Improved Surface Display of Human Hyal1 and Identification of Testosterone Propionate and Chicoric Acid as New Inhibitors




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
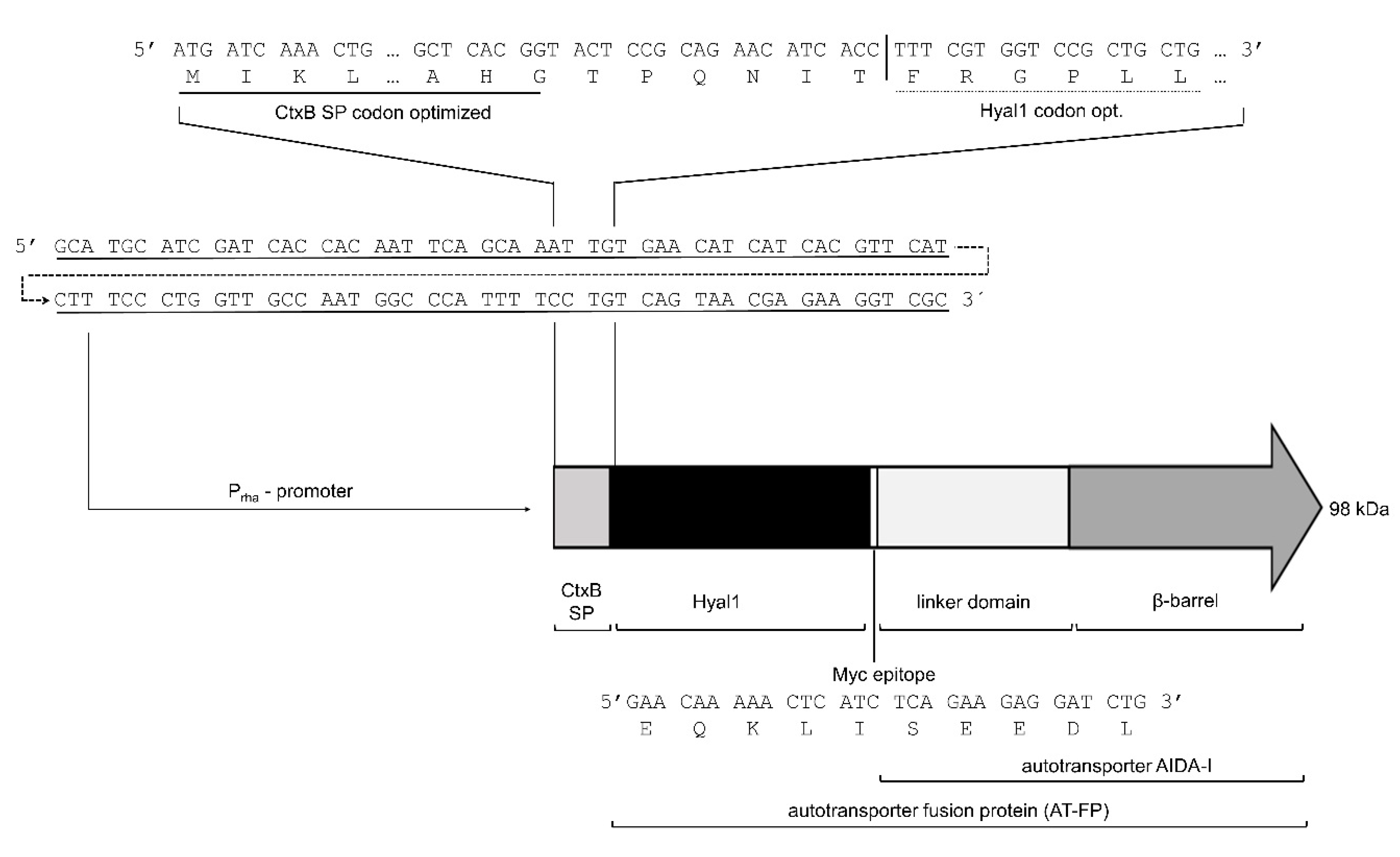
2.1. Surface Expression of Hyal1
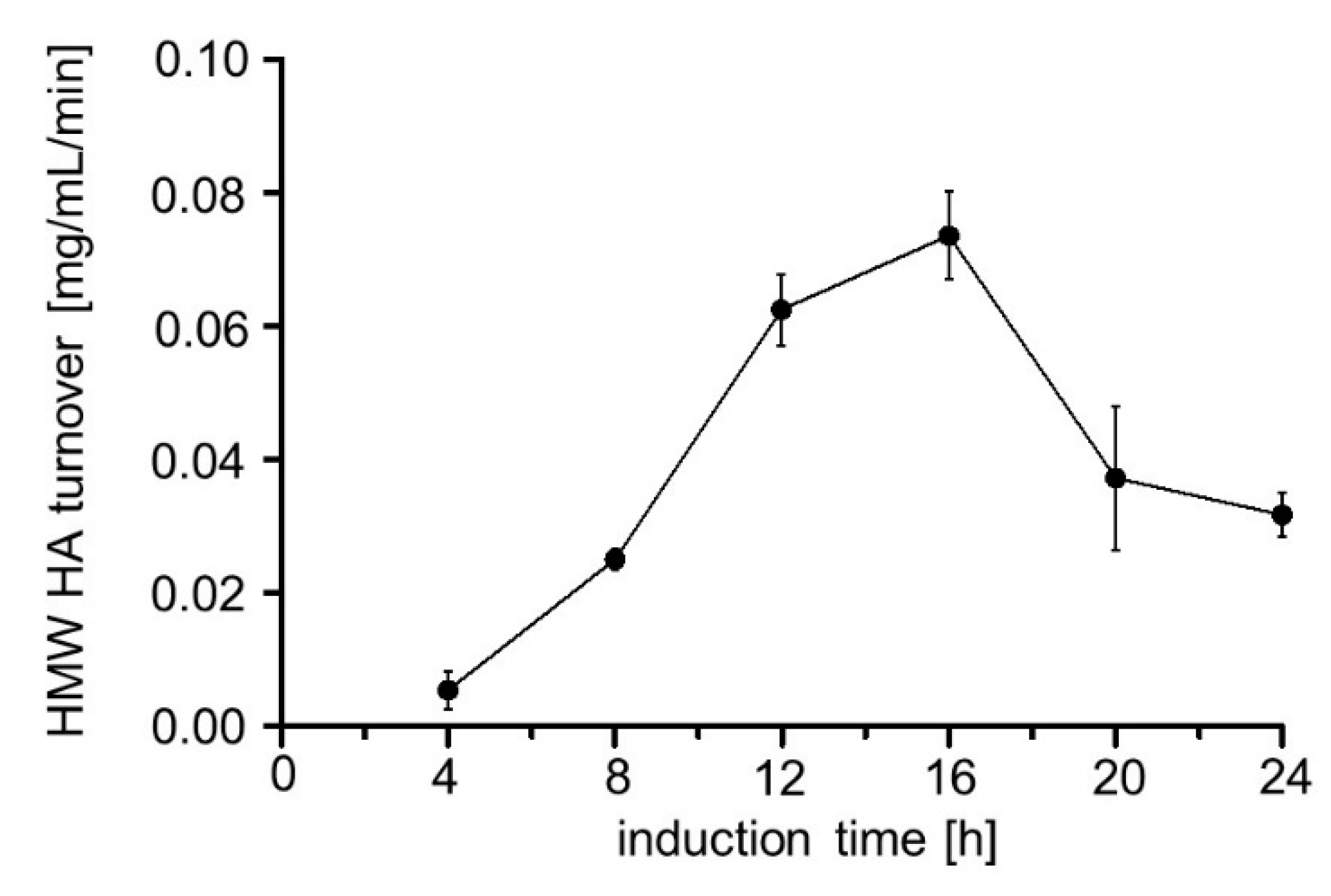
2.2. Influence of Induction Time on Hyal1 Expression and Activity
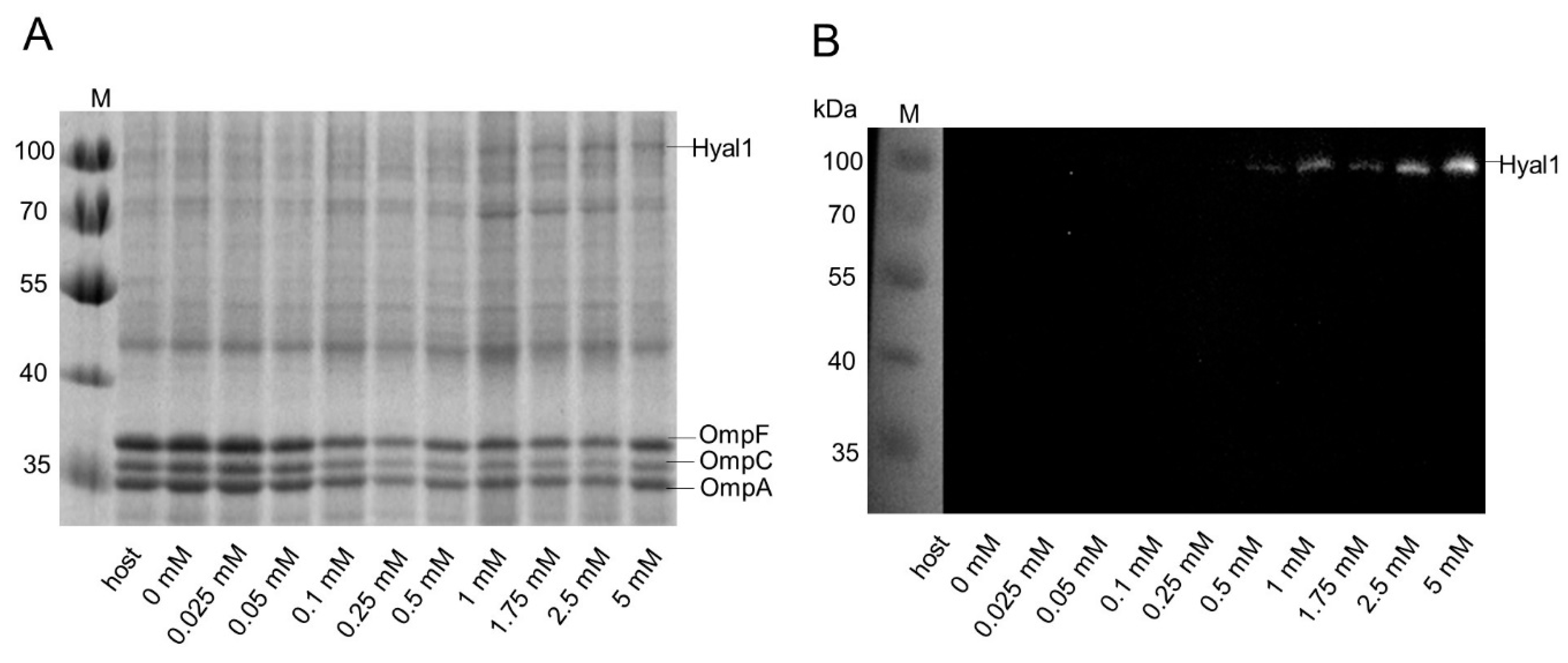
2.3. Influence of Rhamnose Concentration on Hyal1 Expression and Activity
2.4. Influence of pH, Temperature, Substrate Concentration and Sodium Chloride on Enzyme Activity
2.5. Reaction Time
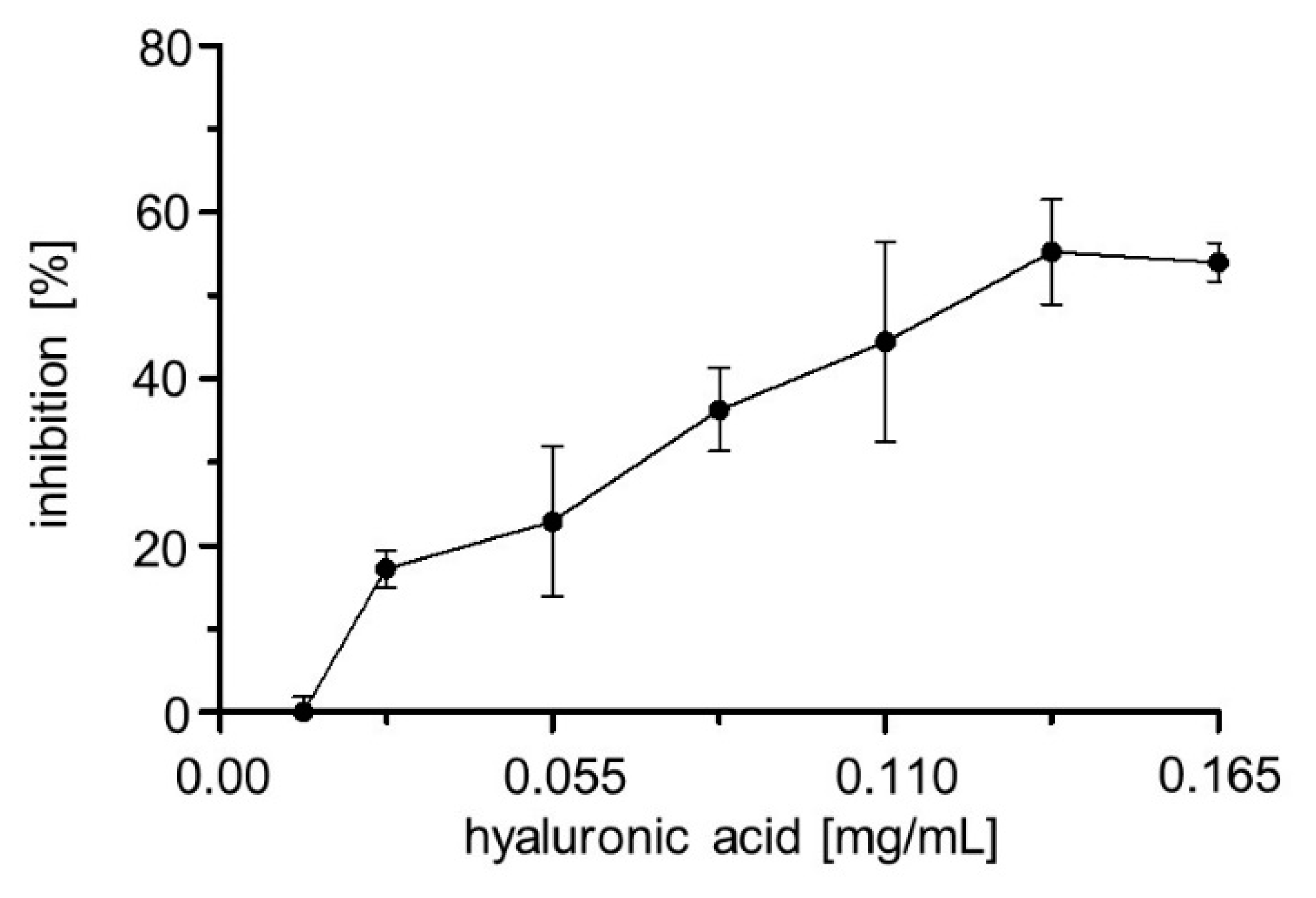
2.6. Inhibitor Screening
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Plasmid Construction
5.3. Escherichia coli F470 Culturing
5.4. Preparation of Outer Membrane Proteins
5.5. SDS-PAGE and Western Blot Analysis
5.6. Flow Cytometry Analysis
5.7. Stains-All Assay
5.8. In Silico Experiments
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cowman, M.K.; Schmidt, T.A.; Raghavan, P.; Stecco, A. Viscoelastic Properties of Hyaluronan in Physiological Conditions. F1000Research 2015, 4, 622. [Google Scholar] [CrossRef]
- Tammi, M.I.; Day, A.J.; Turley, E.A. Hyaluronan and homeostasis: A balancing act. J. Biol. Chem. 2002, 277, 4581–4584. [Google Scholar] [CrossRef] [PubMed]
- Monslow, J.; Govindaraju, P.; Pure, E. Hyaluronan—A functional and structural sweet spot in the tissue microenvironment. Front. Immunol. 2015, 6, 231. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, P.; Kamal, R. Hyaluronic Acid: A Boon in Periodontal Therapy. N. Am. J. Med. Sci. 2013, 5, 309–315. [Google Scholar] [CrossRef] [PubMed]
- El-Safory, N.S.; Fazary, A.E.; Lee, C.K. Hyaluronidases, a group of glycosidases: Current and future perspectives. Carbohydr. Polym. 2010, 81, 165–181. [Google Scholar] [CrossRef]
- Noble, P.W. Hyaluronan and its catabolic products in tissue injury and repair. Matrix Biol. 2002, 21, 25–29. [Google Scholar] [CrossRef]
- Csoka, A.B.; Scherer, S.W.; Stern, R. Expression analysis of six paralogous human hyaluronidase genes clustered on chromosomes 3p21 and 7q31. Genomics 1999, 60, 356–361. [Google Scholar] [CrossRef]
- Csoka, A.B.; Frost, G.I.; Stern, R. The six hyaluronidase-like genes in the human and mouse genomes. Matrix Biol. 2001, 20, 499–508. [Google Scholar] [CrossRef]
- Stern, R.; Jedrzejas, M.J. Hyaluronidases: Their genomics, structures, and mechanisms of action. Chem. Rev. 2006, 106, 818–839. [Google Scholar] [CrossRef]
- Chi, A.; Shirodkar, S.P.; Escudero, D.O.; Ekwenna, O.O.; Yates, T.J.; Ayyathurai, R.; Garcia-Roig, M.; Gahan, J.C.; Manoharan, M.; Bird, V.G.; et al. Molecular characterization of kidney cancer Association of Hyaluronic Acid Family With Histological Subtypes and Metastasis. Cancer 2012, 118, 2394–2402. [Google Scholar] [CrossRef]
- Myles, D.G.; Primakoff, P. Why did the sperm cross the cumulus? To get to the oocyte. Functions of the sperm surface proteins PH-20 and fertilin in arriving at, and fusing with, the egg. Biol. Reprod. 1997, 56, 320–327. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lokeshwar, V.B.; Lokeshwar, B.L.; Pham, H.T.; Block, N.L. Association of elevated levels of hyaluronidase, a matrix-degrading enzyme, with prostate cancer progression. Cancer Res. 1996, 56, 651–657. [Google Scholar] [PubMed]
- Lokeshwar, V.B.; Obek, C.; Pham, H.T.; Wei, D.; Young, M.J.; Duncan, R.C.; Soloway, M.S.; Block, N.L. Urinary hyaluronic acid and hyaluronidase: Markers for bladder cancer detection and evaluation of grade. J. Urol. 2000, 163, 348–356. [Google Scholar] [CrossRef]
- Madan, A.K.; Pang, Y.; Wilkiemeyer, M.B.; Yu, D.; Beech, D.J. Increased hyaluronidase expression in more aggressive prostate adenocarcinoma. Oncol. Rep. 1999, 6, 1431–1433. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.X.; Wang, X.Y.; Li, H.Y.; Su, X.L.; Wang, L.A.; Ran, L.A.; Zheng, K.; Ren, G.S. HYAL1 overexpression is correlated with the malignant behavior of human breast cancer. Int. J. Cancer 2011, 128, 1303–1315. [Google Scholar] [CrossRef]
- Kramer, M.W.; Golshani, R.; Merseburger, A.S.; Knapp, J.; Garcia, A.; Hennenlotter, J.; Duncan, R.C.; Soloway, M.S.; Jorda, M.; Kuczyk, M.A.; et al. HYAL-1 Hyaluronidase: A Potential Prognostic Indicator for Progression to Muscle Invasion and Recurrence in Bladder Cancer. Eur. Urol. 2010, 57, 86–93. [Google Scholar] [CrossRef]
- Neumann, A.; Schinzel, R.; Palm, D.; Riederer, P.; Munch, G. High molecular weight hyaluronic acid inhibits advanced glycation endproduct-induced NF-kappa B activation and cytokine expression. FEBS Lett. 1999, 453, 283–287. [Google Scholar] [CrossRef]
- Jentsch, H.; Pomowski, R.; Kundt, G.; Gocke, R. Treatment of gingivitis with hyaluronan. J. Clin. Periodontol. 2003, 30, 159–164. [Google Scholar] [CrossRef]
- Rayahin, J.E.; Buhrman, J.S.; Zhang, Y.; Koh, T.J.; Gemeinhart, R.A. High and Low Molecular Weight Hyaluronic Acid Differentially Influence Macrophage Activation. ACS Biomater. Sci. Eng. 2015, 1, 481–493. [Google Scholar] [CrossRef]
- Salmen, S.; Hoechstetter, J.; Kasbauer, C.; Paper, D.H.; Bernhardt, G.; Buschauer, A. Sulphated oligosaccharides as inhibitors of hyaluronidases from bovine testis, bee venom and Streptococcus agalactiae. Planta Med. 2005, 71, 727–732. [Google Scholar] [CrossRef]
- Kuppusamy, U.R.; Khoo, H.E.; Das, N.P. Structure-Activity Studies of Flavonoids as Inhibitors of Hyaluronidase. Biochem. Pharm. 1990, 40, 397–401. [Google Scholar] [CrossRef]
- Furuya, T.; Yamagata, S.; Shimoyama, Y.; Fujihara, M.; Morishima, N.; Ohtsuki, K. Biochemical characterization of glycyrrhizin as an effective inhibitor for hyaluronidases from bovine testis. Biol. Pharm. Bull. 1997, 20, 973–977. [Google Scholar] [CrossRef]
- Botzki, A.; Rigden, D.J.; Braun, S.; Nukui, M.; Salmen, S.; Hoechstetter, J.; Bernhardt, G.; Dove, S.; Jedrzejas, M.J.; Buschauer, A. l-Ascorbic Acid 6-Hexadecanoate, a Potent Hyaluronidase Inhibitor: X-RAY structure and molecular modeling of enzyme-inhibitor complexes. J. Biol. Chem. 2004, 279, 45990–45997. [Google Scholar] [CrossRef]
- Isoyama, T.; Thwaites, D.; Selzer, M.G.; Carey, R.I.; Barbucci, R.; Lokeshwar, V.B. Differential selectivity of hyaluronidase inhibitors toward acidic and basic hyaluronidases. Glycobiology 2006, 16, 11–21. [Google Scholar] [CrossRef]
- Afify, A.M.; Stern, M.; Guntenhoner, M.; Stern, R. Purification and Characterization of Human Serum Hyaluronidase. Arch. Biochem. Biophys. 1993, 305, 434–441. [Google Scholar] [CrossRef]
- Csoka, T.B.; Frost, G.I.; Wong, T.; Stern, R. Purification and microsequencing of hyaluronidase isozymes from human urine. FEBS Lett. 1997, 417, 307–310. [Google Scholar] [CrossRef]
- Frost, G.I.; Csoka, T.B.; Wong, T.; Stern, R. Purification, cloning, and expression of human plasma hyaluronidase. Biochem. Biophys. Res. Commun. 1997, 236, 10–15. [Google Scholar] [CrossRef]
- Hofinger, E.S.A.; Spickenreither, M.; Oschmann, J.; Bernhardt, G.; Rudolph, R.; Buschauer, A. Recombinant human hyaluronidase Hyal-1: Insect cells versus Escherichia coli as expression system and identification of low molecular weight inhibitors. Glycobiology 2007, 17, 444–453. [Google Scholar] [CrossRef]
- Orlando, Z.; Lengers, I.; Melzig, M.; Buschauer, A.; Hensel, A.; Jose, J. Autodisplay of Human Hyaluronidase Hyal-1 on Escherichia coli and Identification of Plant-Derived Enzyme Inhibitors. Molecules 2015, 20, 15449–15468. [Google Scholar] [CrossRef]
- Sendker, J.; Böker, I.; Lengers, I.; Brandt, S.; Jose, J.; Stark, T.; Hofmann, T.; Fink, C.; Abdel-Aziz, H.; Hensel, A. Phytochemical Characterization of Low Molecular Weight Constituents from Marshmallow Roots (Althaea officinalis) and Inhibiting Effects of the Aqueous Extract on Human Hyaluronidase-1. J. Nat. Prod. 2017, 80, 290–297. [Google Scholar] [CrossRef]
- Giacalone, M.J.; Gentile, A.M.; Lovitt, B.T.; Berkley, N.L.; Gunderson, C.W.; Surber, M.W. Toxic protein expression in Escherichia coli using a rhamnose-based tightly regulated and tunable promoter system. Biotechniques 2006, 40, 355–364. [Google Scholar] [CrossRef]
- Berrow, N.S.; Alderton, D.; Sainsbury, S.; Nettleship, J.; Assenberg, R.; Rahman, N.; Stuart, D.I.; Owens, R.J. A versatile ligation-independent cloning method suitable for high-throughput expression screening applications. Nucleic Acids Res. 2007, 35, e45. [Google Scholar] [CrossRef]
- Quehl, P.; Hollender, J.; Schuurmann, J.; Brossette, T.; Maas, R.; Jose, J. Co-expression of active human cytochrome P450 1A2 and cytochrome P450 reductase on the cell surface of Escherichia coli. Microb. Cell Factories 2016, 15, 26. [Google Scholar] [CrossRef]
- Park, M.; Yoo, G.; Bong, J.-H.; Jose, J.; Kang, M.-J.; Pyun, J.-C. Isolation and characterization of the outer membrane of Escherichia coli with autodisplayed Z-domains. Biochim. Biophys. Acta 2015, 1848, 842–847. [Google Scholar] [CrossRef]
- Yethon, J.A.; Vinogradov, E.; Perry, M.B.; Whitfield, C. Mutation of the lipopolysaccharide core glycosyltransferase encoded by waaG destabilizes the outer membrane of Escherichia coli by interfering with core phosphorylation. J. Bacteriol. 2000, 182, 5620–5623. [Google Scholar] [CrossRef]
- Vandeputte-Rutten, L.; Kramer, R.A.; Kroon, J.; Dekker, N.; Egmond, M.R.; Gros, P. Crystal structure of the outer membrane protease OmpT from Escherichia coli suggests a novel catalytic site. EMBO J. 2001, 20, 5033–5039. [Google Scholar] [CrossRef]
- Maurer, J.; Jose, J.; Meyer, T.F. Autodisplay: One-component system for efficient surface display and release of soluble recombinant proteins from Escherichia coli. J. Bacteriol. 1997, 179, 794–804. [Google Scholar] [CrossRef]
- Baneyx, F.; Georgiou, G. Invivo Degradation of Secreted Fusion Proteins by the Escherichia-Coli Outer-Membrane Protease Ompt. J. Bacteriol. 1990, 172, 491–494. [Google Scholar] [CrossRef]
- Sichwart, S.; Tozakidis, I.E.P.; Teese, M.; Jose, J. Maximized Autotransporter-Mediated Expression (MATE) for Surface Display and Secretion of Recombinant Proteins in Escherichia coli. Food Technol. Biotechnol. 2015, 53, 251–260. [Google Scholar] [CrossRef]
- Jose, J.; Bernhardt, R.; Hannemann, F. Cellular surface display of dimeric Adx and whole cell P450-mediated steroid synthesis on E.coli. J. Biotechnol 2002, 95, 257–268. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.e.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Homer, K.A.; Denbow, L.; Whiley, R.A.; Beighton, D. Chondroitin sulfate depolymerase and hyaluronidase activities of viridans streptococci determined by a sensitive spectrophotometric assay. J. Clin. Microbiol. 1993, 31, 1648–1651. [Google Scholar] [CrossRef]
- Schlegel, S.; Lofblom, J.; Lee, C.; Hjelm, A.; Klepsch, M.; Strous, M.; Drew, D.; Slotboom, D.J.; de Gier, J.W. Optimizing Membrane Protein Overexpression in the Escherichia coli strain Lemo21(DE3). J. Mol. Biol. 2012, 423, 648–659. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Klagas, I.; Karakiulakis, G.; Hostettler, K.; S’ng, C.T.; Kotoula, V.; Savic, S.; Tamm, M.; Roth, M. Steroids and beta(2)-Agonists Regulate Hyaluronan Metabolism in Asthmatic Airway Smooth Muscle Cells. Am. J. Respir. Cell Mol. 2012, 47, 759–767. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Klagas, I.; Karakiulakis, G.; Tamm, M.; Roth, M.; Stolz, D. Glucocorticoids and β2-agonists regulate the pathologic metabolism of hyaluronic acid in COPD. Pulm. Pharmacol. Ther. 2018, 48, 104–110. [Google Scholar] [CrossRef]
- Raheem, K.A.; Marei, W.F.; Mifsud, K.; Khalid, M.; Wathes, D.C.; Fouladi-Nashta, A.A. Regulation of the hyaluronan system in ovine endometrium by ovarian steroids. Reproduction 2013, 145, 491–504. [Google Scholar] [CrossRef][Green Version]
- Tanyildizi, S.; Bozkurt, T. Effects of progesterone and testosterone on the hyaluronidase activities and sperm characteristics in sheep. Turk. J. Vet. Anim. Sci. 2002, 26, 1137–1143. [Google Scholar]
- Desalegu, M.; Plonska, H.; Pigman, W. A Comparison of Serum and Testicular Hyaluronidase. Arch. Biochem. Biophys. 1967, 121, 548. [Google Scholar] [CrossRef]
- Cramer, J.A.; Bailey, L.C.; Bailey, C.A.; Miller, R.T. Kinetic and Mechanistic Studies with Bovine Testicular Hyaluronidase. Biochim. Biophys. Acta 1994, 1200, 315–321. [Google Scholar] [CrossRef]
- Barsukov, A.K.; Kozhevnikova, O.V.; Khokhryakova, A.V. Isolation and purification of bovine testicular hyaluronidase. Appl. Biochem. Microbiol. 2003, 39, 549–552. [Google Scholar] [CrossRef]
- Kaya, M.O.; Arslan, O.; Guler, O.O. A new affinity method for purification of bovine testicular hyaluronidase enzyme and an investigation of the effects of some compounds on this enzyme. J. Enzym. Inhib. Med. Chem. 2015, 30, 524–527. [Google Scholar] [CrossRef]
- Gold, E.W. Purification and Properties of Hyaluronidase from Human-Liver—Differences from and Similarities to the Testicular Enzyme. Biochem. J. 1982, 205, 69–74. [Google Scholar] [CrossRef]
- Wolf, R.A.; Glogar, D.; Chaung, L.Y.; Garrett, P.E.; Ertl, G.; Tumas, J.; Braunwald, E.; Kloner, R.A.; Feldstein, M.L.; Muller, J.E. Heparin Inhibits Bovine Testicular Hyaluronidase Activity in Myocardium of Dogs with Coronary-Artery Occlusion. Am. J. Cardiol. 1984, 53, 941–944. [Google Scholar] [CrossRef]
- Guerra, F. Hyaluronidase Inhibition by Sodium Salicylate in Rheumatic Fever. Science 1946, 103, 686–687. [Google Scholar] [CrossRef] [PubMed]
- Kaessler, A.; Nourrisson, M.R.; Duflos, M.; Jose, J. Indole carboxamides inhibit bovine testes hyaluronidase at pH 7.0 and indole acetamides activate the enzyme at pH 3.5 by different mechanisms. J. Enzym. Inhib. Med. Chem. 2008, 23, 719–727. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hertel, W.; Peschel, G.; Ozegowski, J.H.; Willer, P.J. Inhibitory effects of triterpenes and flavonoids on the enzymatic activity of hyaluronic acid-splitting enzymes. Arch. Der Pharm. 2006, 339, 313–318. [Google Scholar] [CrossRef]
- Harunari, E.; Imada, C.; Igarashi, Y.; Fukuda, T.; Terahara, T.; Kobayashi, T. Hyaluromycin, a New Hyaluronidase Inhibitor of Polyketide Origin from Marine Streptomyces sp. Mar. Drugs 2014, 12, 491–507. [Google Scholar] [CrossRef]
- Kohi, S.; Sato, N.; Koga, A.; Hirata, K.; Harunari, E.; Igarashi, Y. Hyaluromycin, a Novel Hyaluronidase Inhibitor, Attenuates Pancreatic Cancer Cell Migration and Proliferation. J. Oncol. 2016, 2016, 9063087. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Bonovolias, I.; Karakioulakis, G.; Tamm, M.; Louis, R.; Milenkovic, B.; Boersma, W.; Kostikas, K.; Blasi, F.; Aerts, J.; et al. Serum hyaluronidase-1 is increased in COPD exacerbations and is associated with COPD severity and outcomes. Eur. Respir. J. 2018, 52, PA4969. [Google Scholar] [CrossRef]
- Kakegawa, H.; Mitsuo, N.; Matsumoto, H.; Satoh, T.; Akagi, M.; Tasaka, K. Hyaluronidase-Inhibitory and Anti-Allergic Activities of the Photo-Irradiated Products of Tranilast. Chem. Pharm. Bull. 1985, 33, 3738–3744. [Google Scholar] [CrossRef]
- Halgren, T.A. Merck molecular force field. 1. Basis, form, scope, parameterization, and performance of MMFF94. J. Comput Chem 1996, 17, 490–519. [Google Scholar] [CrossRef]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lengers, I.; Herrmann, F.; Le Borgne, M.; Jose, J. Improved Surface Display of Human Hyal1 and Identification of Testosterone Propionate and Chicoric Acid as New Inhibitors. Pharmaceuticals 2020, 13, 54. https://doi.org/10.3390/ph13040054
Lengers I, Herrmann F, Le Borgne M, Jose J. Improved Surface Display of Human Hyal1 and Identification of Testosterone Propionate and Chicoric Acid as New Inhibitors. Pharmaceuticals. 2020; 13(4):54. https://doi.org/10.3390/ph13040054
Chicago/Turabian StyleLengers, Isabelle, Fabian Herrmann, Marc Le Borgne, and Joachim Jose. 2020. "Improved Surface Display of Human Hyal1 and Identification of Testosterone Propionate and Chicoric Acid as New Inhibitors" Pharmaceuticals 13, no. 4: 54. https://doi.org/10.3390/ph13040054
APA StyleLengers, I., Herrmann, F., Le Borgne, M., & Jose, J. (2020). Improved Surface Display of Human Hyal1 and Identification of Testosterone Propionate and Chicoric Acid as New Inhibitors. Pharmaceuticals, 13(4), 54. https://doi.org/10.3390/ph13040054