
Simultaneous Gut-Brain Electrophysiology Shows Cognition and Satiety Specific Coupling


Abstract
1. Introduction
2. Materials and Methods
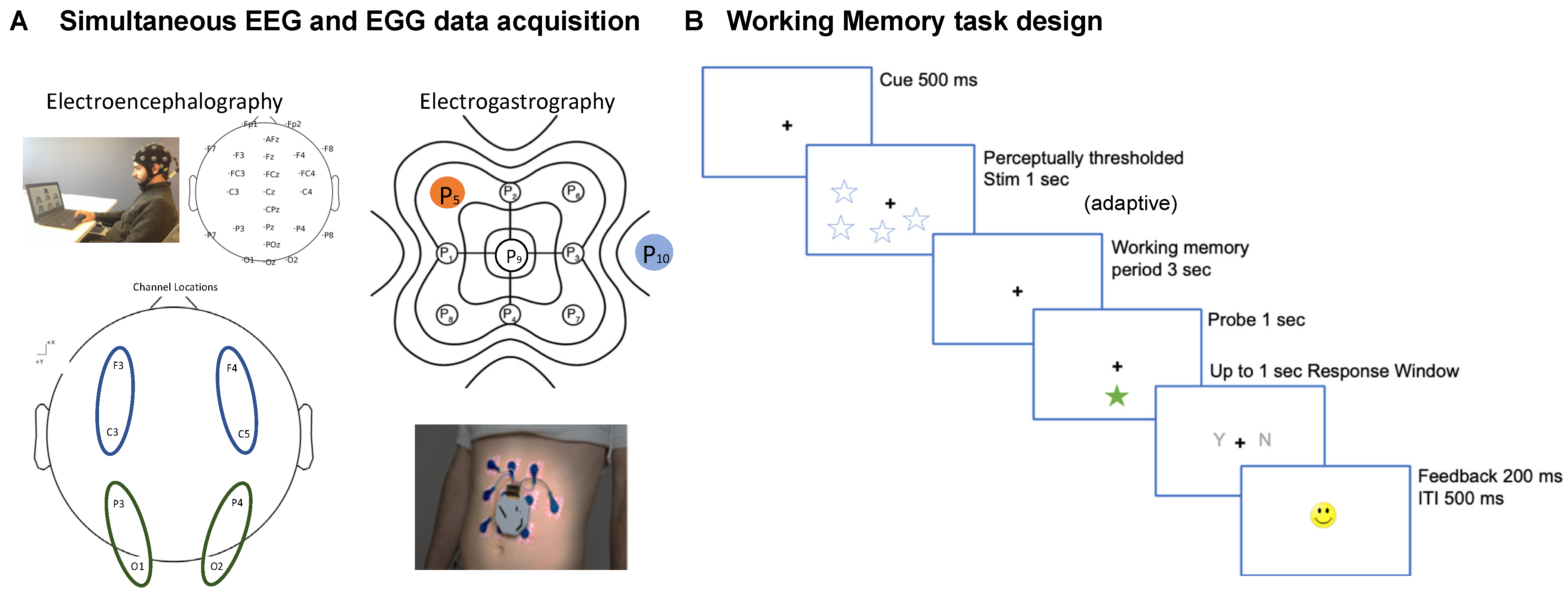
2.1. Procedure and Recording
2.2. Brain-Gut Coupling Analyses
2.3. Statistical Analysis
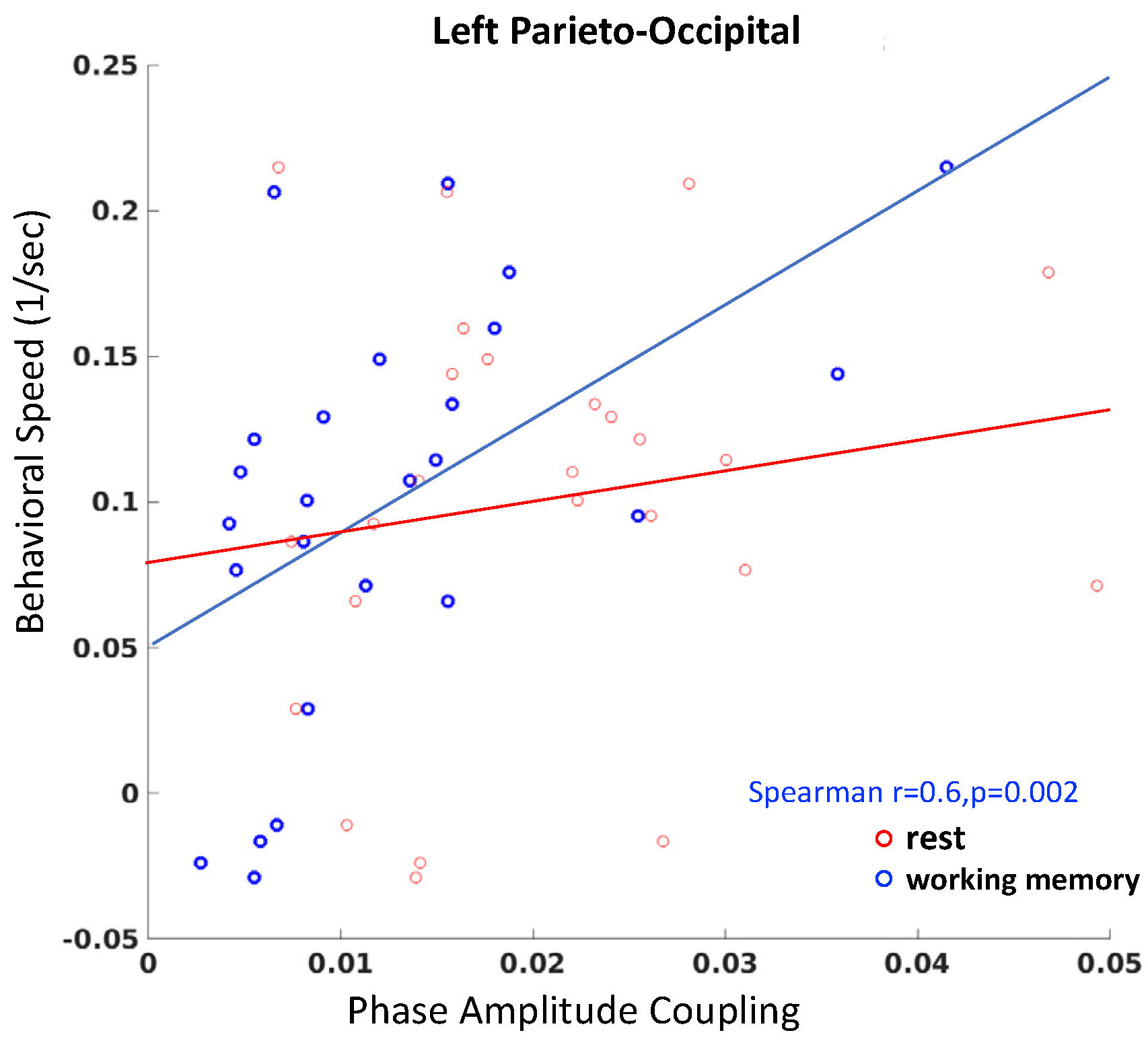
3. Results
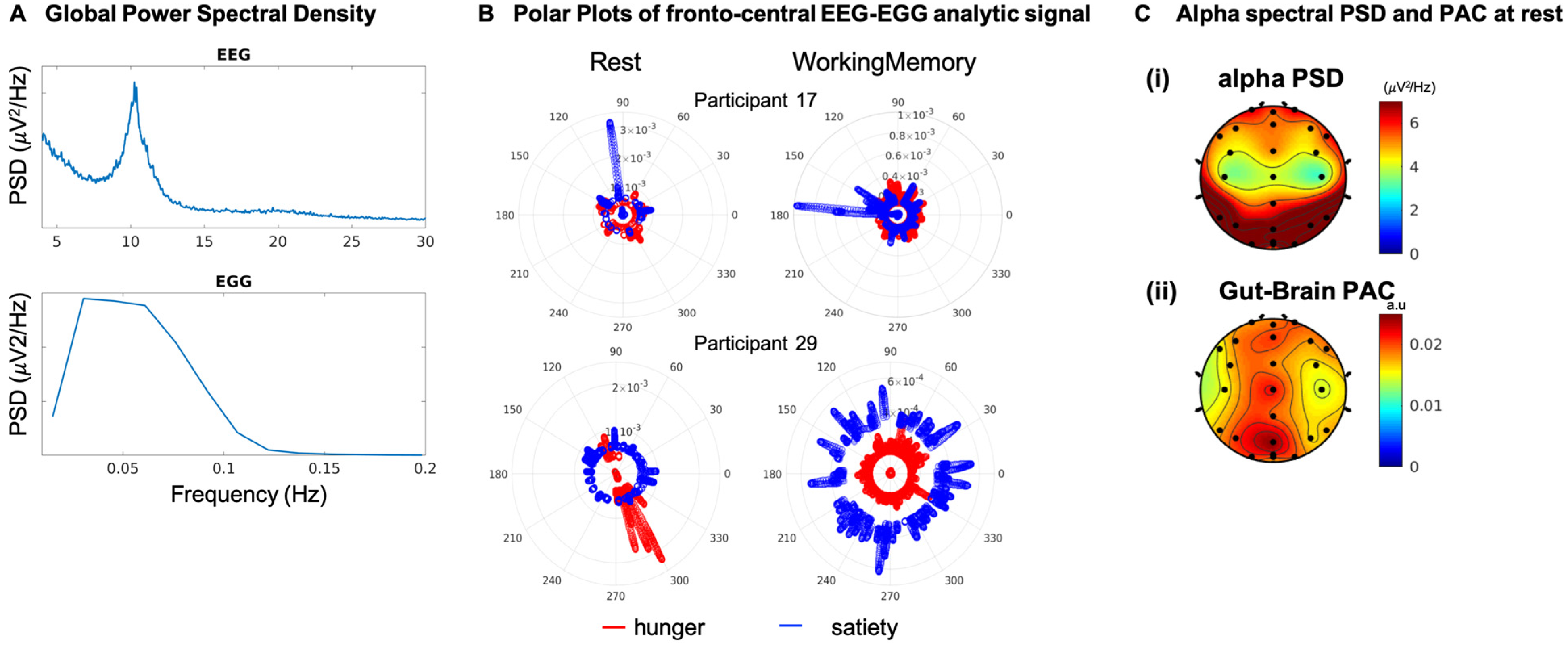
3.1. Permutation Control Analysis Reveal Significant Gut-Brain Coupling by Concurrent EEG-EGG
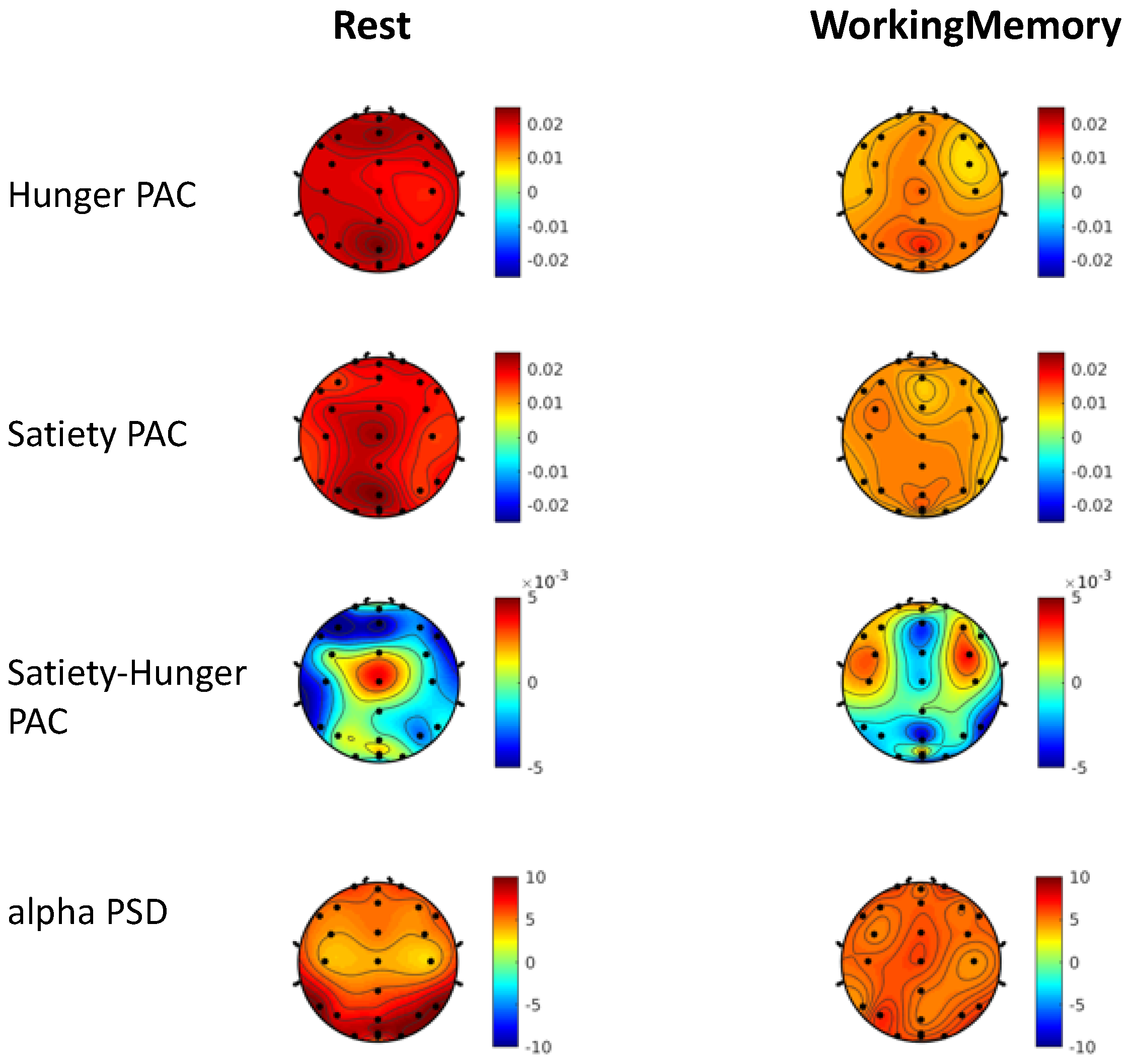
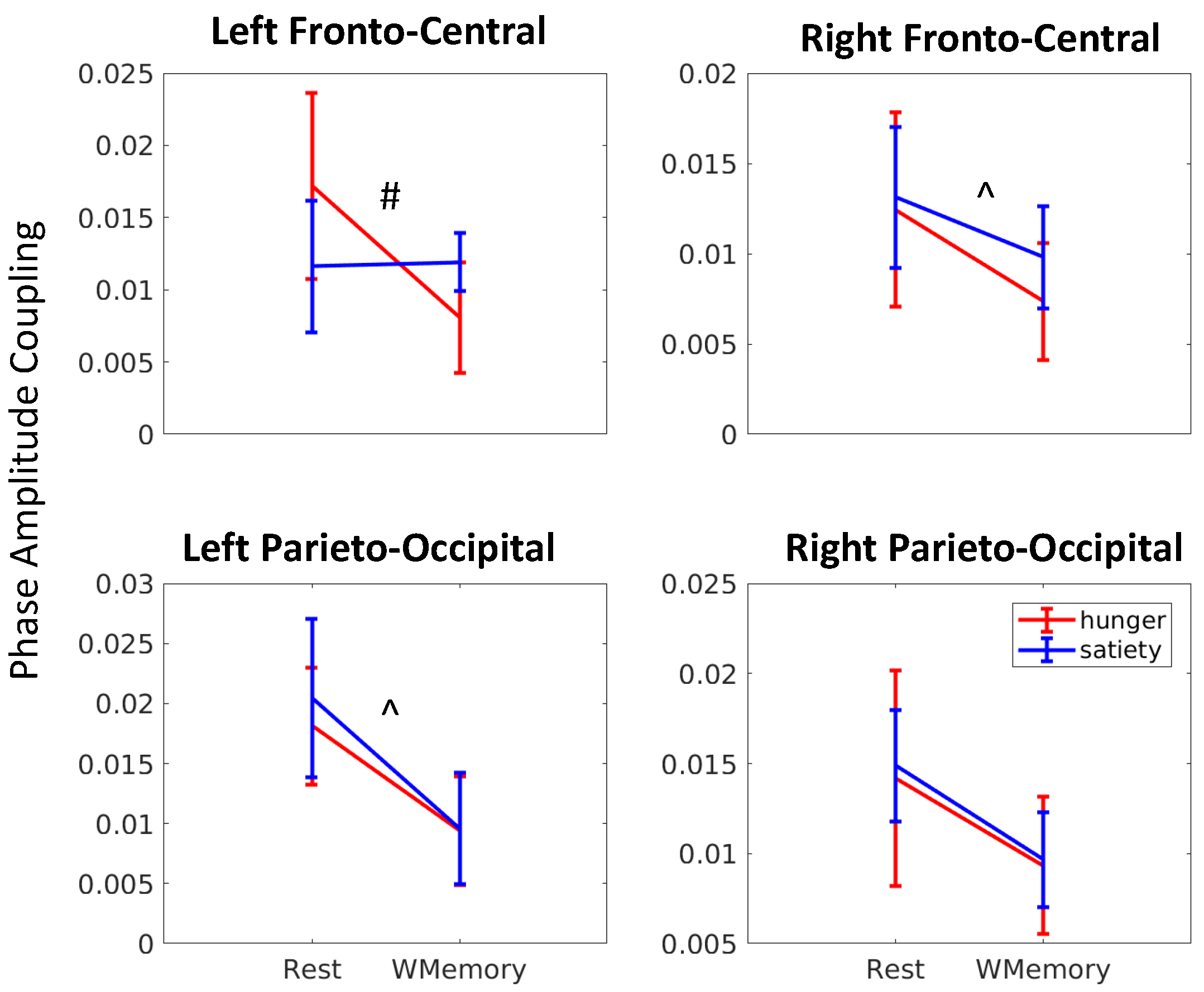
3.2. Gut-Brain Coupling Differs with Cognitive Engagement and Hunger State
3.3. Gut-Brain Coupling Relates to Behavioral Performance Speed
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayer, E.A. Gut feelings: The emerging biology of gut–brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.K.; Chang, R.B.; Strochlic, D.E.; Umans, B.D.; Lowell, B.B.; Liberles, S.D. Sensory neurons that detect stretch and nutrients in the digestive system. Cell 2016, 166, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.G.; Babo-Rebelo, M.; Schwartz, D.; Tallon-Baudry, C. Phase-amplitude coupling at the organism level: The amplitude of spontaneous alpha rhythm fluctuations varies with the phase of the infra-slow gastric basal rhythm. NeuroImage 2017, 146, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Levinthal, D.J.; Strick, P.L. Multiple Areas of the Cerebral Cortex Influence the Stomach. Proc. Natl. Acad. Sci. USA 2020, 117, 13078–13083. Available online: https://www.pnas.org/content/early/2020/05/19/2002737117 (accessed on 5 June 2020). [CrossRef] [PubMed]
- Rebollo, I.; Devauchelle, A.-D.; Béranger, B.; Tallon-Baudry, C. Stomach-brain synchrony reveals a novel, delayed-connectivity resting-state network in humans. Elife 2018, 7, e33321. [Google Scholar] [CrossRef]
- Rebollo, I.; Wolpert, N.; Tallon-Baudry, C. Brain-stomach coupling: Anatomy, functions, and future avenues of research. Curr. Opin. Biomed. Eng. 2021, 18, 100270. [Google Scholar] [CrossRef]
- Azzalini, D.; Rebollo, I.; Tallon-Baudry, C. Visceral Signals Shape Brain Dynamics and Cognition. Trends Cogn. Sci. 2019, 23, 488–509. [Google Scholar] [CrossRef]
- Aftanas, L.I.; Golocheikine, S.A. Human anterior and frontal midline theta and lower alpha reflect emotionally positive state and internalized attention: High-resolution EEG investigation of meditation. Neurosci. Lett. 2001, 310, 57–60. [Google Scholar] [CrossRef]
- Arns, M.; Etkin, A.; Hegerl, U.; Williams, L.M.; DeBattista, C.; Palmer, D.M.; Fitzgerald, P.B.; Harris, A.; de Beuss, R.; Gordon, E. Frontal and rostral anterior cingulate (rACC) theta EEG in depression: Implications for treatment outcome? Eur. Neuropsychopharmacol. 2015, 25, 1190–1200. [Google Scholar] [CrossRef]
- Balasubramani, P.P.; Ojeda, A.; Grennan, G.; Maric, V.; Le, H.; Alim, F.; Zafar-Khan, M.; Diaz-Delgado, J.; Silveira, S.; Ramanathan, D.; et al. Mapping Cognitive Brain Functions at Scale. NeuroImage 2021, 231, 117641. [Google Scholar] [CrossRef] [PubMed]
- Gasser, T.; Verleger, R.; Bächer, P.; Sroka, L. Development of the EEG of school-age children and adolescents. I. Analysis of band power. Electroencephalogr. Clin. Neurophysiol. 1988, 69, 91–99. [Google Scholar] [CrossRef]
- HajiHosseini, A.; Holroyd, C.B. Reward feedback stimuli elicit high-beta EEG oscillations in human dorsolateral prefrontal cortex. Sci. Rep. 2015, 5, 13021. [Google Scholar] [CrossRef]
- Massar, S.A.A.; Rossi, V.; Schutter, D.; Kenemans, J.L. Baseline EEG theta/beta ratio and punishment sensitivity as biomarkers for feedback-related negativity (FRN) and risk-taking. Clin. Neurophysiol. 2012, 123, 1958–1965. [Google Scholar] [CrossRef]
- Agrusa, A.S.; Allegra, A.B.; Kunkel, D.C.; Coleman, T.P. Robust Methods to Detect Abnormal Initiation in the Gastric Slow Wave from Cutaneous Recordings. In Proceedings of the 2020 42nd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Montreal, QC, Canada, 20–24 July 2020; pp. 225–231. [Google Scholar]
- Gharibans, A.A.; Kim, S.; Kunkel, D.C.; Coleman, T.P. High-Resolution Electrogastrogram: A Novel, Noninvasive Method for Determining Gastric Slow-Wave Direction and Speed. IEEE Trans Biomed. Eng. 2017, 64, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Wolpert, N.; Rebollo, I.; Tallon-Baudry, C. Electrogastrography for psychophysiological research: Practical considerations, analysis pipeline, and normative data in a large sample. Psychophysiology 2019, 57, e13599. [Google Scholar] [CrossRef] [PubMed]
- Balasubramani, P.P.; Chakravarthy, V.S. Bipolar oscillations between positive and negative mood states in a computational model of Basal Ganglia. Cogn. Neurodyn. 2020, 14, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Meule, A.; Skirde, A.K.; Freund, R.; Vögele, C.; Kübler, A. High-calorie food-cues impair working memory performance in high and low food cravers. Appetite 2012, 59, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.P.; Clark, R.; Gallagher, J. Breakfast cereal and caffeinated coffee: Effects on working memory, attention, mood, and cardiovascular function. Physiol. Behav. 1999, 67, 9–17. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Mahony, S.M. The microbiome-gut-brain axis: From bowel to behavior. Neurogastroenterol. Motil. 2011, 23, 187–192. [Google Scholar] [CrossRef]
- Bloemendaal, M.; Szopinska-Tokov, J.; Belzer, C.; Boverhoff, D.; Papalini, S.; Michels, F.; van Hemert, S.; Vasquez, A.A.; Aarts, E. Probiotics-induced changes in gut microbial composition and its effects on cognitive performance after stress: Exploratory analyses. Transl. Psychiatry 2021, 11, 300. [Google Scholar] [CrossRef]
- Özkurt, T.E.; Schnitzler, A. A critical note on the definition of phase–amplitude cross-frequency coupling. J. Neurosci. Methods 2011, 201, 438–443. [Google Scholar] [CrossRef]
- Evrensel, A.; Ceylan, M.E. The Gut-Brain Axis: The Missing Link in Depression. Clin. Psychopharmacol. Neurosci. 2015, 13, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.-J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2014; pp. 39–71. [Google Scholar]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyses using G* Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods. 2009, 41, 1149–1160. [Google Scholar] [CrossRef]
- Boudreau, B.; Poulin, C. An examination of the validity of the Family Affluence Scale II (FAS II) in a general adolescent population of Canada. Soc. Indic. Res. 2008, 94, 29. [Google Scholar] [CrossRef]
- Babo-Rebelo, M.; Richter, C.G.; Tallon-Baudry, C. Neural Responses to Heartbeats in the Default Network Encode the Self in Spontaneous Thoughts. J. Neurosci. 2016, 36, 7829–7840. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Hsu, S.-H.; Pion-Tonachini, L.; Jung, T.-P. Evaluation of artifact subspace reconstruction for automatic EEG artifact removal. In Proceedings of the 2018 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Honolulu, HI, USA, 18–21 July 2018; pp. 1242–1245. [Google Scholar]
- Sternberg, S. High-speed scanning in human memory. Science 1966, 153, 652–654. [Google Scholar] [CrossRef]
- Lavie, N.; Hirst, A.; Fockert, J.W.D. Load Theory of Selective Attention and Cognitive Control. J. Exp. Psychol. Gen. 2004, 133, 339–354. [Google Scholar] [CrossRef]
- Lenartowicz, A.; Delorme, A.; Walshaw, P.D.; Cho, A.L.; Bilder, R.; McGough, J.J.; McCracken, J.T.; Makeig, S.; Loo, S.K. Electroencephalography Correlates of Spatial Working Memory Deficits in Attention-Deficit/Hyperactivity Disorder: Vigilance, Encoding, and Maintenance. J. Neurosci. 2014, 34, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Leek, M.R. Adaptive procedures in psychophysical research. Percept Psychophys. 2001, 63, 1279–1292. [Google Scholar] [CrossRef]
- Mishra, J.; Anguera, J.A.; Gazzaley, A. Video games for neuro-cognitive optimization. Neuron 2016, 90, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Kothe, C.; Medine, D.; Boulay, C.; Grivich, M.; Stenner, T. “Lab Streaming Layer” Copyright. 2019. Available online: https://labstreaminglayer.readthedocs.io/ (accessed on 18 January 2018).
- Jurkiewicz, G.J.; Hunt, M.J.; Żygierewicz, J. Addressing pitfalls in phase-amplitude coupling analysis with an extended modulation index toolbox. Neuroinformatics 2021, 19, 319–345. [Google Scholar] [CrossRef] [PubMed]
- Tort, A.B.L.; Kramer, M.A.; Thorn, C.; Gibson, D.J.; Kubota, Y.; Graybiel, A.M.; Kopell, N.J. Dynamic cross-frequency couplings of local field potential oscillations in rat striatum and hippocampus during performance of a T-maze task. Proc. Natl. Acad. Sci. USA 2008, 105, 20517–20522. [Google Scholar] [CrossRef] [PubMed]
- Tort, A.B.; Komorowski, R.; Eichenbaum, H.; Kopell, N. Measuring phase-amplitude coupling between neuronal oscillations of different frequencies. J. Neurophysiol. 2010, 104, 1195–1210. [Google Scholar] [CrossRef]
- Good, P. Permutation Tests: A Practical Guide to Resampling Methods for Testing Hypotheses; Springer Science & Business Media: New York, NY, USA, 2013. [Google Scholar]
- Dumouchel, W.; O’Brien, F. Integrating a robust option into a multiple regression computing environment. In Computer Science and Statistics, Proceedings of the 21st Symposium on the Interface, Monterey, CA, USA, 19–22 October 2008; American Statistical Association: Alexandria, VA, USA, 1989; pp. 297–302. [Google Scholar]
- Holland, P.W.; Welsch, R.E. Robust regression using iteratively reweighted least-squares. Commun. Stat. Theory Methods 1977, 6, 813–827. [Google Scholar] [CrossRef]
- Huber, P.J. Robust Statistics; John Wiley & Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- Street, J.O.; Carroll, R.J.; Ruppert, D. A note on computing robust regression estimates via iteratively reweighted least squares. Am. Stat. 1988, 42, 152–154. [Google Scholar]
- Avissar, M.; Powell, F.; Ilieva, I.; Respino, M.; Gunning, F.M.; Liston, C.; Dubin, M.J. Functional Connectivity of the Left DLPFC to Striatum Predicts Treatment Response of Depression to TMS. Brain Stimulat. 2017, 10, 919–925. [Google Scholar] [CrossRef]
- McClintock, S.M.; Reti, I.M.; Carpenter, L.L.; McDonald, W.M.; Dubin, M.; Taylor, S.F.; Cook, I.A.; O’Reardon, J.; Husain, M.M.; Wall, C.; et al. Consensus Recommendations for the Clinical Application of Repetitive Transcranial Magnetic Stimulation (rTMS) in the Treatment of Depression. J. Clin. Psychiatry 2018, 79, 35–48. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5846193/ (accessed on 3 May 2020). [CrossRef]
- Mayberg, H.S.; Brannan, S.K.; Mahurin, R.K.; Jerabek, P.A.; Brickman, J.S.; Tekell, J.L.; Silva, J.A.; McGinnis, S.; Glass, T.G.; Martin, C.C.; et al. Cingulate function in depression: A potential predictor of treatment response. Neuroreport 1997, 8, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- McGrath, C.L.; Kelley, M.E.; Dunlop, B.W.; Holtzheimer, P.E., III; Craighead, W.E.; Mayberg, H.S. Pretreatment brain states identify likely nonresponse to standard treatments for depression. Biol. Psychiatry 2014, 76, 527–535. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Regions | Factor | Robust Linear Fit Estimate ± SE | Robust Linear Fit tstat | Robust Linear Fit p-Value |
|---|---|---|---|---|
| Left fronto-central | Satiety | −0.11 × 10−2 ± 0.5 × 10−3 | −2.19 | 0.03 |
| Cognition | −0.20 × 10−2 ± 0.5 × 10−3 | −3.90 | 0.0003 | |
| interaction | 0.16 × 10−2 ± 0.7 × 10−3 | 2.19 | 0.03 | |
| Right fronto-central | Satiety | −0.15 × 10−3 ± 0.4 × 10−3 | −0.32 | 0.74 |
| Cognition | −0.10 × 10−2 ± 0.4 × 10−3 | −2.21 | 0.03 | |
| interaction | 0.56 × 10−3 ± 0.6 × 10−3 | 0.83 | 0.41 | |
| Left parieto-occipital | Satiety | 0.44 × 10−3 ± 0.7 × 10−3 | 0.63 | 0.52 |
| Cognition | −0.14 × 10−2 ± 0.7 × 10−3 | −2.05 | 0.04 | |
| interaction | −0.26 × 10−3 ± 0.9 × 10−3 | −0.26 | 0.79 | |
| Right parieto-occipital | Satiety | −0.36 × 10−3 ± 0.6 × 10−3 | 0.58 | 0.56 |
| Cognition | −0.80 × 10−3 ± 0.6 × 10−3 | −1.29 | 0.20 | |
| interaction | 0.23 × 10−3 ± 0.8 × 10−3 | −0.27 | 0.78 |
| Brain Regions | Main Effects and Interaction | rm-ANOVA (df = 13) Fstat | rm-ANOVA (df = 13) p-Value |
|---|---|---|---|
| Left fronto-central | Satiety | 1.48 | 0.25 |
| Cognition | 9.11 | 0.009 | |
| interaction | 7.21 | 0.02 | |
| Right fronto-central | Satiety | 0.04 | 0.84 |
| Cognition | 7.39 | 0.02 | |
| interaction | 0.26 | 0.61 | |
| Left parieto-occipital | Satiety | 0.10 | 0.76 |
| Cognition | 8.65 | 0.01 | |
| interaction | 0.45 | 0.51 | |
| Right parieto-occipital | Satiety | 0.03 | 0.87 |
| Cognition | 3.98 | 0.07 | |
| interaction | 2.32 | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balasubramani, P.P.; Walke, A.; Grennan, G.; Perley, A.; Purpura, S.; Ramanathan, D.; Coleman, T.P.; Mishra, J. Simultaneous Gut-Brain Electrophysiology Shows Cognition and Satiety Specific Coupling. Sensors 2022, 22, 9242. https://doi.org/10.3390/s22239242
Balasubramani PP, Walke A, Grennan G, Perley A, Purpura S, Ramanathan D, Coleman TP, Mishra J. Simultaneous Gut-Brain Electrophysiology Shows Cognition and Satiety Specific Coupling. Sensors. 2022; 22(23):9242. https://doi.org/10.3390/s22239242
Chicago/Turabian StyleBalasubramani, Pragathi Priyadharsini, Anuja Walke, Gillian Grennan, Andrew Perley, Suzanna Purpura, Dhakshin Ramanathan, Todd P. Coleman, and Jyoti Mishra. 2022. "Simultaneous Gut-Brain Electrophysiology Shows Cognition and Satiety Specific Coupling" Sensors 22, no. 23: 9242. https://doi.org/10.3390/s22239242
APA StyleBalasubramani, P. P., Walke, A., Grennan, G., Perley, A., Purpura, S., Ramanathan, D., Coleman, T. P., & Mishra, J. (2022). Simultaneous Gut-Brain Electrophysiology Shows Cognition and Satiety Specific Coupling. Sensors, 22(23), 9242. https://doi.org/10.3390/s22239242