Abstract
Cataracts, one of the leading causes of preventable blindness worldwide, refers to lens degradation that is characterized by clouding, with consequent blurry vision. As life expectancies improve, the number of people affected with cataracts is predicted to increase worldwide, especially in low-income nations with limited access to surgery. Although cataract surgery is considered safe, it is associated with some complications such as retinal detachment, warranting a search for cheap, pharmacological alternatives to the management of this ocular disease. The lens is richly endowed with a complex system of non-enzymatic and enzymatic antioxidants which scavenge reactive oxygen species to preserve lens proteins. Depletion and/or failure in this primary antioxidant defense system contributes to the damage observed in lenticular molecules and their repair mechanisms, ultimately causing cataracts. Several attempts have been made to counteract experimentally induced cataract using in vitro, ex vivo, and in vivo techniques. The majority of the anti-cataract compounds tested, including plant extracts and naturally-occurring compounds, lies in their antioxidant and/or free radical scavenging and/or anti-inflammatory propensity. In addition to providing an overview of the pathophysiology of cataracts, this review focuses on the role of various categories of natural and synthetic compounds on experimentally-induced cataracts.
1. Introduction
Cataracts, one of the leading causes of preventable blindness worldwide, refers to lens degradation that is characterized by clouding, with consequent blurry or hazy vision [1]. There is evidence that cataracts account for 10.8 million out of 32.4 million blind individuals, and that 35.1 million out of 191 million people with impaired vision globally have this debilitating disease [2]. The prevalence of cataracts increases exponentially after 40 years of age, ranging from 3.9% among 55–64-year-olds to 92.6% among those 80 years and older [3,4,5]. In the US, the number of people suffering from cataracts is projected to double from 24.4 to 50 million by the year 2050 [6]. Epidemiological studies have elucidated common cataract risk factors to include age, smoking, ultraviolet (UV) radiation, female gender, steroid consumption, diabetes mellitus, and high body mass index [7,8]. As life expectancies improve throughout the globe, the number of people suffering from cataracts is predicted to increase worldwide [9], especially in low-income nations that lack easy access to cataract surgery, warranting a search for cheap, pharmacological alternatives to the management of this disease.
1.1. Classification, Signs and Symptoms of Cataracts
Cataracts are classified based upon the etiology of disease and anatomical location of opacity. Table 1 provides the etiological classification of various forms of cataracts as summarized by Gupta et al. (2014) [10]. Congenital and developmental cataracts occur during fetal growth or growth of children. Age-related cataracts are associated with old age and are mainly attributed to oxidative stress. Traumatic, complicated, and metabolic cataracts are attributed to physical trauma, inflammatory and degenerative eye disease, and metabolic disease, respectively. On the contrary, toxic, radiation and electrical cataracts occur due to exposure to toxicants, electromagnetic waves, and high electrical currents.

Table 1.
Type of cataract and their respective causes and risks [10].
Age-related cataracts can be further classified based upon the anatomical location of opacity within the lens into nuclear, cortical, and posterior subcapsular cataracts (Figure 1A–D) [11]. Nuclear cataracts affect the center of the lens, with the lens becoming yellow or brown after hardening (nuclear sclerosis). Nuclear cataracts are common with older age and are associated with myopia. In contrast, cortical cataracts affect outer fibers of the lens (around edges of the nucleus) and assume a wedge-shaped appearance. Cortical cataracts can result in glares but do not alter vision as much as nuclear cataracts do. Posterior subcapsular cataracts, which affect the posterior cortex of the lens, are observed in relatively younger patients. This form of cataracts is associated with hyperopia and progresses faster than nuclear and cortical cataracts. Like cortical cataracts patients, patients with posterior subcapsular cataracts may experience glare. Posterior subcapsular cataracts are also associated with corticosteroid use [12]. Shared symptoms across different types of age-related cataracts include clouded or blurred vision (Figure 2), faded colors, glare, a halo around lights, poor night vision, and frequent prescription changes for corrective lenses [6,13].
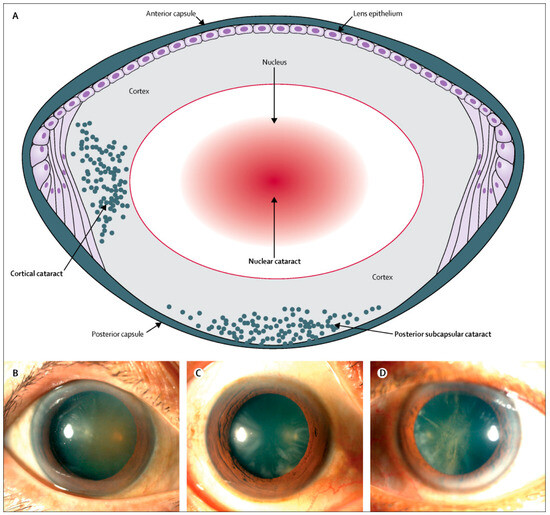
Figure 1.
Characteristics of lens structures and major types of cataracts for location-based classification. (A) A schematic view of lens structures and corresponding types of cataracts. Slit lamp biomicroscopy photos showing (B) nuclear cataract, (C) cortical cataract, and (D) subcapsular posterior cataract. Copyright Elsevier (used with permission from Elsevier) [11].

Figure 2.
Scene viewed by normal vision (A) and a person with cataracts (B); sourced from NEI Media Library [13].
1.2. Lens Anatomy and Physiology
A review of the anatomy and physiology of the lens is necessary to understand the pathophysiology of the disease. The lens is a transparent structure that is devoid of any blood supply. Anteriorly, the lens surface is covered by a monolayer of epithelial cells. In addition to maintaining lens metabolic activity, epithelial cells replicate to produce daughter cells, which migrate and differentiate into fiber cells [14]. Lens fiber cells make up greater than 95% of the lens and are stretched out to form compact, concentric layers (“shells”), thereby reducing intercellular space (Figure 3) [15]. Superficial lens fibers are nucleated and are metabolically active while deeper fibers, which make up most of the lens, are organelle-free with minimal metabolic activity. Interiorly, fiber cells have a high expression of soluble crystallin proteins but are devoid of nuclei, mitochondria, endoplasm reticulum, ribosomes, and other organelles [16]. Lens crystallins make up almost 90% of proteins in the mature lens [17]. In humans, the non-nucleated human lens fiber cells consist of α-crystallins, β-crystallins, and γ-crystallins (Table 2) [17]. Purification of high-molecular-weight α-crystallin fraction from human lenses yielded two homologous α-crystallin polypeptides: αA-crystallins and αB-crystallins [18,19]. The α-crystallin proteins account for up to one-third of the total protein in the lens [20]. However, three γ-crystallins (γC, γD, γS-crystallin), which are also found with five β-crystallin polypeptides (βB1, βB2, βB3, β A1/A3, βA4) predominate [21]. Table 2 provides a summary of crystallins identified in human lens according to size, amino acid residues, Gibbs free energy, their encoding genes, and chromosomal location [17].
Table 2.
Crystallins identified in human lens [17].
The stability of soluble crystallins is regulated by several protective systems within and surrounding the lens. Acetylation of N-terminal residues in β- and γ-crystallins confer inherent protection of these lens proteins from exopeptidases. Additionally, acetylated N-terminal residues of α-crystallin subunits are buried in the interior of high-molecular-weight α-crystallin aggregate [22], rendering them inaccessible. The α-crystallin proteins prevent heat induced precipitation of β- and γ-crystallins [20] and serve as molecular chaperones that cloister misfolded proteins to mitigate pervasive protein aggregation [19]. Crystallin stability is further enhanced by the temperature difference in the anterior segment. Anterior portion of the eye is 2 °C lower than total body temperature due to evaporative cooling from cornea and distance from blood supply [23]. Additionally, ion pumps such as the Na+/K+ ATPase and Ca2+ATPase in pre-equatorial epithelium, together with gap junctions and channels such as aquaporin tightly regulate the homeostatic environment, thereby contributing the stability of lens crystallins [24]. Since deeper fibers which make up most of the lens are organelle-free, there is minimal metabolic activity and lower oxygen levels in the central part of the lens, thereby enhancing stability of lens proteins. Indeed, oxygen concentration in nucleus of lens has been reported to be as low as less than 10 µM in mammalian lens [25,26]. To this end, studies have shown that exposure of mammalian eyes to hyperbaric oxygen is associated with the development of nuclear opacities [27,28].
1.3. Lens Transparency
The lens is essential for focusing light onto the retina and can perform this function due to its transparent and dioptric properties. Transparency of the lens depends on avascularity, the paucity of organelles, narrow inter-fiber spaces, and regular organization of cells and proteins [29]. At the cellular level, there is a limited light-scattering by organelles due to their limited presence in the lens. Moreover, organelles are located away from the light path, exiled to the equator in the fibers from the central epithelium, thereby reducing light scattering in the lens. Transparency is also achieved by the short-range spatial order of proteins [30,31]. In fiber cells, crystallins are densely packed in a short-range order of about 250–400 mg/mL. The small protein size (<10 nm diameter), together with the close packing at high concentration, renders their wavelength less than that of light [17]. Furthermore, dense packing of protein aggregates reduces fluctuations of protein density and reduces the refractive index below wavelength of light [32]. Protein crystallization and precipitation are further deterred through a specialized mixture of crystallin protein forms (e.g., α, β, and γ forms), which confer superior solubility and native protein conformations in the lens [17]. In addition to their structural function within the lens, by increasing the refractive index, β- and γ-crystallins exhibit high solubility and thermodynamic stability to prevent scattering of light [22]. The α-crystallins serve as chaperones by partially binding to denatured proteins within the lens cells to form high-molecular-weight aggregates that maintain protein solubility and transparency [33].
In the cortex of the lens, transparency is enhanced by a high spatial order of fiber architecture with narrow intercellular spaces, which then compensates for light-scattering due to refractive index differences between membranes and cytoplasm. In the nucleus, high spatial order is not required due to minimal light scattering and negligible differences in the refractive index between fiber membranes and cytoplasm [15,34]. The cornea traps light with wavelength below 310 nm [35]. Interestingly, the mammalian lens possesses small-molecular-weight UV filters such as tryptophan metabolites (e.g., kynurenine, 3-hydroxykynurenine, 3-hydroxykynurenine glucoside) that remove UV radiation between 300–400 nm. Indeed, pathways for the biosynthesis of 3-hydroxykynurenine glucoside have also been identified in the mammalian lens [36,37,38].

Figure 3.
Schematic presentation of the cross-sectional view of mammalian lens. Used with permission from Roberts J [39].
Figure 3.
Schematic presentation of the cross-sectional view of mammalian lens. Used with permission from Roberts J [39].

2. Antioxidant Systems in the Lens
The lens is richly endowed with a complex antioxidant system consisting of non-enzymatic and enzymatic pathways, which further contribute to lens transparency. The major non-enzymatic antioxidants are glutathione (GSH), vitamin C (ascorbic acid), vitamin E, and carotenoids, while the enzymatic antioxidants include superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), glutathione reductase (GSH-Rx), and catalase (CAT). Both non-enzymatic and enzymatic antioxidants scavenge reactive oxygen species to protect the lens. Failure in this primary antioxidant defense system leads to damage to lenticular molecules and their repair mechanisms. Ultimately, the degradation of lenticular molecules leads to cataracts [40,41,42,43].
2.1. Non-Enzymatic Antioxidants
GSH is a tripeptide thiol with a sulfhydryl residue that scavenges hydroperoxides and electrophilic compounds [44]. Lenticular GSH synthesis occurs in the cortex from the amino acids, glycine, cysteine, and glutamate. The enzyme, γ-glutamylcysteine synthase catalyzes the first and rate-limiting step in the biosynthesis of GSH and exerts feedback inhibition in response to GSH levels [44,45]. Incidentally, the lens has the highest concentration of GSH in the body [46], with the highest quantities being localized in the cortex, its site of synthesis [47]. GSH has a significant role in lens transparency by guarding membrane thiol groups which are vital in cation transport and regulation of the electrolyte balance [48]. Vitamin C is another non-enzymatic antioxidant that plays a vital role in the physiology of the lens. In a normal human lens, it is present at high concentrations of up to 3 mmol/L, a level which is 20–60-fold higher than that of the plasma content [49]. Similarly, α-tocopherol is also found in the normal human lens at 1.6 μg/g of lens tissue [49]. The only carotenoids found in the human lens are lutein and zeaxanthin. About 74% of these carotenoids are localized in the epithelium and cortex [50].
2.2. Enzymatic Antioxidants
Among enzymatic antioxidants, SOD removes superoxides through one electron dismutation of superoxide into H2O2 and oxygen. Three mammalian SOD isoforms have been described based on their structure, location, and metal-cofactors: the cytosolic copper-zinc (CuZn)-SOD (EC 1.15.1.1) or SOD-1, the mitochondrial (Mn)-SOD (EC 1.15.1.1) or SOD-2, and the extracellular (EC)-SOD (EC 1.15.1.1) or SOD-3, which also contains copper and zinc [51]. SOD-1 enzyme is the most prominent isotype in the lens [52]. The copper ion in the enzyme is involved in dismutation reactions undertaking alternate oxidation and reduction, while the zinc ion has only a stabilizing role with no activity in these reactions [53]. The H2O2 produced as a result of dismutation reaction is removed through the CAT and GSH-Px systems. GSH-Px removes H2O2 by using it to oxidize GSH, whereas CAT removes the H2O2 through the formation of oxygen and water [45,54,55]. Moreover, GSH disulfide (GSSG) is converted to a reduced form, GSH using GSH-Px enzyme and the co factor NADPH, from the pentose phosphate pathway [45,56]. Primate lenses have such robust protective antioxidant mechanisms that they can withstand incubation in H2O2 (1 mM), partly due to the high level of GSH-reductase (GSH-Rx) activity [57]. Additionally, there is a GSH-dependent thioltransferase system, including a GSH-S-transferase (GST) enzyme that repairs lens protein oxidation by cleaving protein–thiol groups to maintain a reduced state [58,59,60]. Lenticular thiol/disulfide homeostasis is upheld and the protein–protein disulfide bond is also maintained at low levels by the NADPH-dependent thioredoxin/thioredoxin reductase system [43,58,61,62]. Collectively, these protein and enzyme antioxidant systems play a vital role in further prevention of crystallin aggregation and the consequent development of cataracts [30,45,46].
3. Molecular Mechanisms of Cataract Formation
The molecular mechanism of cataract formation has been explained by the “protein aggregation” and/or “loss of protein solubility” disease models. The “protein aggregation” disease model hypothesizes that covalent bond damages lens crystallin proteins due to insult to systems that maintain redox, ionic, and other physiological environment leading to a partial unfolding of crystallin proteins. Research suggests that several mechanisms, including deamidation, oxidation, glycation, and truncation of crystallins, contribute to these covalent modifications associated with damage to lens crystallins [17,63,64,65,66,67,68,69,70,71,72,73,74]. Consequently, the crystallin proteins lose their native stability to become unstable molecules that aggregate, thereby shifting the refractive index. Thus, this model is based on loss of crystallin stability and the tendency of proteins to partially or fully unfold [17,75]. In support of this hypothesis, studies on materials from cataractous lens have revealed the presence of multiple species of lens proteins with higher molecular weight forms due to aggregation and polymerization. These high molecular weight aggregates cause light scattering in the lens, thereby accounting for lens opacity [63,64,65,66,67,68,69,70,71,72,73,74]. The “protein aggregation” cataract disease model is contrary to that proposed for other protein aggregation diseases such as Alzheimer’s disease, Huntington’s disease and Parkinson’s disease, where the high molecular weight proteins further aggregate to form plaque deposits in the affected parts of the CNS [17].
The “loss of solubility” model hypothesizes that aberrant interactions of native crystallins lead to loss of solubility of crystallins in their native conformations, with consequent precipitation into protein-rich and protein-poor regions. This ultimately disrupts the short-range order to alter the refractive index of the lens [75,76]. Some studies have proposed that altered solubility and crystallization propensity account for hereditary cataract in certain disease states [76,77,78,79].
4. Treatments Strategies for Cataracts
4.1. Current Cataract Treatments
Up to date, there is no pharmacological intervention available for the management of cataracts, leaving surgery as the main option for the treatment of this disease. Among the different surgical approaches, phacoemulsification is a technique that involves the use of a probe that emits ultrasound to break up the lens, which is then aspirated through a thin cannula introduced with a smaller incision in the cornea. Extracapsular cataract extraction, which involves the removal of nucleus and cortical materials, leaving behind the posterior capsule, is commonly used in developed countries. This procedure is used for advanced cataract that cannot be removed with phacoemulsification [80]. While intracapsular surgery is not routinely used, it involves surgical removal of the entire lens with its capsule [81]. Even though cataractous lenses can be surgically removed and replaced with an artificial intraocular lens, a procedure that has been successful in restoring sight, there are still barriers to cataracts surgery and eye care in many countries. Indeed, access to surgery remains a challenge in developing nations, where cataracts are estimated to accounts for 90% of total blindness among those of lower socioeconomic status [9]. Additionally, intra-operative (e.g., posterior capsular rapture, zonular weakness, suprachoroidal hemorrhage) and post-operative complications (e.g., posterior capsular opacification, elevated IOP, persistent anterior uveitis, retinal detachment) still occur [82]. Thus, pharmacological strategies to management of cataracts may play a significant role in regions of the world where surgical options are limited.
4.2. Potential Pharmacological Treatments for Cataracts
Based on the models of cataract development and the proposed mechanism for its formation, there have been attempts to use herbal and/or antioxidants, amino acids, and minerals to attenuate the development of cataracts. Interference with protein aggregation pathway and modulation of chaperone activity to promote refolding are also among the efforts that have been attempted. Furthermore, patented approaches involving the following mechanisms are available: inhibition of glycation-specific mechanisms, phase separation inhibitors, modulators of the TGF-β pathway, and inhibition of matrix metalloproteinase [83]. This section provides a review of different categories of compounds that have been evaluated for anti-cataract potential, in vitro, ex vivo and in vivo.
4.2.1. Antioxidants
The role of oxidative stress in the onset and progression of cataractogenesis has been well described in literature [40,70]. Consequently, antioxidants and free radical scavengers present a potential therapeutic strategy to the management of cataracts.
(i) In vitro and ex vivo studies
Typical antioxidants
There is evidence that depletion of GSH affects lens transparency and leads to the development of cataracts. Indeed, lenticular GSH is essential in preventing oxidative stress elicited by H2O2, where it serves to maintain lens protein sulfhydryl groups in a reduced state, thereby averting formation of inter- and intra-molecular disulfide cross-linkages. GSH also plays a protective role in membrane permeability and active transport of lens protein sulfhydryl groups. It regulates calcium influx and protects lenticular protein from oxidative and osmotic stress [84,85,86]. Indeed, GSH mitigated lipid peroxidation and enhanced the activity of SOD, CAT, and GSH-Px enzymes in H2O2 (10 mM)-induced cataract formation in cultured goat lenses, ex vivo [87]. There is evidence that endogenous antioxidants, vitamin C, and GSH attenuated structural crosslinking, oligomerization, and proteolytic instability of lens crystallins in a lens protein solution exposed to sunlight in the presence of a photosensitizer, riboflavin [88]. Taken together, these studies affirm the protective role of GSH in the pathophysiology of the lens.
The carotenoids, lutein and zeaxanthin, and the vitamin E derivative, α-tocopherol, mitigated an H2O2-induced increase in protein oxidation, lipid peroxidation, and DNA damage in human lens epithelial cells. Moreover, these antioxidants also increased GSH and the GSH:GSSG ratio [89,90]. Interestingly, the vitamins C and E but not the carotenoids, lutein and zeaxanthin, conferred resistance in H2O2-induced depletion of GSH in epithelial cells, in vitro, suggesting that protective effect of carotenoids was not dependent on the elevation of GSH [89,90]. The vitamin E derivative, α-tocopherol, prevented the formation and progression of glucose-induced cataract in goat lenses incubated in artificial aqueous humor. This activity was further supported by the higher Na+-K+-ATPase activity and low lipid peroxidation compared to glucose-only treated lenses [91].
Amifostine, an organic thiophosphate prodrug, is clinically used as a cytoprotective adjuvant in cancer chemotherapy due to its free radical scavenging capacity and ability to detoxify reactive drug metabolites [92]. Belkacémi et al. (2001) reported that the active metabolite of amifostin, aminothiol WR-1065, protected bovine epithelial cells against X-ray radiation of 10 Gy (2 Gy per minute) as evidenced by the elevation of GSH content, enhanced cell viability, and a decrease in both nuclear condensation and epithelial cell apoptosis [93].
In the past few decades, an endogenously-derived organosulfur compound that serves as an essential co-factor for several enzymes in the body, α-lipoic acid, has emerged as a potent antioxidant, nutritional supplement, and an aldose reductase inhibitor [94,95]. There is evidence that α-lipoic acid elicits its antioxidant activity by serving as a free radical scavenger and/or stimulating the biosynthesis of endogenous antioxidants such as GSH [96,97,98]. To this end, α-lipoic acid mitigated H2O2 (0.2 mM)-induced lens opacification, apoptosis and lipid peroxidation in cultured Sprague–Dawley rat lenses, ex vivo. Moreover, in biochemical studies, it enhanced the activity of the endogenous antioxidants, SOD and GSH-peroxidase in peroxide-treated lens homogenates [99].
Keto acids and amino acid antioxidants
There is evidence that the ketoacids, pyruvate and α-ketoglutarate, which are intricately involved in several metabolic pathways in the body, possess antioxidant activity in various biological systems [100,101,102]. Oxaloacetate functions as an intermediate in several metabolic pathways, including gluconeogenesis, the glyoxylate cycle, amino acid synthesis and fatty acid synthesis [103]. Both pyruvate and oxaloacetate serve as precursors for the synthesis of several essential and non-essential amino acids. Interestingly, pyruvate (10 mM), α-ketoglutarate(20 mM), and oxaloacetate (20 mM) decreased the level of lenticular lipid peroxidation and enhanced activity of GSH-peroxidase in H2O2 (10 mM)-induced cataract formation in cultured goat lenses [104]. Similarly, pyruvate attenuated in H2O2 (10 mM)-induced lipid peroxidation and enhanced SOD, CAT, and GSH-peroxidase activity in cultured goat lenses [87]. Moreover, pyruvate prevented H2O2-induced insolubilization of ɣ-crystallin content in rat organ culture system [105], thereby affirming the protective role of ketoacids in mammalian lens physiology.
In addition to the ketoacids, amino acids and their derivatives have also been reported to exert a protective effect on mammalian lenses. Amino acids make a vital contribution to lens clarity because they serve as precursors for the biosynthesis of lens proteins (such as GSH, crystallins). Indeed, insufficient ingestion of certain amino acids has been linked to cataractogenesis in rats [106]. Rathore and Gupta (2010) evaluated the effect several amino acids, glycine, l-tryptophan, l-tyrosine, l-phenylalanine, l-histidine, l-lysine, l-arginine, l-cysteine, l-glutamic acid, l-aspartic acid, and l-proline on H2O2 (1 mM)-induced cataract formation in goat lenses for 24 h. With the exception of l-tyrosine and l-phenylalanine, all amino acids tested conferred significant protection from peroxide-induced GSH depletion. Moreover, all amino acids except l-aspartic acid elevated total soluble protein content in peroxide-treated lenses. Taken together, these data support a protective function for amino acids in the mammalian lens, ex vivo [107].
l-cysteine is an essential amino acid that serves as a precursor for the biosynthesis of the tripeptide thiol antioxidant, GSH. N-acetylcysteine, a prodrug for l-cysteine, is well established as a thiol antioxidant with better bioavailability compared to the parent amino acid. N-acetyl-cysteine is clinically versatile, being used as an antidote for acetaminophen poisoning and a mucolytic agent, amongst others [108]. In experimental animals, N-acetylcysteine has been shown to replete hyperoxia-induced reduction of GSH in rat liver and mitigate hyperglycemia-mediated protein oxidation in cultured rabbit lens cells, in vitro [109,110]. N-acetylcysteine amide, a derivative of N-acetylcysteine, exhibits similar antioxidant activity but superior accessibility to intracellular tissues due to its lipophilicity [111]. Wang et al. (2009) investigated the effect of N-acetylcysteine amide on hyperoxia-induced cortical opacification in adult New Zealand rabbit lenses, ex vivo. These workers reported that N-acetylcysteine amide (20 and 40 mM) prevented hyperoxia-induced cortical opacification and reduction in Na+-K+-ATPase activity. Additionally, this compound elicited an elevation in soluble proteins, GSH and GSH-Rx activity [112]. In other studies, N-acetylcysteine amide protected rat lenses from dexamethasone-induced cataract formation with a corresponding elevation in the GSH:GSSG ratio, GSH-Rx activity, and a reduction in lipid peroxidation [113], thereby affirming the protective role of N-acetylcysteine amide on cataract formation, ex vivo.
Acetyl-l-carnitine is an amine precursor for carnitine, a key player in fatty acid metabolism within the cell. There is evidence that acetyl-l-carnitine confers protection from oxidative stress in several biological systems [114,115]. Acetyl-l-carnitine mitigated selenite-induced cataractogenesis in Wistar rat lenses, ex vivo. Moreover, the activities of CAT, GSH-Px were higher and lipid peroxidation lower in acetyl-l-carnitine treated lenses, affirming its protective function in selenite-induced cataractogenesis [116]. Propolis, a compound produced by bees, reduced reactive oxygen species and increased cell viability in rat epithelial cell cultured with glucose, implicating the role of propolis in inhibiting hyperglycemia insult-initiated oxidative and osmotic stress [117].
Plant-derived compounds and herbal extracts
There is a large body of evidence supporting the anti-cataract potential of numerous medicinal plants, presumably via mechanisms that involve antioxidant potency of these plant-derived compounds and/or extracts. Quercetin, a flavonoid that is present in many fruits, vegetables, and grains, has been identified as a potent antioxidant and anticancer agent [118,119]. Quercetin prevented cataract progression and formation in glucose-induced goat lenses incubated in artificial aqueous humor in a manner that was characterized by a decrease in lipid peroxidation and an increase in Na+-K+-ATPase activity [91]. Chrysin, a flavone found in honey, prevented selenite-induced cataract formation in cultured Wister rat lenses by modulating genes responsible for calcium transport, calpain activation, and apoptosis [120]. Chaudhury et al. (2017) also showed that flavonoids from green tea prevented UV radiation-induced oxidative damage to human γB-crystallin, in vitro [121]. Moreover, a polyphenol derived from green tea, epigallocatechin-3-gallate (EGCG) attenuated and reversed the aggregation of αA(66–80), a peptide fragment derived from αA-crystallin peptide. Since αA(66–80) aggregates play a role in the destabilization of native crystallin structures, EGCG can potentially prevent onset and reverse cataract formation [122]. In corroboration, green tea (Camellia sinensis) leaf extract mitigated Na selenite-induced reduction on SOD, GSH-Px and CAT activities, and increase in lipid peroxidation in Wistar rat lenses, ex vivo [123]. In general, these studies suggest a protective role for natural polyphenols and flavonoids in cataract formation.
Drevogenin D is a triterpenoid aglycone derived from Dregea volubilis, a plant which belongs to the Asclepiedaceae family. Extracts from Dregea volubilis have found medicinal application for various conditions such as inflammation, asthma, dyspepsia, tumors [124,125,126]. Biju et al. (2007) demonstrated an anti-cataract activity of the triterpenoid aglycone, drevogenin D from Dregea volubilis, in a selenite-induced Sprague–Dawley rat cataract model, ex vivo. Not only did the presence of drevogenin D mitigate selenite-induced inhibition of SOD, CAT, and GSH-Px enzyme activities, it also elicited an increase in GSH and sulfhydryl levels and a decrease in lipid peroxidation [127].
Fenugreek (Trigonella foenum-graecum), a leguminous plant that is used globally as a spice, has been shown to possess multiple beneficial medicinal properties such as antidiabetic, anticarcinogenic, hypocholesterolemic, antioxidant, and immunological effects [128]. Moreover, the extracts of fenugreek seed have been shown to possess potent antioxidant and free radical scavenging activities in vitro and in vivo [129,130]. Using Wistar rat lenses, Gupta et al. (2010a) demonstrated that the aqueous extract of fenugreek (Trigonella foenum-graecum seeds) could sustain lens transparency and restore lenticular activities of SOD, GSH-Px, CAT, and GST enzymes in a selenite-induced lens model. Moreover, this plant extract also blocked selenite-induced depletion of GSH and higher lipid peroxidation in Wistar rat lenses, ex vivo [131]. Using the same experimental model, Gupta and co-workers also found that the aqueous extract of the herbal preparation Triphala, made from three fruits of Asian origin, Emblica officinalis, Terminalia chebula, and Terminalia belerica protected cultured rat lenses from selenite-insult by maintaining optical clarity, restoring the activity of SOD, GSH-Px, CAT, and GST and attenuating lipid peroxidation [132]. Although the exact composition of Triphala is variable, this herbal mixture has been reported to exert multiple beneficial effects in humans [133].
Moringa oleifera, a plant belonging to the Moringaceae family, has found therapeutic application in a wide range of conditions including epilepsy, fever, hypertension, inflammation, tumors, hyperlipidemia, and diabetes. Due to enrichment of vitamins A and C and minerals, the Moringa oleifera plant cans serve as a nutritional supplement and a source of natural antioxidants in malnourished individuals [134,135]. The ethanol extract of Moringa oliefera prevented lens opacification of hyperglycemia (55 mM)-induced cataract formation up to 72 h. Moreover, Moringa oleifera extracts restored GSH, soluble protein content, CAT activity, and mitigated lipid peroxidation, ex vivo [136].
Abutilon indicum, a plant that belongs to the Malvaceae family, is an indigenous shrub found in tropical and subtropical climates in Asia. It is used as a herbal remedy for a wide range of ailments such as fever, infection, and inflammation [137,138,139]. Hydroethanolic extracts of Abutilon indicum effectively attenuated lens opacification in glucose (55 mM)-induced cataract formation in cultured goat lenses. In these studies, the extract-treated lenses displayed a reduction in malondialdehyde content and an increase in total protein content and SOD activity. Moreover, the extracts attenuated aldose reductase activity in hyperglycemia-treated lenses, thereby affirming its anti-cataract activity in an acute diabetic cataract lens model, ex vivo [140]. In other studies, the ethanol extract of ginger (Zingiber officinale), a well-known spice, delayed hyperglycemia-induced opacification in cultured goat lenses, ex vivo. The ginger extract treated lenses also displayed a higher Na+-K+-ATPase activity and total protein content, and a reduction in lipid peroxidation [141].
Hippophae rhamnoides L. is a deciduous shrub of the family Elaeagnaceae that is also known as Seabuckthorn. Although exact composition varies with geographical location of the plant, the seabuckthorn plant is enriched with minerals, vitamins, and micronutrients with medicinal and nutritional value [142]. Leaf extracts of seabucthorn have been shown to possess potent antioxidant activity in C-6 glioma cells, in vitro [143]. The aqueous extract of seabuckthorn leaf delayed initiation and progression of H2O2-induced cataract formation and restored GSH, SOD activity in goat lenses, ex vivo [144].
Abies pindrow, commonly known as west Himalayan fir, is an evergreen tree that belongs to the Pinaceae family. Leaf extracts of Abies pindrow with demonstrated potent antioxidant activity [145], have been used as a herbal remedy for various ailments, including fever, diabetes, bronchitis, and antispasmodic [146]. Dubey et al. (2015a) investigated the anti-cataract activity of the aqueous leaf extract of Abies pindrow on H2O2-induced cataract formation in goat lenses, ex vivo. These investigators reported that the herbal extract prevented clouding caused by peroxide insult and enhanced GSH and total protein contents and SOD activity, when compared to peroxide-only treated lenses [147]. In another study, Dubey et al. (2015b) further delineated the role of Luffa cylindrica on cataract formation in goat lenses, ex vivo. Also known as sponge gourd, Luffa cylindrica is a fibrous plant belonging to the family Cucurbitaceae. In addition to being consumed globally as a vegetable, the fruit possesses potent antioxidant properties and is used as a traditional medication for inflammation and diabetes management, amongst others [148,149,150]. Dubey (2015b) and coworkers showed that a standardized preparation of Luffa cylindrica fruit extract protected goat lenses from H2O2-induced opacification, lipid peroxidation, and enhanced total protein content, ex vivo [151].
Ocimum tenuiflorum (also known as Ocimum sanctum) is a perennial plant that is indigenous to Indian subcontinent that belongs to the Lamiaceae family. Ocimum sanctum is used as a herbal remedy for various ailments, including inflammation (e.g., arthritis), respiratory problems (e.g., cough, asthma, bronchitis), malaria, stomach problems (e.g., diarrhea), and ocular pain in some Asian nations [152]. Gupta et al. (2005) found that aqueous extracts of Ocimum sanctum could mitigate selenite-induced damage to proteins and antioxidant enzymes in rat lenses, ex vivo [153].
In summary, a wide range of compounds and herbal mixtures that possess antioxidant properties have been shown to protect lenses from oxidative damage, ex vivo and in vitro. For these compounds to be clinically significant, these studies need to be replicated in vivo. Furthermore, extensive chemical characterization of the active compounds in the plant-derived products needs to be performed to determine their pharmacological mechanism/s of action. Table 3 provides a summary of antioxidant drugs that elicit an anti-cataract effect on various models of the disease, in vitro and ex vivo.
Table 3.
Summary of in vitro and ex vivo studies of cataractogenesis.
(ii) In vivo studies
Various experimental animal models, including the inducible or genetic models of cataracts have been employed in studies of cataractogenesis, in vivo [154,155,156]. The induced cataract types include models for oxidant stress-induced (selenite, naphthalene, I-buthionine-(S,R)-sulfoximine (BSO), hyperbaric oxygen), diabetic cataract (streptozotocin, galactose, and rat strains with spontaneous onset diabetes); UV-induced cataract (exposure to UVA and UVB) and steroid-induced opacification in different animal species [154,155,156]. Using these in vivo models, different antioxidants have been evaluated for their anti-cataract activity.
Typical antioxidants
Lenticular vitamin C is a crucial antioxidant and ultraviolet-filter that mitigates entry of light rays into the lens, thereby minimizing consequences of oxidative damage to the lens [157,158]. Insufficient consumption of vitamin C elicited a reduction in lenticular levels of ascorbic acid in guinea pigs [159], while dietary intake of the vitamin was found to increase lenticular ascorbic acid levels in rats [160,161]. Peighmbarzadeh and Tavana (2014) investigated the effect of vitamin C on selenite-induced cataractogenesis in New Zealand albino rabbits, in vivo. In this study, cataract was induced by a single subcutaneous injection of selenite (20 μmol/kg) on the 10th postpartum day. Daily subcutaneous injection of vitamin C (2.5 mg/rabbit) from the eighth day (postpartum) for 14 days attenuated cataract formation in 40% of animals [162]. In support of these observations, biochemical studies revealed a higher level of total soluble protein content compared to selenite-treated animals, thereby affirming its anti-cataract activity [162]. Using a similar experimental model, Jahadi Hosseini et al. (2008) demonstrated a 50% protective action for vitamin C (0.3 mM) from selenite-induced cataract formation in Sprague–Dawley rat pups, in vivo. Biochemical assays confirmed a higher concentration of total proteins and soluble proteins in vitamin C-treated animal lenses, compared to that of selenite-only treated animals [163]. Moreover, vitamin C attenuated galactose-induced cataract formation in guinea pigs [164]. Recently, this vitamin was reported to mitigate selenite-induced cataract formation and the corresponding loss in chaperone activity in Sprague–Dawley rat pups, in vivo [165]. Taken together, these studies suggest a partial protective role of vitamin C in experimental cataractogenesis, thereby affirming the multifactorial etiology of the disease. On the contrary, higher levels of vitamin C have been shown to serve a pro-oxidant role in mammalian lens, presumably due to its oxidative stress metabolites such as dehydroascorbic acid [166].
The fat-soluble vitamin, vitamin E, has also been reported to serve as a crucial antioxidant in the lens defense system. Not only is a deficiency of vitamin E linked to an increase in the risk of cataractogensis in rats [167,168], but its anti-cataract activity has been demonstrated against selenite-, galactose-, steroid-, streptozocin- and UV radiation-induced cataract in experimental animals [165,167,168,169,170,171]. Amongst possible mechanisms, vitamin E has been shown to elevate GSH levels and protect the chaperone activity of crystallin proteins, thereby preserving the antioxidative defense mechanism within the lens [165,169].
Khan et al. 2017 showed that the potent, endogenous disulfide antioxidant, α-lipoic acid, prevented the onset and progression of fructose-induced cataract in Sprague–Dawley albino rats. These workers also found an increase in the level of several antioxidants (GSH-Px, CAT, SOD, and GSH) while lipid peroxidation was reduced, compared to that of fructose-only treated animals. Interestingly, α-lipoic acid treatment reinstated lens protein Ca2+ ATPase and Ca2+ in the lens [172]. In other studies, the anti-cataract effect of α-lipoic acid together with pantethine, penicillamine, and deferoxamine were attributed to their antioxidant effects [173,174,175,176].
The potent pyridoindole antioxidant and free radical scavenger, stobadine, deferred the progression of streptozocin-induced diabetic cataract in Wistar rats by blocking lipid peroxidation, in vivo [171,177]. In other studies, the endogenous free radical scavenger, melatonin, attenuated BSO-induced cataract formation in neonatal rats in vivo and enhanced total lenticular GSH content, suggesting that either the free radical scavenging and/or a stimulatory effect on GSH content was responsible for the anti-cataract activity [178]. Taken together, these studies affirm the protective role of antioxidants on the pathophysiology of the lens, in vivo.
The dietary minerals, zinc and copper, have been reported to prevent cataract formation. Indeed, copper and zinc are crucial for the activity of the lenticular antioxidant systems such as the cytosolic SOD-1 enzyme [179,180]. Low lenticular concentrations of zinc have been observed in patients with mature age-related cataracts and high levels in those with traumatic cataracts [181]. In a recent study, dietary supplementation with zinc for a period of six weeks mitigated the formation of streptozocin-induced cataracts in experimental rats [182]. In addition to enhanced levels of α-crystallin proteins, zinc supplementation attenuated the lenticular expression of polyol pathway enzymes [182]. Topical application of zinc sulfate eye drops has also been reported to attenuate lens opacity induced by intravitreal injection of sodium selenite in adult rabbits, in vivo [183], supporting an important role for zinc in lens clarity.
Although selenium is a trace element that plays a significant role in lenses where it supports the activity of antioxidant selenoproteins such as GSH-Px [184], toxic levels of this element produce oxidant stress-mediated cataract formation, leading to its application in the induction of experimental cataracts [154,156]. Karakucuk et al. observed a decrease in the concentration of selenium in the aqueous humor and lens of patients with age-related cataracts, an important finding that supports the significant role of selenium in the activity of GSH-Px enzyme in maintaining lens clarity [185]. Ebselen, an organoselenium molecule that possesses antioxidant, neuroprotective, and anti-inflammatory activities, protected Sprague–Dawley rat pups from selenite-induced cataracts, in vivo [186,187]. Moreover, the ebselen-treated lenses exhibited clearer lenses, higher lenticular GSH content, and less lipid peroxidation when compared to that of selenite-only treated animals [187]. Taken together, these studies confirm the vital antioxidant role of trace elements in maintaining lens transparency, in vivo.
Ketoacids and amino acids antioxidants
The ketoacid, pyruvate, attenuated the progression of streptozotocin- and selenite-induced cataracts in experimental animal models, in vivo [105,188]. In addition to exhibiting anti-cataract activity, ex vivo [112], N-acetylcysteine, a prodrug for the amino acid and thiol antioxidant l-cysteine, has been shown to replicate its anti-cataract activity in vivo. N-acetyl-cysteine inhibited selenite-induced nuclear opacities in Sprague–Dawley rats, in vivo [189]. Lenticular malondialdehyde and protein carbonyl levels were lower in lens homogenates from N-acetylcysteine-treated rats while GSH content was significantly higher compared to that of control animals [189]. Tuzcu et al. (2014) sought to determine the role of intravitreal triamcinolone acetonide injection on cataract formation in albino Wistar rats, in vivo. Although there was no visible lens opacification in response to steroid exposure, N-acetylcysteine protected lenses from the steroid-induced decline in GSH and GSH-Px activity and a corresponding decline in malondialdehyde and protein carbonyl content, compared to those of control animals [190].
N-acetylcysteine amide is a more lipophilic derivative of N-acetylcysteine, with superior bioavailability into intracellular sites. Both N-acetylcysteine amide and glutathione ethyl ester eye drops delayed onset of streptozotocin-induced cataract formation in Sprague–Dawley rats, as evidenced by the high level of thiol and catalase activity in lens homogenates. Interestingly, both drugs were unable to delay progression or reverse lens opacity in the advanced stages of the disease [191], presumably due to the depletion of the total lenticular antioxidant pool. Maddirala et al. (2017) also investigated the effect of both topically- and intraperitoneally-administered N-acetylcysteine amide on selenite-induced cataract formation in male Wistar rat pups [192]. In this experimental model, Na selenite (19 μM/kg body weight) was administered intraperitoneally on 10th postpartum day, intraperitoneal N-acetylcysteine amide (250 mg/kg body weight) on ninth, 11th and 13th postpartum days, while topical N-acetylcysteine drops (1%) was instilled into the eyes from postpartum 15th and 30th days. Interestingly, both intraperitoneal and topical N-acetylcysteine amide attenuated the severity of cataracts formation, in vivo. Furthermore, the amide-treated lens homogenates exhibited higher levels of GSH content, thioltransferase and m-calpain activity, while lipid peroxidation, GSH-Rx activity, and calcium levels were reduced when compared to selenite-only treated lens homogenates [192]. Carey et al. (2011) also investigated the effect of N-acetylcysteine amide on cataracts induced by the GSH synthesis inhibitor, BSO, in a Wister rat pup model of the disease. Slit lamp examination of animal lenses revealed that N-acetyl-cysteine amide prevented cataract formation in 80% of lenses. Biochemical studies revealed a reduction in lenticular protein carbonylation and lipid peroxidation while antioxidant defense enzymes were replenished, compared to that of BSO-only treated lenses [193]. In agreement with data from ex vivo studies [116], intraperitoneal administration of acetyl-l-carnitine prevented selenite-induced cataractogenesis in Wister rat pups, via mechanisms involving inhibition of lipid peroxidation and restoration of antioxidant content and activity, in vivo [194]. N-acetylcarnosine, a prodrug of l-carnosine also attenuated onset and progression of age-related cataracts in humans and dogs [195], corroborating its effect in lenticular epithelial cells, propolis, delayed onset and progression of galactose-induced cataract Sprague–Dawley rats, in vivo [117].
Plant-derived compounds and herbal remedies
Plant-derived compounds and herbal remedies have also been reported to demonstrated anti-cataract potential in studies using experimental animals with this disease. The fruit and vegetable-derived flavonoid, quercetin, is a potent antioxidant and free radical scavenger that possess several health benefits, including cardioprotective, anti-diabetic, anti-inflammatory, and anti-cancer effects, amongst others [196]. As observed in ex vivo studies [91], quercetin prevented the onset and progression of selenite-induced cataracts and sustained lens chaperone activity in Sprague–Dawley rats, in vivo [165]. In other studies, intraperitoneal injection of a citrus flavonoid, rutin blocked selenite-induced lenticular opacification in Wistar rats, with a corresponding elevation in the activity of the antioxidant enzymes, CAT, SOD, GSH-Px, GST, and GSH-Rx and a reduction in lipid peroxidation, compared to selenite-only treated lenses [197].
Hesperetin is a flavonoid found in orange rinds that has been reported to elicit beneficial cardiovascular and anti-inflammatory action effects [198,199]. Hesperetin prevented selenite-induced cataract formation in rats by attenuating the loss of the lens protein, filensin, and replenishing GSH and ascorbic acid levels [200]. Furthermore, hesperetin and its derivatives, hesperetin stearic acid ester and hesperetin oleic acid ester, counteracted the decrease in α-crystallin chaperone activity due to selenite, in vivo [201,202].
Ellagic acid, a polyphenol found in several plants and fruits such as grapes, nuts, strawberries can elicit antiproliferative and antioxidant effects, in vitro and in vivo [203,204,205]. Sakthivel et al. (2008) showed that ellagic acid mitigated selenite-induced opacification in Wistar rats, in vivo. Ellagic acid treated lenses exhibited superior CAT, GSH-Px, SOD, GST activities, compared to selenite-only treated control lenses. Moreover, malondialdehyde and calcium level was lower in ellagic acid treated lenses compared to untreated selenite group [206].
The leaves and leaf buds of the evergreen shrub, Camellia sinens of the family Theaceae, have been used for centuries to produce the beverage known as tea. Both green tea and black tea, which are rich in polyphenols, prevented the onset and progression of lens opacification by restoring antioxidants in selenite-induced and streptozotocin-induced cataract [121,123,207,208]. The biological effects of caffeine, a methylxanthine alkaloid found in plants and used as a beverage worldwide, has been extensively described in literature [209,210]. There is evidence supporting a protective role for caffeine from UV radiation-induced cataractogenesis, in vivo. Indeed, topically administered caffeine mitigated UV radiation-induced cataractogenesis and lens apoptosis in Sprague–Dawley rats [211,212,213]. Moreover, coffee delayed onset of selenite-induced cataract in rats and preserved antioxidant enzymes and lens protein [165,214]. Caffeine and pyrocatechol also sustained lens chaperone activity in selenite-induced Sprague–Dawley rat lenses, in vivo [165].
Other dietary antioxidants have been extensively explored in the prevention of cataract formation, in vivo. The carotenoids (α-carotene, β-carotene, lutein, lycopene, and cryptoxanthin) were shown to elicit a protective action from both oxidative stress- and hyperglycemia-induced cataract formation, in vivo [165,215,216,217]. Nakazawa et al. (2017) also demonstrated that both oil-soluble (e.g., β-carotene, lutein, zeaxanthin, anthocynain) and water-soluble (e.g., cyanidin) antioxidants could impede the onset and progression of selenite-induced cataracts and sustain lens chaperone activity in Sprague–Dawley rats, in vivo [165]. Curcumin is a bright yellow compound with antioxidant properties and is produced from the Curcuma longa plant belonging to the Zingiberaceae family. Curcumin delayed the initiation and progression of galactose-, oxidative stress- and streptozotocin-induced cataract formation, in vivo, by mechanisms that involved the preservation of lenticular antioxidant, lipid peroxidation, and soluble protein content [218,219,220,221].
Resveratrol (3,5,4′-trihydroxy-trans-stilbene) is a natural phenolic stilbenoid present in the skin of grapes, blueberries, raspberries, mulberries, and peanuts [222,223]. Resveratrol has been reported to possess health benefits in several chronic diseases such as diabetes mellitus, various cancers, metabolic syndrome, cardiovascular diseases, and Alzheimer’s disease, to mention a few [223,224]. Resveratrol prevented selenite-induced cataract in Sprague–Dawley rat pups, as evidenced by the optical clarity of lenses and higher GSH and reduction in lipid peroxidation in resveratrol-treated lenses [225].
Heliotropium indicum, also known as Indian heliotrope, is a plant that belongs to the family Boraginaceae. In some parts of Asia, its leaf extracts are used as a traditional remedy for wounds, skin ulcers, conjunctivitis and cataracts, amongst others [226,227]. Whole plant aqueous extracts of Heliotropium indicum prevented selenite-induced cataractogenesis in Sprague–Dawley rats, in vivo. In addition to preserving aquaporin 0, crystallins (αA &αB), total lens proteins, and GSH contents, its aqueous extracts attenuated lipid peroxidation, suggesting an antioxidant basis for its anti-cataract activity [228].
Echium amoenum, a perennial plant that also belongs to the Boraginaceae family, has been shown to possess antimicrobial, antidiabetic, and anxiolytic properties, amongst others [229]. Hydroalcoholic extract of Echium amoenum also elicited a protective effect against selenite-induced lens opacification in rats, in vivo. These investigators concluded that the antioxidant phytochemicals within the plant accounted for the anti-cataract activity of Echium amoenum [230].
IH636 grape seed proanthocyanidin extract (GSPE), an oligomeric proanthocyanidin extracted from grape seeds, has been shown to possess potent free radical scavenging and antioxidant properties with several potential health benefits [231,232]. IH636 GSPE attenuated cataract formation in Sprague–Dawley rat pups, most likely through the antioxidant actions of the extract [233].
Cassia tora, also known as sennatora, is a herbaceous plant that belongs to the Fabaceae family, has a wide application ranging from edible vegetable, food additive, medicinal (laxative, treatment for skin disease, antiparasitic) to industrial (cassia gum—a thickener and gelling agent) use [234]. Cassia tora leaves delayed the onset and progression of cataract formation in neonatal rats. In addition to preserving the antioxidant defense system, lens morphology and the expression of lens crystallins, Cassia tora attenuated lipid peroxidation, compared to selenite-only treated lenses.
Vaccinium corymbosum (northern highbush blueberry), a deciduous shrub that belongs to the family Ericaceae, has been shown to be rich in vitamin C and polyphenol antioxidants [235]. A decoction of Vaccinium corymbosum leaf attenuated the onset and progression of selenite-induced cataracts in neonatal rats [236]. Lenses from the decoction-treated animals exhibited significantly higher levels of antioxidant markers and soluble proteins while protein aggregation and activation of calpain were mitigated [236].
Emilia sonchifolina (also known as “lilac tassel flower”), a tropical plant that belongs to the Asteraceae family, is used as a herbal remedy for ocular ailments, fever, sore throat, diarrhea, eczema, and as an antidote for snake bites in some Asian countries [237,238]. Lija et al. (2006) found that Emilia sonchifolia-derived flavonoids prevented the initiation and progression of selenite-cataract formation, in vivo. In addition to enhancing activities of SOD, CAT, GSH enzymes, the flavonoids reduced lipid peroxidation compared to selenite-only treated animals, suggesting that antioxidant mechanisms account for the anti-cataract effect of Emilia sonchifolia-derived flavonoids.
Broccoli (Brassica oleracea var. italic), a common vegetable that belongs to the family Brassicaceaeis, is naturally enriched with nutritional antioxidants and flavonoids [239]. Broccoli has been reported to possess several health benefits such as decreasing the risk of cancer [240]. Broccoli-derived flavonoid fraction attenuated the onset and progression of selenite-induced lens opacification with corresponding higher GSH content, activity of antioxidant enzymes (CAT, SOD) and a reduction in lipid peroxidation, calcium, and calpains when compared to selenite-only treated animals [241].
Lupeol, a pentacyclic triterpenoid present in the flavonoid fraction of Vernonia cinereal plant that belongs to the Asteraceae family, possesses antimicrobial, anti-inflammatory, and anticancer activities [242]. Asha et al. (2016) demonstrated an anti-cataract action for lupeol in selenite-induced Sprague–Dawley rat pups, in vivo. Compared to selenite-only treated animals, lupeol attenuated selenite-induced lipid peroxidation as well as the decline in the activity of the antioxidant enzymes, CAT, GSH-Px, GSH-Rx, GST, and Ca2+ATPase [243].
Allium sativum (garlic) and Allium cepa (onion) are bulbous plants that belong to the Amaryllidaceae family. Commonly used as a seasoning worldwide, the therapeutic potential of garlic has been reported to include anti-tumor, cardioprotective, anti-diabetic, antimicrobial, amongst others [244]. In addition to delaying onset of streptozotocin-induced cataract in Wistar rats, the methanolic Allium sativum (garlic) extract mitigated hypoglycemia-induced oxidative stress associated with the disease in a dose-dependent manner and restored lenticular SOD and GSH-Px activities and GSH content [245]. In other studies, the flavonoid-rich, aqueous extract of Allium cepa (onion), preserved optical clarity in selenite-treated Wistar Albino rats, in vivo. Moreover, the extract treated lenses exhibited higher total antioxidant level, GSH content, and higher activity of GSH-Px and SOD enzymes compared to untreated lenses, in vivo [246].
Pinus densiflora, a pine tree that belongs to the family Pinaceae, is naturally enriched with antioxidant procyanidins and polyphenols [247]. Pinus densiflora has been used as a traditional remedy for inflammation and pain in some Asian countries [248]. The bark extract of Pinus densiflora prevented selenite-induced cataract development in Sprague–Dawley rat pups in a dose-dependent manner [249], with a corresponding enhancement of lenticular water-soluble protein and GSH contents; antioxidant activities of SOD, GSH-Px, and CAT enzymes; and a reduction in water-insoluble protein content and lipid peroxidation. Furthermore, the extract counteracted the selenite-induced downregulation of αA-crystalline, lens-specific m-calpain (Lp84), filensin and phakinin, and the antiapoptotic factor (Bcl-2), and the upregulation of apoptotic proteins [249]. In other studies, Ocimum sanctum delayed the onset and incidence of selenite-induced cataracts, and preserved antioxidants and soluble protein contents in rat lenses, in vivo [153].
Crocus sativus, commonly known as saffron crocus, is a flowering plant that belongs to the Iridaceae family. The Crocus sativus stigma, which is enriched with nutrients such as flavonoids, crocetin, and crocin is widely used as a spice and a traditional medicinal remedy for various ailments such as stomach pain [250]. Aqueous extract of Crocus sativus stigma prevented selenite-induced cataracts in rats and enhanced the antioxidant enzymes SOD, CAT, GSH-Px, and GSH content. Additionally, the extract attenuated lenticular selenite-induced lipid peroxidation, protein oxidation, and proteolysis, thereby affirming the antioxidant-based mechanism for the anti-cataract action of the plant [251].
The plants, Pterocarpus marsupium and Trigonellafoenum-graceum, which belong to the Fabaceae family are found to have traditional application as anti-diabetic remedies [252]. Therefore, Vat et al. (2004) sought to investigate the possible application of these plants to prevent diabetic cataracts and found that the aqueous extracts of the bark of Pterocarpus marsupium and the alcoholic extract of the Trigonellafoenum-graceum seeds displayed an anti-cataract effect in alloxan-diabetic rats, in vivo. Moreover, the anti-cataract potency was directly proportional to the hypoglycemic potency of the plants, leading the authors to conclude that the hypoglycemic capacity of the plants was responsible for the anti-cataract effect observed [253].
Ginkgo biloba, a tree from the family Ginkgoaceae, has been used for centuries as a traditional remedy for various ailments such as cough and asthma in some Asian countries [254,255]. Oral supplementation with an extract of Ginkgo biloba delayed the onset and progression of cranial radiation-induced cataract formation and abolished the corresponding reduction in SOD and GSH-Px activities and increase in lipid peroxidation in Sprague–Dawley rats, in vivo [256]. For further details, the reader is referred to the rigorous review by Thiagarajan and Manikandan, focused on the anti-cataract activity of vitamin C, vitamin E, and curcumin along with prototype plant materials with antioxidant activities tested against cataracts [257].
Gupta et al. (2010) investigated the pharmacological actions of Triphala, consisting of Emblica officinalis, Terminalia chebula, and Terminalia belerica on selenite-induced cataract in Wistar rats in vivo. In corroboration with its ex vivo effects, the herbal combination blocked the progression and intensity of selenite-induced cataracts, in vivo [132]. In summary, various plant extracts and products that possess antioxidant properties have tremendous potential in the development of anti-cataract medications. Table 4 provides a summary of antioxidant drugs that elicit an anti-cataract effect on various models of the disease, in vivo.
Table 4.
Summary of in vivo studies of cataractogenesis.
4.2.2. Non-Steroidal Anti-Inflammatory Drugs (NSAIDs)
In the 80′s NSAIDs emerged as potential anti-cataract agents after aspirin was found to have anti-cataract effect in a rheumatoid arthritis study [258]. Subsequently, aspirin and variety of NSAIDs such as ibuprofen, naproxen, sulindac, bendazac, 5-hydroxybendazac, and bendazac-lysine were pursued for their anti-cataract activity [259,260,261,262,263,264,265,266,267,268]. These observed anti-cataract activities of NSAIDS were due to different mechanisms such as acetylation, inhibition of glycosylation, and carbamylation of lens proteins [259]. Sulindac possessed anti-cataract activity, possibly due to aldose reductase inhibition [269] while naproxen had effect on galactose-induced and selenite-induced cataract in rats [270,271]. Naproxen reversed depletion of GSH and high malondialdehyde levels due to oxidant stress induced lipid peroxidation in isolated rat lens [272]. Although some studies implicate the inhibition of aldose reductase for the anti-cataract activity of NSAIDs other studies indicate antioxidant properties, suggesting an array of pathways [7,259,260,261,262,263,264,265,266,267,268,269,270,271]. However, no recent studies have surfaced in the past few years in supporting an anti-cataract role for NSAIDs besides their common post-cataract surgery use to prevent cystoid macular edema [273,274].
4.2.3. Miscellaneous Drugs
In addition to antioxidants, various miscellaneous compounds have exhibited an anti-cataract effect. Pharmacological chaperons that bind α-crystallins (cryAA and cryAB) reversed protein aggregation in various models of cataracts. For example, compound 29 (5-cholesten-3b,25-diol) prevented protein aggregation and enhanced protein solubility, thereby improving lens clarity in severe age-related cataracts (R120G cryAB knock-in mice), hereditary cataract due to a mutation (R49C cryAA knock-in mice), and age-related cataract (C57BL/6J wild-type mice) [275].
Angiotensin-converting enzyme (ACE) inhibitors, lisinopril and enalapril, have been reported to elicit anti-cataract activities in glucose-induced cataract, probably due to the antioxidant and free radical scavenging mechanisms [276]. Enalapril and lisinopril were also able to retard the glucose (55 mM)-induced opacification in goat lenses cultured in artificial aqueous humor, ex vivo. Contrary to ACE-treated lenses in which Na+-K+-ATPase activity and total protein content was higher, the glucose-only treated lenses displayed a higher level of lipid peroxidation [276].
Ursodeoxycholic acid, a component of bear bile, similarly prevented selenite-induced cataract formation on M-199 cultured Wister rat lenses, ex vivo [277]. Hydrogen saline and ursodeoxycholic acid were able to elevate the total antioxidative capabilities and activity of SOD, CAT, GSH-Px, GSH-Rx, and GST compared to untreated lenses following selenite insult, ex vivo. Moreover, the contents of sulfhydryl groups and GSH were higher in hydrogen saline- and ursodeoxycholic acid-treated lenses as compared to the selenite-only treated group, which displayed higher lenticular malondialdehyde [277,278].
Posterior capsular opacification is a common secondary complication to cataract surgery that involves intraocular lens implantation with an artificial intraocular lens [279]. Sternberg et al. (2010) investigated a method to prevent secondary cataract in New Zealand rabbits after implantation of intraocular lens, in vivo [280]. Compared to control animals that developed opacification in six weeks, these investigators demonstrated that treatment with methotrexate and actinomycin D mixture loaded hyaluronic acid delayed opacification up to six months, postoperatively [280]. In other studies, a sustained cyclosporin A delivery microsphere prevented postoperative posterior subcapsular cataract development in New Zealand white rabbits, in vivo [281]. Brown and Akaichi (2015) observed an association between vitamin D deficiency and posterior subcapsular cataract patients and suggested that intake of vitamin D might reduce incidence of posterior subcapsular cataracts [282]. In corroboration, lanosterol, a crucial intermediate in the biosynthesis of steroids and vitamin D, reduced protein aggregation due to mutant crystallin in in vitro tests and cell transfection methods. Moreover, lanosterol attenuated cataract and increased lens transparency in cataractous lenses from rabbits and dogs, thus diminishing severity of cataract, in vivo [283]. Taken together, these studies suggest a significant role for vitamin D in lens transparency.
5. Conclusions
Although cataract has been the leading cause of preventable blindness worldwide for many decades, pharmacological strategies to mitigate, prevent, or cure this blinding disease have remained elusive. With the projected increase in life expectancies, the number of people affected with cataract is predicted to increase worldwide. The prognosis of the disease is at best oblique in low income nations that lack easy and affordable access to cataract surgery. Therefore, the potential therapeutic and economic benefits of pharmacological cataract treatments are immeasurable, ranging from a reduction in economic burden to better quality of life. This review presents a wide range of compounds, including antioxidants and herbal remedies/extracts that have exhibited anti-cataract activity in vitro, ex vivo, and in vivo. So far, compounds that possess antioxidant and free radical scavenging activity have shown tremendous potential in these experimental studies. The ready accessibility of plant-derived compounds/mixtures renders them an attractive treatment choice, especially in developing nations where cost is likely to hinder the development and patient use of medication. However, none of these drug candidates have translated into US FDA-approved anti-cataract eye drops or remedies that can prevent, deter, or cure cataracts in humans. Therefore, further research is necessary to establish efficacy and safety profile of these herbal remedies and antioxidants, standardize drug formulations, and conduct double-blinded studies to justify clinical application in humans. It is interesting to note that the Age-Related Eye Disease Study 2 (AREDS2) failed to demonstrate a significant deterrent in the progression of age-related cataract using lutein/zeaxanthine supplementation [284]. Similarly, an extensive Cochrane review did not identify a significant anti-cataract effect of vitamin supplementation in humans [285]. It is conceivable that improvement in drug delivery strategies could enhance anti-cataract efficacy in the prevention and/or treatment of cataracts. Additionally, there is a need to delineate the exact mechanisms involved in the process of formation of cataracts in order to identify new therapeutic targets and new drug candidates that can be refined for therapeutic and/or prophylactic use in humans.
Author Contributions
All authors contributed to the writing, editing and review of this article. All authors have read and agreed to the published version of the manuscript.
Funding
This project received no external funding.
Acknowledgments
We acknowledge the Pharmaceutical Sciences Masters Program, Department of Pharmacy Sciences, School of Pharmacy and Health Professions, Creighton University for their support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ACE | Angiotensin converting enzyme |
| ATPase | Adenosine triphosphatase |
| BSO | Buthionine sulfoximine |
| CAT | Catalase |
| CNS | Central nervous system |
| Cu | Copper |
| EC | Extracellular |
| EGCG | Epigallocatechin-3-gallate |
| GSH | Glutathione |
| GSH-Px | Glutathione peroxidase |
| GSPE | Grape seed proanthocyanidin extract |
| GSH-Rx | Glutathione reductase |
| GSSG | Glutathione disulfide |
| GST | Glutathione-S-Transferase |
| K+ | Potassium cation |
| Mn-SOD | Mitochondrial-superoxide dismutase |
| Na+ | Sodium cation |
| Na+-K+-ATPase | sodium-potassium adenosine triphosphatase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NSAIDs | Non-steroidal anti-inflammatory drugs |
| SOD | Superoxide dismutase |
| TGF-β | Transforming growth factor beta |
| UV | Ultraviolet |
| UVA | Ultraviolet A |
| UVB | Ultraviolet B |
| Zn | Zinc |
References
- Lee, C.M.; Afshari, N.A. The global state of cataract blindness. Curr. Opin. Ophthalmol. 2017, 28, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Khairallah, M.; Kahloun, R.; Bourne, R.; Limburg, H.; Flaxman, S.R.; Jonas, J.B.; Keeffe, J.; Leasher, J.; Naidoo, K.; Pesudovs, K.; et al. Number of people blind or visually impaired by cataract worldwide and in world regions, 1990 to 2010. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6762–6769. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.; Cumming, R.G.; Attebo, K.; Panchapakesan, J. Prevalence of cataract in Australia: The Blue Mountains eye study. Ophthalmology 1997, 104, 581–588. [Google Scholar] [CrossRef]
- Chua, J.; Koh, J.Y.; Tan, A.G.; Zhao, W.; Lamoureux, E.; Mitchell, P.; Wang, J.J.; Wong, T.Y.; Cheng, C.Y. Ancestry, socioeconomic status, and age-related cataract in Asians: The Singapore Epidemiology of Eye Diseases Study. Ophthalmology 2015, 122, 2169–2178. [Google Scholar] [CrossRef]
- Varma, R.; Torres, M. Prevalence of lens opacities in Latinos: The Los Angeles Latino Eye Study. Ophthalmology 2004, 111, 1449–1456. [Google Scholar] [CrossRef]
- National Institute of Health, National Eye Institute. Cataracts. Available online: https://nei.nih.gov/eyedata/cataract (accessed on 20 August 2019).
- Gupta, S.K.; Selvan, V.K.; Agrawl, S.S.; Saxena, R. Advances in pharmacological strategies for the prevention of cataract development. Indian J. Ophthalmol. 2009, 57, 175–183. [Google Scholar] [CrossRef]
- Thompson, J.; Lakhani, N. Cataracts. Prim. Care 2015, 42, 409–423. [Google Scholar] [CrossRef]
- World Health Organization. Cataract. 2019. Available online: https://www.who.int/blindness/causes/priority/en/index1.html (accessed on 29 August 2019).
- Gupta, V.B.; Rajagopala, M.; Ravishankar, B. Etiopathogenesis of cataract: An appraisal. Indian J. Ophthalmol. 2014, 62, 103–110. [Google Scholar] [CrossRef]
- Liu, Y.C.; Wilkins, M.; Kim, T.; Malyugin, B.; Mehta, J.S. Cataracts. Lancet 2017, 390, 600–612. [Google Scholar] [CrossRef]
- Bollinger, K.E.; Langston, R.H. What can patients expect from cataract surgery? Clevel. Clin. J. Med. 2008, 75, 193–200. [Google Scholar] [CrossRef]
- National Institute of Health, National Eye Institute. Facts about Cataracts. Available online: https://nei.nih.gov/health/cataract/cataract_facts (accessed on 20 August 2019).
- Augusteyn, R.C. On the growth and internal structure of the human lens. Exp. Eye Res. 2010, 90, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Michael, R.; Van Marle, J.; Vrensen, G.F.; Van den Berg, T.J. Changes in the refractive index of lens fibre membranes during maturation--impact on lens transparency. Exp. Eye Res. 2003, 77, 93–99. [Google Scholar] [CrossRef]
- Bassnett, S. Lens organelle degradation. Exp. Eye Res. 2002, 74, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Moreau, K.L.; King, J.A. Protein Misfolding and Aggregation in Cataract Disease and Prospects for Prevention. Trends Mol. Med. 2012, 18, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.; Chiesa, R.; Sredy, J.; Garner, W. cAMP-dependent phosphorylation of bovine lens alpha-crystallin. Proc. Natl. Acad. Sci. USA 1985, 8, 4712–4716. [Google Scholar] [CrossRef]
- Aquilina, J.A.; Benesch, J.L.P.; Ding, L.L.; Yaron, O.; Horwitz, J.; Robinson, C.V. Phosphorylation of αB-Crystallin Alters Chaperone Function through Loss of Dimeric Substructure. J. Boil. Chem. 2004, 279, 28675–28680. [Google Scholar] [CrossRef]
- Horwitz, J. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA 1992, 89, 10449–10453. [Google Scholar] [CrossRef]
- Lampi, K.J.; Ma, Z.; Shih, M.; Shearer, T.R.; Smith, J.B.; Smith, D.L.; David, L.L. Sequence analysis of betaA3, betaB3, and betaA4 crystallins completes the identification of the major proteins in young human lens. J. Boil. Chem. 1997, 272, 2268–2275. [Google Scholar] [CrossRef]
- Jaenicke, R.; Slingsby, C. Lens crystallins and their microbial homologs: Structure, stability, and function. Crit. Rev. Biochem. Mol. Biol. 2001, 36, 435–499. [Google Scholar] [CrossRef]
- Weale, R.A. Human ocular aging and ambient temperature. Br. J. Ophthalmol. 1981, 65, 869–870. [Google Scholar] [CrossRef]
- Mathias, R.T.; White, T.W.; Gong, X. Lens gap junctions in growth, differentiation, and homeostasis. Physiol. Rev. 2010, 90, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Barbazetto, I.A.; Liang, J.; Chang, S.; Zheng, L.; Spector, A.; Dillon, J.P. Oxygen tension in the rabbit lens and vitreous before and after vitrectomy. Exp. Eye Res. 2004, 78, 917–924. [Google Scholar] [CrossRef] [PubMed]
- McNulty, R.; Wang, H.; Mathias, R.; Ortwerth, B.J.; Truscott, R.J.W.; Bassnett, S. Regulation of tissue oxygen levels in the mammalian lens. J. Physiol. 2004, 559, 883–898. [Google Scholar] [CrossRef] [PubMed]
- Evanger, K.; Haugen, O.H.; Irgens, Å.; Aanderud, L.; Thorsen, E. Ocular refractive changes in patients receiving hyperbaric oxygen administered by oronasal mask or hood. Acta Ophthalmol. Scand. 2004, 82, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Simpanya, M.F.; Ansari, R.R.; Suh, K.I.; Leverenz, V.R.; Giblin, F.J. Aggregation of Lens Crystallins in an In Vivo Hyperbaric Oxygen Guinea Pig Model of Nuclear Cataract: Dynamic Light-Scattering and HPLC Analysis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4641–4651. [Google Scholar] [CrossRef]
- Bassnett, S.; Shi, Y.; Vrensen, G.F. Biological glass: Structural determinants of eye lens transparency. Philos. Trans. R. Soc. B 2011, 366, 1250–1264. [Google Scholar] [CrossRef]
- Benedek, G.B. Theory of transparency of the eye. Appl. Opt. 1971, 10, 459–473. [Google Scholar] [CrossRef]
- Delaye, M.; Tardieu, A. Short-range order of crystallin proteins accounts for eye lens transparency. Nature 1983, 302, 415–417. [Google Scholar] [CrossRef]
- Michael, R.; Bron, A.J. The ageing lens and cataract: A model of normal and pathological ageing. Philos. Trans. R. Soc. B 2011, 366, 1278–1292. [Google Scholar] [CrossRef]
- Horwitz, J. Alpha-crystallin. Exp. Eye Res. 2003, 76, 145–153. [Google Scholar] [CrossRef]
- Costello, M.J.; Johnsen, S.; Metlapally, S.; Gilliland, K.O.; Ramamurthy, B.; Krishna, P.V.; Balasubramanian, R. Ultrastructural analysis of damage to nuclear fiber cell membranes in advanced age-related cataracts from India. Exp. Eye Res. 2008, 87, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Ringvold, A. Corneal epithelium and UV-protection of the eye. Acta Ophthalmol. Scand. 1998, 76, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.M.; Truscott, R.J.W. Ultraviolet filter compounds in human lenses: 3-hydroxykynurenine glucoside formation. Vis. Res. 1994, 34, 1369–1374. [Google Scholar] [CrossRef]
- Hains, P.G.; Simpanya, M.F.; Giblin, F.; Truscott, R.J.W. UV filters in the lens of the thirteen lined ground squirrel (Spermophilus tridecemlineatus). Exp. Eye Res. 2006, 82, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Tsentalovich, Y.P.; Sherin, P.S.; Kopylova, L.V.; Cherepanov, I.V.; Grilj, J.; Vauthey, E. Photochemical properties of UV filter molecules of the human eye. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7687–7696. [Google Scholar] [CrossRef]
- Roberts, J. Photobiology of the Human Lens. 2011. Available online: http://photobiology.info/Roberts.html (accessed on 11 January 2020).
- Spector, A. Oxidative stress-induced cataract: Mechanism of action. FASEB J. 1995, 9, 1173–1182. [Google Scholar] [CrossRef]
- Andley, U.P.; Liang, J.J.N.; Lou, M.F. Biochemical mechanisms of age-related cataract. In Principles and Practice of Ophthalmology, 2nd ed.; Albert, D.M., Jakobiec, F.A., Eds.; Saunders: Philadelphia, PA, USA, 2000; pp. 1428–1449. [Google Scholar]
- Jahngen-Hodge, J.; Cyr, D.; Laxman, E.; Taylor, A. Ubiquitin and ubiquitin conjugates in human lens. Exp. Eye Res. 1992, 55, 897–902. [Google Scholar] [CrossRef]
- Lou, M.F. Redox regulation in the lens. Prog. Retin. Eye Res. 2003, 22, 657–682. [Google Scholar] [CrossRef]
- Rahman, I.; MacNee, W. Lung glutathione and oxidative stress: Implications in cigarette smoke-induced airway disease. Am. J. Physio. Lung Cell. Mol. Physiol. 1999, 277, 1067–1088. [Google Scholar] [CrossRef]
- Reddy, V.N.; Giblin, F.J. Metabolism and function of glutathione in the lens. Ciba Found Symp. 2000, 106, 65–87. [Google Scholar]
- Giblin, F.J. Glutathione: A vital lens antioxidant. J. Ocul. Pharmacol. Ther. 2000, 16, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.H.; Truscott, R.J. An impediment to glutathione diffusion in older normal human lenses: A possible precondition for nuclear cataract. Exp. Eye Res. 1998, 67, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Epstein, D.L.; Kinoshita, J.H. The effect of diamide on lens glutathione and lens membrane function. Investig. Ophthalmol. Vis. Sci. 1970, 9, 629–638. [Google Scholar]
- Jacques, P.F.; Taylor, A.; E Hankinson, S.; Willett, W.C.; Mahnken, B.; Lee, Y.; Vaid, K.; Lahav, M. Long-term vitamin C supplement use and prevalence of early age-related lens opacities. Am. J. Clin. Nutr. 1997, 66, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Yeum, K.J.; Taylor, A.; Tang, G.; Russell, R.M. Measurement of carotenoids, retinoids, and tocopherols in human lenses. Investig. Ophthalmol. Vis. Sci. 1995, 36, 2756–2761. [Google Scholar]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Behndig, A.; Karlsson, K.; Reaume, A.G.; Sentman, M.L.; Marklund, S.L. In vitro photochemical cataract in mice lacking copper-zinc superoxide dismutase. Free Radic. Biol. Med. 2001, 31, 738–744. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Ageing, nutrition, disease and therapy: A role for antioxidants? In Free Radical Biology and Medicine; Halliwell, B., Gutteridge, J.M.C., Eds.; Oxford University Press: Oxford, UK, 2015. [Google Scholar] [CrossRef]
- Giblin, F.J.; McCready, J.P.; Reddy, V.N. The role of glutathione metabolism in the detoxification of H2O2 in rabbit lens. Investig. Ophthalmol. Vis. Sci. 1982, 22, 330–335. [Google Scholar]
- Slaughter, M.R.; Thakkar, H.; O’Brien, P.J. Differential expression of the lenticular antioxidant system in laboratory animals: A determinant of species predilection to oxidative stress-induced ocular toxicity? Curr. Eye Res. 2003, 26, 15–23. [Google Scholar] [CrossRef]
- Chaudière, J.; Ferrari-Iliou, R. Intracellular antioxidants: From chemical to biochemical mechanisms. Food Chem. Toxicol. 1999, l37, 949–962. [Google Scholar] [CrossRef]
- Rathbun, W.B.; Bovis, M.G.; Holleschau, A.M. Species survey of glutathione peroxidase and glutathione reductase: Search for an animal model of the human lens. Ophthalmic Res. 1986, 18, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin and Glutaredoxin systems. J. Biol. Chem. 1989, 264, 13963–13966. [Google Scholar] [PubMed]
- Mieyal, J.J.; Srinivasan, U.; Starke, D.W.; Gravina, S.A.; Mieyal, P.A. Glutathionyl specificity of thioltransferases: Mechanistic and physiological implications. In Biothiols in Health and Disease; CRC Press: Boca Raton, FL, USA, 1995; pp. 305–372. [Google Scholar]
- Raghavachari, N.; Lou, M.F. Evidence for the presence of thioltransferase in the lens. Exp. Eye Res. 1996, 63, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, K.C.; Reddy, P.G.; Bhuyan, D.K. Thioredoxin genes in lens: Regulation by oxidative stress. Methods Enzymol. 2002, 347, 421–435. [Google Scholar] [PubMed]
- Yegorova, S.; Liu, A.; Lou, M.F. Human lens thioredoxin: Cloning, overexpression, characterization and H2O2-upregulation. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2365. [Google Scholar]
- Hains, P.G.; Truscott, R.J. Post-translational modifications in the nuclear region of young, aged, and cataract human lenses. J. Proteome Res. 2007, 6, 3935–3943. [Google Scholar] [CrossRef]
- Hains, P.G.; Truscott, R.J. Proteomic analysis of the oxidation of cysteine residues in human age-related nuclear cataract lenses. Biochim. Biophys. Acta 2008, 1784, 1959–1964. [Google Scholar] [CrossRef]
- Lampi, K.J.; Ma, Z.; Hanson, S.R.; Azuma, M.; Shih, M.; Shearer, T.R.; Smith, D.L.; Smith, J.B.; David, L.L. Age-related Changes in Human Lens Crystallins Identified by Two-dimensional Electrophoresis and Mass Spectrometry. Exp. Eye Res. 1998, 67, 31–43. [Google Scholar] [CrossRef]
- Ma, Z.; Hanson, S.R.; Lampi, K.J.; David, L.L.; Smith, D.L.; Smith, J.B. Age-Related Changes in Human Lens Crystallins Identified by HPLC and Mass Spectrometry. Exp. Eye Res. 1998, 67, 21–30. [Google Scholar] [CrossRef]
- Zhang, Z.; Smith, D.L.; Smith, J.B. Human beta-crystallins modified by backbone cleavage, deamidation and oxidation are prone to associate. Exp. Eye Res. 2003, 77, 259–272. [Google Scholar] [CrossRef]
- Hanson, S.R.; Hasan, A.; Smith, D.L.; Smith, J.B. The major in vivo modifications of the human water-insoluble lens crystallins are disulfide bonds, deamidation, methionine oxidation and backbone cleavage. Exp. Eye Res. 2000, 71, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Flaugh, S.L.; Mills, I.A.; King, J. Glutamine deamidation destabilizes human γD-crystallin and lowers the kinetic barrier to unfolding. J. Biol. Chem. 2006, 281, 30782–30793. [Google Scholar] [CrossRef] [PubMed]
- Truscott, R.J. Age-related nuclear cataract-oxidation is the key. Exp. Eye Res. 2005, 80, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Santhoshkumar, P.; Udupa, P.; Murugesan, R.; Sharma, K.K. Significance of Interactions of Low Molecular Weight Crystallin Fragments in Lens Aging and Cataract Formation. J. Biol. Chem. 2008, 283, 8477–8485. [Google Scholar] [CrossRef]
- Santhoshkumar, P.; Raju, M.; Sharma, K.K. αA-Crystallin Peptide 66SDRDKFVIFLDVKHF80 Accumulating in Aging Lens Impairs the Function of α-Crystallin and Induces Lens Protein Aggregation. PLoS ONE 2011, 6, e19291. [Google Scholar] [CrossRef]
- Udupa, E.G.; Sharma, K.K. Effect of oxidized betaB3-crystallin peptide on lens betaL-crystallin: Interaction with betaB2-crystallin. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2514–2521. [Google Scholar] [CrossRef]
- Udupa, P.E.; Sharma, K.K. Effect of oxidized betaB3-crystallin peptide (152–166) on thermal aggregation of bovine lens gamma-crystallins: Identification of peptide interacting sites. Exp. Eye Res. 2005, 80, 185–196. [Google Scholar] [CrossRef]
- Benedek, G.B. Cataract as a protein condensation disease: The Proctor Lecture. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1911–1921. [Google Scholar]
- Evans, P.; Wyatt, K.; Wistow, G.; Bateman, O.; Wallace, B.; Slingsby, C. The P23T Cataract Mutation Causes Loss of Solubility of Folded γD-Crystallin. J. Mol. Boil. 2004, 343, 435–444. [Google Scholar] [CrossRef]
- Kmoch, S.; Brynda, J.; Asfaw, B.; Bezouška, K.; Novák, P.; Řezáčová, P.; Ondrová, L.; Filipec, M.; Sedláček, J.; Elleder, M. Link between a novel human γD-crystallin allele and a unique cataract phenotype explained by protein crystallography. Hum. Mol. Genet. 2000, 9, 1779–1786. [Google Scholar] [CrossRef]
- Pande, A.; Pande, J.; Asherie, N.; Lomakin, A.; Ogun, O.; King, J.A.; Lubsen, N.H.; Walton, D.; Benedek, G.B. Molecular basis of a progressive juvenile-onset hereditary cataract. Proc. Natl. Acad. Sci. USA 2000, 97, 1993–1998. [Google Scholar] [CrossRef] [PubMed]
- Pande, A.; Pande, J.; Asherie, N.; Lomakin, A.; Ogun, O.; King, J.; Benedek, G.B. Crystal cataracts: Human genetic cataract caused by protein crystallization. Proc. Natl. Acad. Sci. USA 2001, 98, 6116–6120. [Google Scholar] [CrossRef] [PubMed]
- American Optometric Association. Cataract Surgery. 2017. Available online: https://www.aoa.org/patients-and-public/eye-and-vision-problems/glossary-of-eye-and-vision-conditions/cataract/cataract-surgery (accessed on 11 January 2020).
- Powe, N.R.; Schein, O.D.; Gieser, S.C.; Tielsch, J.M.; Luthra, R.; Javitt, J.; Steinberg, E.P. Synthesis of the Literature on Visual Acuity and Complications Following Cataract Extraction With Intraocular Lens Implantation. Arch. Ophthalmol. 1994, 112, 239. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.; Mahroo, O.A.R.; Spalton, D.J. Complications of cataract surgery. Clin. Exp. Optom. 2010, 93, 379–389. [Google Scholar] [CrossRef]
- Mucke, H.A.; Mucke, P.; Mucke, E. Pharmacological therapies for cataract and refractive errors: Landscaping niches of ocular drug patenting. Pharm. Pat. Anal. 2012, 1, 165–175. [Google Scholar] [CrossRef]
- Mohanty, I.; Joshi, S.; Trivedi, D.; Srivastava, S.; Gupta, S.K. Lycopene prevents sugar-induced morphological changes and modulates antioxidant status of human lens epithelial cells. Br. J. Nutr. 2002, 88, 347–354. [Google Scholar] [CrossRef]
- Reddy, V.N. Glutathione and its function in the lens—An overview. Exp. Eye Res. 1990, 50, 771–778. [Google Scholar] [CrossRef]
- Varma, S.D.; Hegde, K.R. Effect of α-ketoglutarate against selenite cataract formation. Exp. Eye Res. 2004, 79, 913–918. [Google Scholar] [CrossRef]
- Padalkar, P.; Bulakh, P.M.; Melinkeri, R.R. Effect of Endogenous Antioxidants on Hydrogen Peroxide Induced Experimental Cataract. Int. J. Health Sci. Res. 2013, 3, 11–15. [Google Scholar]
- Anbaraki, A.; Khoshaman, K.; Ghasemi, Y.; Yousefi, R. Preventive role of lens antioxidant defense mechanism against riboflavin-mediated sunlight damaging of lens crystallins. Int. J. Biol. Macromol. 2016, 91, 895–904. [Google Scholar] [CrossRef]
- Gao, S.; Qin, T.; Liu, Z.; Caceres, M.A.; Ronchi, C.F.; Chen, C.-Y.O.; Yeum, K.-J.; Taylor, A.; Blumberg, J.B.; Liu, Y.; et al. Lutein and zeaxanthin supplementation reduces H2O2-induced oxidative damage in human lens epithelial cells. Mol. Vis. 2011, 17, 3180–3190. [Google Scholar] [PubMed]
- Shang, F.; Lu, M.; Dudek, E.; Reddan, J.; Taylor, A. Vitamin C and vitamin E restore the resistance of GSH-depleted lens cells to H2O2. Free Radic. Biol. Med. 2003, 34, 521–530. [Google Scholar] [CrossRef]
- Shetty, L.L.; Harikiran, H.; Sharma, A. In vitro prophylactic cataract prevention study on glucose induced cataract by quercetin and alpha tocopherol. Int. J. Pharm. Sci. Res. 2010, 1, 41–45. [Google Scholar]
- Kouvaris, J.; Kouloulias, V.E.; Vlahos, L.J. Amifostine: The first selective-target and broad-spectrum radioprotector. Oncology 2007, 12, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, Y.; Rat, P.; Piel, G.; Christen, M.-O.; Touboul, E.; Warnet, J.-M. Lens epithelial cell protection by aminothiol WR-1065 and anetholedithiolethione from ionizing radiation. Int. J. Cancer 2001, 96, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Witt, E.H.; Tritschle, H.J. Alpha-lipoic acid as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef]
- Packer, L.; Kraemer, K.; Rimbach, G. Molecular aspects of lipoic acid in the prevention of diabetes complications. Nutrition 2001, 17, 888–895. [Google Scholar] [CrossRef]
- Yan, W.; Li, N.; Hu, X.; Huang, Y.; Zhang, W.; Wang, Q.; Wang, F.; Wang, C.; Zhai, X.; Xu, R.; et al. Effect of oral ALA supplementation on oxidative stress and insulin sensitivity among overweight/obese adults: A double-blinded, randomized, controlled, cross-over intervention trial. Int. J. Cardiol. 2013, 167, 602–603. [Google Scholar] [CrossRef]
- Cremer, D.R.; Rabeler, R.; Roberts, A.; Lynch, B. Safety evaluation of α-lipoic acid (ALA). Regul. Toxicol. Pharmacol. 2006, 46, 29–41. [Google Scholar] [CrossRef]
- Cremer, D.R.; Rabeler, R.; Roberts, A.; Lynch, B. Long-term safety of α-lipoic acid (ALA) consumption: A 2-year study. Regul. Toxicol. Pharmacol. 2006, 46, 193–201. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.Z.; Shi, J.M.; Jia, S.B. Alpha lipoic acid protects lens from H2O2-induced cataract by inhibiting apoptosis of lens epithelial cells and inducing activation of anti-oxidative enzymes. Asian Pac. J. Trop. Med. 2013, 6, 548–551. [Google Scholar] [CrossRef]
- Ramos-Ibeas, P.; Barandalla, M.; Colleoni, S.; Lazzari, G. Pyruvate antioxidant roles in human fibroblasts and embryonic stem cells. Mol. Cell. Biochem. 2017, 429, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Nagatome, M.; Kondo, Y.; Kadowaki, D.; Saishyo, Y.; Irikura, M.; Irie, T.; Ishitsuka, Y. Ethyl pyruvate attenuates acetaminophen-induced liver injury and prevents cellular injury induced by N-acetyl-p-benzoquinone imine. Heliyon 2018, 4, e00521. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Halliwell, B. Artefacts in cell culture: α-Ketoglutarate can scavenge hydrogen peroxide generated by ascorbate and epigallocatechin gallate in cell culture media. Biochem. Biophys. Res. Commun. 2011, 406, 20–24. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Principles of Biochemistry, 4th ed.; W. H. Freeman: New York, NY, USA, 2005; ISBN 0-7167-4339-6. [Google Scholar]
- Jain, R.M.; Bulakh, P.M. Effect of ketoacids on H2O2 induced cataract. Indian J. Clin. Biochem. 2003, 8, 91–95. [Google Scholar] [CrossRef]
- Varma, S.D.; Ramachandran, S.; Devamanoharan, P.S.; Morris, S.M.; Ali, A.H. Prevention of oxidative damage to rat lens by pyruvate in vitro: Possible attenuation in vivo. Curr. Eye Res. 1995, 14, 643–649. [Google Scholar] [CrossRef]
- Bunce, G.E.; Hess, J.L.; Davis, D. Cataract formation following limited amino acid intake during gestation and lactation. Proc. Soc. Exp. Biol. Med. 1984, 176, 485–489. [Google Scholar] [CrossRef]
- Rathore, M.S.; Gupta, V.B. Protective effect of amino acids on eye lenses against oxidative stress induced by hydrogen peroxide. Asian J. Pharm. Clin. Res. 2010, 3, 166–169. [Google Scholar]
- Kelly, G.S. Clinical applications of N-acetylcysteine. Altern. Med. Rev. 1998, 3, 114–127. [Google Scholar]
- Shattuck, K.E.; Rassin, D.K.; Grinnell, C.D. N-acetylcysteine protects from glutathione depletion in rats exposed to hyperoxia. JPEN J. Parenter. Enter. Nutr. 1998, 22, 228–233. [Google Scholar] [CrossRef]
- Jain, A.K.; Lim, G.; Langford, M.; Jain, S.K. Effect of high-glucose levels on protein oxidation in cultured lens cells, and in crystalline and albumin solution and its inhibition by vitamin B6 and N-acetylcysteine: Its possible relevance to cataract formation in diabetes. Free Radic. Biol. Med. 2002, 33, 1615–1621. [Google Scholar] [CrossRef]
- Sunitha, K.; Hemshekhar, M.; Thushara, R.M.; Santhosh, M.S.; Yariswamy, M.; Kemparaju, K.; Girish, K.S. N-Acetylcysteine amide: A derivative to fulfill the promises of N-Acetylcysteine. Free. Radic. Res. 2013, 47, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, X.C.; Yan, H.; Li, M.Y. Hyperoxia-induced lens damage in rabbit: Protective effects of N-acetylcysteine. Mol. Vis. 2009, 15, 2945–2952. [Google Scholar] [PubMed]
- Tobwala, S.; Pinarci, E.Y.; Maddirala, Y.; Ercal, N. N-acetylcysteine amide protects against dexamethasone-induced cataract related changes in cultured rat lenses. Adv. Biol. Chem. 2014, 4, 26–34. [Google Scholar] [CrossRef][Green Version]
- Liu, J.; Head, E.; Kuratsune, H.; Cotman, C.W.; Ames, B.N. Comparison of the effects of l-carnitine and acetyl-l-carnitine on carnitine levels, ambulatory activity, and oxidative stress biomarkers in the brain of old rats. Ann. N. Y. Acad. Sci. 2004, 1033, 117–131. [Google Scholar] [CrossRef]
- Calabrese, V.; Ravagna, A.; Colombrita, C.; Scapagnini, G.; Guagliano, E.; Calvani, M.; Butterfield, D.A.; Stella, A.M.G. Acetylcarnitine induces heme oxygenase in rat astrocytes and protects against oxidative stress: Involvement of the transcription factor Nrf2. J. Neurosci. Res. 2005, 79, 509–521. [Google Scholar] [CrossRef]
- Geraldine, P.; Sneha, B.B.; Elanchezhian, R.; Ramesh, E.; Kalavathy, C.; Kaliamurthy, J.; Thomas, P. Prevention of selenite-induced cataractogenesis by acetyl-l-carnitine: An experimental study. Exp. Eye Res. 2006, 83, 1340–1349. [Google Scholar] [CrossRef]
- Shibata, T.; Shibata, S.; Shibata, N.; Kiyokawa, E.; Sasaki, H.; Singh, D.P.; Kubo, E. Propolis, a Constituent of Honey, Inhibits the Development of Sugar Cataracts and High-Glucose-Induced Reactive Oxygen Species in Rat Lenses. J. Ophthalmol. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Williams, R.J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef]
- Russo, G.L.; Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Iannitti, R.; Palumbo, R. Quercetin: A pleiotropic kinase inhibitor against cancer. Cancer Treat. Res. 2014, 159, 185–205. [Google Scholar]
- Sundararajan, M.; Thomas, P.A.; Teresa, P.A.; Anbukkarasi, M.; Geraldine, P. Regulatory effect of chrysin on expression of lenticular calcium transporters, calpains, and apoptotic-cascade components in selenite-induced cataract. Mol. Vis. 2016, 22, 401–423. [Google Scholar] [PubMed]
- Chaudhury, S.; Roy, P.; Dasgupta, S. Green tea flavanols protect human γB-crystallin from oxidative photodamage. Biochemie 2017, 137, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Gour, S.; Peter, O.S.; Gandhi, S.; Goyal, P.; Pandey, J.; Harsolia, R.S.; Yadav, J.K. Effect of Green Tea Polyphenol Epigallocatechin-3-gallate on the Aggregation of αA(66-80) Peptide, a Major Fragment of αA-crystallin Involved in Cataract Development. Curr. Eye Res. 2017, 42, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Halder, N.; Srivastava, S.; Trivedi, D.; Joshi, S.; Varma, S. Green Tea (Camellia sinensis) Protects against Selenite-Induced Oxidative Stress in Experimental Cataractogenesis. Ophthalmic Res. 2002, 34, 258–263. [Google Scholar] [CrossRef]
- Kirtikar, R.K.; Basu, D.B. Indian Medicinal Plants, 2nd ed.; International Book Distributors: Uttarakhand, India, 1935; Volume 3, pp. 1635–1636. [Google Scholar]
- Kirtikar, R.K.; Basu, D.B.; Chopra, R.N.; Nayar, S.L.; Chopra, I.C. Glossary of Indian Medicinal Plants; CSIR: New Delhi, India, 1935; p. 258. [Google Scholar]
- Anjaria, J.; Parabia, M.; Bhatt, G.; Khamar, R. Nature Heals: A Glossary of Selected Medicinal Plants of India, 2nd ed.; SRISTI Innovations: Ahmedabad, India, 2002; p. 26. [Google Scholar]
- Biju, P.G.; Devi, V.G.; Lija, Y.; Abraham, A. Protection against selenite cataract in rat lens by drevogenin D, a triterpenoid aglycone from Dregea volubilis. J. Med. Food 2007, 10, 308–315. [Google Scholar] [CrossRef]
- Wani, S.A.; Kumar, P. Fenugreek: A review on its nutraceutical properties and utilization in various food products. J. Saudi Soc. Agric. Sci. 2018, 17, 97–106. [Google Scholar] [CrossRef]
- Naidu, M.M.; Shyamala, B.N.; Naik, P.J.; Sulochanamma, G.; Srinivas, P. Chemical composition and antioxidant activity of the husk and endosperm of fenugreek seeds. Food Sci. Technol. 2010, 44, 451–456. [Google Scholar]
- Pathak, N.; Pant, N.; Singh, J.P.; Agrawal, S. Antioxidant activity of Trigonella foenum graecum L. using various in vitro models. Int. J. Herb. Med. 2014, 2, 53–57. [Google Scholar]
- Gupta, S.K.; Kalaiselvan, V.; Srivastava, S.; Saxena, R.; Agrawal, S.S. Trigonella foenum-graecum (Fenugreek) protects against selenite-induced oxidative stress in experimental cataractogenesis. Biol. Trace Elem. Res. 2010, 136, 258–268. [Google Scholar] [CrossRef]
- Gupta, S.K.; Kalaiselvan, V.; Srivastava, S.; Agrawal, S.S.; Saxena, R. Evaluation of anti-cataract potential of Triphala in selenite-induced cataract: In vitro and in vivo studies. J. Ayurveda Integr. Med. 2010, 1, 280–286. [Google Scholar] [CrossRef]
- Peterson, C.T.; Denniston, K.; Chopra, D. Therapeutic Uses of Triphala in Ayurvedic Medicine. J. Altern. Complement. Med. 2017, 23, 607–614. [Google Scholar] [CrossRef]
- Anwar, F.; Latif, S.; Ashraf, M.; Gilani, A.H. Moringa oleifera: A food plant with multiple medicinal uses. Phytother. Res. 2007, 21, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Thurber, M.D.; Fahey, J.W. Adoption of Moringa oleifera to combat under-nutrition viewed through the lens of the “Diffusion of Innovations” theory. Ecol. Food Nutr. 2009, 48, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Kurmi, R.; Ganeshpurkar, A.; Bansal, D.; Agnihotri, A.; Dubey, N. Ethanol extract of Moringa oliefera prevents in vitro glucose induced cataract on isolated goat eye lens. Indian J. Ophthalmol. 2014, 62, 154–157. [Google Scholar]
- Yoganarasimhan, S.N. Medicinal Plants of India; Cyber Media: HongKong, China, 2000; Volume 2, p. 10. [Google Scholar]
- Ahmed, M.; Amin, S.; Islam, M.; Takahashi, M.; Okuyama, E.; Hossain, C.F. Analgesic principle from Abutilon indicum. Die Pharm. 2000, 55, 314–316. [Google Scholar]
- Rajakaruna, N.; Harris, C.S.; Towers, G.H.N. Antimicrobial activity of plants collected from serpentine outcrops in Sri Lanka. Pharm. Biol. 2002, l40, 235–244. [Google Scholar] [CrossRef]
- Rosheni, N.; Nithya, K.; Brindha, S.; Elango, N.; Gokila, V.; Nishmitha, S.; Kokila, S. Evaluation of anti-cataract potential of Abutilon indicum: An in vitro study. Indian J. Sci. Res. 2016, 7, 81–84. [Google Scholar]
- Kumar, M.; Singh, T.; Ali, J.; Tyagi, L.K. In vitro anti-cataract activity of Zingiber officinale on goat lenses. Int. J. Pharm. Biol. Arch. 2011, 2, 1430–1433. [Google Scholar]
- Zeb, A. Chemical and nutritional constituents of Seabuckthorn juice. Pak. J. Nutr. 2004, 3, 99–106. [Google Scholar]
- Narayanan, S.; Ruma, D.; Gitika, B.; Sharma, S.K.; Pauline, T.; Ram, M.S.; Ilavazhagan, G.; Sawhney, R.C.; Kumar, D.; Banerjee, P.K. Antioxidant activities of seabuckthorn (Hippophae rhamnoides) during hypoxia induced oxidative stress in glial cells. Mol. Cell. Biochem. 2005, 278, 9–14. [Google Scholar] [CrossRef]
- Dubey, S.; Deep, P.; Singh, A.K. Phytochemical characterization and evaluation of anti-cataract potential of seabuckthorn leaf extract. Vet. Ophthalmol. 2016, 19, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Bhardwaj, R.; Gupta, R.K. In vitro antioxidant activity of extracts from the leaves of Abies pindrow Royle. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Majeed, H.; Bokhari, T.Z.; Sherwani, S.K.; Younis, U.; Shah, M.H.R.; Khaliq, B. An Overview of Biological, Phytochemical, and Pharmacological Values of Abies pindrow. J. Pharm. Phytochem. 2013, 2, 182–187. [Google Scholar]
- Dubey, S.; Saha, S.; Saraf, S.A. In vitro anti-cataract evaluation of standardised Abies pindrow leaf extract using isolated goat lenses. Nat. Prod. Res. 2015, 29, 1145–1148. [Google Scholar] [CrossRef]
- Azeez, M.A.; Bello, O.S.; Adedeji, A.O. Traditional and medicinal uses of Luffa cylindrica: A review. J. Med. Plants 2013, 1, 102–111. [Google Scholar]
- Du, Q.; Xu, Y.; Li, L.; Zhao, Y.; Jerz, G.; Winterhalter, P. Antioxidant Constituents in the Fruits of Luffa cylindrica (L.) Roem. J. Agric. Food Chem. 2006, 54, 4186–4190. [Google Scholar] [CrossRef]
- Hazra, M.; KunduSen, S.; Bhattacharya, S.; Haldar, P.K.; Gupta, M.; Mazumder, U.K. Evaluation of hypoglycemic and antihyperglycemic effects of Luffa cylindrica fruit extract in rats. J. Adv. Pharm. Educ. Res. 2011, 2, 138–146. [Google Scholar]
- Dubey, S.; Saha, S.; Kaithwas, G.; Saraf, S.A. Effect of standardized fruit extract of Luffa cylindrica on oxidative stress markers in hydrogen peroxide induced cataract. Indian J. Pharmacol. 2015, 47, 644–648. [Google Scholar]
- Cohen, M.M. Tulsi—Ocimum sanctum: A herb for all reasons. J. Ayurveda Integr. Med. 2014, 5, 251–259. [Google Scholar] [CrossRef]
- Gupta, S.K.; Srivastava, S.; Trivedi, D.; Joshi, S.; Halder, N. Ocimum sanctum modulates selenite-induced cataractogenic changes and prevents rat lens opacification. Curr. Eye Res. 2005, 30, 583–591. [Google Scholar] [CrossRef]
- Lim, J.C.; Umapathy, A.; Donaldson, P.J. Tools to fight the cataract epidemic: A review of experimental animal models that mimic age related nuclear cataract. Exp. Eye Res. 2016, 145, 432–443. [Google Scholar] [CrossRef] [PubMed]
- West-Mays, J.; Bowman, S. Animal Models of Cataracts. In Animal Models of Ophthalmic Diseases. Essentials in Ophthalmology; Springer: Cham, Switzerland, 2016; pp. 11–29. [Google Scholar]
- Kyselova, Z. Different experimental approaches in modelling cataractogenesis. Interdis. Toxicol. 2010, 3, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Ringvold, A. Quenching of UV-induced fluorescence by ascorbic acid in the aqueous humor. Acta Ophthalmol. Scand. 1995, 73, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.N.; Giblin, F.J.; Lin, L.R.; Chak-rapani, B. The effect of aqueous hu-morscorbate on ultraviolet-B-induced DNA damage in lens epi-thelium. Investig. Ophthalmol. Vis. Sci. 1998, 39, 344–350. [Google Scholar]
- Ohta, Y.; Niwa, T.; Yamasaki, T. Effect of prolonged marginal ascorbic acid deficiency on lenticular levels of antioxidants and lipid peroxide in guinea pigs. Int. J. Vitam. Nutr. Res. 2001, 71, 103–109. [Google Scholar] [CrossRef]
- Mody, V.C.; Kakar, M.; Elfving, Å.; Söderberg, P.G.; Löfgren, S. Ascorbate in the rat lens: Dependence on dietary intake. Ophthalmic Res. 2005, 37, 142–149. [Google Scholar] [CrossRef]
- Harding, J.J. Can drugs or micronutrients prevent cataract? Drugs Aging 2001, 18, 473–486. [Google Scholar] [CrossRef]
- Peighmbarzadeh, S.Z.; Tavana, M. Attenuation of experimental cataract by vitamin c in rabbits. WALIA J. 2014, 30, 204–207. [Google Scholar]
- Hosseini, H.J.; Aminlari, M.; Khalili, M.R. Prevention of selenite-induced cataract by l-cysteine and vitamin C in rats. Iran. Red Cres. Med. 2008, 10, 281–287. [Google Scholar]
- Yokoyama, T.; Sasaki, H.; Giblin, F.J.; Reddy, V.N. A physiological level of ascorbate inhibits galactose cataract in guinea pigs by decreasing polyol accumulation in the lens epithelium: A dehydroascorbate-linked mechanism. Exp. Eye Res. 1994, 58, 207–218. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Nagai, N.; Ishimori, N.; Oguchi, J.; Tamura, H. Administration of antioxidant compounds affects the lens chaperone activity and prevents the onset of cataracts. Biomed. Pharmacother. 2017, 95, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Giblin, F.J.; Winkler, B.S.; Chakrapani, B.; Leverenz, V.; Shu, C.C. A protective role for glutathione-dependent reduction of dehydroascorbic acid in lens epithelium. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1807–1817. [Google Scholar]
- Kojima, M.; Shui, Y.B.; Murano, H.; Nagata, M.; Hockwin, O.; Sasaki, K.; Takahashi, N. Low vitamin E level as a subliminal risk factor in a rat model of prednisolone-induced cataract. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1116–1120. [Google Scholar]
- Längle, U.W.; Wolf, A.; Cordier, A. Enhancement of SDZ ICT 322-induced cataracts and skin changes in rats following vitamin E-and selenium-deficient diet. Arch. Toxicol. 1997, 71, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Ayala, M.N.; Söderberg, P.G. Vitamin E can protect against ultraviolet radiation-induced cataract in albino rats. Ophthalm. Res. 2004, 36, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.B.; Nayak, S.; Reddy, P.Y.; Bhat, K.S. Reduced levels of rat lens antioxidant vitamins upon in vitro UVB irradiation. J. Nutr. Biochem. 2001, 12, 121–124. [Google Scholar] [CrossRef]
- Kyselova, Z.; Gajdosik, A.; Gajdosikova, A.; Ulicna, O.; Mihalova, D.; Karasu, C.; Stefek, M. Effect of the pyridoindole antioxidant stobadine on development of experimental diabetic cataract and on lens protein oxidation in rats: Comparison with vitamin E and BHT. Mol. Vis. 2005, 11, 56–65. [Google Scholar]
- Khan, S.B.; Choudhary, R.; Vishwakarma, P.K.; Singh, A.; Shree, J.; Bodakhe, S.H. Protective effect of alpha-lipoic acid on progression of cataract formation in fructose-induced experimental cataract. Pharma Nutr. 2017, 5, 127–132. [Google Scholar] [CrossRef]
- Maitra, I.; Serbinova, E.; Trischler, H.; Packer, L. α-Lipoic acid prevents buthionine sulfoximine-induced cataract formation in newborn rats. Free Radic. Biol. Med. 1995, 18, 823–829. [Google Scholar] [CrossRef]
- Hiraoka, T.; Clark, J.I.; Li, X.Y.; Thurston, G.M. Effect of selected anti-cataract agents on opacification in the selenite cataract model. Exp. Eye Res. 1996, 62, 11–20. [Google Scholar] [CrossRef]
- Kuck, J.F.R.; KUCK, K.D. The Emory mouse cataract: The effects on cataractogenesis of α-tocopherol, penicillamine, triethylenetetramine, and mercaptopropionylglycine. J. Ocul. Pharmacol. Ther. 1988, 4, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hess, J.L.; Bunce, G.E. Deferoxamine effect on selenite-induced cataract formation in rats. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2511–2519. [Google Scholar]
- Juranek, I.; Rackova, L.; Stefek, M. Stobadine—An Indole Type Alternative to the Phenolic Antioxidant Reference Trolox. In Biochemistry; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef]
- Abe, M.; Reiter, R.J.; Orhii, P.B.; Hara, M.; Poeggeler, B. Inhibitory effect of melatonin on cataract formation in newborn rats: Evidence for an antioxidative role for melatonin. J. Pineal Res. 1994, 17, 94–100. [Google Scholar] [CrossRef]
- Nath, R.; Srivastava, S.; Singh, K. Accumulation of copper and inhibition of lactate dehydrogenase activity in human senile cataractous lens. Indian J. Exp. Biol. 1969, l7, 25–26. [Google Scholar]
- Behndig, A.; Karlsson, K.; Johansson, B.O.; Brannstrom, T.; Marklund, S.L. Superoxide dismutase isoenzymes in the normal and diseased human cornea. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2293–2296. [Google Scholar]
- Jeru, I. The role of zinc in the appearance of cataract. Ophthalmology 1997, 41, 329–332. [Google Scholar]
- Barman, S.; Srinivasan, K. Zinc Supplementation Ameliorates Diabetic Cataract Through Modulation of Crystallin Proteins and Polyol Pathway in Experimental Rats. Biol. Trace Elem. Res. 2019, 41, 329–332. [Google Scholar] [CrossRef]
- Baha’a, A.; Alzubaidy, A.A. Role of Topically-Applied Zinc Sulfate in Prevention of Sodium Selenite-Induced Cataract in Rabbits. Int. J. Adv. Res. 2014, 2, 1014–1022. [Google Scholar]
- Letavayova, L.; Vlčková, V.; Brozmanova, J. Selenium: From cancer prevention to DNA damage. Toxicological 2006, 227, 1–14. [Google Scholar] [CrossRef]
- Karaküçük, S.; Mirza, G.E.; Ekinciler, O.F.; Saraymen, R.; Karaküçük, I.; Ustdal, M. Selenium concentrations in serum, lens and aqueous humour of patients with senile cataract. Acta Ophthalmol. Scand. 1995, 73, 329–332. [Google Scholar] [CrossRef]
- Lapchak, P.A.; Zivin, J.A. Ebselen, a seleno-organic antioxidant, is neuroprotective after embolic strokes in rabbits: Synergism with low-dose tissue plasminogen activator. Stroke 2003, 34, 2013–2018. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aydemir, O.; Güler, M.; Kaya, M.K.; Deniz, N.; Üstündağ, B. Protective effects of ebselen on sodium-selenite-induced experimental cataract in rats. J. Cataract Refract. Surg. 2012, 38, 2160–2166. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Devamanoharan, P.S.; Henein, M.; Ali, A.H.; Varma, S.D. Diabetes-induced biochemical changes in rat lens: Attenuation of cataractogenesis by pyruvate. Diabetes Obes. Metab. 2000, 2, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Aydin, B.; Yagci, R.; Yılmaz, F.M.; Erdurmuş, M.; Karadag, R.; Keskin, U.; Durmuş, M.; Yigitoglu, R. Prevention of Selenite-Induced Cataractogenesis by N-Acetylcysteine in Rats. Curr. Eye Res. 2009, 34, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Tuzcu, E.A.; Tuzcu, K.; Basarslan, F.; Motor, S.; Coskun, M.; Keskin, U.; Ayintap, E.; Ilhan, Ö.; Oksuz, H.; Tuzcu, K.; et al. Protective effects of N-acetylcysteine on triamcinolone acetonide-induced lens damage in rats. Cutan. Ocul. Toxicol. 2014, 33, 294–298. [Google Scholar] [CrossRef]
- Zhang, S.; Chai, F.Y.; Yan, H.; Guo, Y.; Harding, J.J. Effects of N-acetylcysteine and glutathione ethyl ester drops on streptozotocin-induced diabetic cataract in rats. Mol. Vis. 2008, 14, 862–870. [Google Scholar]
- Maddirala, Y.; Tobwala, S.; Karacal, H.; Ercal, N. Prevention and reversal of selenite-induced cataracts by N-acetylcysteine amide in Wistar rats. BMC Ophthalmol. 2017, 17, 54. [Google Scholar] [CrossRef]
- Carey, J.W.; Pinarci, E.Y.; Penugonda, S.; Karacal, H.; Ercal, N. In vivo inhibition of l-buthionine-(S,R)-sulfoximine-induced cataracts by a novel antioxidant, N-acetylcysteine amide. Free Radic. Biol. Med. 2011, 50, 722–729. [Google Scholar] [CrossRef]
- Elanchezhian, R.; Ramesh, E.; Sakthivel, M.; Isai, M.; Geraldine, P.; Rajamohan, M.; Jesudasan, C.N.; Thomas, P.A. Acetyl-l-Carnitine Prevents Selenite-Induced Cataractogenesis in an Experimental Animal Model. Curr. Eye Res. 2007, 32, 961–971. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Deyev, A.I.; Yermakova, V.N.; Brikman, I.V.; Bours, J. Lipid peroxidation and cataracts. Drugs R D 2004, 5, 125–139. [Google Scholar] [CrossRef]
- Anand, D.A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharm. Rev. 2016, 10, 84–89. [Google Scholar]
- Isai, M.; Sakthivel, M.; Ramesh, E.; Thomas, P.A.; Geraldine, P. Prevention of selenite-induced cataractogenesis by rutin in Wistar rats. Mol. Vis. 2009, 15, 2570–2577. [Google Scholar] [PubMed]
- Ohtsuki, K.; Abe, A.; Mitsuzumi, H.; Kondo, M.; Uemura, K.; Iwasaki, Y.; Kondo, Y. Effects of Long-Term Administration of Hesperidin and Glucosyl Hesperidin to Spontaneously Hypertensive Rats. J. Nutr. Sci. Vitaminol. 2002, 48, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Miwa, Y.; Mitsuzumi, H.; Sunayama, T.; Yamada, M.; Okada, K.; Kubota, M.; Chaen, H.; Mishima, Y.; Kibata, M. Glucosyl hesperidin lowers serum triglyceride level in hypertriglyceridemic subjects through the improvement of very low-density lipoprotein metabolic abnormality. J. Nutr. Sci. Vitaminol. 2005, 51, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, Y.; Oka, M.; Bando, M.; Takehana, M. Hesperetin prevents selenite-induced cataract in rats. Mol. Vis. 2015, 21, 804–810. [Google Scholar] [PubMed]
- Nakazawa, Y.; Oka, M.; Tamura, H.; Takehana, M. Effect of hesperetin on chaperone activity in selenite-induced cataract. Open Med. 2016, 11, 183–189. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Pauze, M.; Fukuyama, K.; Nagai, N.; Funakoshi-Tago, M.; Sugai, T.; Tamura, H. Effect of hesperetin derivatives on the development of selenite-induced cataracts in rats. Mol. Med. Rep. 2018, 18, 1043–1050. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Henning, S.M.; Niu, Y.; Zhang, Y.; Nair, M.G.; Heber, D. In vitro antiproliferative, apoptotic and antioxidant activities of punicalagin, ellagic acid and a total pomegranate tannin extract are enhanced in combination with other polyphenols as found in pomegranate juice. J. Nutr. Biochem. 2005, 16, 360–367. [Google Scholar] [CrossRef]
- Narayanan, B.A.; Geoffroy, O.; Willingham, M.C.; Re, G.G.; Nixon, D.W. p53/p21(WAF1/CIP1) expression and its possible role in G1 arrest and apoptosis in ellagic acid treated cancer cells. Cancer Lett. 1999, 136, 215–221. [Google Scholar] [CrossRef]
- Zhang, H.-M.; Zhao, L.; Li, H.; Xu, H.; Chen, W.-W.; Tao, L. Research progress on the anticarcinogenic actions and mechanisms of ellagic acid. Cancer Boil. Med. 2014, 11, 92–100. [Google Scholar]
- Sakthivel, M.; Elanchezhian, R.; Ramesh, E.; Isai, M.; Jesudasan, C.N.; Thomas, P.; Geraldine, P. Prevention of selenite-induced cataractogenesis in Wistar rats by the polyphenol, ellagic acid. Exp. Eye Res. 2008, 86, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Thiagarajan, G.; Chandani, S.; Sundari, C.; Rao, S.; Kulkarni, A.V.; Balasubramanian, D. Antioxidant Properties of Green and Black Tea, and their Potential Ability to Retard the Progression of Eye Lens Cataract. Exp. Eye Res. 2001, 73, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Zhang, J. Black and green teas equally inhibit diabetic cataracts in a streptozotocin-induced rat model of diabetes. J. Agric. Food Chem. 2005, 53, 3710–3713. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.; Grgic, J. Caffeine and exercise: What next? Sports Med. 2019, 49, 1007–1030. [Google Scholar] [CrossRef] [PubMed]
- De Giuseppe, R.; Di, N.I.; Granata, F.; Mottolese, A.; Cena, H. Caffeine and blood pressure: A critical review perspective. Nutr. Res. Rev. 2019, 32, 169–175. [Google Scholar] [CrossRef]
- Kronschläger, M.; Galichanin, K.; Ekström, J.; Lou, M.F.; Söderberg, P.G. Protective Effect of the Thioltransferase Gene on In Vivo UVR-300 nm–Induced Cataract. Investig. Ophthalmol. Vis. Sci. 2012, 53, 248–252. [Google Scholar] [CrossRef]
- Kronschläger, M.; Löfgren, S.; Yu, Z.; Talebizadeh, N.; Varma, S.D.; Söderberg, P. Caffeine eye drops protect against UV-B cataract. Exp. Eye Res. 2013, 113, 26–31. [Google Scholar] [CrossRef]
- Kronschläger, M.; Forsman, E.; Yu, Z.; Talebizadeh, N.; Löfgren, S.; Meyer, L.M.; Bergquist, J.; Söderberg, P. Pharmacokinetics for topically applied caffeine in the rat. Exp. Eye Res. 2014, 122, 94–101. [Google Scholar] [CrossRef]
- El Okda, E.A.; Mohamed, M.M.; Shaheed, E.B.; Abdel-Moemin, A.R. Switching to instant black coffee modulates sodium selenite-induced cataract in rats. Ger. Med. Sci. 2016, 14, Doc 05. [Google Scholar]
- Mares-Periman, J.A.; Brady, W.E.; Klein, B.E.K.; Klein, R.; Haus, G.J.; Palta, M.; Ritter, L.L.; Shoff, S.M.; Mares-Perlman, J.A. Diet and Nuclear Lens Opacities. Am. J. Epidemiol. 1995, 141, 322–334. [Google Scholar] [CrossRef]
- Cumming, R.G.; Mitchell, P.; Smith, W. Diet and cataract: The blue mountains eye study. Ophthalmology 2000, 107, 450–456. [Google Scholar] [CrossRef]
- Gupta, S.K.; Trivedi, D.; Srivastava, S.; Joshi, S.; Halder, N.; Verma, S.D. Lycopene attenuates oxidative stress induced experimental cataract development: An in vitro and in vivo study. Nutrition 2003, 19, 794–799. [Google Scholar] [CrossRef]
- Suryanarayana, P.; Krishnaswamy, K.; Reddy, G.B. Effect of curcumin on galactose-induced cataractogenesis in rats. Mol. Vis. 2003, 9, 223–230. [Google Scholar] [PubMed]
- Padmaja, S.; Raju, T.N. Antioxidant effect of curcumin in selenium induced cataract of Wistar rats. Indian J. Exp. Biol. 2004, 42, 601–603. [Google Scholar]
- Manikandan, R.; Thiagarajan, R.; Beulaja, S.; Sudhandiran, G.; Arumugam, M. Effect of curcumin on selenite-induced cataractogenesis in Wistar rat pups. Curr. Eye Res. 2010, 35, 122–129. [Google Scholar] [CrossRef]
- Suryanarayana, P.; Saraswat, M.; Mrudula, T.; Krishna, T.P.; Krishnaswamy, K.; Reddy, G.B. Curcumin and Turmeric Delay Streptozotocin-Induced Diabetic Cataract in Rats. Investig. Opthalmol. Vis. Sci. 2005, 46, 2092–2099. [Google Scholar] [CrossRef]
- Phenol-Explorer Showing All Foods in Which the Polyphenol Resveratrol Is Found. 2019. Available online: http://phenol-explorer.eu/contents/polyphenol/592 (accessed on 11 January 2020).
- Vang, O.; Ahmad, N.; Baile, C.A.; Baur, J.A.; Brown, K.; Csiszar, A.; Das, D.K.; Delmas, M.; Gottfried, C.; Lin, H.-Y.; et al. What Is New for an Old Molecule? Systematic Review and Recommendations on the Use of Resveratrol. PLoS ONE 2011, 6, e19881. [Google Scholar] [CrossRef]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef]
- Doganay, S.; Borazan, M.; Iraz, M.; Cigremis, Y. The effect of resveratrol in experimental cataract model formed by sodium selenite. Curr. Eye Res. 2006, 31, 147–153. [Google Scholar] [CrossRef]
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Asp. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef]
- Rahmatullah, M.; Das, A.K.; Mollik, M.A.H.; Jahan, R.; Khan, M.; Rahman, T.; Chowdhury, M.H. An ethnomedicinal survey of Dhamrai sub-district in Dhaka District, Bangladesh. Am. Eurasian J. Sustain. Agric. 2009, 3, 881–888. [Google Scholar]
- Kyei, S.; Koffuor, G.A.; Ramkissoon, P.; Afari, C.; Asiamah, E.A. The Claim of Anti-Cataract Potential of Heliotropiumindicum: A Myth or Reality? Ophthalmol. Ther. 2015, 4, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Shojaii, A.; Azizi, H.; Ghafari, S.; Ghods, R.; Salmanian, M.; Ghafarzadeh, J. A review study on pharmacological activities, chemical constituents, and traditional uses of Echium amoenum. Pharmacogn. Rev. 2018, 12, 208. [Google Scholar] [CrossRef]
- NoroozpourDailami, K.; Azadbakht, M.; Lashgari, M.; Rashidi, Z. Prevention of selenite-induced cataractogenesis by hydroalchoholic extract of Echium amoenum: An experimental evaluation of the Iranian traditional eye medication. Pharm. Biomed. Res. 2015, 1, 40–47. [Google Scholar]
- Bagchi, D.; Bagchi, M.; Stohs, S.J.; Das, D.K.; Ray, S.D.; A Kuszynski, C.; Joshi, S.S.; Pruess, H.G. Free radicals and grape seed proanthocyanidin extract: Importance in human health and disease prevention. Toxicology 2000, 148, 187–197. [Google Scholar] [CrossRef]
- Bashir, N.; Shagirtha, K.; Manoharan, V.; Miltonprabu, S. The molecular and biochemical insight view of grape seed proanthocyanidins in ameliorating cadmium-induced testes-toxicity in rat model: Implication of PI3K/Akt/Nrf-2 signaling. Biosci. Rep. 2019, 39, 39. [Google Scholar] [CrossRef]
- Durukan, A.H.; Evereklioglu, C.; Hurmeric, V.; Kerimoglu, H.; Erdurman, C.; Bayraktar, M.Z.; Mumcuoğlu, T. Ingestion of IH636 grape seed proanthocyanidin extract to prevent selenite-induced oxidative stress in experimental cataract. J. Cataract. Refract. Surg. 2006, 32, 1041–1045. [Google Scholar] [CrossRef]
- Shukla, S.K.; Kumar, A.; Terrence, M.; Yusuf, J.; Singh, V.P.; Mishra, M. The probable use medicinal usage of Cassia Tora: An overview. OnLine J. Biol. Sci. 2013, 13, 13–17. [Google Scholar] [CrossRef]
- Ferlemi, A.-V.; Mermigki, P.G.; Makri, O.E.; Anagnostopoulos, D.; Koulakiotis, N.S.; Margarity, M.; Tsarbopoulos, A.; Georgakopoulos, C.D.; Lamari, F.N. Cerebral Area Differential Redox Response of Neonatal Rats to Selenite-Induced Oxidative Stress and to Concurrent Administration of Highbush Blueberry Leaf Polyphenols. Neurochem. Res. 2015, 40, 2280–2292. [Google Scholar] [CrossRef] [PubMed]
- Ferlemi, A.V.; Makri, O.E.; Mermigki, P.G.; Lamari, F.N.; Georgakopoulos, C.D. Quercetin glycosides and chlorogenic acid in highbush blueberry leaf decoction prevent cataractogenesis in vivo and in vitro: Investigation of the effect on calpains, antioxidant and metal chelating properties. Exp. Eye Res. 2016, 145, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Nguyen, V.K. Edible Wild Plants of Vietnam; Orchid Press: Bangkok, Thailand, 2007; ISBN 9745240893. [Google Scholar]
- Chopra, R.N.; Nayar, S.L.; Chopra, I.C. Glossary of Indian Medicinal Plants; National Institute of Science Communication, CSIR: New Delhi, India, 1996. [Google Scholar]
- Vallejo, F.; Tomas-Barberan, F.A.; Garcia-Viguera, C. Effect of climatic and sulphur fertilization conditions, on phenolic compounds and vitamin C in the influorescence of eight broccoli cultivars. Eur. Food Res. Technol. 2003, 216, 395–401. [Google Scholar] [CrossRef]
- Beecher, C. Cancer preventative properties of varieties of Brassica oleracea: A review. Am. J. Clin. Nut. 1994, 59, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Vibin, M.; Priya, S.G.S.; Rooban, B.N.; Sasikala, V.; Sahasranamam, V.; Abraham, A. Broccoli Regulates Protein Alterations and Cataractogenesis in Selenite Models. Curr. Eye Res. 2010, 35, 99–107. [Google Scholar] [CrossRef]
- Gallo, M.B.; Sarachine, M.J. Biological activities of lupeol. Int. J. Biomed. Pharm. Sci. 2009, 3, 46–66. [Google Scholar]
- Asha, R.; Devi, V.G.; Abraham, A. Lupeol, a pentacyclic triterpenoid isolated from Vernonia cinerea attenuate selenite induced cataract formation in Sprague Dawley rat pups. Chem. Biol. Interact. 2016, 245, 20–29. [Google Scholar] [CrossRef]
- Bayan, L.; Koulivand, P.H.; Gorji, A. Garlic: A review of potential therapeutic effects. Avicenna J. Phytomed. 2014, 4, 1–14. [Google Scholar] [PubMed]
- Raju, T.N.; Kanth, V.R.; Lavanya, K. Effect of methanolic extract of Allium sativum (AS) in delaying cataract in STZ-induced diabetic rats. J. Ocul. Biol. Dis. Inform. 2008, 1, 46–54. [Google Scholar] [CrossRef]
- Javadzadeh, A.; Ghorbanihaghjo, A.; Arami, S.; Rashtchizadeh, N.; Mesgari, M.; Rafeey, M.; Omidi, Y. Prevention of Selenite-Induced Cataractogenesis in Wistar Albino Rats by Aqueous Extract of Garlic. J. Ocul. Pharmacol. Ther. 2009, 25, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Rimbach, G.; Virgili, F. Antioxidant activity and biologic properties of a procyanidin-rich extract from pine (Pinus maritima) bark, pycnogenol. Free Radic. Biol. Med. 1999, 27, 704–724. [Google Scholar] [CrossRef]
- Choi, E.M. Antinociceptive and antiinflammatory activities of pine (Pinus densiflora) pollen extract. Phytother. Res. 2007, 21, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choung, S.Y. Pinus densiflora bark extract prevents selenite-induced cataract formation in the lens of Sprague Dawley rat pups. Mol. Vis. 2017, 23, 638–648. [Google Scholar] [PubMed]
- Al-Snafi, A.E. The pharmacology of Crocus sativus—A review. IOSR J. Pharm. 2016, 6, 8–38. [Google Scholar]
- Makri, O.E.; Ferlemi, A.V.; Lamari, F.N.; Georgakopoulos, C.D. Saffron administration prevents selenite-induced cataractogenesis. Mol. Vis. 2013, 19, 1188–1197. [Google Scholar]
- Grover, J.K.; Yadav, S.; Vats, V. Medicinal plants of India with anti-diabetic potential. J. Ethnopharmacol. 2002, 81, 81–100. [Google Scholar] [CrossRef]
- Vats, V.; Yadav, S.P.; Biswas, N.R.; Grover, J.K. Anti-cataract activity of Pterocarpus marsupium bark and Trigonellafoenum-graecum seeds extract in alloxan diabetic rats. J. Ethnopharmacol. 2004, 93, 289–294. [Google Scholar] [CrossRef]
- Zimmermann, M.; Colciaghi, F.; Cattabeni, F.; Di, M.L. Ginkgo biloba extract: From molecular mechanisms to the treatment of Alzhelmer’s disease. Cell. Mol. Biol. 2002, 48, 613–623. [Google Scholar]
- Smith, J.V.; Luo, Y. Studies on molecular mechanisms of Ginkgo biloba extract. Appl. Microbiol. Biotechnol. 2004, 64, 465–472. [Google Scholar]
- Ertekin, M.V.; Koçer, I.; Taysi, S.; Gepdiremen, A.; Sezen, O.; Balcı, E.; Bakan, N.; Karslıoğlu, I. Effects of Oral Ginkgo biloba Supplementation on Cataract Formation and Oxidative Stress Occurring in Lenses of Rats Exposed to Total Cranium Radiotherapy. Jpn. J. Ophthalmol. 2004, 48, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Thiagarajan, R.; Manikandan, R. Antioxidants and cataract. Free Radic. Res. 2013, 47, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Cotlier, E.; Sharma, Y. Aspirin and senile cataracts in rheumatoid arthritis. Lancet 1981, 317, 338–339. [Google Scholar] [CrossRef]
- Harding, J.J.; Egerton, M.; Harding, R.S. Protection against cataract by aspirin, paracetamol and ibuprofen. Acta Ophthalmol. 1989, 67, 518–524. [Google Scholar] [CrossRef]
- Gupta, S.K.; Joshi, S. Relationship between Aldose Reductase Inhibiting Activity and Anti-Cataract Action of Various Non-Steroidal Anti-Inflammatory Drugs1. Dev. Ophthalmol. 1991, 21, 151–156. [Google Scholar] [PubMed]
- Bono, A.; Militello, A.; Bongiorno, A.; Livrea, M.A.; Pandolfo, L. Effects of bendazac l-lysine salt on some metabolic enzymes of glutathione in the rabbit lens after X-irradiation. Ital. J. Biochem. 1987, 36, 153–165. [Google Scholar]
- Gupta, S.K.; Joshi, S.; Tandon, R.; Mathur, P. Topical aspirin provides protection against galactosemic cataract. Indian J. Ophthalmol. 1997, 45, 221–225. [Google Scholar]
- Eckerskorn, U.; Hockwin, O.; Müller-Breitenkamp, R.; Chen, T.T.; Knowles, W.; E Dobbs, R. Evaluation of cataract-related risk factors using detailed classification systems and multivariate statistical methods. Dev. Ophthalmol. 1987, 15, 82–91. [Google Scholar]
- Silvestrini, B. Recent Developments in the Pharmacological Treatment of Cataract; Dermo, F., Ponte, F., Laties, A.M., Eds.; Kugler: Amsterdam, The Netherlands, 1987; pp. 1–9. [Google Scholar]
- Guglielmotti, A.; De Joannon, A.C.; Cazzolla, N.; Marchetti, M.; Soldo, L.; Cavallo, G.; Pinza, M. Radical scavenger activity of bendazac, an anti-cataract non-steroidal anti-inflammatory agent. Pharmacol. Res. 1995, 32, 369–373. [Google Scholar] [CrossRef]
- Lewis, B.S.; Harding, J.J. The major metabolite of bendazac inhibits the glycosylation of soluble lens proteins: A possible mechanism for a delay in cataractogenesis. Exp. Eye Res. 1988, 47, 217–225. [Google Scholar] [CrossRef]
- Pandolfo, L.; Livrea, M.A.; Bono, A. Effects of bendazac l-lysine salt on X-ray-induced cataract in the rabbit lens. Exp. Eye Res. 1986, 42, 167–175. [Google Scholar] [CrossRef]
- Testa, M.; Iuliano, G.; Silvestrini, B. Pilot study of bendazac for treatment of cataract. Lancet 1982, 1, 849–850. [Google Scholar] [CrossRef]
- Gupta, S.K.; Agnihotri, S.; Joshi, S. Anti-cataract action of topical sulindac (IH-indene-3-acetic acid, 5 fluoro 2 methly methylene) in galactosemic rats. Afro Asian J. Ophthalmol. 1989, 8, 57–61. [Google Scholar]
- Gupta, S.K.; Joshi, S. Naproxen: An Aldose Reductase Inhibitor and Potential Anti-Cataract Agent. Dev. Ophthalmol. 1991, 21, 170–178. [Google Scholar] [PubMed]
- Gupta, S.K.; Joshi, S. Role of naproxen as anti-oxidant in selenite cataract. Ophthalmic Res. 1994, 26, 226–231. [Google Scholar] [CrossRef]
- Gupta, S.K.; Joshi, S. Prevention of photoperoxidation of lens lipids by naproxen. Afro Asian J. Ophthalmol. 1993, 12, 295–297. [Google Scholar]
- Lim, B.X.; Lim, C.H.; Lim, D.K.; Evans, J.R.; Bunce, C.; Wormald, R. Prophylactic Non-Steroidal Anti-Inflammatory Drugs for the Prevention of Macular Oedema after Cataract Surgery. Cochrane Database Syst. Rev. 2016, 11, CD006683. [Google Scholar] [CrossRef]
- Kessel, L.; Tendal, B.; Jørgensen, K.J.; Erngaard, D.; Flesner, P.; Andresen, J.L.; Hjortdal, J. Post-cataract prevention of inflammation and macular edema by steroid and nonsteroidal anti-inflammatory eye drops: A systematic review. Ophthalmology 2014, 121, 1915–1924. [Google Scholar] [CrossRef]
- Makley, L.N.; McMenimen, K.A.; DeVree, B.T.; Goldman, J.W.; McGlasson, B.N.; Rajagopal, P.; Dunyak, B.M.; McQuade, T.J.; Thompson, A.D.; Sunahara, R.; et al. Pharmacological chaperone for α-crystallin partially restores transparency in cataract models. Science 2015, 350, 674–677. [Google Scholar] [CrossRef]
- Langade, D.G.; Rao, G.; Girme, R.C.; Patki, P.S.; Bulakh, P.M. In vitro prevention by ACE inhibitors of cataract induced by glucose. Indian J. Pharm. 2006, l38, 107–110. [Google Scholar] [CrossRef]
- Qi, H.-P.; Wei, S.-Q.; Gao, X.-C.; Yu, N.-N.; Hu, W.-Z.; Bi, S.; Cui, H. Ursodeoxycholic acid prevents selenite-induced oxidative stress and alleviates cataract formation: In vitro and in vivo studies. Mol. Vis. 2012, 18, 151–160. [Google Scholar] [PubMed]
- Yang, C.X.; Yan, H.; Ding, T.B. Hydrogen saline prevents selenite-induced cataract in rats. Mol. Vis. 2013, 19, 1684–1693. [Google Scholar] [PubMed]
- Raj, S.M.; Vasavada, A.R.; Johar, S.R.; Vasavada, V.A.; Vasavada, V.A. Post-operative capsular opacification: A review. Int. J. Biomed. Sci. IJBS 2007, 3, 237–250. [Google Scholar] [PubMed]
- Sternberg, K.; Terwee, T.; Stachs, O.; Guthoff, R.; Löbler, M.; Schmitz, K.-P. Drug-Induced Secondary Cataract Prevention: Experimental ex vivo and in vivo Results with Disulfiram, Methotrexate and Actinomycin D. Ophthalmic Res. 2010, 44, 225–236. [Google Scholar] [CrossRef]
- Pei, C.; Xu, Y.; Jiang, J.X.; Cui, L.-J.; Li, L.; Qin, L. Application of sustained delivery microsphere of cyclosporine A for preventing posterior capsular opacification in rabbits. Int. J. Ophthalmol. 2013, 6, 1–7. [Google Scholar]
- Brown, C.J.; Akaichi, F. Vitamin D deficiency and posterior subcapsular cataract. Clin. Ophthalmol. 2015, 9, 1093–1098. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, X.-J.; Zhu, J.; Xi, Y.-B.; Yang, X.; Hu, L.-D.; Ouyang, H.; Patel, S.H.; Jin, X.; Lin, D.; et al. Lanosterol reverses protein aggregation in cataracts. Nature 2015, 523, 607–611. [Google Scholar] [CrossRef]
- Chew, E.Y.; Clemons, T.; SanGiovanni, J.P.; Danis, R.; Domalpally, A.; McBee, W.; Sperduto, R.; Ferris, F.L. AREDS2 Research Group The Age-Related Eye Disease Study 2 (AREDS2): Study design and baseline characteristics (AREDS2 report number 1). Ophthalmology 2012, 119, 2282–2289. [Google Scholar] [CrossRef]
- Mathew, M.C.; Ervin, A.-M.; Tao, J.; Davis, R.M. Antioxidant vitamin supplementation for preventing and slowing the progression of age-related cataract. Cochrane Database Syst. Rev. 2012, 6, CD004567. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).