ASA Suppresses PGE2 in Plasma and Melanocytic Nevi of Human Subjects at Increased Risk for Melanoma

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
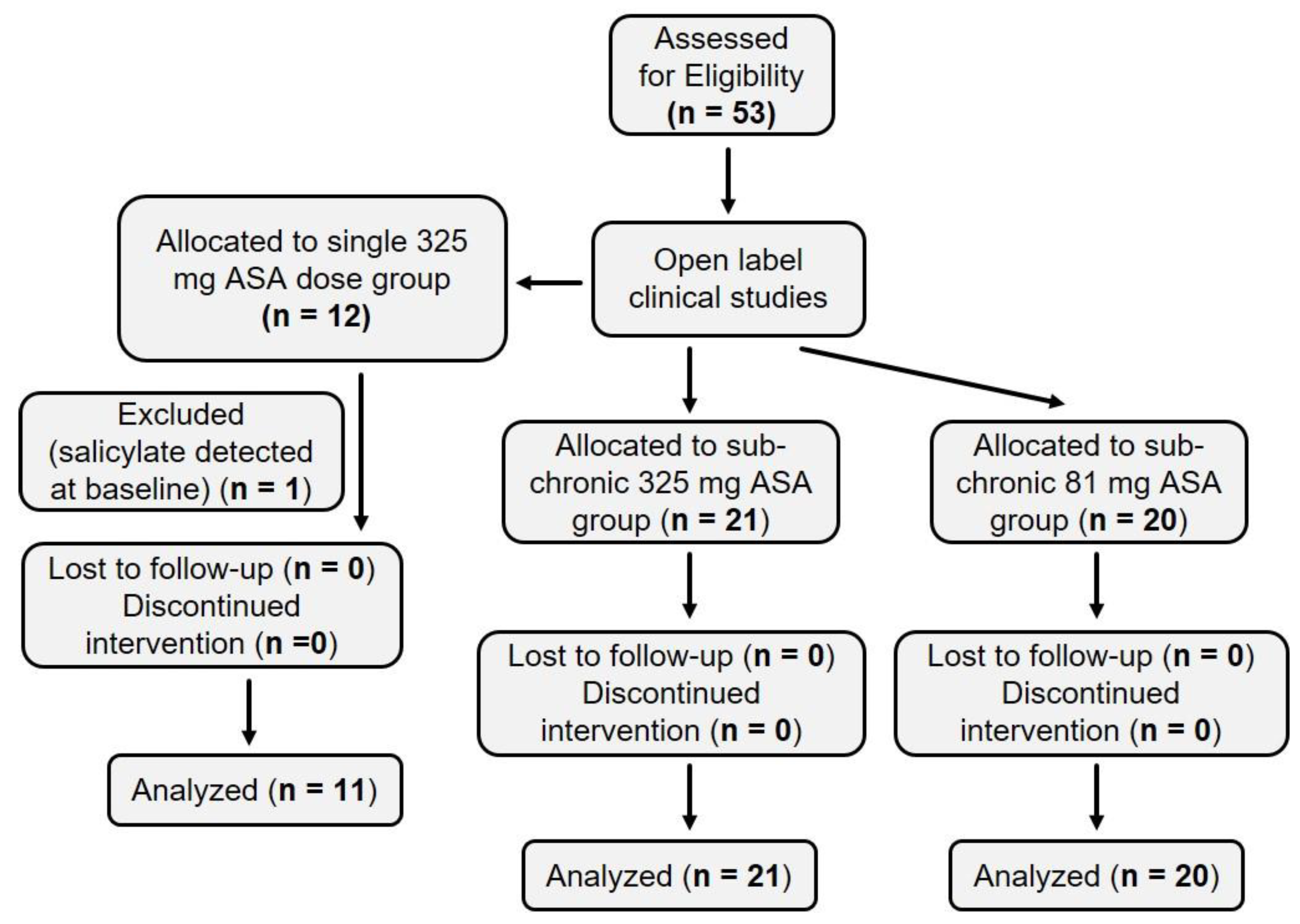
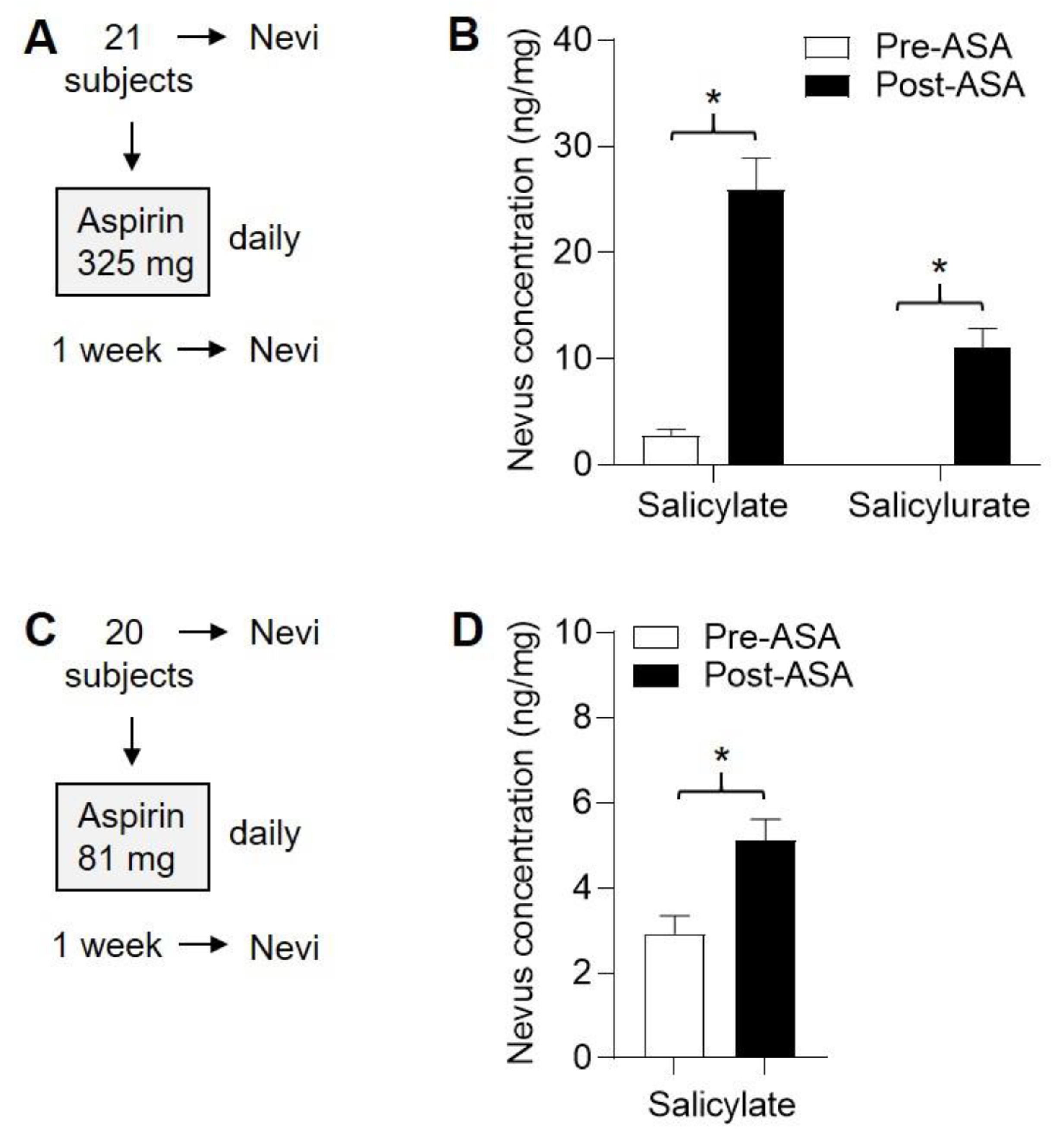
2.1. Summary of Human Subject Enrollment
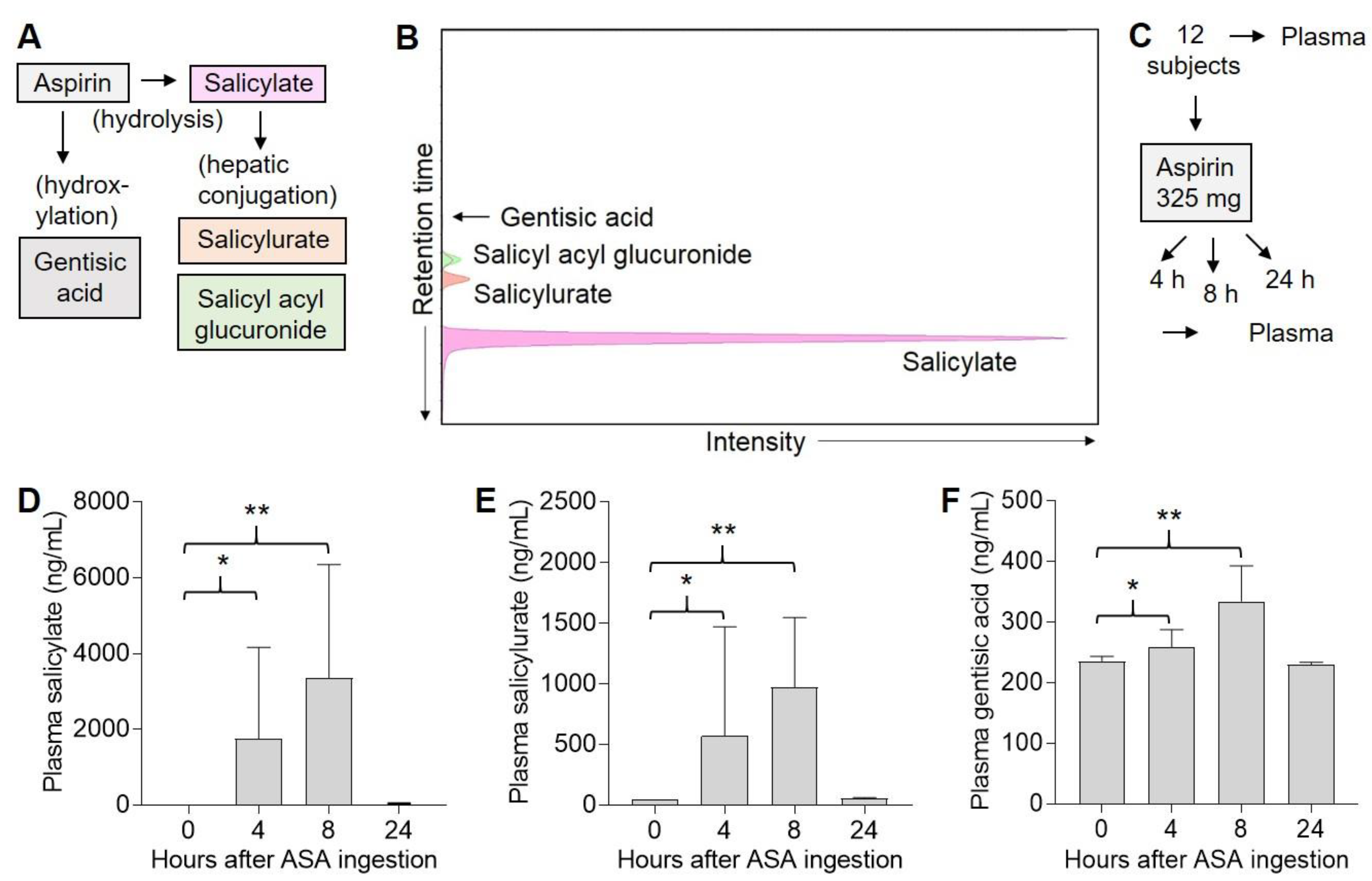
2.2. Three ASA Metabolites Detected in Plasma Following Single Dose of ASA
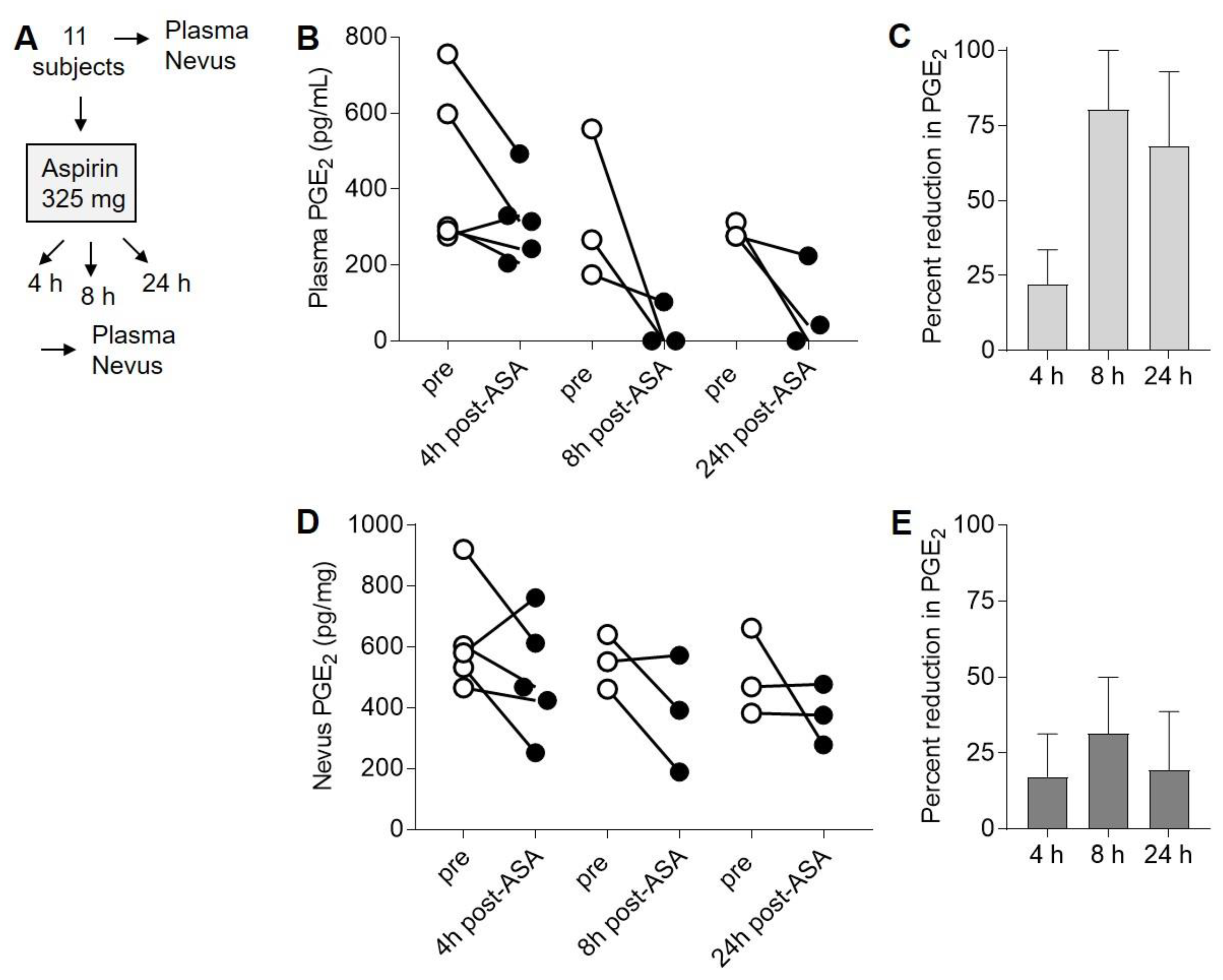
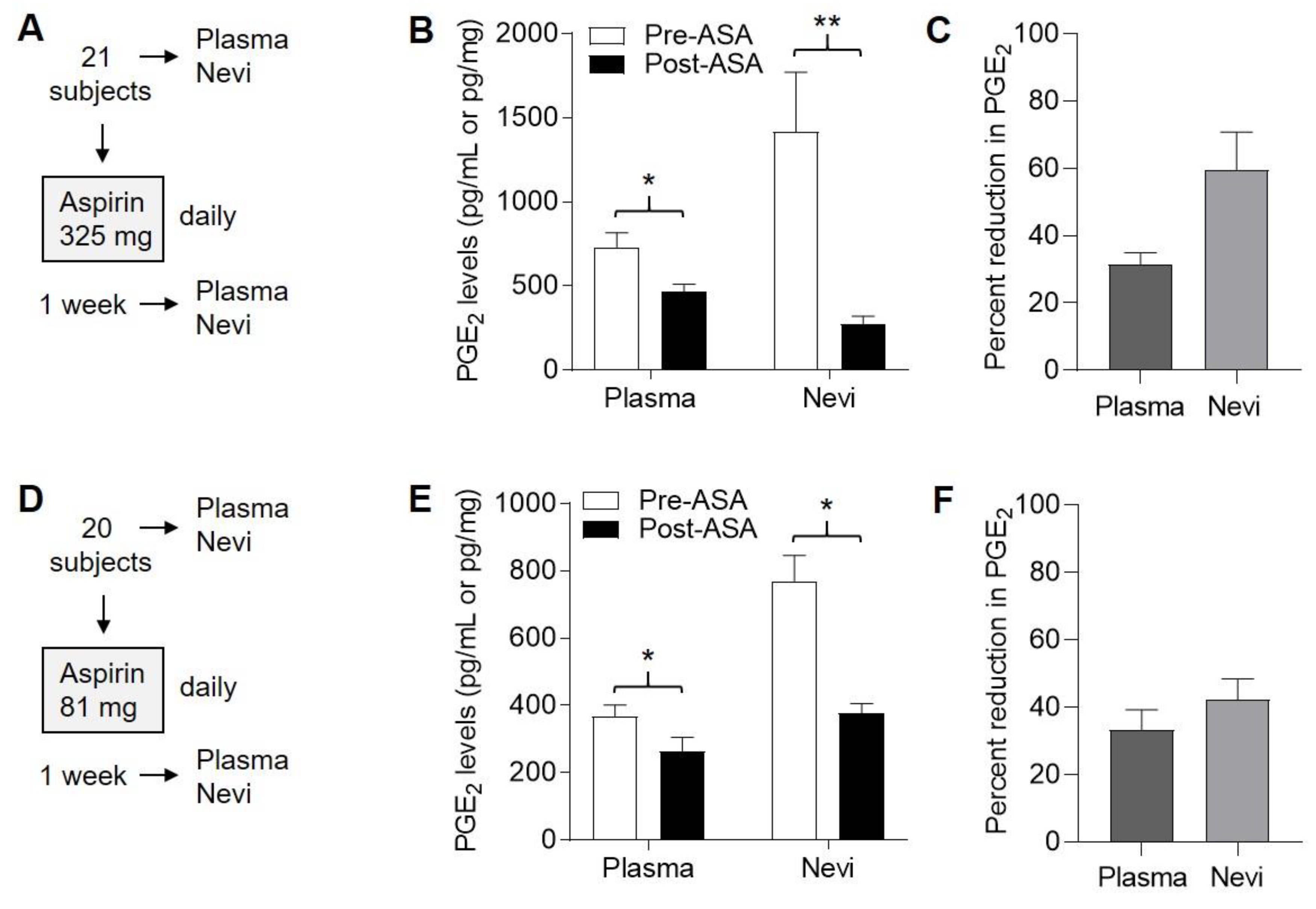
2.3. Suppression of PGE2 in Plasma and Nevi Following Single Dose of ASA
2.4. ASA Suppresses PGE2 in Nevi Expressing Mutant BRAFV600E
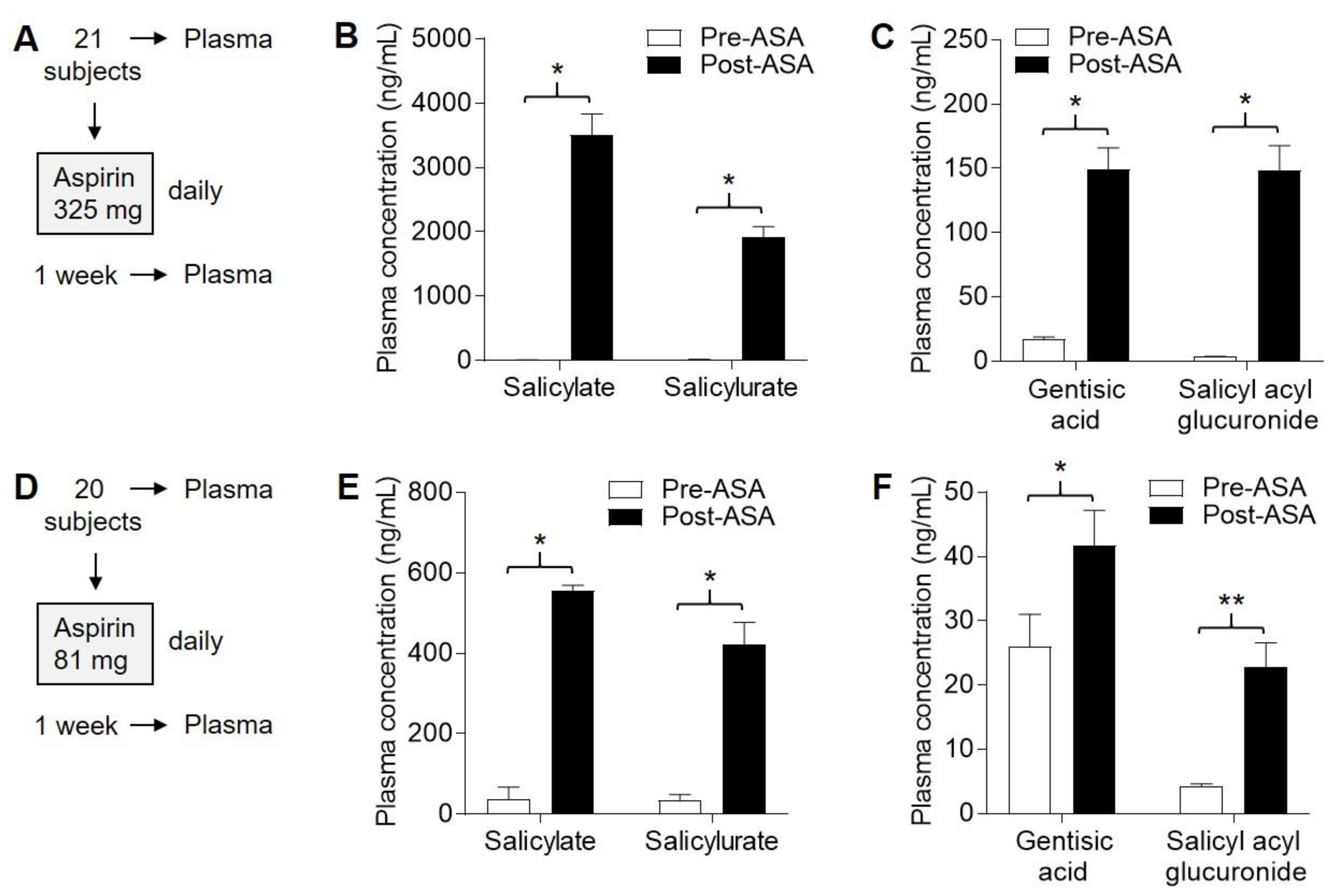
2.5. Four ASA Metabolites Detected in Plasma Following Sub-Chronic ASA Dosing
2.6. Detection of ASA Metabolites in Nevi Following Subchronic ASA Dosing
2.7. Suppression of PGE2 in Plasma and Nevi Following Subchronic ASA Dosing
2.8. Variable Effect on AMPK Activation in Nevi Following Subchronic ASA Dosing
2.9. ASA Reduces Inflammatory Cytokines without Affecting Leukocyte Subsets
2.10. Metabolites Suppressed by ASA
3. Discussion
3.1. Quantitative Detection of ASA Metabolites
3.2. Anti-Inflammatory Effects of ASA
3.3. Modulation of PGE2 in Relevant Target Tissues
3.4. Looking Toward Future Melanoma Chemoprevention Studies
4. Materials and Methods
4.1. Human Subjects
4.2. ASA
4.3. Nevus Tissues
4.4. Blood Collection
4.5. Detection of ASA Metabolites and PGE2 by LC–MS
4.6. Detection of PGE2 in Plasma and Nevi by ELISA
4.7. Western Blotting
4.8. Cytokine Analysis
4.9. Metabolomics
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Desborough, M.J.; Keeling, D.M. The aspirin story-from willow to wonder drug. Br. J. Haematol. 2017, 177, 674–683. [Google Scholar] [CrossRef]
- Barnard, M.E.; Poole, E.M.; Curhan, G.C.; Eliassen, A.H.; Rosner, B.A.; Terry, K.L.; Tworoger, S.S. Association of Analgesic Use With Risk of Ovarian Cancer in the Nurses’ Health Studies. JAMA Oncol. 2018, 4, 1675–1682. [Google Scholar] [CrossRef]
- Simon, T.G.; Ma, Y.; Ludvigsson, J.F.; Chong, D.Q.; Giovannucci, E.L.; Fuchs, C.S.; Meyerhardt, J.A.; Corey, K.E.; Chung, R.T.; Zhang, X.; et al. Association Between Aspirin Use and Risk of Hepatocellular Carcinoma. JAMA Oncol. 2018, 4, 1683–1690. [Google Scholar] [CrossRef] [PubMed]
- Salinas, C.A.; Kwon, E.M.; FitzGerald, L.M.; Feng, Z.; Nelson, P.S.; Ostrander, E.A.; Peters, U.; Stanford, J.L. Use of aspirin and other nonsteroidal antiinflammatory medications in relation to prostate cancer risk. Am. J. Epidemiol. 2010, 172, 578–590. [Google Scholar] [CrossRef]
- Swede, H.; Mirand, A.L.; Menezes, R.J.; Moysich, K.B. Association of regular aspirin use and breast cancer risk. Oncology 2005, 68, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, P.M.; Wilson, M.; Elwin, C.E.; Norrving, B.; Algra, A.; Warlow, C.P.; Meade, T.W. Long-term effect of aspirin on colorectal cancer incidence and mortality: 20-year follow-up of five randomised trials. Lancet 2010, 376, 1741–1750. [Google Scholar] [CrossRef]
- McNeil, J.J.; Nelson, M.R.; Woods, R.L.; Lockery, J.E.; Wolfe, R.; Reid, C.M.; Kirpach, B.; Shah, R.C.; Ives, D.G.; Storey, E.; et al. Effect of Aspirin on All-Cause Mortality in the Healthy Elderly. N. Engl. J. Med. 2018, 379, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- McNeil, J.J.; Wolfe, R.; Woods, R.L.; Tonkin, A.M.; Donnan, G.A.; Nelson, M.R.; Reid, C.M.; Lockery, J.E.; Kirpach, B.; Storey, E.; et al. Effect of Aspirin on Cardiovascular Events and Bleeding in the Healthy Elderly. N. Engl. J. Med. 2018, 379, 1509–1518. [Google Scholar] [CrossRef]
- Boutaud, O.; Sosa, I.R.; Amin, T.; Oram, D.; Adler, D.; Hwang, H.S.; Crews, B.C.; Milne, G.; Harris, B.K.; Hoeksema, M.; et al. Inhibition of the Biosynthesis of Prostaglandin E2 By Low-Dose Aspirin: Implications for Adenocarcinoma Metastasis. Cancer Prev. Res. 2016, 9, 855–865. [Google Scholar] [CrossRef]
- Grilli, M.; Pizzi, M.; Memo, M.; Spano, P. Neuroprotection by aspirin and sodium salicylate through blockade of NF-kappaB activation. Science 1996, 274, 1383–1385. [Google Scholar] [CrossRef]
- Hawley, S.A.; Fullerton, M.D.; Ross, F.A.; Schertzer, J.D.; Chevtzoff, C.; Walker, K.J.; Peggie, M.W.; Zibrova, D.; Green, K.A.; Mustard, K.J.; et al. The ancient drug salicylate directly activates AMP-activated protein kinase. Science 2012, 336, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Goodman, J.R.; Grossman, D. Aspirin and other NSAIDs as chemoprevention agents in melanoma. Cancer Prev. Res. 2014, 7, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.E.; Gledhill, K.; Masoodi, M.; Haylett, A.K.; Brownrigg, M.; Thody, A.J.; Tobin, D.J.; Nicolaou, A. The sunburn response in human skin is characterized by sequential eicosanoid profiles that may mediate its early and late phases. FASEB J. 2009, 23, 3947–3956. [Google Scholar] [CrossRef] [PubMed]
- Gresham, A.; Masferrer, J.; Chen, X.; Leal-Khouri, S.; Pentland, A.P. Increased synthesis of high-molecular-weight cPLA2 mediates early UV-induced PGE2 in human skin. Am. J. Physiol. 1996, 270, C1037–C1050. [Google Scholar] [CrossRef]
- Gledhill, K.; Rhodes, L.E.; Brownrigg, M.; Haylett, A.K.; Masoodi, M.; Thody, A.J.; Nicolaou, A.; Tobin, D.J. Prostaglandin-E2 is produced by adult human epidermal melanocytes in response to UVB in a melanogenesis-independent manner. Pigment. Cell Melanoma Res. 2010, 23, 394–403. [Google Scholar] [CrossRef]
- Prasad, R.; Katiyar, S.K. Prostaglandin E2 Promotes UV radiation-induced immune suppression through DNA hypermethylation. Neoplasia 2013, 15, 795–804. [Google Scholar] [CrossRef]
- Becker, M.R.; Siegelin, M.D.; Rompel, R.; Enk, A.H.; Gaiser, T. COX-2 expression in malignant melanoma: A novel prognostic marker? Melanoma Res. 2009, 19, 8–16. [Google Scholar] [CrossRef]
- Denkert, C.; Kobel, M.; Berger, S.; Siegert, A.; Leclere, A.; Trefzer, U.; Hauptmann, S. Expression of cyclooxygenase 2 in human malignant melanoma. Cancer Res. 2001, 61, 303–308. [Google Scholar]
- Kuzbicki, L.; Lange, D.; Straczynska-Niemiec, A.; Chwirot, B.W. The value of cyclooxygenase-2 expression in differentiating between early melanomas and histopathologically difficult types of benign human skin lesions. Melanoma Res. 2012, 22, 70–76. [Google Scholar] [CrossRef]
- Zhang, Y.; Daaka, Y. PGE2 promotes angiogenesis through EP4 and PKA Cgamma pathway. Blood 2011, 118, 5355–5364. [Google Scholar] [CrossRef]
- Jain, S.; Chakraborty, G.; Raja, R.; Kale, S.; Kundu, G.C. Prostaglandin E2 regulates tumor angiogenesis in prostate cancer. Cancer Res. 2008, 68, 7750–7759. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Rahman, H.; Tyagi, E.; Liu, T.; Li, C.; Lu, R.; Lum, D.; Holmen, S.L.; Maschek, J.A.; Cox, J.E.; et al. Aspirin Suppresses PGE2 and Activates AMP Kinase to Inhibit Melanoma Cell Motility, Pigmentation, and Selective Tumor Growth In Vivo. Cancer Prev. Res. 2018, 11, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Y.; Kim, W.J.; Daaka, Y. PGE2 promotes renal carcinoma cell invasion through activated RalA. Oncogene 2013, 32, 1408–1415. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Davison, C. Salicylate metabolism inet al. man. Ann. N. Y. Acad. Sci. 1971, 179, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Liesenfeld, D.B.; Botma, A.; Habermann, N.; Toth, R.; Weigel, C.; Popanda, O.; Klika, K.D.; Potter, J.D.; Lampe, J.W.; Ulrich, C.M. Aspirin Reduces Plasma Concentrations of the Oncometabolite 2-Hydroxyglutarate: Results of a Randomized, Double-Blind, Crossover Trial. Cancer Epidemiol. Biomarkers Prev. 2016, 25, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Chhonker, Y.S.; Pandey, C.P.; Chandasana, H.; Laxman, T.S.; Prasad, Y.D.; Narain, V.S.; Dikshit, M.; Bhatta, R.S. Simultaneous quantitation of acetylsalicylic acid and clopidogrel along with their metabolites in human plasma using liquid chromatography tandem mass spectrometry. Biomed. Chromatogr. 2016, 30, 466–473. [Google Scholar] [CrossRef]
- Li, J.P.; Guo, J.M.; Shang, E.X.; Zhu, Z.H.; Liu, Y.; Zhao, B.C.; Tang, Z.S.; Duan, J.A. Quantitative determination of five metabolites of aspirin by UHPLC-MS/MS coupled with enzymatic reaction and its application to evaluate the effects of aspirin dosage on the metabolic profile. J. Pharm. Biomed. Anal. 2017, 138, 109–117. [Google Scholar] [CrossRef]
- Navarro, S.L.; Saracino, M.R.; Makar, K.W.; Thomas, S.S.; Li, L.; Zheng, Y.; Levy, L.; Schwarz, Y.; Bigler, J.; Potter, J.D.; et al. Determinants of aspirin metabolism in healthy men and women: Effects of dietary inducers of UDP-glucuronosyltransferases. J. Nutr. Nutr. 2011, 4, 110–118. [Google Scholar] [CrossRef]
- Pirola, R.; Bareggi, S.R.; de Benedittis, G. Determination of acetylsalicylic acid and salicylic acid in skin and plasma by high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1998, 705, 309–315. [Google Scholar] [CrossRef]
- Bae, S.K.; Seo, K.A.; Jung, E.J.; Kim, H.S.; Yeo, C.W.; Shon, J.H.; Park, K.M.; Liu, K.H.; Shin, J.G. Determination of acetylsalicylic acid and its major metabolite, salicylic acid, in human plasma using liquid chromatography-tandem mass spectrometry: Application to pharmacokinetic study of Astrix in Korean healthy volunteers. Biomed. Chromatogr. 2008, 22, 590–595. [Google Scholar] [CrossRef]
- Sample, D.; Wargovich, M.; Fischer, S.M.; Inamdar, N.; Schwartz, P.; Wang, X.; Do, K.; Sinicrope, F.A. A dose-finding study of aspirin for chemoprevention utilizing rectal mucosal prostaglandin E(2) levels as a biomarker. Cancer Epidemiol. Biomarkers Prev. 2002, 11, 275–279. [Google Scholar]
- Ferreira, P.M.; Gagliano-Juca, T.; Zaminelli, T.; Sampaio, M.F.; Blackler, R.W.; Trevisan Mda, S.; Novaes Magalhães, A.F.; De Nucci, G. Acetylsalicylic Acid Daily vs Acetylsalicylic Acid Every 3 Days in Healthy Volunteers: Effect on Platelet Aggregation, Gastric Mucosa, and Prostaglandin E2 Synthesis. J. Clin. Pharmacol. 2016, 56, 862–868. [Google Scholar] [CrossRef]
- Ruffin, M.T., 4th; Normolle, D.; Vaerten, M.A.; Peters-Golden, M.; Brenner, D.E.; Krishnan, K.; Rock, C.L.; Boland, C.R.; Crowell, J.; Kelloff, G. Suppression of human colorectal mucosal prostaglandins: Determining the lowest effective aspirin dose. J. Natl. Cancer Inst. 1997, 89, 1152–1160. [Google Scholar] [CrossRef]
- Mullangi, R.; Sharma, K.; Srinivas, N.R. Review of HPLC methods and HPLC methods with mass spectrometric detection for direct determination of aspirin with its metabolite(s) in various biological matrices. Biomed. Chromatogr. 2012, 26, 906–941. [Google Scholar] [CrossRef]
- Kim, J.E.; Kwon, J.Y.; Seo, S.K.; Son, J.E.; Jung, S.K.; Min, S.Y.; Hwang, M.K.; Heo, Y.S.; Lee, K.W.; Lee, H.J. Cyanidin suppresses ultraviolet B-induced COX-2 expression in epidermal cells by targeting MKK4, MEK1, and Raf-1. Biochem. Pharmacol. 2010, 79, 1473–1482. [Google Scholar] [CrossRef]
- Chang, M.S.; Chen, B.C.; Yu, M.T.; Sheu, J.R.; Chen, T.F.; Lin, C.H. Phorbol 12-myristate 13-acetate upregulates cyclooxygenase-2 expression in human pulmonary epithelial cells via Ras, Raf-1, ERK, and NF-kappaB, but not p38 MAPK, pathways. Cell Signal. 2005, 17, 299–310. [Google Scholar] [CrossRef]
- Poynter, J.N.; Elder, J.T.; Fullen, D.R.; Nair, R.P.; Soengas, M.S.; Johnson, T.M.; Redman, B.; Thomas, N.E.; Gruber, S.B. BRAF and NRAS mutations in melanoma and melanocytic nevi. Melanoma Res. 2006, 16, 267–273. [Google Scholar] [CrossRef]
- Nishihara, R.; Lochhead, P.; Kuchiba, A.; Jung, S.; Yamauchi, M.; Liao, X.; Imamura, Y.; Qian, Z.R.; Morikawa, T.; Wang, M.; et al. Aspirin use and risk of colorectal cancer according to BRAF mutation status. JAMA 2013, 309, 2563–2571. [Google Scholar]
- Gao, M.; Kong, Q.; Hua, H.; Yin, Y.; Wang, J.; Luo, T.; Jiang, Y. AMPK-mediated up-regulation of mTORC2 and MCL-1 compromises the anti-cancer effects of aspirin. Oncotarget 2016, 7, 16349–16361. [Google Scholar] [CrossRef][Green Version]
- Din, F.V.; Valanciute, A.; Houde, V.P.; Zibrova, D.; Green, K.A.; Sakamoto, K.; Alessi, D.R.; Dunlop, M.G. Aspirin inhibits mTOR signaling, activates AMP-activated protein kinase, and induces autophagy in colorectal cancer cells. Gastroenterology 2012, 142, 1504–1515. e1503. [Google Scholar] [CrossRef]
- Buskin, J.N.; Upton, R.A.; Williams, R.L. Improved liquid-chromatography of aspirin, salicylate, and salicyluric acid in plasma, with a modification for determining aspirin metabolites in urine. Clin. Chem. 1982, 28, 1200–1203. [Google Scholar]
- Bakar, S.K.; Niazi, S. High-performance liquid chromatographic determination of aspirin and its metabolites in plasma and urine. J. Pharm. Sci. 1983, 72, 1020–1023. [Google Scholar] [CrossRef]
- Yamamoto, E.; Takakuwa, S.; Kato, T.; Asakawa, N. Sensitive determination of aspirin and its metabolites in plasma by LC-UV using on-line solid-phase extraction with methylcellulose-immobilized anion-exchange restricted access media. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 846, 132–138. [Google Scholar] [CrossRef]
- Bareggi, S.R.; Pirola, R.; de Benedittis, G. Skin and plasma levels of acetylsalicylic acid: A comparison between topical aspirin/diethyl ether mixture and oral aspirin in acute herpes zoster and postherpetic neuralgia. Eur. J. Clin. Pharmacol. 1998, 54, 231–235. [Google Scholar] [CrossRef]
- Claria, J.; Serhan, C.N. Aspirin triggers previously undescribed bioactive eicosanoids by human endothelial cell-leukocyte interactions. Proc. Natl. Acad. Sci. USA 1995, 92, 9475–9479. [Google Scholar] [CrossRef]
- Morris, T.; Stables, M.; Hobbs, A.; de Souza, P.; Colville-Nash, P.; Warner, T.; Newson, J.; Bellingan, G.; Gilroy, D.W. Effects of low-dose aspirin on acute inflammatory responses in humans. J. Immunol. 2009, 183, 2089–2096. [Google Scholar] [CrossRef]
- Esquivel-Velazquez, M.; Ostoa-Saloma, P.; Palacios-Arreola, M.I.; Nava-Castro, K.E.; Castro, J.I.; Morales-Montor, J. The role of cytokines in breast cancer development and progression. J. Interferon Cytokine Res. 2015, 35, 1–16. [Google Scholar] [CrossRef]
- Crucioli, V.; Stilwell, J. The histogenesis of malignant melanoma in relation to pre-existing pigmented lesions. J. Cutan. Pathol. 1982, 9, 396–404. [Google Scholar] [CrossRef]
- Kaur, J.; Sanyal, S.N. PI3-kinase/Wnt association mediates COX-2/PGE(2) pathway to inhibit apoptosis in early stages of colon carcinogenesis: Chemoprevention by diclofenac. Tumour. Biol. 2010, 31, 623–631. [Google Scholar] [CrossRef]
- Cassidy, P.B.; Liu, T.; Florell, S.R.; Honeggar, M.; Leachman, S.A.; Boucher, K.M.; Grossman, D. A Phase II Randomized Placebo-Controlled Trial of Oral N-acetylcysteine for Protection of Melanocytic Nevi against UV-Induced Oxidative Stress In Vivo. Cancer Prev. Res. 2017, 10, 36–44. [Google Scholar] [CrossRef]
- Affolter, K.; Samowitz, W.; Tripp, S.; Bronner, M.P. BRAF V600E mutation detection by immunohistochemistry in colorectal carcinoma. Genes Chromosomes Cancer 2013, 52, 748–752. [Google Scholar] [CrossRef]
- Washburn, R.L.; Cox, J.E.; Muhlestein, J.B.; May, H.T.; Carlquist, J.F.; Le, V.T.; Anderson, J.L.; Horne, B.D. Pilot Study of Novel Intermittent Fasting Effects on Metabolomic and Trimethylamine N-oxide Changes during 24-hour Water-only Fasting in the FEELGOOD Trial. Nutrients 2019, 11, 246. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varedi, A.; Rahman, H.; Kumar, D.; Catrow, J.L.; Cox, J.E.; Liu, T.; Florell, S.R.; Boucher, K.M.; Okwundu, N.; Burnett, W.J.; et al. ASA Suppresses PGE2 in Plasma and Melanocytic Nevi of Human Subjects at Increased Risk for Melanoma. Pharmaceuticals 2020, 13, 7. https://doi.org/10.3390/ph13010007
Varedi A, Rahman H, Kumar D, Catrow JL, Cox JE, Liu T, Florell SR, Boucher KM, Okwundu N, Burnett WJ, et al. ASA Suppresses PGE2 in Plasma and Melanocytic Nevi of Human Subjects at Increased Risk for Melanoma. Pharmaceuticals. 2020; 13(1):7. https://doi.org/10.3390/ph13010007
Chicago/Turabian StyleVaredi, Amir, Hafeez Rahman, Dileep Kumar, Jonathan L. Catrow, James E. Cox, Tong Liu, Scott R. Florell, Kenneth M. Boucher, Nwanneka Okwundu, William J. Burnett, and et al. 2020. "ASA Suppresses PGE2 in Plasma and Melanocytic Nevi of Human Subjects at Increased Risk for Melanoma" Pharmaceuticals 13, no. 1: 7. https://doi.org/10.3390/ph13010007
APA StyleVaredi, A., Rahman, H., Kumar, D., Catrow, J. L., Cox, J. E., Liu, T., Florell, S. R., Boucher, K. M., Okwundu, N., Burnett, W. J., VanBrocklin, M. W., & Grossman, D. (2020). ASA Suppresses PGE2 in Plasma and Melanocytic Nevi of Human Subjects at Increased Risk for Melanoma. Pharmaceuticals, 13(1), 7. https://doi.org/10.3390/ph13010007