

Luminescent Bacteria as Bioindicators in Screening and Selection of Enzymes Detoxifying Various Mycotoxins
 , , , , , and
, , , , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Production of Immobilized Photobacteria
2.3. Analysis of Toxicity by Luminescence Measurements
2.4. Treatment of Mycotoxins with Enzymes
2.5. Computer Modeling of Mycotoxin–Enzyme Interaction
3. Results
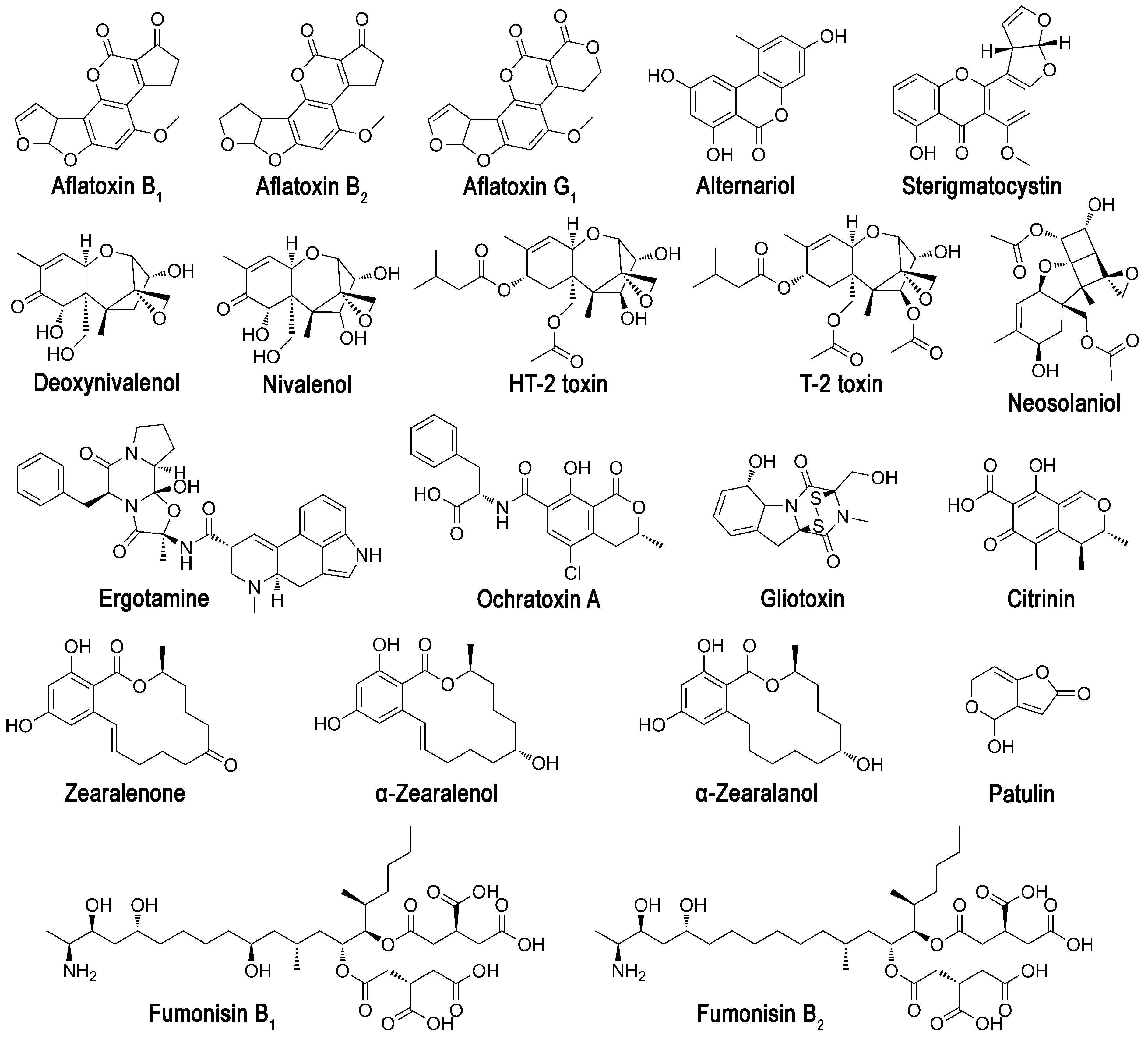
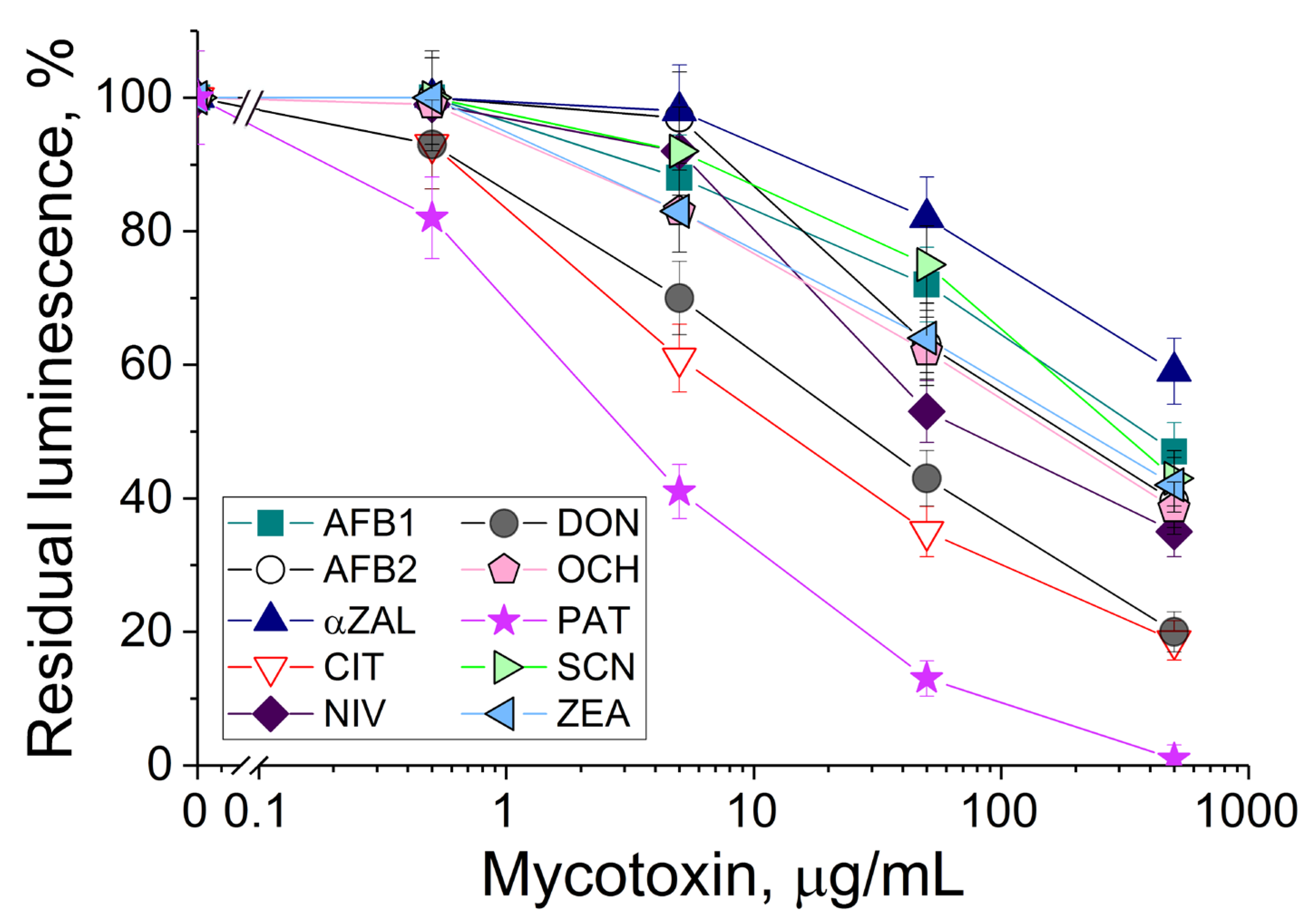
3.1. The Response of Bioluminescent Cells to the Presence of Different Mycotoxins
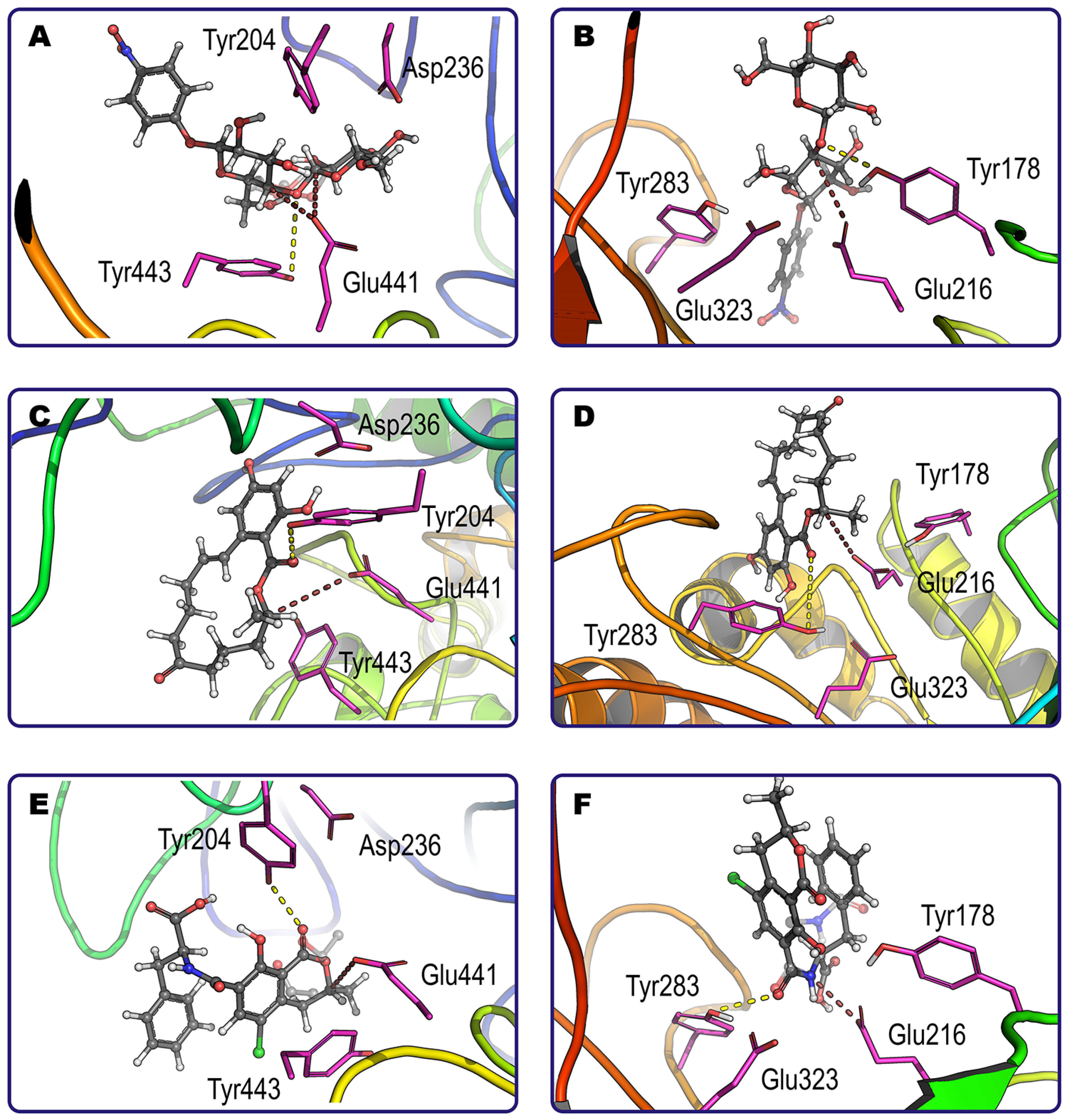
3.2. In silico Investigation of Interactions between Mycotoxins and Enzymes
3.3. Bioluminescent Sensing of Results of Enzymatic Action on Mycotoxins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats posed by the fungal kingdom to humans, wildlife, and agriculture. mBio 2020, 11, e00449-20. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.Z. Mycotoxins in food, recent development in food analysis and future challenges; a review. Curr. Opin. Food Sci. 2021, 42, 237–247. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Stela, M.; Saluk-Bijak, J.; Siadkowski, A.; Bijak, M. Molecular aspects of mycotoxins—A serious problem for human health. Int. J. Mol. Sci. 2020, 21, e8187. [Google Scholar] [CrossRef] [PubMed]
- Nešić, K.; Habschied, K.; Mastanjević, K. Possibilities for the biological control of mycotoxins in food and feed. Toxins 2021, 13, 198. [Google Scholar] [CrossRef] [PubMed]
- Nahle, S.; El Khoury, A.; Savvaidis, I.; Chokr, A.; Louka, N.; Atoui, A. Detoxification approaches of mycotoxins: By microorganisms, biofilms and enzymes. Int. J. Food Contam. 2022, 9, 9. [Google Scholar] [CrossRef]
- Lyagin, I.; Efremenko, E. Enzymes for detoxification of various mycotoxins: Origins and mechanisms of catalytic action. Molecules 2019, 24, 2362. [Google Scholar] [CrossRef] [PubMed]
- Lyagin, I.; Stepanov, N.; Maslova, O.; Senko, O.; Aslanli, A.; Efremenko, E. Not a mistake but a feature: Promiscuous activity of enzymes meeting mycotoxins. Catalysts 2022, 12, 1095. [Google Scholar] [CrossRef]
- Singh, G.; Verma, A.K.; Kumar, V. Catalytic properties, functional attributes and industrial applications of β-glucosidases. 3 Biotech 2016, 6, 3. [Google Scholar] [CrossRef]
- Cence, K.; dos Santos, P.; Garcia, M.V.; Copetti, M.V.; Valduga, E.; Cansian, R.L.; Zeni, J.; Backes, G.T. Enzymatic biocontrol of spoilage fungi from salami. LWT 2019, 115, e108457. [Google Scholar] [CrossRef]
- Liu, L.; Xie, M.; Wei, D. Biological detoxification of mycotoxins: Current status and future advances. Int. J. Mol. Sci. 2022, 23, 1064. [Google Scholar] [CrossRef]
- Lyagin, I.; Maslova, O.; Stepanov, N.; Efremenko, E. Degradation of mycotoxins in mixtures by combined proteinous nanobiocatalysts: In silico, in vitro and in vivo. Int. J. Biol. Macromol. 2022, 218, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Sandlin, N.; Kish, D.R.; Kim, J.; Zaccaria, M.; Momeni, B. Current and emerging tools of computational biology to improve the detoxification of mycotoxins. Appl. Environ. Microbiol. 2022, 88, 0210221. [Google Scholar] [CrossRef]
- Zaccaria, M.; Dawson, W.; Kish, D.R.; Reverberi, M.; Bonaccorsi di Patti, M.C.; Domin, M.; Cristiglio, V.; Chan, B.; Dellafiora, L.; Gabel, F.; et al. Experimental–theoretical study of laccase as a detoxifier of aflatoxins. Sci. Rep. 2023, 13, 860. [Google Scholar] [CrossRef]
- Hao, W.-B.; Gu, X.; Yu, X.; Zhao, Y.; Li, C.; Jia, M.; Du, X.-D. Laccase Lac-W detoxifies aflatoxin B1 and degrades five other major mycotoxins in the absence of redox mediators. Environ. Pollut. 2023, 338, 122581. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Cortés, P.C.; Flores-Ortíz, C.M.; Hernández-Portilla, L.B.; Vázquez Medrano, J.; Rodríguez-Peña, O.N. Molecular docking and in vitro studies of ochratoxin A (OTA) biodetoxification testing three endopeptidases. Molecules 2023, 28, 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xing, H.; Dongliang, L.; Han, M.; Cai, P.; Lin, H.; Tian, Y.; Guo, Y.; Sun, B.; Tian, Y.; et al. Deep learning enables rapid identification of mycotoxin-degrading enzymes. ChemRxiv 2023. [Google Scholar] [CrossRef]
- Karadurmus, L.; Kaya, S.I.; Ozkan, S.A. Recent advances of enzyme biosensors for pesticide detection in foods. Food Measure 2021, 15, 4582–4595. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Ma, L.; Yan, X.; Lei, Q. Dose-effect and structure-activity relationships of haloquinoline toxicity towards Vibrio fischeri. Environ. Sci. Pollut. Res. Int. 2022, 29, 10858–10864. [Google Scholar] [CrossRef]
- Efremenko, E.; Maslova, O.; Stepanov, N.; Ismailov, A. Using cholinesterases and immobilized luminescent Photobacteria for the express-analysis of mycotoxins and estimating the efficiency of their enzymatic hydrolysis. Toxins 2021, 13, 34. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, T.J.; Czodrowski, P.; Li, H.; Nielsen, J.E.; Jensen, J.H.; Klebe, G.; Baker, N.A. PDB2PQR: Expanding and upgrading automated preparation of biomolecular structures for molecular simulations. Nucleic Acids Res. 2007, 35, W522–W525. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Schmidt, M.W.; Baldridge, K.K.; Boatz, J.A.; Elbert, S.T.; Gordon, M.S.; Jensen, J.H.; Koseki, S.; Matsunaga, N.; Nguyen, K.A.; Su, S.; et al. General atomic and molecular electronic structure system. J. Comput. Chem. 1993, 14, 1347–1363. [Google Scholar] [CrossRef]
- Voevodin, V.; Antonov, A.; Nikitenko, D.; Shvets, P.; Sobolev, S.; Sidorov, I.; Stefanov, K.; Voevodin, V.; Zhumatiy, S. Supercomputer Lomonosov-2: Large Scale, Deep Monitoring and Fine Analytics for the User Community. Supercomput. Front. Innov. 2019, 6, 4–11. [Google Scholar] [CrossRef]
- Jian, Q.; Gong, L.; Li, T.; Wang, Y.; Wu, Y.; Chen, F.; Qu, H.; Duan, X.; Jiang, Y. Rapid assessment of the toxicity of fungal compounds using luminescent Vibrio qinghaiensis sp. q67. Toxins 2017, 9, 335. [Google Scholar] [CrossRef]
- Orobchenko, O.; Kurbatska, O.; Paliy, A.; Palii, A. Toxicological evaluation of feed contaminated with mycotoxins using a luminescent microorganism: Photobacterium phosphoreum. Veterinarska Stanica 2023, 54, 147–164. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef]
- Pauliukaite, R.; Voitechovič, E. Multisensor Systems and Arrays for Medical Applications Employing Naturally-Occurring Compounds and Materials. Sensors 2020, 20, 3551. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Efremenko, E.; Lyagin, I.; Stepanov, N.; Senko, O.; Maslova, O.; Aslanli, A.; Ugarova, N. Luminescent Bacteria as Bioindicators in Screening and Selection of Enzymes Detoxifying Various Mycotoxins. Sensors 2024, 24, 763. https://doi.org/10.3390/s24030763
Efremenko E, Lyagin I, Stepanov N, Senko O, Maslova O, Aslanli A, Ugarova N. Luminescent Bacteria as Bioindicators in Screening and Selection of Enzymes Detoxifying Various Mycotoxins. Sensors. 2024; 24(3):763. https://doi.org/10.3390/s24030763
Chicago/Turabian StyleEfremenko, Elena, Ilya Lyagin, Nikolay Stepanov, Olga Senko, Olga Maslova, Aysel Aslanli, and Natalia Ugarova. 2024. "Luminescent Bacteria as Bioindicators in Screening and Selection of Enzymes Detoxifying Various Mycotoxins" Sensors 24, no. 3: 763. https://doi.org/10.3390/s24030763
APA StyleEfremenko, E., Lyagin, I., Stepanov, N., Senko, O., Maslova, O., Aslanli, A., & Ugarova, N. (2024). Luminescent Bacteria as Bioindicators in Screening and Selection of Enzymes Detoxifying Various Mycotoxins. Sensors, 24(3), 763. https://doi.org/10.3390/s24030763