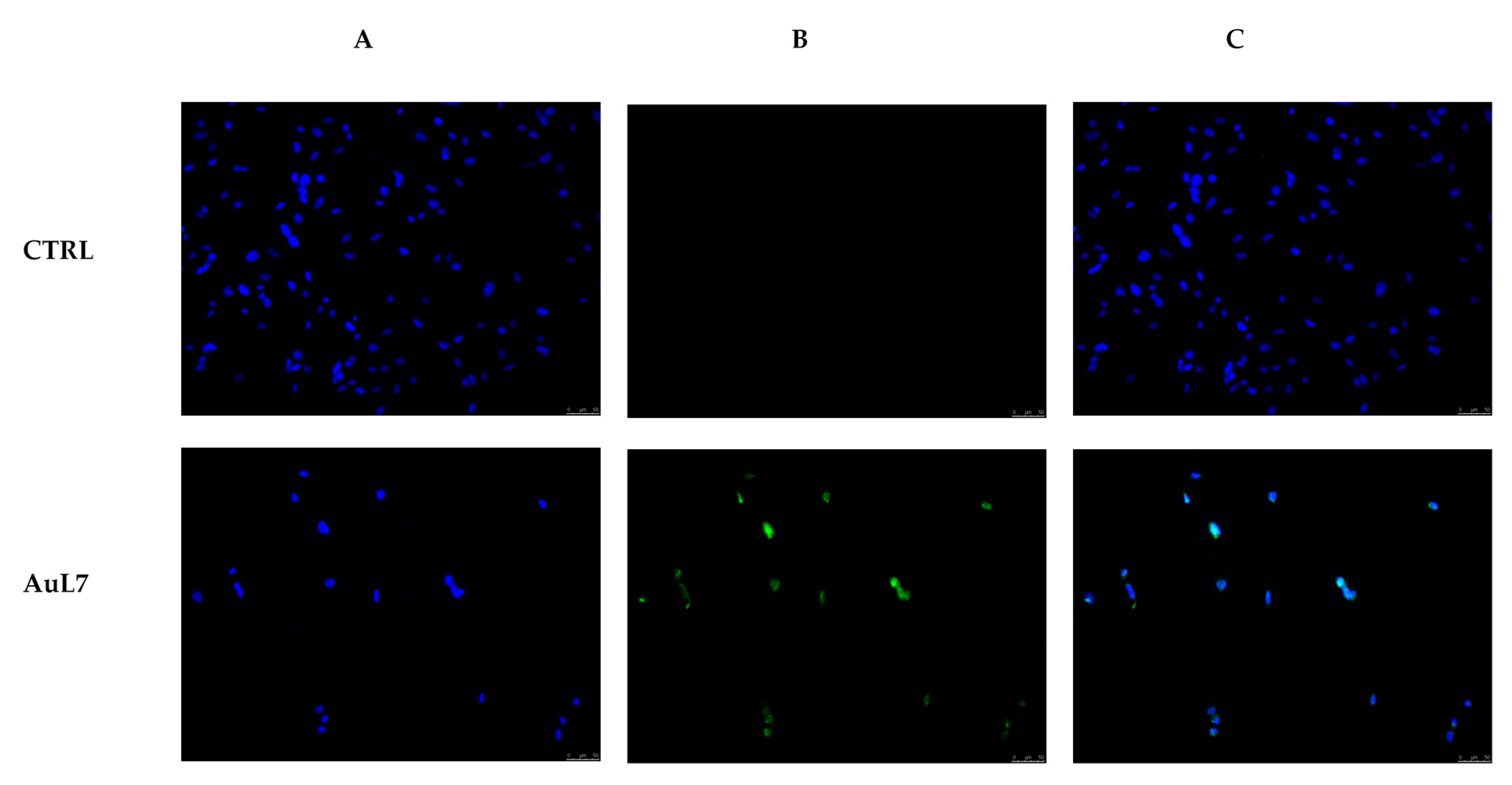
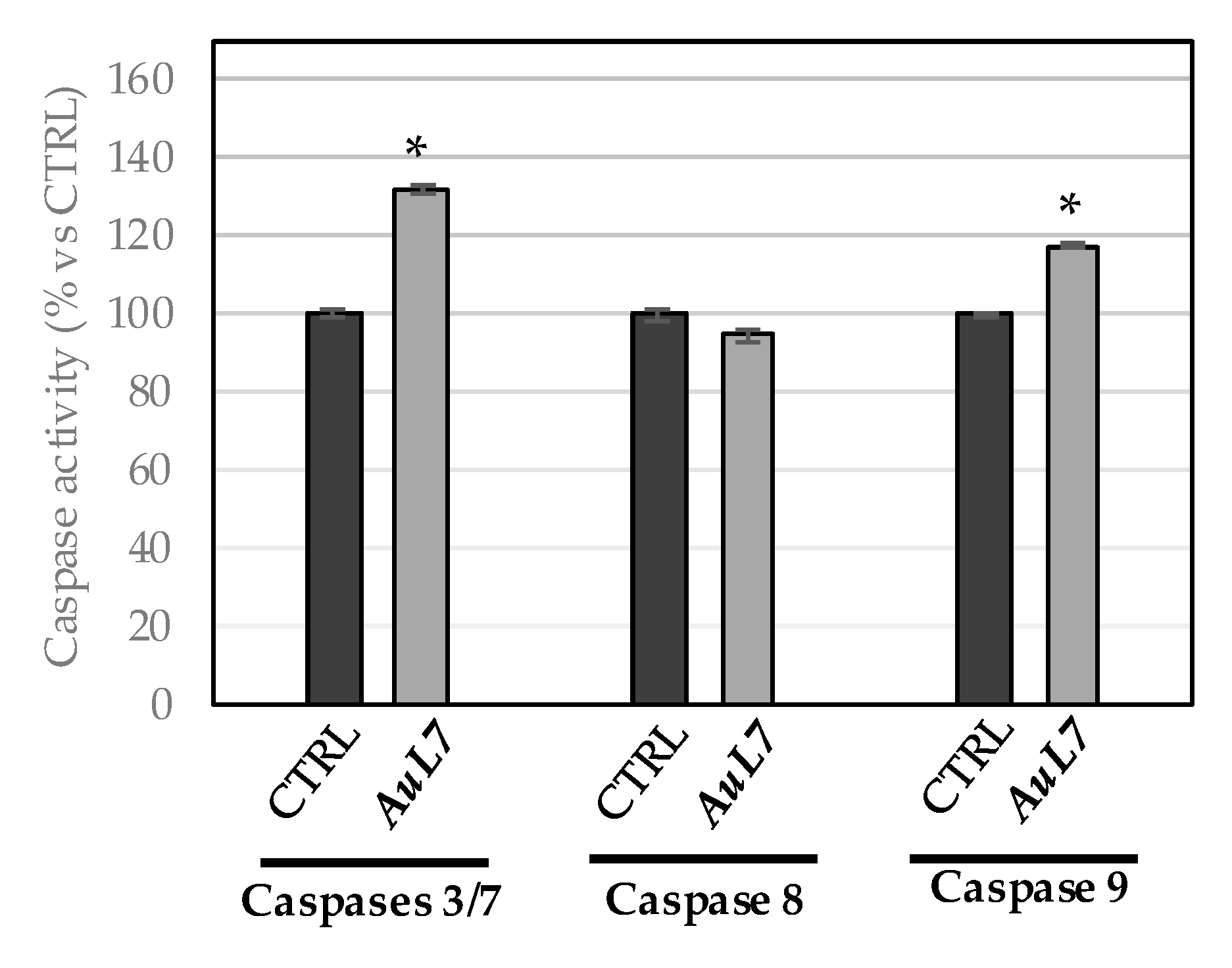
2.2.6. AuL7 Induces Cancer Cells Apoptosis
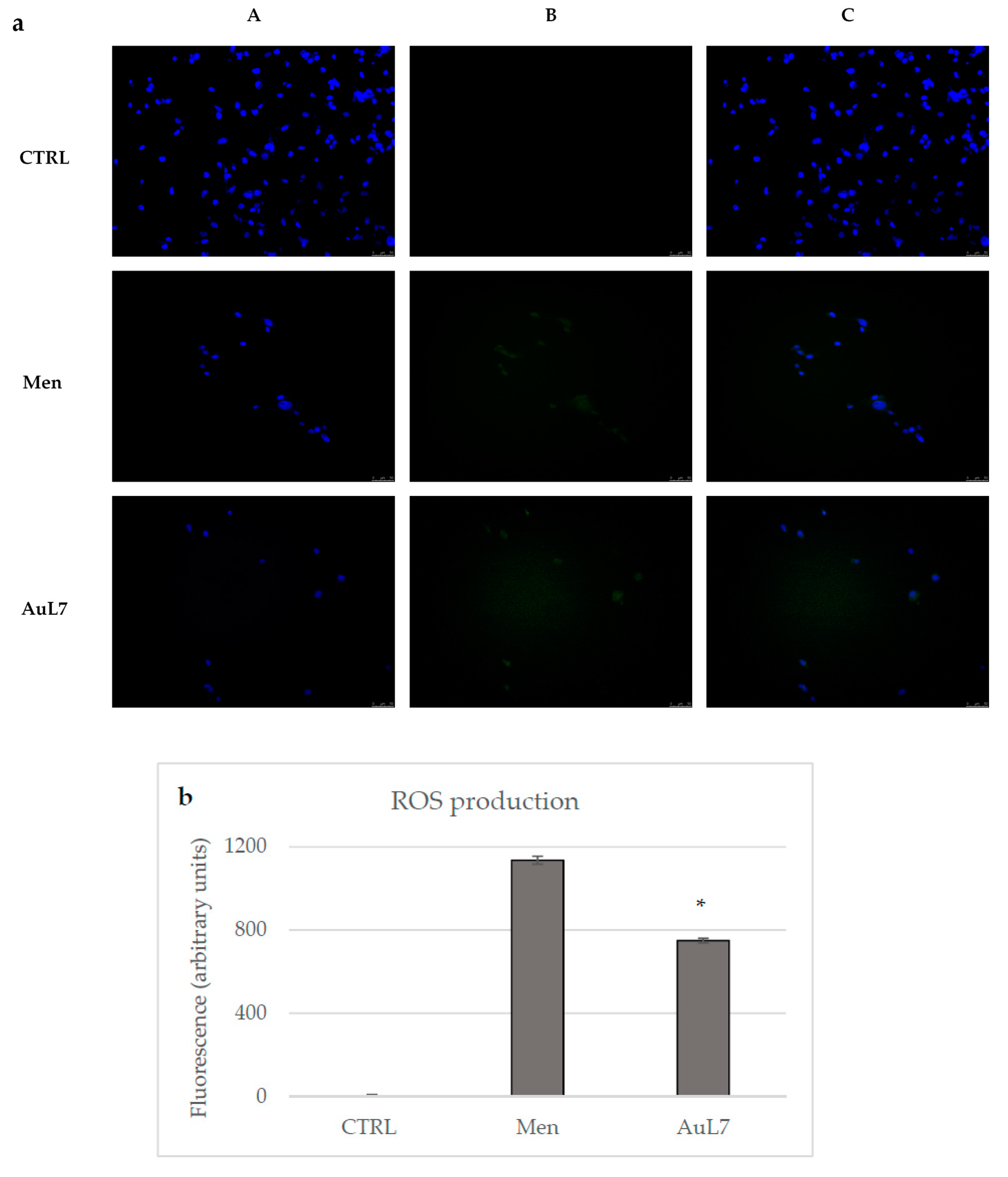
2.2.7. AuL7 Induces Reactive Oxygen Species Formation and the Mitochondria-Mediated Apoptosis Pathway
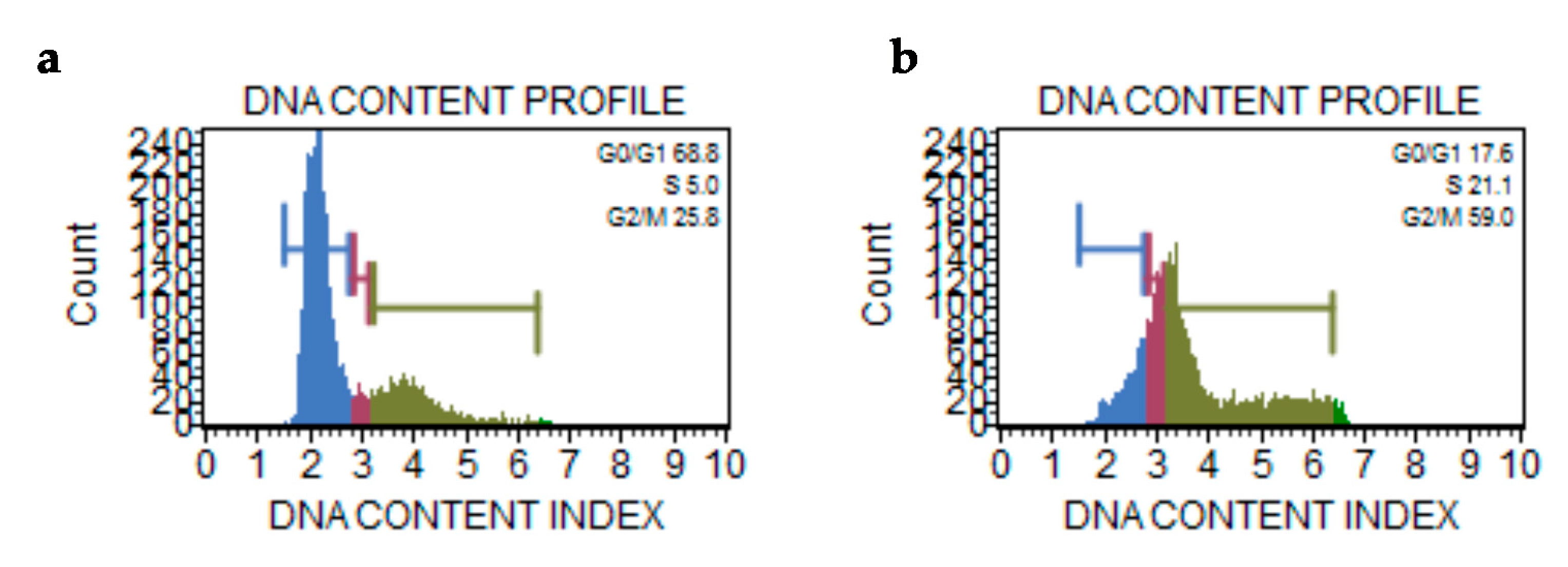
2.2.8. Cell Cycle Assay
3. Discussion
4. Materials and Methods
4.1. Chemistry
4.1.1. Synthesis of N-substituted bis-imidazolium Salts (L5, L6)
4.1.2. Synthesis of Complexes AgL5, AuL5, AgL6, and AuL6
4.2. Docking Simulations
4.3. Biology
4.3.1. Cell Culture
4.3.2. Cell Viability
4.3.3. Detection of Intracellular H2O2 (ROS)
4.3.4. TUNEL Assay
4.3.5. Caspase Assay
4.3.6. Cytoskeleton and Mitochondrial Staining and Immunofluorescence
4.3.7. In Vitro Tubulin-Polymerization Assay
4.3.8. Human Topoisomerase I Relaxation Assay
4.3.9. Human Topoisomerase II Decatenation Assay
4.3.10. Cell Cycle
4.3.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Lu, J.J.; Pan, W.; Hu, Y.J.; Wang, Y.T. Multi-target drugs: The trend of drug research and development. PLoS ONE 2012, 7, e40262. [Google Scholar] [CrossRef] [PubMed]
- Ceramella, J.; Caruso, A.; Occhiuzzi, M.A.; Iacopetta, D.; Barbarossa, A.; Rizzuti, B.; Dallemagne, P.; Rault, S.; El-Kashef, H.; Saturnino, C.; et al. Benzothienoquinazolinones as new multi-target scaffolds: Dual inhibition of human Topoisomerase I and tubulin polymerization. Eur. J. Med. Chem. 2019, 181, 111583. [Google Scholar] [CrossRef] [PubMed]
- Gatzka, M.V. Targeted Tumor Therapy Remixed-An Update on the Use of Small-Molecule Drugs in Combination Therapies. Cancers 2018, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Ghoncheh, M.; Momenimovahed, Z.; Salehiniya, H. Epidemiology, Incidence and Mortality of Breast Cancer in Asia. Asian Pac. J. Cancer Prev. 2016, 17, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Navarro Rosenblatt, D.A.; Chan, D.S.; Abar, L.; Vingeliene, S.; Vieira, A.R.; Greenwood, D.C.; Norat, T. Anthropometric factors and ovarian cancer risk: A systematic review and nonlinear dose-response meta-analysis of prospective studies. Int. J. Cancer 2015, 136, 1888–1898. [Google Scholar] [CrossRef] [PubMed]
- Zucca-Matthes, G.; Urban, C.; Vallejo, A. Anatomy of the nipple and breast ducts. Gland Surg. 2016, 5, 32–36. [Google Scholar]
- Gehrig, P.A.; Bae-Jump, V.L.; Boggess, J.F.; Groben, P.A.; Fowler, W.C., Jr.; Van Le, L. Association between uterine serous carcinoma and breast cancer. Gynecol. Oncol. 2004, 94, 208–211. [Google Scholar] [CrossRef]
- Khanna, S.; Jana, B.; Saha, A.; Kurkute, P.; Ghosh, S.; Verma, S. Targeting cytotoxicity and tubulin polymerization by metal-carbene complexes on a purine tautomer platform. Dalton Trans. 2014, 43, 9838–9842. [Google Scholar] [CrossRef]
- Ceramella, J.; Mariconda, A.; Iacopetta, D.; Saturnino, C.; Barbarossa, A.; Caruso, A.; Rosano, C.; Sinicropi, M.S.; Longo, P. From coins to cancer therapy: Gold, silver and copper complexes targeting human topoisomerases. Bioorg Med. Chem Lett 2020, 30, 126905. [Google Scholar] [CrossRef]
- Saturnino, C.; Barone, I.; Iacopetta, D.; Mariconda, A.; Sinicropi, M.S.; Rosano, C.; Campana, A.; Catalano, S.; Longo, P.; Ando, S. N-heterocyclic carbene complexes of silver and gold as novel tools against breast cancer progression. Future Med. Chem. 2016, 8, 2213–2229. [Google Scholar] [CrossRef]
- Iacopetta, D.; Mariconda, A.; Saturnino, C.; Caruso, A.; Palma, G.; Ceramella, J.; Muia, N.; Perri, M.; Sinicropi, M.S.; Caroleo, M.C.; et al. Novel Gold and Silver Carbene Complexes Exert Antitumor Effects Triggering the Reactive Oxygen Species Dependent Intrinsic Apoptotic Pathway. ChemMedChem 2017, 12, 2054–2065. [Google Scholar] [CrossRef] [PubMed]
- Mariconda, A.; Sirignano, M.; Costabile, C.; Longo, P. New NHC- silver and gold complexes active in A3-coupling (aldehyde-alkyne-amine) reaction. Mol. Catal 2020, 480, 110570. [Google Scholar] [CrossRef]
- Arnold, P.L.; Rodden, M.; Davis, K.M.; Scarisbrick, A.C.; Blake, A.J.; Wilson, C. Asymmetric lithium(I) and copper(II) alkoxy-N-heterocyclic carbene complexes; crystallographic characterisation and Lewis acid catalysis. Chem. Commun. (Camb) 2004. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.L.; Liddle, S.T. F-block N-heterocyclic carbene complexes. Chem. Commun. (Camb) 2006. [Google Scholar] [CrossRef] [PubMed]
- Pugh, D.; Danopoulos, A.A. Metal complexes with ‘pincer’-type ligands incorporating N-heterocyclic carbene functionalities. Coord Chem. Rev. 2007, 251, 610–641. [Google Scholar] [CrossRef]
- Zhou, Y.B.; Chen, W.Z. Synthesis and characterization of square-planar tetranuclear silver and gold clusters supported by a pyrazole-linked bis(N-heterocyclic carbene) ligand. Organometallics 2007, 26, 2742–2746. [Google Scholar] [CrossRef]
- Bocchino, C.; Napoli, M.; Costabile, C.; Longo, P. Synthesis of Octahedral Zirconium Complex Bearing [NHC-O] Ligands, and Its Behavior as Catalyst in the Polymerization of Olefins. J. Polym. Sci. A Polym. Chem. 2011, 49, 862–870. [Google Scholar] [CrossRef]
- Lee, C.K.; Lee, K.M.; Lin, I.J.B. Inorganic-organic hybrid lamella of di- and tetranuclear silver-carbene complexes. Organometallics 2002, 21, 10–12. [Google Scholar] [CrossRef]
- Arduengo, A.J.; Dias, H.V.R.; Calabrese, J.C.; Davidson, F. Homoleptic Carbene Silver(I) and Carbene Copper(I) Complexes. Organometallics 1993, 12, 3405–3409. [Google Scholar] [CrossRef]
- Ku, R.Z.; Huang, J.C.; Cho, J.Y.; Kiang, F.M.; Reddy, K.R.; Chen, Y.C.; Lee, K.J.; Lee, J.H.; Lee, G.H.; Peng, S.M.; et al. Metal ion mediated transfer and cleavage of diaminocarbene ligands. Organometallics 1999, 18, 2145–2154. [Google Scholar] [CrossRef]
- Liu, Q.X.; Hu, Z.L.; Yu, S.C.; Zhao, Z.X.; Wei, D.C.; Li, H.L. NHC Pd(II) and Ag(I) Complexes: Synthesis, Structure, and Catalytic Activity in Three Types of C-C Coupling Reactions. ACS Omega 2018, 3, 4035–4047. [Google Scholar] [CrossRef] [PubMed]
- Mariconda, A.; Grisi, F.; Costabile, C.; Falcone, S.; Bertolasi, V.; Longo, P. Synthesis, characterization and catalytic behaviour of a palladium complex bearing a hydroxy-functionalized N-heterocyclic carbene ligand. New J. Chem. 2014, 38, 762–769. [Google Scholar] [CrossRef]
- Jakubowicz-Gil, J.; Paduch, R.; Piersiak, T.; Glowniak, K.; Gawron, A.; Kandefer-Szerszen, M. The effect of quercetin on pro-apoptotic activity of cisplatin in HeLa cells. Biochem. Pharmacol. 2005, 69, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Aldemir, M.; Okulu, E.; Kosemehmetoglu, K.; Ener, K.; Topal, F.; Evirgen, O.; Gurleyik, E.; Avci, A. Evaluation of the protective effect of quercetin against cisplatin-induced renal and testis tissue damage and sperm parameters in rats. Andrologia 2014, 46, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Gonzalez, P.D.; Lopez-Hernandez, F.J.; Duenas, M.; Prieto, M.; Sanchez-Lopez, E.; Thomale, J.; Ruiz-Ortega, M.; Lopez-Novoa, J.M.; Morales, A.I. Differential effect of quercetin on cisplatin-induced toxicity in kidney and tumor tissues. Food Chem. Toxicol. 2017, 107, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Choie, D.D.; Longnecker, D.S.; del Campo, A.A. Acute and chronic cisplatin nephropathy in rats. Lab. Invest. 1981, 44, 397–402. [Google Scholar]
- Matsushima, H.; Yonemura, K.; Ohishi, K.; Hishida, A. The role of oxygen free radicals in cisplatin-induced acute renal failure in rats. J. Lab. Clin. Med. 1998, 131, 518–526. [Google Scholar] [CrossRef]
- Iacopetta, D.; Grande, F.; Caruso, A.; Mordocco, R.A.; Plutino, M.R.; Scrivano, L.; Ceramella, J.; Muia, N.; Saturnino, C.; Puoci, F.; et al. New insights for the use of quercetin analogs in cancer treatment. Future Med. Chem. 2017, 9, 2011–2028. [Google Scholar] [CrossRef]
- Hatia, S.; Septembre-Malaterre, A.; Le Sage, F.; Badiou-Beneteau, A.; Baret, P.; Payet, B.; Lefebvre d’hellencourt, C.; Gonthier, M.P. Evaluation of antioxidant properties of major dietary polyphenols and their protective effect on 3T3-L1 preadipocytes and red blood cells exposed to oxidative stress. Free Radic. Res. 2014, 48, 387–401. [Google Scholar] [CrossRef]
- Majewska, M.; Skrzycki, M.; Podsiad, M.; Czeczot, H. Evaluation of antioxidant potential of flavonoids: An in vitro study. Acta Pol. Pharm. 2011, 68, 611–615. [Google Scholar]
- Lappano, R.; Rosano, C.; Pisano, A.; Santolla, M.F.; De Francesco, E.M.; De Marco, P.; Dolce, V.; Ponassi, M.; Felli, L.; Cafeo, G.; et al. A calixpyrrole derivative acts as an antagonist to GPER, a G-protein coupled receptor: Mechanisms and models. Dis Model. Mech. 2015, 8, 1237–1246. [Google Scholar] [CrossRef]
- Sinicropi, M.S.; Lappano, R.; Caruso, A.; Santolla, M.F.; Pisano, A.; Rosano, C.; Capasso, A.; Panno, A.; Lancelot, J.C.; Rault, S.; et al. (6-bromo-1,4-dimethyl-9H-carbazol-3-yl-methylene)-hydrazine (carbhydraz) acts as a GPER agonist in breast cancer cells. Curr. Top. Med. Chem. 2015, 15, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Stec-Martyna, E.; Ponassi, M.; Miele, M.; Parodi, S.; Felli, L.; Rosano, C. Structural comparison of the interaction of tubulin with various ligands affecting microtubule dynamics. Curr. Cancer Drug Targets 2012, 12, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.L.; Zhu, X.J.; Feng, Y.M.; Huang, N.; Luo, G.F.; Mao, Y.J.; Han, X.F.; Tian, W.; Wang, G.R.; Han, X.B.; et al. Development of a Novel Class of Tubulin Inhibitors with Promising Anticancer Activities. Mol. Cancer Res. 2013, 11, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.A. Microtubule inhibitors: Differentiating tubulin-inhibiting agents based on mechanisms of action, clinical activity, and resistance. Mol. Cancer Ther. 2009, 8, 2086–2095. [Google Scholar] [CrossRef]
- You, F.; Gao, C. Topoisomerase Inhibitors and Targeted Delivery in Cancer Therapy. Curr. Top. Med. Chem. 2019, 19, 713–729. [Google Scholar] [CrossRef]
- Iacopetta, D.; Rosano, C.; Puoci, F.; Parisi, O.I.; Saturnino, C.; Caruso, A.; Longo, P.; Ceramella, J.; Malzert-Freon, A.; Dallemagne, P.; et al. Multifaceted properties of 1,4-dimethylcarbazoles: Focus on trimethoxybenzamide and trimethoxyphenylurea derivatives as novel human topoisomerase II inhibitors. Eur. J. Pharm. Sci. 2017, 96, 263–272. [Google Scholar] [CrossRef]
- Sinicropi, M.S.; Iacopetta, D.; Rosano, C.; Randino, R.; Caruso, A.; Saturnino, C.; Muia, N.; Ceramella, J.; Puoci, F.; Rodriquez, M.; et al. N-thioalkylcarbazoles derivatives as new anti-proliferative agents: Synthesis, characterisation and molecular mechanism evaluation. J. Enzyme Inhib. Med. Chem. 2018, 33, 434–444. [Google Scholar] [CrossRef]
- Saturnino, C.; Caruso, A.; Iacopetta, D.; Rosano, C.; Ceramella, J.; Muia, N.; Mariconda, A.; Bonomo, M.G.; Ponassi, M.; Rosace, G.; et al. Inhibition of Human Topoisomerase II by N,N,N-Trimethylethanammonium Iodide Alkylcarbazole Derivatives. ChemMedChem 2018, 13, 2635–2643. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef]
- Mizutani, H.; Tada-Oikawa, S.; Hiraku, Y.; Oikawa, S.; Kojima, M.; Kawanishi, S. Mechanism of apoptosis induced by a new topoisomerase inhibitor through the generation of hydrogen peroxide. J. Biol. Chem. 2002, 277, 30684–30689. [Google Scholar] [CrossRef] [PubMed]
- Nandy, A.; Samanta, T.; Mallick, S.; Mitra, P.; Seth, S.K.; Saha, K.D.; Al-Deyabe, S.S.; Dinda, J. Synthesis of gold(III) ← gold(I)–NHC through disproportionation: The role of gold(i)–NHC in the induction of apoptosis in HepG2 cells. New J. Chem. 2016, 40, 6289–6298. [Google Scholar] [CrossRef]
- Nandy, A.; Dey, S.K.; Das, S.; Munda, R.N.; Dinda, J.; Saha, K.D. Gold (I) N-heterocyclic carbene complex inhibits mouse melanoma growth by p53 upregulation. Mol. Cancer 2014, 13, 57. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.; Casagrande, N.; Mariotto, E.; Baig, N.; Kawde, A.N.; Corona, G.; Larcher, R.; Borghese, C.; Pavan, C.; Seliman, A.A.; et al. Potent In Vitro and In Vivo Anticancer Activity of New Bipyridine and Bipyrimidine Gold (III) Dithiocarbamate Derivatives. Cancers 2019, 11, 474. [Google Scholar] [CrossRef]
- Magherini, F.; Fiaschi, T.; Valocchia, E.; Becatti, M.; Pratesi, A.; Marzo, T.; Massai, L.; Gabbiani, C.; Landini, I.; Nobili, S.; et al. Antiproliferative effects of two gold(I)-N-heterocyclic carbene complexes in A2780 human ovarian cancer cells: A comparative proteomic study. Oncotarget 2018, 9, 28042–28068. [Google Scholar] [CrossRef]
- Figueroa, D.; Asaduzzaman, M.; Young, F. Real time monitoring and quantification of reactive oxygen species in breast cancer cell line MCF-7 by 2′,7′-dichlorofluorescin diacetate (DCFDA) assay. J. Pharmacol. Toxicol. Methods 2018, 94, 26–33. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, X. Cytochrome C-mediated apoptosis. Annu Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Lobrich, M.; Jeggo, P.A. The impact of a negligent G2/M checkpoint on genomic instability and cancer induction. Nat. Rev. Cancer 2007, 7, 861–869. [Google Scholar] [CrossRef]
- Jackson, J.R.; Patrick, D.R.; Dar, M.M.; Huang, P.S. Targeted anti-mitotic therapies: Can we improve on tubulin agents? Nat. Rev. Cancer 2007, 7, 107–117. [Google Scholar] [CrossRef]
- Nitiss, J.L. Targeting DNA topoisomerase II in cancer chemotherapy. Nat. Rev. Cancer 2009, 9, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, E.; Cervinka, M. Topoisomerases and tubulin inhibitors: A promising combination for cancer treatment. Curr. Med. Chem. Anticancer. Agents 2003, 3, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Castelli, S.; Vassallo, O.; Katkar, P.; Che, C.M.; Sun, R.W.; Desideri, A. Inhibition of human DNA topoisomerase IB by a cyclometalated gold III compound: Analysis on the different steps of the enzyme catalytic cycle. Arch. Biochem. Biophys 2011, 516, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Katkar, P.; Wai-Yin Sun, R.; Desideri, A. Transition Metal Complex Gold (III) Inhibits Potential Anticancer Target Human DNA Topoisomerase IB. Int. J. Chem Phys. Sci. 2015, 4, 195–201. [Google Scholar]
- Chen, Z.F.; Shi, Y.F.; Liu, Y.C.; Hong, X.; Geng, B.; Peng, Y.; Liang, H. TCM active ingredient oxoglaucine metal complexes: Crystal structure, cytotoxicity, and interaction with DNA. Inorg. Chem. 2012, 51, 1998–2009. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.R.; Fagenson, A.M.; Ruangpradit, W.; Muller, M.T.; Munro, O.Q. Gold(III) complexes of pyridyl- and isoquinolylamido ligands: Structural, spectroscopic, and biological studies of a new class of dual topoisomerase I and II inhibitors. Inorg. Chem. 2013, 52, 7889–7906. [Google Scholar] [CrossRef]
- Akerman, K.J.; Fagenson, A.M.; Cyril, V.; Taylor, M.; Muller, M.T.; Akerman, M.P.; Munro, O.Q. Gold(III) macrocycles: Nucleotide-specific unconventional catalytic inhibitors of human topoisomerase I. J. Am. Chem. Soc. 2014, 136, 5670–5682. [Google Scholar] [CrossRef]
- Samia, L.B.P.; Parrilha, G.L.; Da Silva, J.G.; Ramos, J.P.; Souza-Fagundes, E.M.; Castelli, S.; Vutey, V.; Desideri, A.; Beraldo, H. Metal complexes of 3-(4-bromophenyl)-1-pyridin-2-ylprop-2-en-1-one thiosemicarbazone: Cytotoxic activity and investigation on the mode of action of the gold(III) complex. Biometals 2016, 29, 515–526. [Google Scholar] [CrossRef]
- Yan, J.J.; Chow, A.L.; Leung, C.H.; Sun, R.W.; Ma, D.L.; Che, C.M. Cyclometalated gold(III) complexes with N-heterocyclic carbene ligands as topoisomerase I poisons. Chem. Commun. (Camb) 2010, 46, 3893–3895. [Google Scholar] [CrossRef]
- Huang, X.; Huang, R.; Gou, S.; Wang, Z.; Liao, Z.; Wang, H. Platinum(IV) complexes conjugated with phenstatin analogue as inhibitors of microtubule polymerization and reverser of multidrug resistance. Bioorg. Med. Chem. 2017, 25, 4686–4700. [Google Scholar] [CrossRef] [PubMed]
- Acharya, S.; Maji, M.; Ruturaj; Purkait, K.; Gupta, A.; Mukherjee, A. Synthesis, Structure, Stability, and Inhibition of Tubulin Polymerization by Ru(II)-p-Cymene Complexes of Trimethoxyaniline-Based Schiff Bases. Inorg. Chem. 2019, 58, 9213–9224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Y.; Yi, Q.Y.; Wang, Y.J.; Du, F.; He, M.; Tang, B.; Wan, D.; Liu, Y.J.; Huang, H.L. Photoinduced anticancer activity studies of iridium(III) complexes targeting mitochondria and tubules. Eur. J. Med. Chem. 2018, 151, 568–584. [Google Scholar] [CrossRef]
- Waight, A.B.; Bargsten, K.; Doronina, S.; Steinmetz, M.O.; Sussman, D.; Prota, A.E. Structural Basis of Microtubule Destabilization by Potent Auristatin Anti-Mitotics. PLoS ONE 2016, 11, e0160890. [Google Scholar] [CrossRef] [PubMed]
- Redinbo, M.R.; Stewart, L.; Kuhn, P.; Champoux, J.J.; Hol, W.G. Crystal structures of human topoisomerase I in covalent and noncovalent complexes with DNA. Science 1998, 279, 1504–1513. [Google Scholar] [CrossRef]
- Schmidt, B.H.; Osheroff, N.; Berger, J.M. Structure of a topoisomerase II-DNA-nucleotide complex reveals a new control mechanism for ATPase activity. Nat. Struct Mol. Biol. 2012, 19, 1147–1154. [Google Scholar] [CrossRef]
- Abbassi, N.; Rakib el, M.; Chicha, H.; Bouissane, L.; Hannioui, A.; Aiello, C.; Gangemi, R.; Castagnola, P.; Rosano, C.; Viale, M. Synthesis and antitumor activity of some substituted indazole derivatives. Arch. Pharm. (Weinheim) 2014, 347, 423–431. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Sanner, M.F.; Duncan, B.S.; Carrillo, C.J.; Olson, A.J. Integrating computation and visualization for biomolecular analysis: An example using python and AVS. Pac. Symp. Biocomput 1999. [Google Scholar] [CrossRef]
- Cesarini, S.; Spallarossa, A.; Ranise, A.; Schenone, S.; Rosano, C.; La Colla, P.; Sanna, G.; Busonera, B.; Loddo, R. N-acylated and N,N′-diacylated imidazolidine-2-thione derivatives and N,N′-diacylated tetrahydropyrimidine-2(1H)-thione analogues: Synthesis and antiproliferative activity. Eur. J. Med. Chem. 2009, 44, 1106–1118. [Google Scholar] [CrossRef]
- Rosano, C.; Lappano, R.; Santolla, M.F.; Ponassi, M.; Donadini, A.; Maggiolini, M. Recent advances in the rationale design of GPER ligands. Curr. Med. Chem. 2012, 19, 6199–6206. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Chimento, A.; Sala, M.; Gomez-Monterrey, I.M.; Musella, S.; Bertamino, A.; Caruso, A.; Sinicropi, M.S.; Sirianni, R.; Puoci, F.; Parisi, O.I.; et al. Biological activity of 3-chloro-azetidin-2-one derivatives having interesting antiproliferative activity on human breast cancer cell lines. Bioorg. Med. Chem. Lett. 2013, 23, 6401–6405. [Google Scholar] [CrossRef] [PubMed]
- Sinicropi, M.S.; Caruso, A.; Conforti, F.; Marrelli, M.; El Kashef, H.; Lancelot, J.C.; Rault, S.; Statti, G.A.; Menichini, F. Synthesis, inhibition of NO production and antiproliferative activities of some indole derivatives. J. Enzyme Inhib. Med. Chem. 2009, 24, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Iacopetta, D.; Sinicropi, M.S.; Bonesi, M.; Leporini, M.; Passalacqua, N.G.; Ceramella, J.; Menichini, F.; Loizzo, M.R. Assessment of antioxidant, antitumor and pro-apoptotic effects of Salvia fruticosa Mill. subsp. thomasii (Lacaita) Brullo, Guglielmo, Pavone & Terrasi (Lamiaceae). Food Chem. Toxicol. 2017, 106, 155–164. [Google Scholar]
- Iacopetta, D.; Carocci, A.; Sinicropi, M.S.; Catalano, A.; Lentini, G.; Ceramella, J.; Curcio, R.; Caroleo, M.C. Old Drug Scaffold, New Activity: Thalidomide-Correlated Compounds Exert Different Effects on Breast Cancer Cell Growth and Progression. ChemMedChem 2017, 12, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Iacopetta, D.; Lappano, R.; Cappello, A.R.; Madeo, M.; De Francesco, E.M.; Santoro, A.; Curcio, R.; Capobianco, L.; Pezzi, V.; Maggiolini, M.; et al. SLC37A1 gene expression is up-regulated by epidermal growth factor in breast cancer cells. Breast Cancer Res. Treat. 2010, 122, 755–764. [Google Scholar] [CrossRef][Green Version]
- Chimento, A.; Saturnino, C.; Iacopetta, D.; Mazzotta, R.; Caruso, A.; Plutino, M.R.; Mariconda, A.; Ramunno, A.; Sinicropi, M.S.; Pezzi, V.; et al. Inhibition of human topoisomerase I and II and anti-proliferative effects on MCF-7 cells by new titanocene complexes. Bioorg Med. Chem. 2015, 23, 7302–7312. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
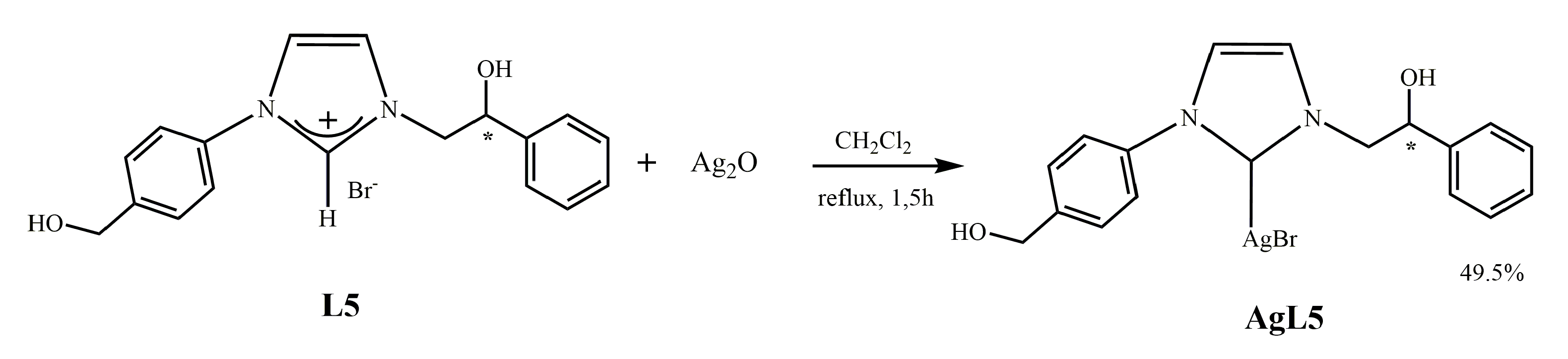{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µM) | ||||||
|---|---|---|---|---|---|---|
| Compounds | MCF-7 | MDA-MB-231 | HeLa | ISHIKAWA | MCF-10A | Hek-293 |
| L5 | >200 | >200 | >200 | >200 | >200 | >200 |
| AgL5 | >200 | >200 | >200 | >200 | >200 | >200 |
| AuL5 | 18.2 ± 0.5 | 31.7 ± 0.8 | 44.5 ± 0.9 | 54.9 ± 1.0 | >200 | >200 |
| L6 | >200 | >200 | >200 | >200 | >200 | >200 |
| AgL6 | 29.7 ± 0.3 | 57.9 ± 0.5 | 47.7 ± 0.4 | 26.7 ± 0.5 | 143.4 ± 0.8 | >200 |
| AuL6 | 9.38 ± 0.2 | 53.7 ± 0.7 | 98.6 ± 1.2 | 12.6 ± 0.3 | 158.8 ± 1.0 | >200 |
| L7 | >200 | >200 | >200 | >200 | >200 | >200 |
| AgL7 | 10.5 ± 0.8 | 3.22 ± 1.2 | 9.76 ± 0.6 | 6.55 ± 0.9 | >200 | >200 |
| AuL7 | 5.18 ± 0.4 | 2.1 ± 0.7 | 31.9 ± 0.5 | 29.9 ± 0.9 | >200 | 65.0 ± 0.7 |
| AuL7 + Quercetin (4 h) | - | - | - | - | - | 96.9 ± 1.1 |
| Cis-platin | 35.8 ± 1.3 | 28.7 ± 1.0 | 15.7 ± 1.1 | 15.1 ± 0.8 | 81.3 ± 1.2 | 16.8 ± 0.4 |
| Compounds | Tubulin | Topoisomerase I | Topoisomerase II | |||
|---|---|---|---|---|---|---|
| Eb (kcal/mol) | Ki (μM) | Eb (kcal/mol) | Ki (μM) | Eb (kcal/mol) | Ki (μM) | |
| AuL4 | −7.4 | 3.78 | −5.83 | 53.0 | −6.39 | 20.86 |
| AgL5 | −7.66 | 2.43 | −7.44 | 3.51 | −7.72 | 2.21 |
| AuL5 | −8.45 | 0.6629 | −7.55 | 2.92 | −7.49 | 3.03 |
| AgL6 | −7.73 | 2.17 | −7.73 | 2.17 | −6.44 | 18.91 |
| AuL6 | −6.76 | 8.26 | −8.9 | 0.3219 | −7.84 | 4.16 |
| AgL7 | −13.27 | 1.3 × 10−6 | −9.18 | 0.1853 | −10.06 | 0.0423 |
| AuL7 | −11.3 | 0.0052 | −8.42 | 0.6694 | −6.84 | 9.64 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iacopetta, D.; Rosano, C.; Sirignano, M.; Mariconda, A.; Ceramella, J.; Ponassi, M.; Saturnino, C.; Sinicropi, M.S.; Longo, P. Is the Way to Fight Cancer Paved with Gold? Metal-Based Carbene Complexes with Multiple and Fascinating Biological Features. Pharmaceuticals 2020, 13, 91. https://doi.org/10.3390/ph13050091
Iacopetta D, Rosano C, Sirignano M, Mariconda A, Ceramella J, Ponassi M, Saturnino C, Sinicropi MS, Longo P. Is the Way to Fight Cancer Paved with Gold? Metal-Based Carbene Complexes with Multiple and Fascinating Biological Features. Pharmaceuticals. 2020; 13(5):91. https://doi.org/10.3390/ph13050091
Chicago/Turabian StyleIacopetta, Domenico, Camillo Rosano, Marco Sirignano, Annaluisa Mariconda, Jessica Ceramella, Marco Ponassi, Carmela Saturnino, Maria Stefania Sinicropi, and Pasquale Longo. 2020. "Is the Way to Fight Cancer Paved with Gold? Metal-Based Carbene Complexes with Multiple and Fascinating Biological Features" Pharmaceuticals 13, no. 5: 91. https://doi.org/10.3390/ph13050091
APA StyleIacopetta, D., Rosano, C., Sirignano, M., Mariconda, A., Ceramella, J., Ponassi, M., Saturnino, C., Sinicropi, M. S., & Longo, P. (2020). Is the Way to Fight Cancer Paved with Gold? Metal-Based Carbene Complexes with Multiple and Fascinating Biological Features. Pharmaceuticals, 13(5), 91. https://doi.org/10.3390/ph13050091