Localization of 99mTc-GRP Analogs in GRPR-Expressing Tumors: Effects of Peptide Length and Neprilysin Inhibition on Biological Responses

 , and
, and 
Abstract
1. Introduction
2. Results
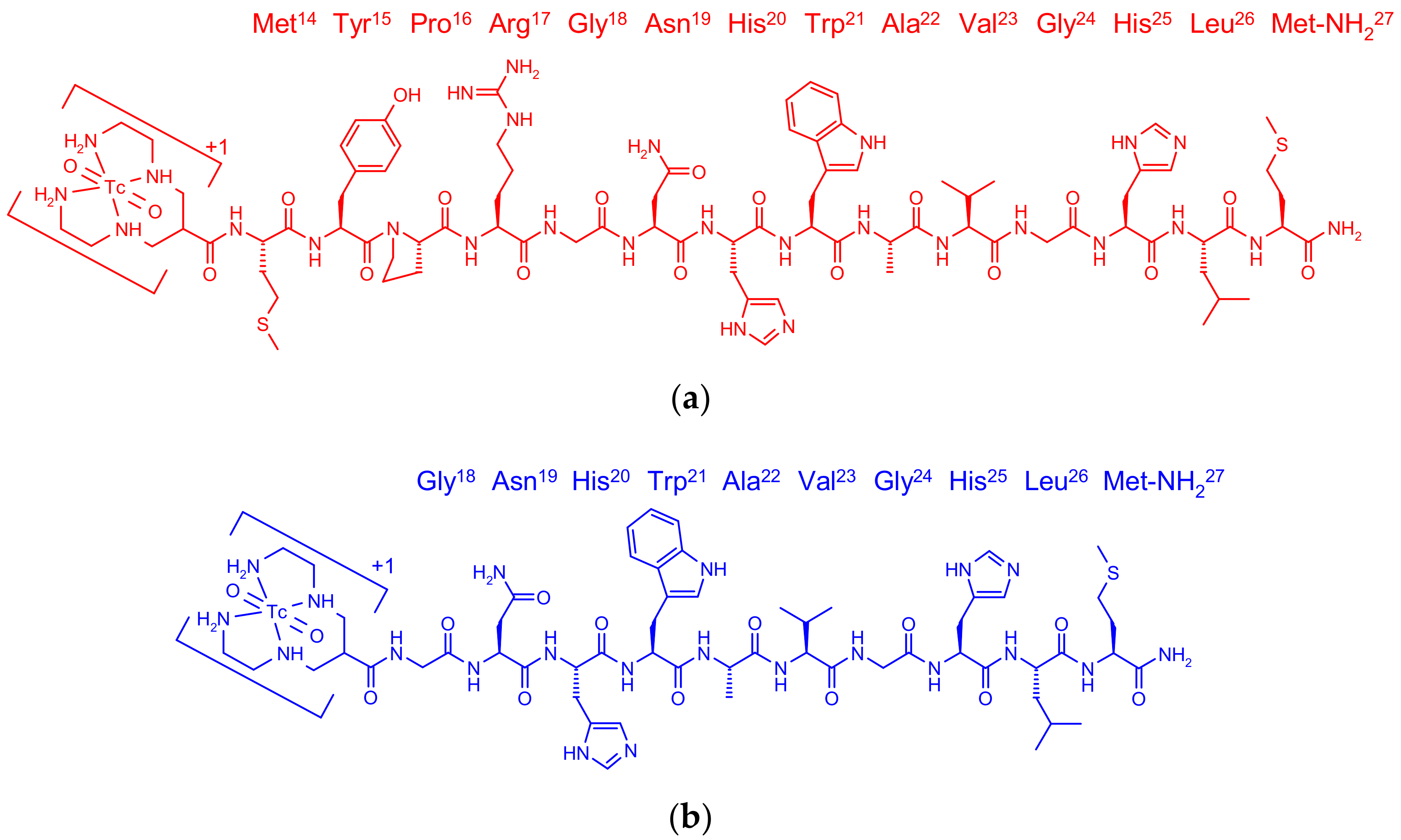
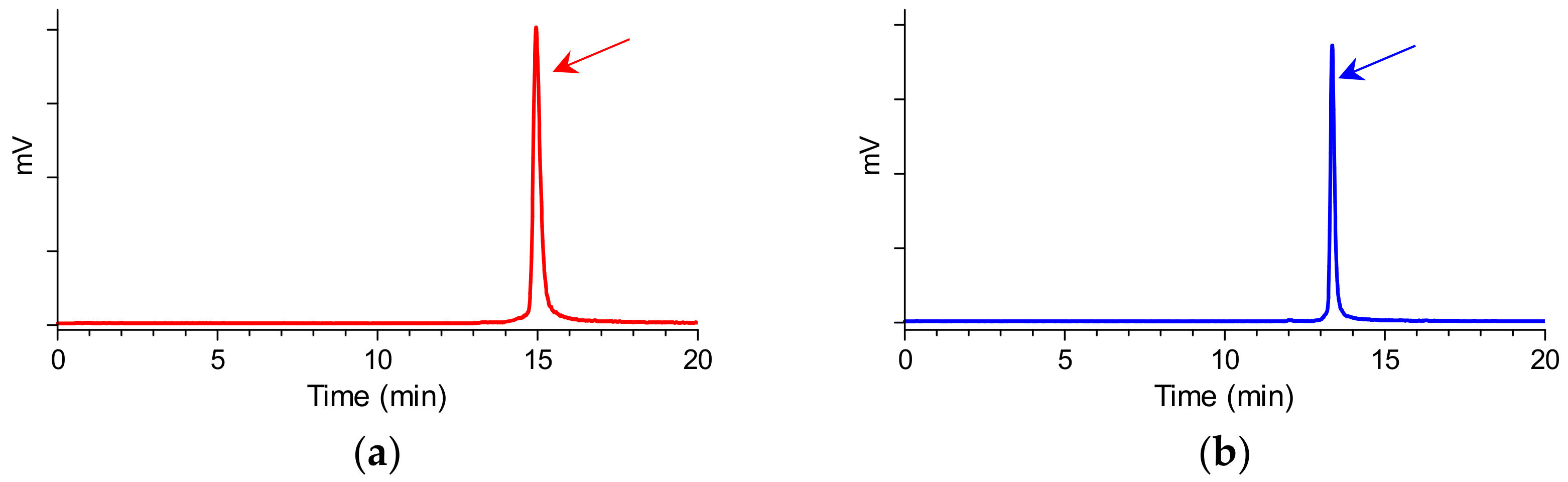
2.1. Peptides and Radioligands
2.2. In Vitro Assays
2.2.1. Receptor Autoradiography in Human Tumor Samples
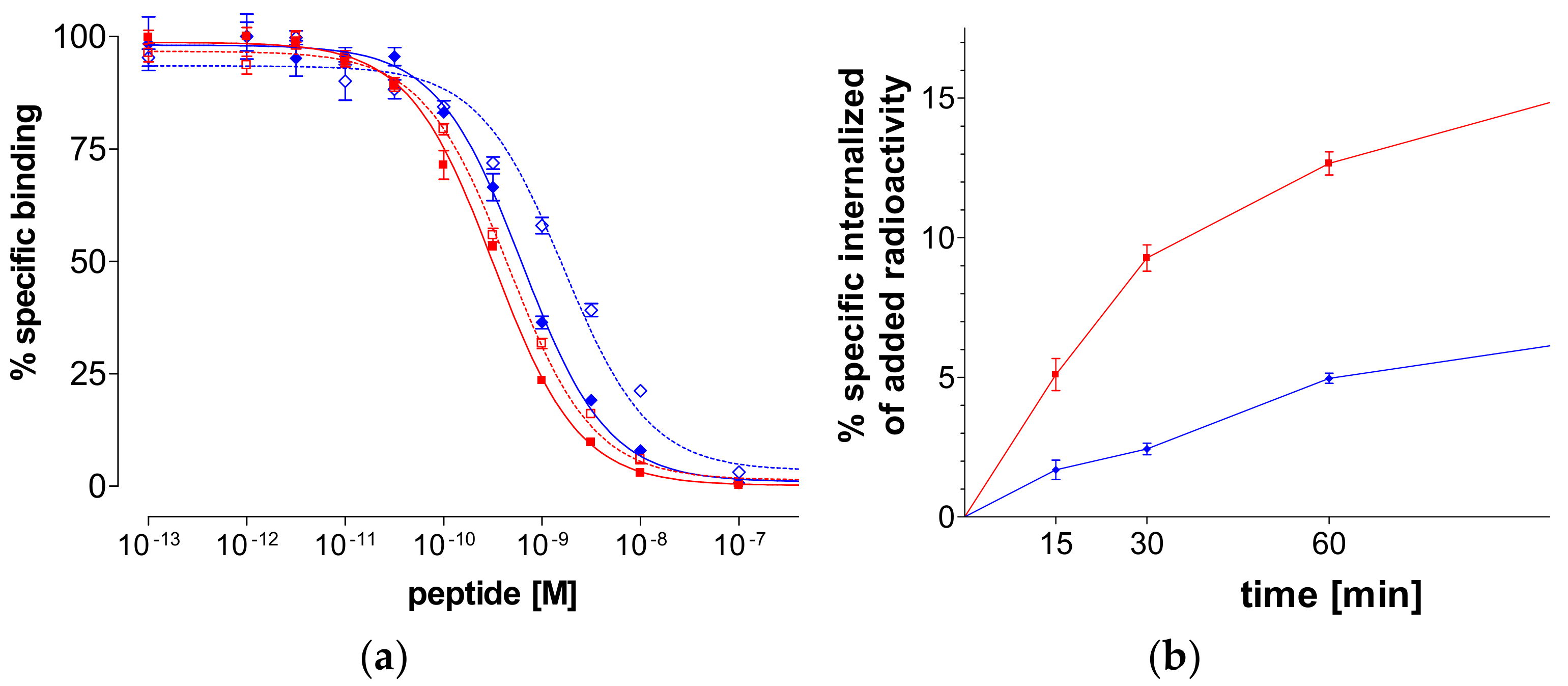
2.2.2. Binding Affinity for the Human GRPR
2.2.3. Internalization of 99mTc-N4-GRP(14–27) and 99mTc-N4-GRP(18–27) in PC-3 Cells
2.3. In Vivo Comparison of 99mTc-N4-GRP(14–27) and 99mTc-N4-GRP(18–27)
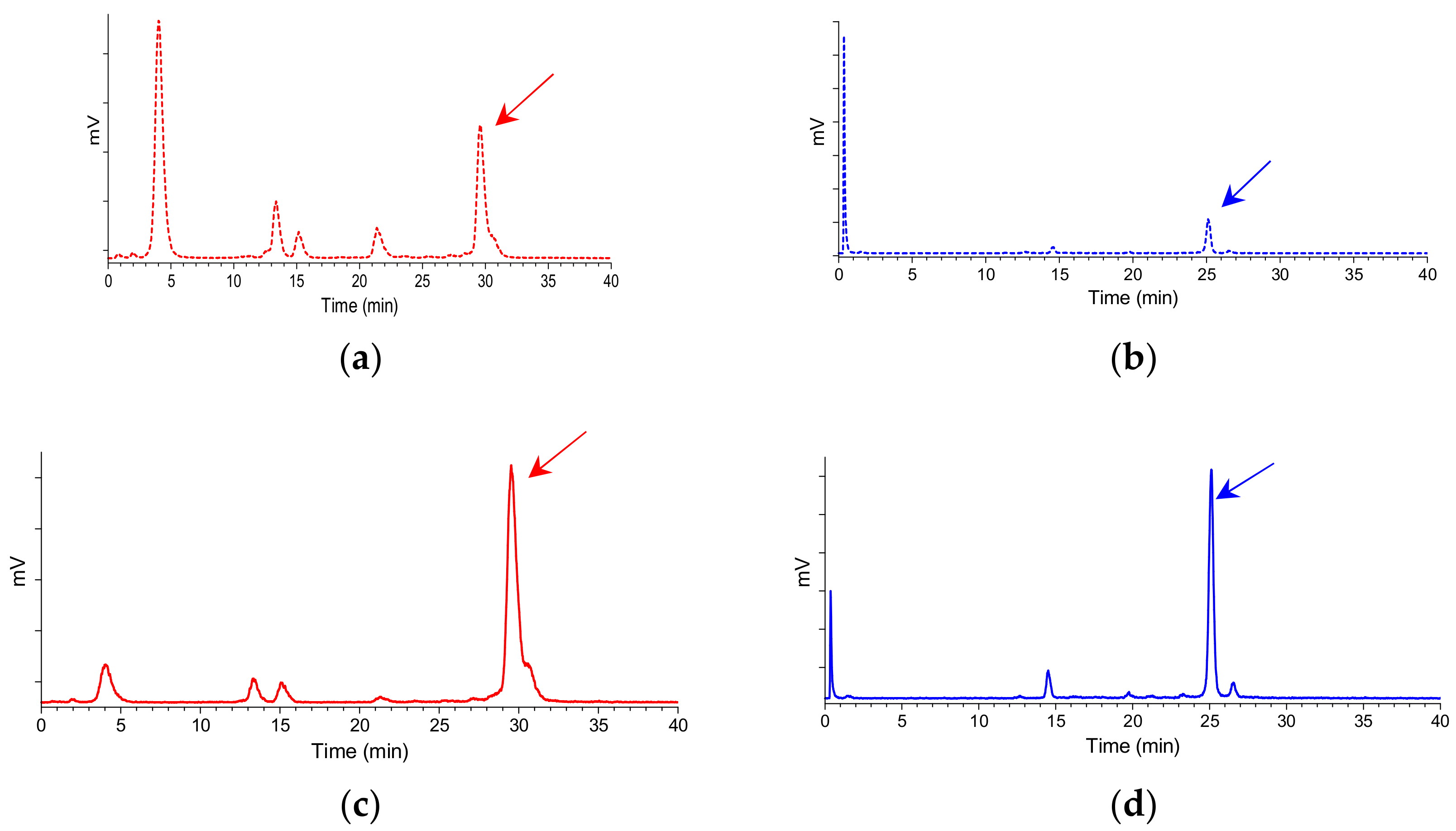
2.3.1. Stability of 99mTc-N4-GRP(14–27) and 99mTc-N4-GRP(18–27) in Mice
2.3.2. Biodistribution in PC-3 Xenograft-Bearing Mice
3. Discussion
4. Materials and Methods
4.1. Peptides and Reagents
Preparation and Quality Control of 99mTc-N4-GRP(14–27) and 99mTc-N4-GRP(18–27)
4.2. In Vitro Assays
4.2.1. Cell Lines and Culture
4.2.2. Receptor Autoradiography
4.2.3. Competition Binding in PC-3 Cell-Membranes
4.2.4. Internalization Assay in PC-3 Cells
4.3. Animal Studies
4.3.1. In Vivo Stability Tests
4.3.2. Induction of PC-3 Xenografts in SCID Mice
4.3.3. Biodistribution in PC-3 Xenograft-Bearing SCID Mice
Author Contributions
Funding
Conflicts of Interest
References
- Kroog, G.S.; Jensen, R.T.; Battey, J.F. Mammalian bombesin receptors. Med. Res. Rev. 1995, 15, 389–417. [Google Scholar] [CrossRef]
- Jensen, R.T.; Battey, J.F.; Spindel, E.R.; Benya, R.V. International union of pharmacology. LXVIII. Mammalian bombesin receptors: Nomenclature, distribution, pharmacology, signaling, and functions in normal and disease states. Pharmacol. Rev. 2008, 60, 1–42. [Google Scholar] [CrossRef]
- Markwalder, R.; Reubi, J.C. Gastrin-releasing peptide receptors in the human prostate: Relation to neoplastic transformation. Cancer Res. 1999, 59, 1152–1159. [Google Scholar] [PubMed]
- Körner, M.; Waser, B.; Rehmann, R.; Reubi, J.C. Early over-expression of GRP receptors in prostatic carcinogenesis. Prostate 2014, 74, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Beer, M.; Montani, M.; Gerhardt, J.; Wild, P.J.; Hany, T.F.; Hermanns, T.; Muntener, M.; Kristiansen, G. Profiling gastrin-releasing peptide receptor in prostate tissues: Clinical implications and molecular correlates. Prostate 2012, 72, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Reubi, J.C.; Wenger, S.; Schmuckli-Maurer, J.; Schaer, J.C.; Gugger, M. Bombesin receptor subtypes in human cancers: Detection with the universal radioligand 125I-[d-Tyr6,beta-Ala11,Phe13,Nle14]bombesin(6-14). Clin. Cancer Res. 2002, 8, 1139–1146. [Google Scholar]
- Halmos, G.; Wittliff, J.L.; Schally, A.V. Characterization of bombesin/gastrin-releasing peptide receptors in human breast cancer and their relationship to steroid receptor expression. Cancer Res. 1995, 55, 280–287. [Google Scholar] [PubMed]
- Gugger, M.; Reubi, J.C. Gastrin-releasing peptide receptors in non-neoplastic and neoplastic human breast. Am. J. Pathol. 1999, 155, 2067–2076. [Google Scholar] [CrossRef]
- Mattei, J.; Achcar, R.D.; Cano, C.H.; Macedo, B.R.; Meurer, L.; Batlle, B.S.; Groshong, S.D.; Kulczynski, J.M.; Roesler, R.; Dal Lago, L.; et al. Gastrin-releasing peptide receptor expression in lung cancer. Arch. Pathol. Lab. Med. 2014, 138, 98–104. [Google Scholar] [CrossRef]
- Guinee, D.G., Jr.; Fishback, N.F.; Koss, M.N.; Abbondanzo, S.L.; Travis, W.D. The spectrum of immunohistochemical staining of small-cell lung carcinoma in specimens from transbronchial and open-lung biopsies. Am. J. Clin. Pathol. 1994, 102, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Moreno, P.; Ramos-Alvarez, I.; Moody, T.W.; Jensen, R.T. Bombesin related peptides/receptors and their promising therapeutic roles in cancer imaging, targeting and treatment. Expert. Opin. Ther. Targets 2016, 20, 1055–1073. [Google Scholar] [CrossRef] [PubMed]
- Maina, T.; Nock, B.A. From bench to bed: New gastrin-releasing peptide receptor-directed radioligands and their use in prostate cancer. PET Clin. 2017, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Uehara, H.; Gonzalez, N.; Sancho, V.; Mantey, S.A.; Nuche-Berenguer, B.; Pradhan, T.; Coy, D.H.; Jensen, R.T. Pharmacology and selectivity of various natural and synthetic bombesin related peptide agonists for human and rat bombesin receptors differs. Peptides 2011, 32, 1685–1699. [Google Scholar] [CrossRef] [PubMed]
- Vigna, S.R.; Mantyh, C.R.; Giraud, A.S.; Soll, A.H.; Walsh, J.H.; Mantyh, P.W. Localization of specific binding sites for bombesin in the canine gastrointestinal tract. Gastroenterology 1987, 93, 1287–1295. [Google Scholar] [CrossRef]
- Chave, H.S.; Gough, A.C.; Palmer, K.; Preston, S.R.; Primrose, J.N. Bombesin family receptor and ligand gene expression in human colorectal cancer and normal mucosa. Br. J. Cancer 2000, 82, 124–130. [Google Scholar] [CrossRef]
- Fleischmann, A.; Laderach, U.; Friess, H.; Buechler, M.W.; Reubi, J.C. Bombesin receptors in distinct tissue compartments of human pancreatic diseases. Lab. Investig. 2000, 80, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Delle Fave, G.; Annibale, B.; de Magistris, L.; Severi, C.; Bruzzone, R.; Puoti, M.; Melchiorri, P.; Torsoli, A.; Erspamer, V. Bombesin effects on human GI functions. Peptides 1985, 6 (Suppl. 3), 113–116. [Google Scholar] [CrossRef]
- Bruzzone, R.; Tamburrano, G.; Lala, A.; Mauceri, M.; Annibale, B.; Severi, C.; de Magistris, L.; Leonetti, F.; Delle Fave, G. Effect of bombesin on plasma insulin, pancreatic glucagon, and gut glucagon in man. J. Clin. Endocrinol. Metab. 1983, 56, 643–647. [Google Scholar] [CrossRef]
- Delle Fave, G.; Kohn, A.; De Magistris, L.; Annibale, B.; Bruzzone, R.; Sparvoli, C.; Severi, C.; Torsoli, A. Effects of bombesin on gastrin and gastric acid secretion in patients with duodenal ulcer. Gut 1983, 24, 231–235. [Google Scholar] [CrossRef]
- Severi, C.; Jensen, R.T.; Erspamer, V.; D’Arpino, L.; Coy, D.H.; Torsoli, A.; Delle Fave, G. Different receptors mediate the action of bombesin-related peptides on gastric smooth muscle cells. Am. J. Physiol. 1991, 260, G683–G690. [Google Scholar] [CrossRef]
- Bitar, K.N.; Zhu, X.X. Expression of bombesin-receptor subtypes and their differential regulation of colonic smooth muscle contraction. Gastroenterology 1993, 105, 1672–1680. [Google Scholar] [CrossRef]
- Falconieri Erspamer, G.; Severini, C.; Erspamer, V.; Melchiorri, P.; Delle Fave, G.; Nakajima, T. Parallel bioassay of 27 bombesin-like peptides on 9 smooth muscle preparations. Structure-activity relationships and bombesin receptor subtypes. Regul. Pept. 1988, 21, 1–11. [Google Scholar] [CrossRef]
- Nock, B.A.; Cescato, R.; Ketani, E.; Waser, B.; Reubi, J.C.; Maina, T. [99mTc]Demomedin C, a radioligand based on human gastrin releasing peptide(18–27): Synthesis and preclinical evaluation in gastrin releasing peptide receptor-expressing models. J. Med. Chem. 2012, 55, 8364–8374. [Google Scholar] [CrossRef] [PubMed]
- Marsouvanidis, P.J.; Maina, T.; Sallegger, W.; Krenning, E.P.; de Jong, M.; Nock, B.A. 99mTc Radiotracers based on human GRP(18–27): Synthesis and comparative evaluation. J. Nucl. Med. 2013, 54, 1797–803. [Google Scholar] [CrossRef] [PubMed]
- Nock, B.A.; Nikolopoulou, A.; Galanis, A.; Cordopatis, P.; Waser, B.; Reubi, J.C.; Maina, T. Potent bombesin-like peptides for GRP-receptor targeting of tumors with 99mTc: A preclinical study. J. Med. Chem. 2005, 48, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Marsouvanidis, P.J.; Maina, T.; Sallegger, W.; Krenning, E.P.; de Jong, M.; Nock, B.A. Tumor diagnosis with new 111In-radioligands based on truncated human gastrin releasing peptide sequences: Synthesis and preclinical comparison. J. Med. Chem. 2013, 56, 8579–8587. [Google Scholar] [CrossRef]
- Roques, B.P.; Noble, F.; Dauge, V.; Fournie-Zaluski, M.C.; Beaumont, A. Neutral endopeptidase 24.11: Structure, inhibition, and experimental and clinical pharmacology. Pharmacol. Rev. 1993, 45, 87–146. [Google Scholar] [PubMed]
- Suda, H.; Aoyagi, T.; Takeuchi, T.; Umezawa, H. Letter: A thermolysin inhibitor produced by actinomycetes: Phosphoramidon. J. Antibiot. (Tokyo) 1973, 26, 621–623. [Google Scholar] [CrossRef]
- Oefner, C.; D’Arcy, A.; Hennig, M.; Winkler, F.K.; Dale, G.E. Structure of human neutral endopeptidase (neprilysin) complexed with phosphoramidon. J. Mol. Biol. 2000, 296, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Marsouvanidis, P.J.; Melis, M.; de Blois, E.; Breeman, W.A.; Krenning, E.P.; Maina, T.; Nock, B.A.; de Jong, M. In vivo enzyme inhibition improves the targeting of [177Lu]DOTA-GRP(13-27) in GRPR-positive tumors in mice. Cancer Biother. Radiopharm. 2014, 29, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Nock, B.A.; Maina, T.; Krenning, E.P.; de Jong, M. “To serve and protect”: Enzyme inhibitors as radiopeptide escorts promote tumor targeting. J. Nucl. Med. 2014, 55, 121–127. [Google Scholar] [CrossRef]
- Chatalic, K.L.; Konijnenberg, M.; Nonnekens, J.; de Blois, E.; Hoeben, S.; de Ridder, C.; Brunel, L.; Fehrentz, J.A.; Martinez, J.; van Gent, D.C.; et al. In vivo stabilization of a gastrin-releasing peptide receptor antagonist enhances pet imaging and radionuclide therapy of prostate cancer in preclinical studies. Theranostics 2016, 6, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Maina, T.; Kaloudi, A.; Valverde, I.E.; Mindt, T.L.; Nock, B.A. Amide-to-triazole switch vs. In vivo NEP-inhibition approaches to promote radiopeptide targeting of GRPR-positive tumors. Nucl. Med. Biol. 2017, 52, 57–62. [Google Scholar] [CrossRef]
- Lymperis, E.; Kaloudi, A.; Sallegger, W.; Bakker, I.L.; Krenning, E.P.; de Jong, M.; Maina, T.; Nock, B.A. Radiometal-dependent biological profile of the radiolabeled gastrin-releasing peptide receptor antagonist SB3 in cancer theranostics: Metabolic and biodistribution patterns defined by neprilysin. Bioconjug. Chem. 2018, 29, 1774–1784. [Google Scholar] [CrossRef]
- Nock, B.; Maina, T. Tetraamine-coupled peptides and resulting 99mTc-radioligands: An effective route for receptor-targeted diagnostic imaging of human tumors. Curr. Top. Med. Chem. 2012, 12, 2655–2667. [Google Scholar] [CrossRef] [PubMed]
- Mather, S.J.; Nock, B.A.; Maina, T.; Gibson, V.; Ellison, D.; Murray, I.; Sobnack, R.; Colebrook, S.; Wan, S.; Halberrt, G.; et al. GRP receptor imaging of prostate cancer using [99mTc]Demobesin 4: A first-in-man study. Mol. Imaging Biol. 2014, 16, 888–895. [Google Scholar] [CrossRef] [PubMed]
- De Castiglione, R.; Gozzini, L. Bombesin receptor antagonists. Crit. Rev. Oncol. Hematol. 1996, 24, 117–151. [Google Scholar] [CrossRef]
- Maina, T.; Nock, B.A.; Kulkarni, H.; Singh, A.; Baum, R.P. Theranostic prospects of gastrin-releasing peptide receptor-radioantagonists in oncology. PET Clin. 2017, 12, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Kassis, A.I. Therapeutic radionuclides: Biophysical and radiobiologic principles. Semin. Nucl. Med. 2008, 38, 358–366. [Google Scholar] [CrossRef]
- Wild, D.; Frischknecht, M.; Zhang, H.; Morgenstern, A.; Bruchertseifer, F.; Boisclair, J.; Provencher-Bolliger, A.; Reubi, J.C.; Maecke, H.R. Alpha- versus beta-particle radiopeptide therapy in a human prostate cancer model (213Bi-DOTA-Pesin and 213Bi-AMBA versus 177Lu-DOTA-Pesin). Cancer Res. 2011, 71, 1009–1018. [Google Scholar] [CrossRef]
- Linder, K.E.; Metcalfe, E.; Arunachalam, T.; Chen, J.; Eaton, S.M.; Feng, W.; Fan, H.; Raju, N.; Cagnolini, A.; Lantry, L.E.; et al. In vitro and in vivo metabolism of Lu-AMBA, a GRP-receptor binding compound, and the synthesis and characterization of its metabolites. Bioconjug. Chem. 2009, 20, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Shipp, M.A.; Tarr, G.E.; Chen, C.Y.; Switzer, S.N.; Hersh, L.B.; Stein, H.; Sunday, M.E.; Reinherz, E.L. CD10/neutral endopeptidase 24.11 hydrolyzes bombesin-like peptides and regulates the growth of small cell carcinomas of the lung. Proc. Natl. Acad. Sci. USA 1991, 88, 10662–10666. [Google Scholar] [CrossRef] [PubMed]
- Reile, H.; Armatis, P.E.; Schally, A.V. Characterization of high-affinity receptors for bombesin/gastrin releasing peptide on the human prostate cancer cell lines PC-3 and DU-145: Internalization of receptor bound 125I-(Tyr4)bombesin by tumor cells. Prostate 1994, 25, 29–38. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Conjugate | IC50s in nM | ||
|---|---|---|---|
| GRPR 1 | NMBR 2 | BB3R 3 | |
| Universal ligand 4 | 1.5 ± 0.1 (3) | 1.5 ± 0.2 (3) | 3.5 ± 0.7 (3) |
| N4-GRP(14–27) | 4.2 ± 1.0 (3) | 72 ± 7.6 (3) | >1000 (3) |
| N4-GRP(18–27) | 2.4 ± 1.0 (3) | 106 ± 13 (2) | >1000 (3) |
| Demobesin 3 | 0.5 (2) | 1.6 (2) | >100 (3) |
| Tissue | 1 h 1 | 4 h 1 | 24 h 1 | 4 h block 2 |
|---|---|---|---|---|
| Blood | 1.54 ± 0.17 | 0.10 ± 0.03 | 0.07 ± 0.01 | 0.08 ± 0.02 |
| Liver | 5.02 ± 0.46 | 3.75 ± 1.11 | 3.23 ± 0.78 | 6.12 ± 2.89 |
| Heart | 0.70 ± 0.07 | 0.11 ± 0.04 | 0.07 ± 0.01 | 0.28 ± 0.13 |
| Kidneys | 14.82 ± 2.22 | 4.83 ± 2.12 | 3.10 ± 0.73 | 4.85 ± 2.66 |
| Stomach | 1.02 ± 0.29 | 1.42 ± 0.93 | 0.78 ± 0.33 | 0.43 ± 0.18 |
| Intestines | 7.21 ± 0.52 | 7.61 ± 2.23 | 1.73 ± 0.23 | 1.45 ± 0.44 |
| Muscle | 0.29 ± 0.02 | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.02 |
| Lungs | 1.96 ± 0.33 | 0.49 ± 0.30 | 0.18 ± 0.04 | 0.65 ± 0.22 |
| Pancreas | 37.85 ± 1.95 | 35.24 ± 4.70 | 13.41 ± 0.77 | 0.83 ± 0.24 |
| Tumor | 10.20 ± 0.72 | 8.41 ± 4.16 | 4.50 ± 0.69 | 0.62 ± 0.24 |
| Tissue | 99mTc-N4-GRP(14–27) | 99mTc-N4-GRP(18–27) | ||
|---|---|---|---|---|
| 4 h 1 | 4 h PA 1,2 | 4 h1 | 4 h PA 1,2 | |
| Blood | 0.10 ± 0.03 | 0.23 ± 0.08 | 0.13 ± 0.04 | 0.21 ± 0.04 |
| Liver | 3.75 ± 1.11 | 4.99 ± 1.53 | 1.02 ± 0.17 | 2.13 ± 0.37 |
| Heart | 0.11 ± 0.04 | 0.89 ± 0.06 | 0.14 ± 0.10 | 0.47 ± 0.27 |
| Kidneys | 4.83 ± 2.12 | 11.68 ± 1.61 | 6.01 ± 1.38 | 7.66 ± 1.39 |
| Stomach | 1.42 ± 0.93 | 3.82 ± 1.10 | 1.12 ± 0.75 | 4.02 ± 0.66 |
| Intestines | 7.61 ± 2.23 | 21.35 ± 1.68 | 7.28 ± 0.60 | 16.23 ± 3.90 |
| Muscle | 0.03 ± 0.01 | 0.06 ± 0.02 | 0.03 ± 0.01 | 0.06 ± 0.01 |
| Lungs | 0.49 ± 0.30 | 1.06 ± 0.19 | 0.28 ± 0.09 | 0.84 ± 0.26 |
| Pancreas | 35.24 ± 4.70 | 110.32 ± 8.76 | 32.18 ± 5.91 | 95.39 ± 20.34 |
| Tumor | 8.41 ± 4.16 | 38.19 ± 4.79 | 7.08 ± 1.29 | 28.37 ± 8.05 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaloudi, A.; Lymperis, E.; Kanellopoulos, P.; Waser, B.; de Jong, M.; Krenning, E.P.; Reubi, J.C.; Nock, B.A.; Maina, T. Localization of 99mTc-GRP Analogs in GRPR-Expressing Tumors: Effects of Peptide Length and Neprilysin Inhibition on Biological Responses. Pharmaceuticals 2019, 12, 42. https://doi.org/10.3390/ph12010042
Kaloudi A, Lymperis E, Kanellopoulos P, Waser B, de Jong M, Krenning EP, Reubi JC, Nock BA, Maina T. Localization of 99mTc-GRP Analogs in GRPR-Expressing Tumors: Effects of Peptide Length and Neprilysin Inhibition on Biological Responses. Pharmaceuticals. 2019; 12(1):42. https://doi.org/10.3390/ph12010042
Chicago/Turabian StyleKaloudi, Aikaterini, Emmanouil Lymperis, Panagiotis Kanellopoulos, Beatrice Waser, Marion de Jong, Eric P. Krenning, Jean Claude Reubi, Berthold A. Nock, and Theodosia Maina. 2019. "Localization of 99mTc-GRP Analogs in GRPR-Expressing Tumors: Effects of Peptide Length and Neprilysin Inhibition on Biological Responses" Pharmaceuticals 12, no. 1: 42. https://doi.org/10.3390/ph12010042
APA StyleKaloudi, A., Lymperis, E., Kanellopoulos, P., Waser, B., de Jong, M., Krenning, E. P., Reubi, J. C., Nock, B. A., & Maina, T. (2019). Localization of 99mTc-GRP Analogs in GRPR-Expressing Tumors: Effects of Peptide Length and Neprilysin Inhibition on Biological Responses. Pharmaceuticals, 12(1), 42. https://doi.org/10.3390/ph12010042